Proteomic Characterization of Two Medically Important Malaysian Snake Venoms, Calloselasma rhodostoma (Malayan Pit Viper) and Ophiophagus hannah (King Cobra)


Abstract
:1. Introduction
2. Results
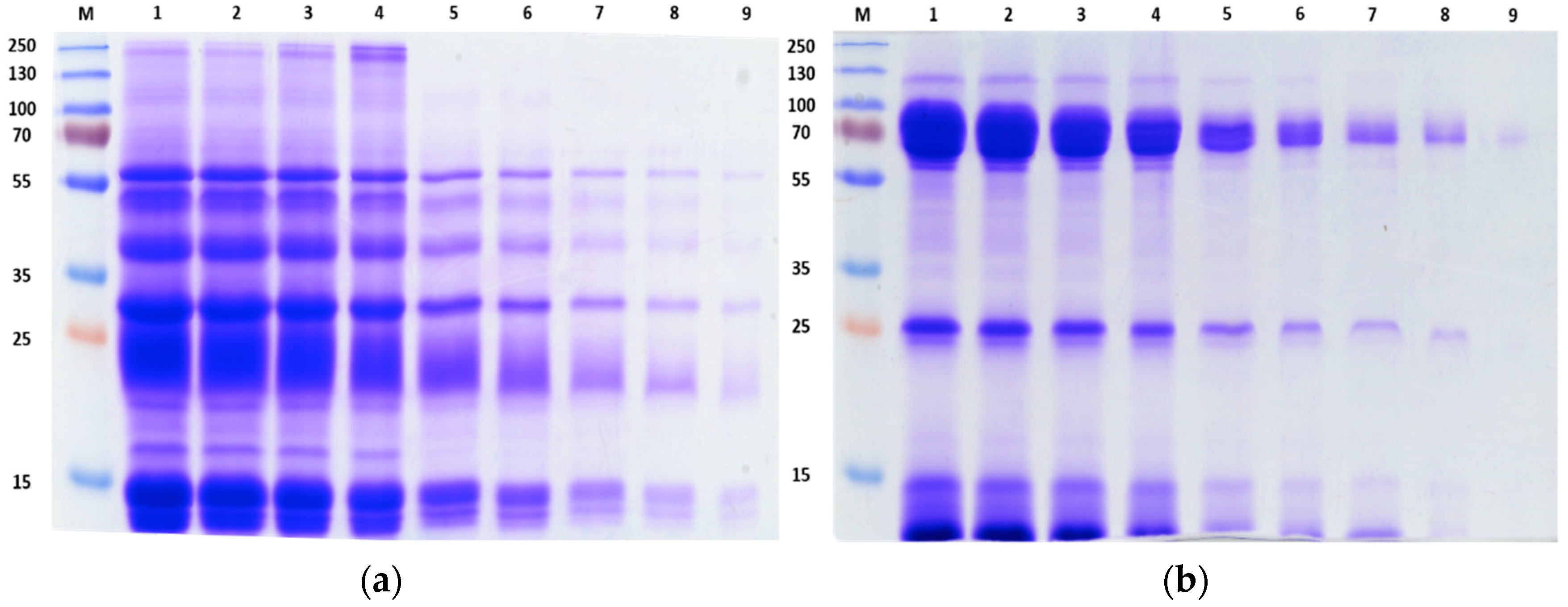
2.1. SDS-PAGE of Crude Venom Proteins
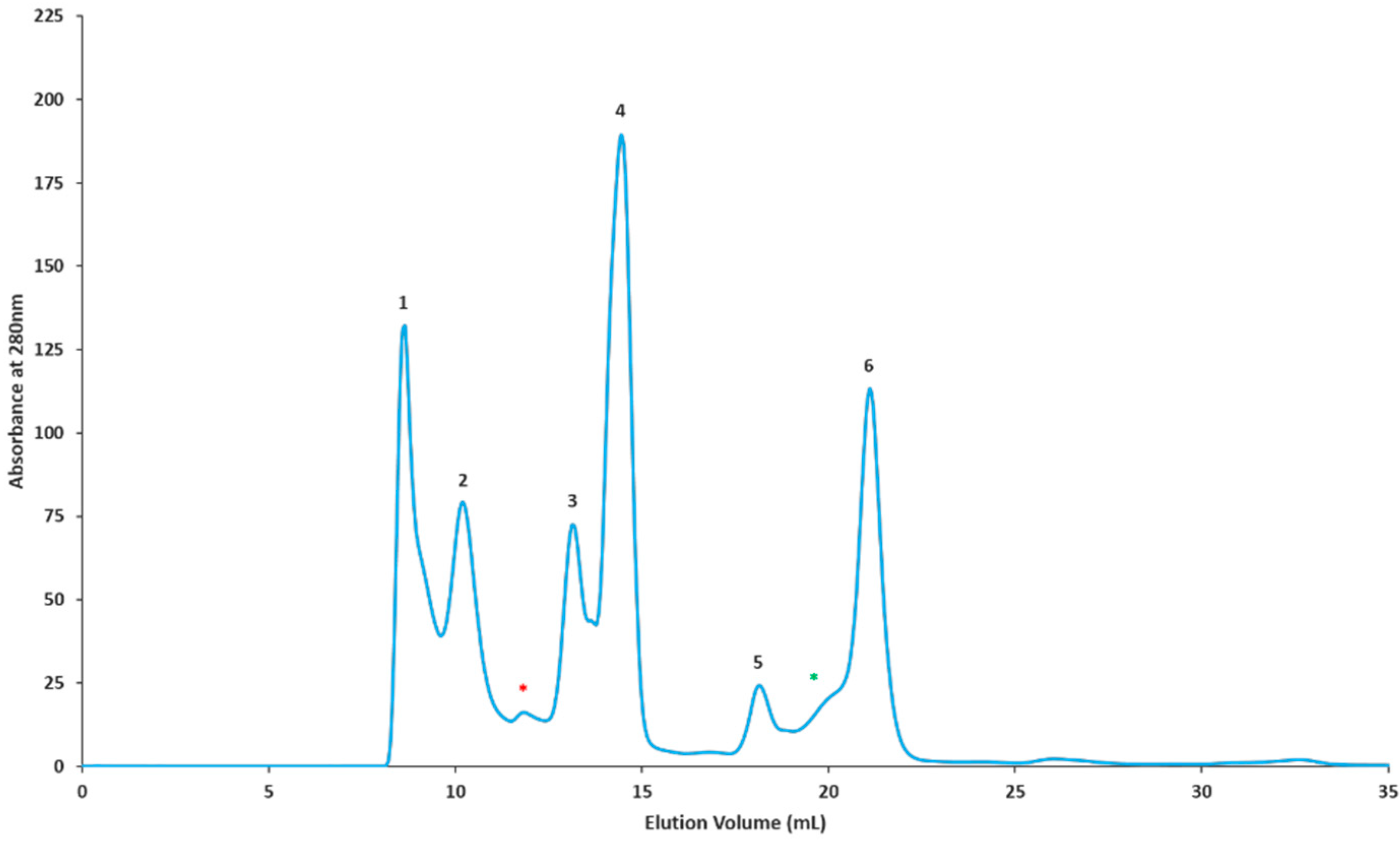
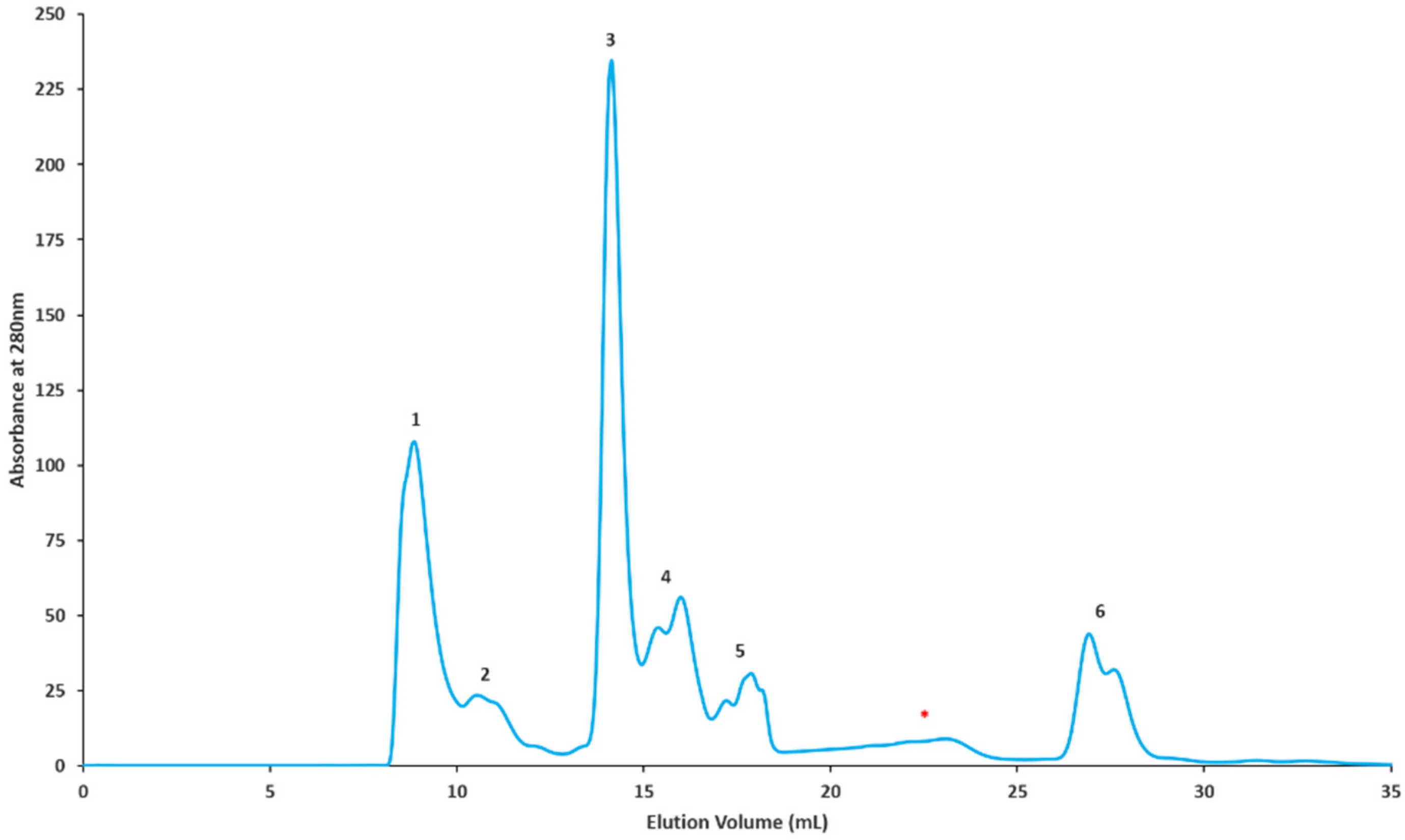
2.2. Gel Filtration Separation of Crude Snake Venom Using Superdex G-75
2.3. Snake Venom Proteomic Characterization Using Shotgun Approach (Shotgun-ESI-LC-MS/MS)
2.4. Snake Venom Proteomic Characterization Using Gel Filtration Protein Fractions (GF-ESI-LC-MS/MS)
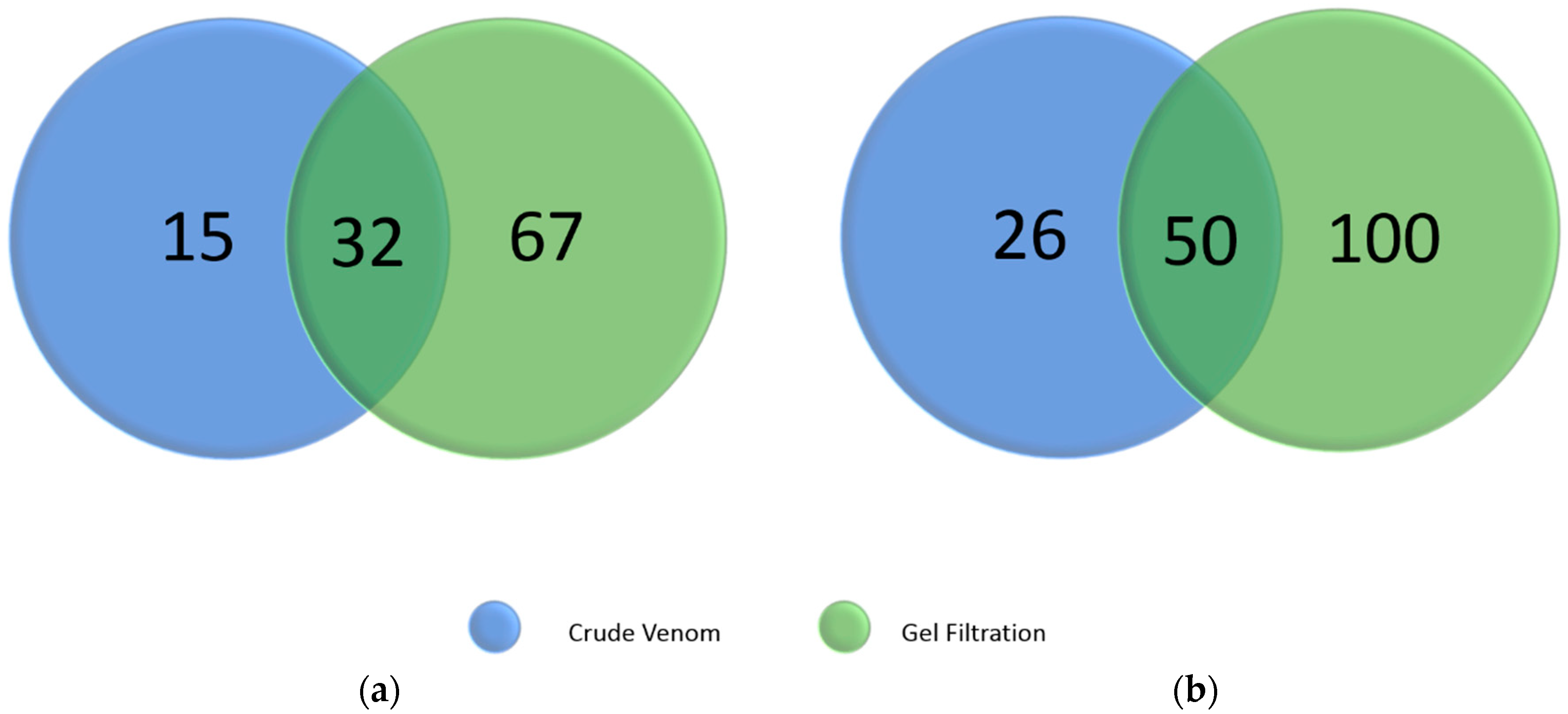
2.5. Summary of Identified C. rhodostoma and O. hannah Snake Venom Proteins
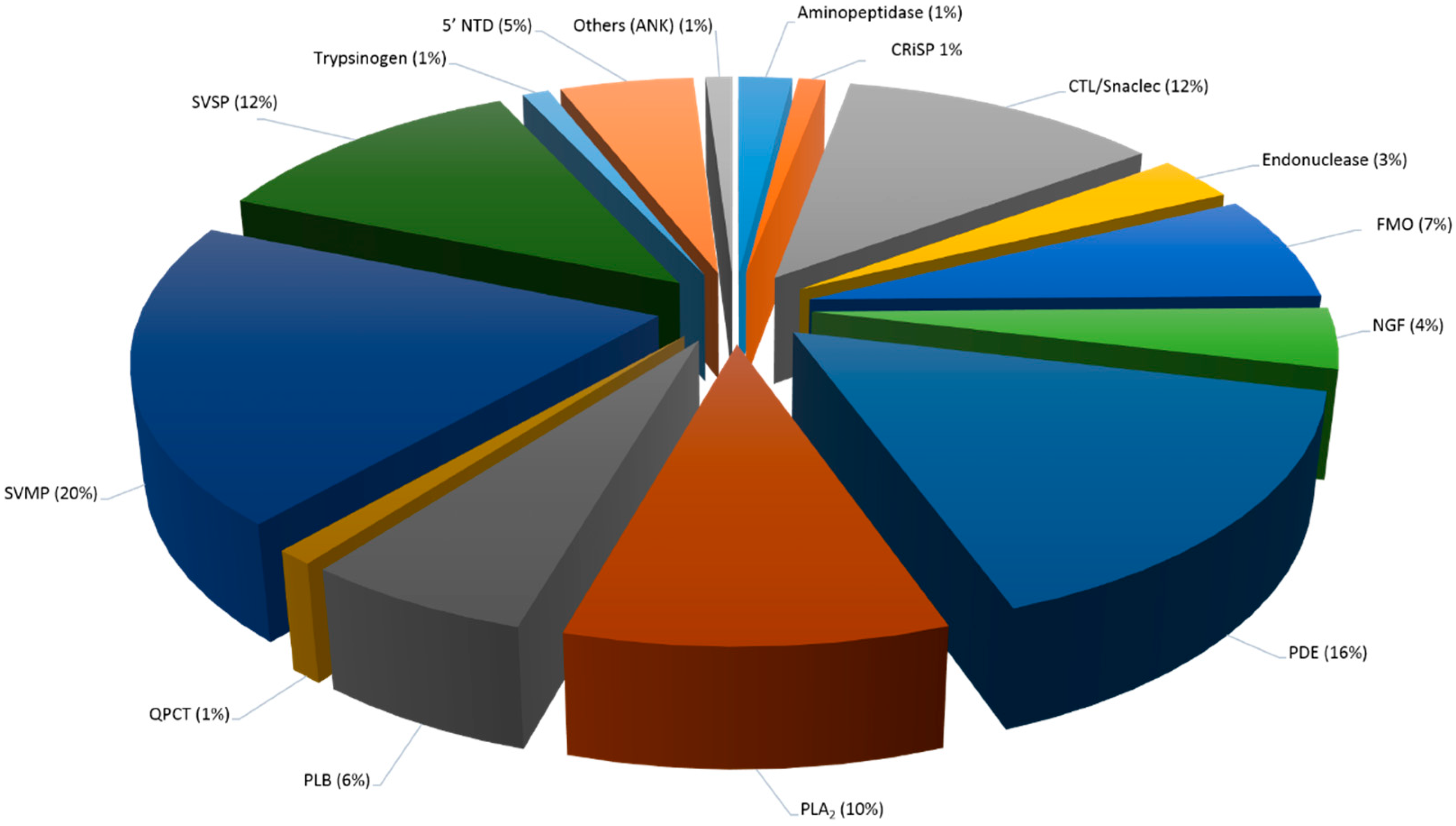
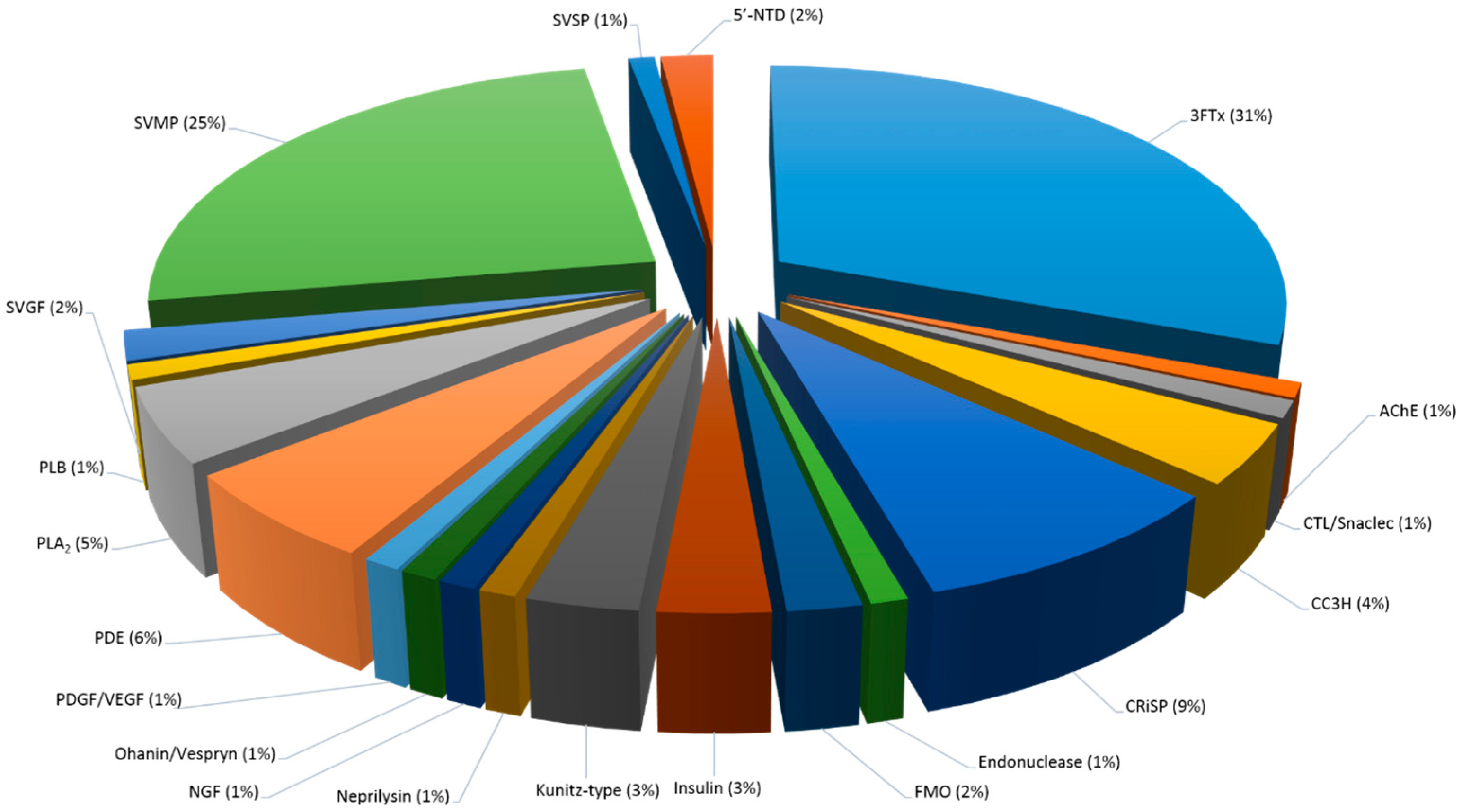
2.5.1. Snake Venom Protein Classes in Both C. rhodostoma and O. hannah Venoms
SVMP
SVSP
CRiSP
Phospholipase A2
PLB
CTL/snaclec
FMO (l-amino Acid Oxidase)
NGF
PDE
Endonuclease
Nucleotidase
2.5.2. Snake Venom Protein Classes in C. rhodostoma Venom
Low Abundance Proteins Only (<2%)
2.5.3. Snake Venom Protein Classes in O. hannah Venom
3FTx
Complement C3 Homolog
Kunitz-Type
Insulin
Low Abundant Proteins in O. hannah Venom (<2%)
3. Conclusions
4. Materials and Methods
4.1. Materials
4.1.1. Snake Venom
4.1.2. Columns, Drugs and Chemicals
4.2. Methods
4.2.1. Sodium Dodedyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
4.2.2. In-solution Protein Digestion
4.2.3. Identification of Crude Venom Proteins Using Shotgun LCMS/MS
4.2.4. Fractionation of C. rhodostoma and O. hannah Crude Venoms by Superdex G-75 Gel Filtration Chromatography
4.2.5. LC-MS/MS Analysis of Crude Venom and Gel-Filtration Protein Fractions
4.2.6. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Williams, D.; Gutiérrez, J.M.; Harrison, R.; Warrell, D.A.; White, J.; Winkel, K.D.; Gopalakrishnakone, P. The global snake bite initiative: An antidote for snake bite. Lancet 2010, 375, 89–91. [Google Scholar] [CrossRef]
- Koh, D.; Armugam, A.; Jeyaseelan, K. Snake venom components and their applications in biomedicine. Cell. Mol. Life Sci. CMLS 2006, 63, 3030–3041. [Google Scholar] [CrossRef] [PubMed]
- Kasai, K.; Ishikawa, T.; Nakamura, T.; Miura, T. Antibacterial properties of l-amino acid oxidase: Mechanisms of action and perspectives for therapeutic applications. Appl. Microbiol. Biotechnol. 2015, 99, 7847–7857. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.; Bhattacharjee, P.; Mishra, R.; Biswas, A.K.; Dasgupta, S.C.; Giri, B.; Debnath, A.; Gupta, S.D.; Das, T.; Gomes, A. Anticancer potential of animal venoms and toxins. Indian J. Exp. Boil. 2010, 48, 93–103. [Google Scholar]
- Iwanaga, S.; Suzuki, T. Enzymes in snake venom. In Snake Venoms; Springer: Berlin/Heidelberg, Germany, 1979; pp. 61–158. [Google Scholar]
- Chellapandi, P. Structural, functional and therapeutic aspects of snake venom metalloproteinases. Mini-Rev. Org. Chem. 2014, 11, 28–44. [Google Scholar] [CrossRef]
- King, G.F. Venoms as a platform for human drugs: Translating toxins into therapeutics. Expert Opin. Boil. Ther. 2011, 11, 1469–1484. [Google Scholar] [CrossRef] [PubMed]
- Oldrati, V.; Arrell, M.; Violette, A.; Perret, F.; Sprüngli, X.; Wolfender, J.-L.; Stöcklin, R. Advances in venomics. Mol. Biosyst. 2016, 12, 3530–3543. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J. Snake venomics: From the inventory of toxins to biology. Toxicon 2013, 75, 44–62. [Google Scholar] [CrossRef] [PubMed]
- De Lima, D.C.; Alvarez Abreu, P.; de Freitas, C.C.; Santos, D.O.; Borges, R.O.; Dos Santos, T.C.; Mendes Cabral, L.; Rodrigues, C.R.; Castro, H.C. Snake venom: Any clue for antibiotics and CAM? Evid.-Based Complement. Altern. Med. 2005, 2, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Das, I.; Ahmed, N.; Liat, L.B. Venomous terrestrial snakes of Malaysia: Their identity and biology. In Clinical Toxinology in Asia Pacific and Africa; Gopalakrishnakone, P., Faiz, M.A., Fernando, R., Gnanathasan, C.A., Habib, A.G., Yang, C.C., Eds.; Springer: Dordrecht, The Netherlands, 2015. [Google Scholar]
- Nolan, C.; Hall, L.; Barlow, G. [17] Ancrod, the coagulating enzyme from Malayan pit viper (Agkistrodon rhodostoma) venom. Methods Enzymol. 1976, 45, 205–213. [Google Scholar] [PubMed]
- Marsh, N.; Williams, V. Practical applications of snake venom toxins in haemostasis. Toxicon 2005, 45, 1171–1181. [Google Scholar] [CrossRef] [PubMed]
- McCleary, R.J.; Kang, T.S.; Kini, R.M. Reptile venoms as a platform for drug development. In Venoms to Drugs: Venom as a Source for the Development of Human Therapeutics, Chapter: Reptile Venoms as a Platform for Drug Development; Royal Society of Chemistry: Cambridge, UK, 2015; pp. 129–162. [Google Scholar]
- Slagboom, J.; Kool, J.; Harrison, R.A.; Casewell, N.R. Haemotoxic snake venoms: Their functional activity, impact on snakebite victims and pharmaceutical promise. Br. J. Haematol. 2017, 177, 947–959. [Google Scholar] [CrossRef] [PubMed]
- Pu, X.; Wong, P.; Gopalakrishnakone, P. A novel analgesic toxin (hannalgesin) from the venom of king cobra (Ophiophagus hannah). Toxicon 1995, 33, 1425–1431. [Google Scholar] [CrossRef]
- Koh, C.Y.; Kini, R.M. From snake venom toxins to therapeutics–cardiovascular examples. Toxicon 2012, 59, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.L. Toxins and drug discovery. Toxicon 2014, 92, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Tang, E.L.; Tan, C.H.; Fung, S.Y.; Tan, N.H. Venomics of Calloselasma rhodostoma, the Malayan pit viper: A complex toxin arsenal unraveled. J. Proteom. 2016, 148, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.A.; Baumann, K.; Jackson, T.N.; Wood, K.; Mason, S.; Undheim, E.A.; Nouwens, A.; Koludarov, I.; Hendrikx, I.; Jones, A.; et al. Proteomic comparison of Hypnale hypnale (Hump-Nosed Pit-Viper) and Calloselasma rhodostoma (Malayan Pit-Viper) venoms. J. Proteom. 2013, 91, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Vejayan, J.; Khoon, T.L.; Ibrahim, H. Comparative analysis of the venom proteome of four important Malaysian snake species. J. Venom. Anim. Toxins Incl. Trop. Dis. 2014, 20, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, C.H.; Tan, K.Y.; Fung, S.Y.; Tan, N.H. Venom-gland transcriptome and venom proteome of the Malaysian king cobra (Ophiophagus hannah). BMC Genom. 2015, 16, 687. [Google Scholar] [CrossRef] [PubMed]
- Vonk, F.J.; Casewell, N.R.; Henkel, C.V.; Heimberg, A.M.; Jansen, H.J.; McCleary, R.J.; Kerkkamp, H.M.; Vos, R.A.; Guerreiro, I.; Calvete, J.J.; et al. The king cobra genome reveals dynamic gene evolution and adaptation in the snake venom system. Proc. Natl. Acad. Sci. USA 2013, 110, 20651–20656. [Google Scholar] [CrossRef] [PubMed]
- Khow, O.; Chulasugandha, P.; Pakmanee, N. Venom protein of the haematotoxic snakes Cryptelytrops albolabris, Calloselasma rhodostoma, and Daboia russelii siamensis. Sci. Asia 2011, 37, 377–381. [Google Scholar] [CrossRef]
- Choudhury, M.; McCleary, R.J.; Kesherwani, M.; Kini, R.M.; Velmurugan, D. Comparison of proteomic profiles of the venoms of two of the ‘Big Four’ snakes of India, the Indian cobra (Naja naja) and the common krait (Bungarus caeruleus), and analyses of their toxins. Toxicon 2017, 135, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Shuting, L.; Jingqiang, W.; Zhang, X.; Yan, R.; Ning, W.; Kang, Z.; Xishu, C.; Caifeng, Z.; Xiaolei, L.; Jianmin, S.; et al. Proteomic characterization of two snake venoms: Naja naja atra and Agkistrodon halys. Biochem. J. 2004, 384, 119–127. [Google Scholar]
- De Oliveira, I.S.; Cardoso, I.A.; Bordon, K.D.C.F.; Carone, S.E.I.; Boldrini-França, J.; Pucca, M.B.; Zoccal, K.F.; Faccioli, L.H.; Sampaio, S.V.; Rosa, J.C.; et al. Global proteomic and functional analysis of Crotalus durissus collilineatus individual venom variation and its impact on envenoming. J. Proteom. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Vaiyapuri, S.; Wagstaff, S.C.; Watson, K.A.; Harrison, R.A.; Gibbins, J.M.; Hutchinson, E.G. Purification and functional characterisation of rhiminopeptidase A, a novel aminopeptidase from the venom of Bitis gabonica rhinoceros. PLoS Negl. Trop. Dis. 2010, 4, e796. [Google Scholar] [CrossRef] [PubMed]
- Markland, F.S., Jr.; Swenson, S. Snake venom metalloproteinases. Toxicon 2013, 62, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.W.; Serrano, S.M. Timeline of key events in snake venom metalloproteinase research. J. Proteom. 2009, 72, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.W.; Serrano, S.M. Structural considerations of the snake venom metalloproteinases, key members of the M12 reprolysin family of metalloproteinases. Toxicon 2005, 45, 969–985. [Google Scholar] [CrossRef] [PubMed]
- Tasoulis, T.; Isbister, G.K. A review and database of snake venom proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef] [PubMed]
- Serrano, S.M. The long road of research on snake venom serine proteinases. Toxicon 2013, 62, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Serrano, S.M.; Maroun, R.C. Snake venom serine proteinases: Sequence homology vs. Substrate specificity, a paradox to be solved. Toxicon 2005, 45, 1115–1132. [Google Scholar] [CrossRef] [PubMed]
- Stubbs, M.T.; Bode, W. A player of many parts: The spotlight falls on thrombin’s structure. Thromb. Res. 1993, 69, 1–58. [Google Scholar] [CrossRef]
- Esnouf, M.; Tunnah, G. The isolation and properties of the thrombin-like activity from Ancistrodon rhodostoma venom. Br. J. Haematol. 1967, 13, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Sanggaard, K.W.; Dyrlund, T.F.; Thomsen, L.R.; Nielsen, T.A.; Brøndum, L.; Wang, T.; Thøgersen, I.B.; Enghild, J.J. Characterization of the gila monster (Heloderma suspectum suspectum) venom proteome. J. Proteom. 2015, 117, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sanggaard, K.W.; Bechsgaard, J.S.; Fang, X.; Duan, J.; Dyrlund, T.F.; Gupta, V.; Jiang, X.; Cheng, L.; Fan, D.; Feng, Y.; et al. Spider genomes provide insight into composition and evolution of venom and silk. Nat. Commun. 2014, 5, 3765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safavi-Hemami, H.; Hu, H.; Gorasia, D.G.; Bandyopadhyay, P.K.; Veith, P.D.; Young, N.D.; Reynolds, E.C.; Yandell, M.; Olivera, B.M.; Purcell, A.W. Combined proteomic and transcriptomic interrogation of the venom gland of Conus geographus uncovers novel components and functional compartmentalization. Mol. Cell. Proteom. 2014, 13, 938–953. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.D.; Safavi-Hemami, H.; McIntosh, L.D.; Purcell, A.W.; Norton, R.S.; Papenfuss, A.T. Diversity of conotoxin gene superfamilies in the venomous snail, Conus victoriae. PLoS ONE 2014, 9, e87648. [Google Scholar] [CrossRef] [PubMed]
- Hook, V.; Yasothornsrikul, S.; Greenbaum, D.; Medzihradszky, K.F.; Troutner, K.; Toneff, T.; Bundey, R.; Logrinova, A.; Reinheckel, T.; Peters, C.; et al. Cathepsin L and Arg/Lys aminopeptidase: A distinct prohormone processing pathway for the biosynthesis of peptide neurotransmitters and hormones. Boil. Chem. 2004, 385, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.S.; Creemers, J.W.; Ohagi, S.; Raffin-Sanson, M.-L.; Sanders, L.; Montague, C.T.; Hutton, J.C.; O’Rahilly, S. Obesity and impaired prohormone processing associated with mutations in the human prohormone convertase 1 gene. Nat. Genet. 1997, 16, 303. [Google Scholar] [CrossRef] [PubMed]
- Milne, T.J.; Abbenante, G.; Tyndall, J.D.; Halliday, J.; Lewis, R.J. Isolation and characterization of a cone snail protease with homology to crisp proteins of the pathogenesis-related protein superfamily. J. Boil. Chem. 2003, 278, 31105–31110. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Morita, T. Structure and function of snake venom cysteine-rich secretory proteins. Toxicon 2004, 44, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Morita, T. Snake venom components affecting blood coagulation and the vascular system: Structural similarities and marked diversity. Curr. Pharm. Des. 2007, 13, 2872–2886. [Google Scholar] [CrossRef] [PubMed]
- Sunagar, K.; Johnson, W.E.; O’Brien, S.J.; Vasconcelos, V.; Antunes, A. Evolution of crisps associated with toxicoferan-reptilian venom and mammalian reproduction. Mol. Boil. Evol. 2012, 29, 1807–1822. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, Y.; Yamazaki, Y.; Hyodo, F.; Sugiyama, Y.; Nozaki, M.; Morita, T. Structural divergence of cysteine-rich secretory proteins in snake venoms. J. Biochem. 2008, 145, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.L.; Haley, T.L.; West, K.A.; Crabb, J.W. Pseudechetoxin: A peptide blocker of cyclic nucleotide-gated ion channels. Proc. Natl. Acad. Sci. USA 1999, 96, 754–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamazaki, Y.; Hyodo, F.; Morita, T. Wide distribution of cysteine-rich secretory proteins in snake venoms: Isolation and cloning of novel snake venom cysteine-rich secretory proteins. Arch. Biochem. Biophys. 2003, 412, 133–141. [Google Scholar] [CrossRef]
- Kini, R.M. Excitement ahead: Structure, function and mechanism of snake venom phospholipase A2 enzymes. Toxicon 2003, 42, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Lomonte, B. Phospholipases A2: Unveiling the secrets of a functionally versatile group of snake venom toxins. Toxicon 2013, 62, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Tsai, I.H.; Wang, Y.M.; Au, L.C.; Ko, T.P.; Chen, Y.H.; Chu, Y.F. Phospholipases A2 from Callosellasma rhodostoma venom gland. FEBS J. 2000, 267, 6684–6691. [Google Scholar] [CrossRef]
- Tsai, S.-H.; Chen, Y.-C.; Chen, L.; Wang, Y.-M.; Tsai, I.-H. Binding of a venom Lys-49 phospholipase A2 to LPS and suppression of its effects on mouse macrophages. Toxicon 2007, 50, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Tan, N.H.; Fung, S.Y.; Tan, K.Y.; Yap, M.K.K.; Gnanathasan, C.A.; Tan, C.H. Functional venomics of the Sri Lankan Russell’s viper (Daboia russelii) and its toxinological correlations. J. Proteom. 2015, 128, 403–423. [Google Scholar] [CrossRef] [PubMed]
- Doery, H.M.; Pearson, J.E. Phospholipase B in snake venoms and bee venom. Biochem. J. 1964, 92, 599. [Google Scholar] [CrossRef] [PubMed]
- Van Deenen, L.L.M.; de Haas, G.H. The substrate specificity of phospholipase A. Biochim. Biophys. Acta (BBA) Spéc. Sect. Lipids Relat. Subj. 1963, 70, 538–553. [Google Scholar]
- Mohamed, A.; Kamel, A.; Ayobe, M. Studies of phospholipase A and B activities of Egyptian snake venoms and a scorpion toxin. Toxicon 1969, 6, 293–298. [Google Scholar] [CrossRef]
- Shiloah, J.; Klibansky, C.; De Vries, A.; Berger, A. Phospholipase B activity of a purified phospholipase A from Vipera palestinae venom. J. Lipid Res. 1973, 14, 267–278. [Google Scholar] [PubMed]
- Bernheimer, A.; Linder, R.; Weinstein, S.; Kim, K.-S. Isolation and characterization of a phospholipase B from venom of collett’s snake, Pseudechis colletti. Toxicon 1987, 25, 547–554. [Google Scholar] [CrossRef]
- Kovalchuk, S.I.; Ziganshin, R.H.; Starkov, V.G.; Tsetlin, V.I.; Utkin, Y.N. Quantitative proteomic analysis of venoms from Russian vipers of Pelias group: Phospholipases A2 are the main venom components. Toxins 2016, 8, 105. [Google Scholar] [CrossRef] [PubMed]
- Ziganshin, R.H.; Kovalchuk, S.I.; Arapidi, G.P.; Starkov, V.G.; Hoang, A.N.; Nguyen, T.T.T.; Nguyen, K.C.; Shoibonov, B.B.; Tsetlin, V.I.; Utkin, Y.N. Quantitative proteomic analysis of Vietnamese krait venoms: Neurotoxins are the major components in Bungarus multicinctus and phospholipases A2 in Bungarus fasciatus. Toxicon 2015, 107, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Zainal Abidin, S.A.; Rajadurai, P.; Chowdhury, M.E.H.; Ahmad Rusmili, M.R.; Othman, I.; Naidu, R. Proteomic characterization and comparison of malaysian Tropidolaemus wagleri and Cryptelytrops purpureomaculatus venom using shotgun-proteomics. Toxins 2016, 8, 299. [Google Scholar] [CrossRef] [PubMed]
- Morita, T. Structures and functions of snake venom CLPs (C-type lectin-like proteins) with anticoagulant-, procoagulant-, and platelet-modulating activities. Toxicon 2005, 45, 1099–1114. [Google Scholar] [CrossRef] [PubMed]
- Clemetson, K.J. Snaclecs (snake C-type lectins) that inhibit or activate platelets by binding to receptors. Toxicon 2010, 56, 1236–1246. [Google Scholar] [CrossRef] [PubMed]
- Qinghua, L.; Xiaowei, Z.; Wei, Y.; Chenji, L.; Yijun, H.; Pengxin, Q.; Xingwen, S.; Songnian, H.; Guangmei, Y. A catalog for transcripts in the venom gland of the Agkistrodon acutus: Identification of the toxins potentially involved in coagulopathy. Biochem. Biophys. Res. Commun. 2006, 341, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.H.; Tan, N.H.; Sim, S.M.; Fung, S.Y.; Gnanathasan, C.A. Proteomic investigation of Sri Lankan hump-nosed pit viper (Hypnale hypnale) venom. Toxicon 2015, 93, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Du, X.-Y.; Clemetson, K.J. Snake venom l-amino acid oxidases. Toxicon 2002, 40, 659–665. [Google Scholar] [CrossRef]
- Fung, S.; Tan, N. Snake venom l-amino acid oxidases. In Handbook of Venoms and Toxins of Reptiles; CRC Press Inc.: London, UK, 2009; pp. 221–235. [Google Scholar]
- Baby, J.; Rajan Sheeja, S.; Jeevitha, M.; Ajisha, S. Pharmacological effects of snake venom l-amino acid oxidases. Int. J. Res. Ayurveda Pharm. 2011, 2, 114–120. [Google Scholar]
- Fry, B. Venomous Reptiles and Their Toxins: Evolution, Pathophysiology and Biodiscovery; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Ponnudurai, G.; Chung, M.C.; Tan, N.-H. Purification and properties of the l-amino acid oxidase from Malayan pit viper (Calloselasma rhodostoma) venom. Arch. Biochem. Biophys. 1994, 313, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Levi-Montalcini, R. A nerve growth-stimulating factor isolated from snake venom. Proc. Natl. Acad. Sci. USA 1956, 42, 571–574. [Google Scholar] [CrossRef] [PubMed]
- McCleary, R.J.; Kini, R.M. Non-enzymatic proteins from snake venoms: A gold mine of pharmacological tools and drug leads. Toxicon 2013, 62, 56–74. [Google Scholar] [CrossRef] [PubMed]
- Trummal, K.; Tõnismägi, K.; Paalme, V.; Järvekülg, L.; Siigur, J.; Siigur, E. Molecular diversity of snake venom nerve growth factors. Toxicon 2011, 58, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Earl, S.T.; Birrell, G.W.; Wallis, T.P.; St Pierre, L.D.; Masci, P.P.; de Jersey, J.; Gorman, J.J.; Lavin, M.F. Post-translational modification accounts for the presence of varied forms of nerve growth factor in Australian elapid snake venoms. Proteomics 2006, 6, 6554–6565. [Google Scholar] [CrossRef] [PubMed]
- Kostiza, T.; Meier, J. Nerve growth factors from snake venoms: Chemical properties, mode of action and biological significance. Toxicon 1996, 34, 787–806. [Google Scholar] [CrossRef]
- Boldrini-França, J.; Cologna, C.T.; Pucca, M.B.; Bordon, K.D.C.F.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cerni, F.A.; Pinheiro-Junior, E.L.; et al. Minor snake venom proteins: Structure, function and potential applications. Biochim. Biophys. Acta (BBA) Gen. Subj. 2017, 1861, 824–838. [Google Scholar] [CrossRef] [PubMed]
- Aird, S.D. Ophidian envenomation strategies and the role of purines. Toxicon 2002, 40, 335–393. [Google Scholar] [CrossRef]
- Dhananjaya, B.L.; D’souza, C.J. The pharmacological role of nucleotidases in snake venoms. Cell Biochem. Funct. 2010, 28, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Danpaiboon, W.; Reamtong, O.; Sookrung, N.; Seesuay, W.; Sakolvaree, Y.; Thanongsaksrikul, J.; Dong-din-on, F.; Srimanote, P.; Thueng-in, K.; Chaicumpa, W. Ophiophagus hannah venom: Proteome, components bound by Naja kaouthia antivenin and neutralization by N. kaouthia neurotoxin-specific human ScFv. Toxins 2014, 6, 1526–1558. [Google Scholar] [CrossRef] [PubMed]
- Dhananjaya, B.; D’souza, C.J. An overview on nucleases (DNase, RNase, and phosphodiesterase) in snake venoms. Biochemistry. 2010, 75, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Aird, S.D. Taxonomic distribution and quantitative analysis of free purine and pyrimidine nucleosides in snake venoms. Comp. Biochem. Physiol. Part B Biochem. Mol. Boil. 2005, 140, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, C.; Huang, T.-F. Inhibition of platelet aggregation by 5′-nucleotidase purified from Trimeresurus gramineus snake venom. Toxicon 1983, 21, 491–501. [Google Scholar] [CrossRef]
- Ouyang, C.; Huang, T.-F. Platelet aggregation inhibitors from Agkistrodon acutus snake venom. Toxicon 1986, 24, 1099–1106. [Google Scholar] [CrossRef]
- Boffa, M.-C.; Boffa, G.A. Correlations between the enzymatic activities and the factors active on blood coagulation and platelet aggregation from the venom of Vipera aspis. Biochim. Biophys. Acta (BBA) Gen. Subj. 1974, 354, 275–290. [Google Scholar] [CrossRef]
- Dhananjaya, B.; Nataraju, A.; Rajesh, R.; Gowda, C.R.; Sharath, B.; Vishwanath, B.; D’Souza, C.J. Anticoagulant effect of Naja naja venom 5′ nucleotidase: Demonstration through the use of novel specific inhibitor, vanillic acid. Toxicon 2006, 48, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Yap, M.K.; Fung, S.Y.; Tan, K.Y.; Tan, N.H. Proteomic characterization of venom of the medically important Southeast Asian Naja sumatrana (Equatorial spitting cobra). Acta Trop. 2014, 133, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Melani, R.D.; Araujo, G.D.; Carvalho, P.C.; Goto, L.; Nogueira, F.C.; Junqueira, M.; Domont, G.B. Seeing beyond the tip of the iceberg: A deep analysis of the venome of the Brazilian Rattlesnake, Crotalus durissus terrificus. EuPA Open Proteom. 2015, 8, 144–156. [Google Scholar] [CrossRef]
- Aird, S.D.; Watanabe, Y.; Villar-Briones, A.; Roy, M.C.; Terada, K.; Mikheyev, A.S. Quantitative high-throughput profiling of snake venom gland transcriptomes and proteomes (Ovophis okinavensis and Protobothrops flavoviridis). BMC Genom. 2013, 14, 790. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, D.; Öhler, M.; Seifert, J.; Von Bergen, M.; Arni, R.K.; Genov, N.; Betzel, C. Snake venomic of Crotalus durissus terrificus—Correlation with pharmacological activities. J. Proteome Res. 2010, 9, 2302–2316. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.W.; Ma, L.; Nelson, K.; Sherman, N.E.; Serrano, S.M. Comparison of indirect and direct approaches using ion-trap and Fourier transform ion cyclotron resonance mass spectrometry for exploring viperid venom proteomes. Toxicon 2006, 47, 700–714. [Google Scholar] [CrossRef] [PubMed]
- Chapeaurouge, A.; Reza, M.A.; Mackessy, S.P.; Carvalho, P.C.; Valente, R.H.; Teixeira-Ferreira, A.; Perales, J.; Lin, Q.; Kini, R.M. Interrogating the venom of the viperid snake Sistrurus catenatus edwardsii by a combined approach of electrospray and MALDI mass spectrometry. PLoS ONE 2015, 10, e0092091. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves-Machado, L.; Pla, D.; Sanz, L.; Jorge, R.J.B.; Leitão-De-Araújo, M.; Alves, M.L.M.; Alvares, D.J.; De Miranda, J.; Nowatzki, J.; de Morais-Zani, K.; et al. Combined venomics, venom gland transcriptomics, bioactivities, and antivenomics of two Bothrops jararaca populations from geographic isolated regions within the Brazilian Atlantic rainforest. J. Proteom. 2016, 135, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-M.; Huang, K.-F.; Tsai, I.-H. Snake venom glutaminyl cyclases: Purification, cloning, kinetic study, recombinant expression, and comparison with the human enzyme. Toxicon 2014, 86, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, J.; Kini, R.M. Snake venom glutaminyl cyclase. Toxicon 2006, 48, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Breeden, L.; Nasmyth, K. Similarity between cell-cycle genes of budding yeast and fission yeast and the Notch gene of Drosophila. Nature 1987, 329, 651–654. [Google Scholar] [CrossRef] [PubMed]
- Foord, R.; Taylor, I.A.; Sedgwick, S.G.; Smerdon, S.J. X-ray structural analysis of the yeast cell cycle regulator Swi6 reveals variations of the ankyrin fold and has implications for Swi6 function. Nat. Struct. Mol. Boil. 1999, 6, 157–165. [Google Scholar]
- He, Q.; Duan, Z.; Yu, Y.; Liu, Z.; Liu, Z.; Liang, S. The venom gland transcriptome of Latrodectus tredecimguttatus revealed by deep sequencing and cDNA library analysis. PLoS ONE 2013, 8, e81357. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M. Molecular moulds with multiple missions: Functional sites in three-finger toxins. Clin. Exp. Pharmacol. Physiol. 2002, 29, 815–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kini, R.M.; Doley, R. Structure, function and evolution of three-finger toxins: Mini proteins with multiple targets. Toxicon 2010, 56, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Dufton, M.; Hider, R. Structure and pharmacology of elapid cytotoxins. Pharmacol. Ther. 1988, 36, 1–40. [Google Scholar] [CrossRef]
- Fry, B.G.; Lumsden, N.G.; Wüster, W.; Wickramaratna, J.C.; Hodgson, W.C.; Kini, R.M. Isolation of a neurotoxin (α-colubritoxin) from a nonvenomous colubrid: Evidence for early origin of venom in snakes. J. Mol. Evol. 2003, 57, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Vogel, C.-W.; Bredehorst, R.; Fritzinger, D.C.; Grunwald, T.; Ziegelmüller, P.; Kock, M.A. Structure and function of cobra venom factor, the complement-activating protein in cobra venom. In Natural Toxins 2; Springer: Boston, MA, USA, 1996; pp. 97–114. [Google Scholar]
- Vogel, C.-W.; Fritzinger, D.C. Cobra venom factor: Structure, function, and humanization for therapeutic complement depletion. Toxicon 2010, 56, 1198–1222. [Google Scholar] [CrossRef] [PubMed]
- Ranasinghe, S.; McManus, D.P. Structure and function of invertebrate Kunitz serine protease inhibitors. Dev. Comp. Immunol. 2013, 39, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Župunski, V.; Kordiš, D.; Gubenšek, F. Adaptive evolution in the snake venom Kunitz/BPTI protein family. FEBS Lett. 2003, 547, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Feng, J.; Wang, B.; Cao, Z.; Li, W.; Wu, Y.; Chen, Z. BF9, the first functionally characterized snake toxin peptide with Kunitz-type protease and potassium channel inhibiting properties. J. Biochem. Mol. Toxicol. 2014, 28, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Shafqat, J.; Beg, O.U.; Yin, S.J.; Zaidi, Z.H.; Jörnvall, H. Primary structure and functional properties of cobra (Naja naja naja) venom Kunitz-type trypsin inhibitor. FEBS J. 1990, 194, 337–341. [Google Scholar] [CrossRef]
- Shabanpoor, F.; Separovic, F.; Wade, J.D. The human insulin superfamily of polypeptide hormones. Vitam. Horm. 2009, 80, 1–31. [Google Scholar] [PubMed]
- Safavi-Hemami, H.; Gajewiak, J.; Karanth, S.; Robinson, S.D.; Ueberheide, B.; Douglass, A.D.; Schlegel, A.; Imperial, J.S.; Watkins, M.; Bandyopadhyay, P.K.; et al. Specialized insulin is used for chemical warfare by fish-hunting cone snails. Proc. Natl. Acad. Sci. USA 2015, 112, 1743–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyengar, N.; Sehra, K.; Mukerji, B.; Chopra, R. Choline esterase in cobra venom. Curr. Sci. 1938, 7, 51–53. [Google Scholar]
- Frobert, Y.; Créminon, C.; Cousin, X.; Rémy, M.-H.; Chatel, J.-M.; Bon, S.; Bon, C.; Grassi, J. Acetylcholinesterases from Elapidae snake venoms: Biochemical, immunological and enzymatic characterization. Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzym. 1997, 1339, 253–267. [Google Scholar] [CrossRef]
- Kerr, M.; Kenny, A. The purification and specificity of a neutral endopeptidase from rabbit kidney brush border. Biochem. J. 1974, 137, 477–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casewell, N.R.; Harrison, R.A.; Wüster, W.; Wagstaff, S.C. Comparative venom gland transcriptome surveys of the saw-scaled vipers (Viperidae: Echis) reveal substantial intra-family gene diversity and novel venom transcripts. BMC Genom. 2009, 10, 564. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.J.; Isaac, R.E.; Coates, D. The neprilysin (NEP) family of zinc metalloendopeptidases: Genomics and function. Bioessays 2001, 23, 261–269. [Google Scholar] [CrossRef]
- Pung, Y.F.; Wong, P.T.; Kumar, P.P.; Hodgson, W.C.; Kini, R.M. Ohanin, a novel protein from king cobra venom, induces hypolocomotion and hyperalgesia in mice. J. Boil. Chem. 2005, 280, 13137–13147. [Google Scholar] [CrossRef] [PubMed]
- Pung, Y.F.; Kumar, S.V.; Rajagopalan, N.; Fry, B.G.; Kumar, P.P.; Kini, R.M. Ohanin, a novel protein from king cobra venom: Its cDNA and genomic organization. Gene 2006, 371, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishnakone, P.; Pu, X.C.; Wong, P.T.-H.; Gwee, M.C.E.; Kini, R.M. Therapeutic Molecules Derived from Snake Venom. U.S. Patent 6613745B1, 2 September 2003. [Google Scholar]
- Wordinger, R.J.; Clark, A.F. Growth factors and neurotrophic factors as targets. Ocul. Ther. 2008, 87–116. [Google Scholar] [CrossRef]
- Mori, T.; Shimizu, A.; Masuda, Y.; Fukuda, Y.; Yamanaka, N. Hepatocyte growth factor-stimulating endothelial cell growth and accelerating glomerular capillary repair in experimental progressive glomerulonephritis. Nephron Exp. Nephrol. 2003, 94, e44–e54. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, J. SDS-polyacrylamide gel electrophoresis of protein. Nature 1970, 2, 680–685. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Family | Protein Description | No. of Proteins | Protein Accession No. | No. of Peptides | No. of Unique Peptides |
|---|---|---|---|---|---|
| 5′-NTD | 5′ nucleotidase (P. flavoviridis) | 5 | T2HRS9 | 4 | 2 |
| 5-nucleotidase (P. elegans) | A0A077L6M5 | 4 | 2 | ||
| 5′ nucleotidase (P. flavoviridis) | A0A077L7M9 | 4 | 2 | ||
| Snake venom 5′-nucleotidase (A. piscivorus) | A0A194APL9 | 3 | 2 | ||
| BATXNUC1 (B. atrox) | A0A1L8D667 | 3 | 2 | ||
| CRiSP | Cysteine-rich secretory protein LCCL 2 (C. adamanteus) | 1 | A0A0F7ZEB6 | 2 | 2 |
| CTL/snaclec | C-type lectin (A. piscivorus leucostoma) | 12 | G8FML6 | 4 | 4 |
| C-type lectin 2 (A. piscivorus) | A0A194APP2 | 6 | 6 | ||
| C-type lectin APL (A. piscivorus piscivorus) | P0DM36 | 4 | 4 | ||
| C-type lectin BjL (B. jararaca) | Q9PRY7 | 4 | 4 | ||
| C-type lectin PAL (B. arietans) | Q9PSN0 | 4 | 4 | ||
| Snaclec clone 2100755 (D. acutus) | Q8JIV8 | 3 | 3 | ||
| Snaclec rhodocetin subunit alpha (C. rhodostoma) | P81397 | 17 | 15 | ||
| Snaclec rhodocetin subunit beta (C. rhodostoma) | P81398 | 15 | 14 | ||
| Snaclec rhodocetin subunit gamma (C. rhodostoma) | D2YW39 | 6 | 4 | ||
| Snaclec rhodocetin subunit delta (C. rhodostoma) | D2YW40 | 6 | 6 | ||
| Snaclec rhodocytin subunit alpha (C. rhodostoma) | Q9I841 | 8 | 8 | ||
| Snaclec rhodocytin subunit beta (C. rhodostoma) | Q9I840 | 14 | 13 | ||
| FMO | Amine oxidase (O. okinavensis) | 2 | T2HQ57 | 18 | 2 |
| L-amino-acid oxidase (C. rhodostoma) | P81382 | 53 | 34 | ||
| PDE | Cadam10_PDE-1 (C. adamanteus) | 7 | A0A1W7RBB0 | 6 | 2 |
| Ectonucleotide pyrophosphatase/phosphodiesterase 3 (C. horridus) | T1DJT5 | 6 | 2 | ||
| Phosphodiesterase (C. horridus) | T1D6P7 | 6 | 3 | ||
| Phosphodiesterase (C. adamanteus) | A0A0F7Z2Q3 | 6 | 3 | ||
| Phosphodiesterase (S. miliarius barbouri) | A0A194AS02 | 6 | 2 | ||
| Venom phosphodiesterase 1 (C. adamanteus) | J3SEZ3 | 6 | 3 | ||
| Venom phosphodiesterase 2 (C. adamanteus) | J3SBP3 | 6 | 3 | ||
| PLA2 | Acidic phospholipase A2 H1E6 (C. rhodostoma) | 6 | Q9PVF2 | 11 | 10 |
| Acidic phospholipase A2 S1E6-b (C. rhodostoma) | Q9PVF0 | 6 | 5 | ||
| Acidic phospholipase A2 Ts-A4 (T. stejnegeri) | Q6H3C7 | 11 | 10 | ||
| Basic phospholipase A2 W6D49 (C. rhodostoma) | Q9PVF4 | 21 | 6 | ||
| Phospholipase A2 (C. rhodostoma) | A0A0H3U266 | 23 | 6 | ||
| Phospholipase A2 (T. sabahi) | A0A0H3U232 | 3 | 2 | ||
| PLB | Phospholipase B (O. okinavensis) | 2 | T2HQ75 | 3 | 2 |
| Phospholipase B (P. regius) | A0A098LY74 | 5 | 3 | ||
| QPCT | Glutaminyl-peptide cyclotransferase (A. contortrix contortrix) | 1 | A0A1W7RH88 | 2 | 2 |
| SVMP | Metalloprotease, mRNA (G. intermedius) | 5 | A0A0C4ZNF1 | 5 | 2 |
| Snake venom metalloproteinase kistomin (C. rhodostoma) | P0CB14 | 86 | 86 | ||
| Zinc metalloproteinase/disintegrin (C. rhodostoma) | P30403 | 38 | 36 | ||
| Zinc metalloproteinase-disintegrin-like halysase (G. halys) | Q8AWI5 | 4 | 2 | ||
| Zinc metalloproteinase-disintegrin-like HV1 (P. flavoviridis) | Q90ZI3 | 4 | 2 | ||
| SVSP | Serine protease 1 (P. elegans) | 4 | A0A077LA46 | 3 | 2 |
| Thrombin-like enzyme ancrod (C. rhodostoma) | P26324 | 21 | 20 | ||
| Thrombin-like enzyme ancrod-2 (C. rhodostoma) | P47797 | 17 | 13 | ||
| Thrombin-like enzyme elegaxobin-2 (P. elegans) | P84787 | 3 | 2 | ||
| Trypsinogen | Trypsinogen homolog (B. jararaca) | 1 | Q9PUF3 | 3 | 3 |
| Others | Ankyrin repeat-containing protein (A. contortrix contortrix) | 1 | A0A1W7RJF1 | 2 | 2 |
| TOTAL PROTEINS | 47 |
| Protein Family | Protein Description | No. of Proteins | Protein Accession No. | No. of Peptides | No. of Unique Peptides |
|---|---|---|---|---|---|
| 3FTx | Alpha-elapitoxin-Dpp2d (D. polylepis polylepis) | 23 | C0HJD7 | 2 | 2 |
| Beta-cardiotoxin CTX27 (O. hannah) | Q69CK0 | 11 | 3 | ||
| Haditoxin (O. hannah) | A8N286 | 3 | 3 | ||
| Long neurotoxin 1 (O. hannah) | P01387 | 8 | 2 | ||
| Long neurotoxin 1 (N. nivea) | P01390 | 2 | 2 | ||
| Long neurotoxin 2 (O. hannah) | P01386 | 7 | 2 | ||
| Long neurotoxin 4 (O. hannah) | P80156 | 2 | 2 | ||
| Long neurotoxin LNTX1 (O. hannah) | Q2VBP8 | 4 | 2 | ||
| Long neurotoxin LNTX-2 homolog (O. hannah) | A8N285 | 8 | 2 | ||
| Long neurotoxin LNTX37 (O. hannah) | Q2VBP3 | 3 | 2 | ||
| Long neurotoxin OH-5 (O. hannah) | P80965 | 2 | 2 | ||
| Long neurotoxin OH-37 (O. hannah) | Q53B59 | 7 | 7 | ||
| Long neurotoxin OH-55 (O. hannah) | Q53B58 | 13 | 6 | ||
| Long neurotoxin OH-56 (O. hannah) | Q53B57 | 4 | 4 | ||
| Long neurotoxin OH-57 (O. hannah) | Q53B56 | 3 | 3 | ||
| Neurotoxin-like protein 1 (C. rhombeatus) | P84716 | 2 | 2 | ||
| Short neurotoxin OH-35 (O. hannah) | Q53B49 | 5 | 5 | ||
| Weak toxin DE-1 (O. hannah) | P01412 | 36 | 29 | ||
| Weak toxin DE-1 homolog 1 (O. hannah) | Q69CJ8 | 2 | 2 | ||
| Weak neurotoxin OH-72 (O. hannah) | Q53B61 | 4 | 3 | ||
| Weak neurotoxin WNTX33 (O. hannah) | Q2VBN3 | 23 | 16 | ||
| Weak neurotoxin WNTX34 (O. hannah) | Q2VBN2 | 5 | 4 | ||
| Uncharacterized protein (O. hannah)—Toxin activity | V8N3G9 | 5 | 5 | ||
| 5′-NTD | Ecto-5′-nucleotidase (M. tener) | 3 | A0A194AS98 | 3 | 2 |
| Ecto-5′-nucleotidase 1 (M. fulvius) | U3FYP9 | 3 | 2 | ||
| Ecto-5′-nucleotidase 1a (M. fulvius) | A0A0F7YZM6 | 3 | 2 | ||
| AChE | Acetylcholinesterase (B. fasciatus) | 1 | Q92035 | 5 | 5 |
| CC3H | O. venom factor (O. hannah) | 1 | I2C090 | 8 | 6 |
| CRiSP | Cysteine-rich venom protein (T. stejnegeri) | 4 | P60623 | 2 | 2 |
| Cysteine-rich venom protein ablomin (G. blomhoffii) | Q8JI40 | 2 | 2 | ||
| Cysteine-rich venom protein mossambin (N. mossambica) | P0DL16 | 2 | 2 | ||
| Cysteine-rich venom protein ophanin (O. hannah) | Q7ZT98 | 13 | 12 | ||
| CTL/snaclec | Snaclec rhodocetin subunit beta (C. rhodostoma) | 1 | P81398 | 2 | 2 |
| Endonuclease | Endonuclease domain-containing 1 protein (O. hannah) | 1 | V8NCE3 | 2 | 2 |
| FMO | Amine oxidase (O. hannah) | 3 | V8N3Q9 | 29 | 2 |
| L-amino-acid oxidase (O. hannah) | P81383 | 16 | 16 | ||
| L-amino-acid oxidase (C. rhodostoma) | P81382 | 8 | 7 | ||
| Insulin | Insulin-like growth factor 1 (P. guttatus) | 1 | I6TIH6 | 7 | 6 |
| Kunitz-type | Kunitz-type serine protease inhibitor TCI (O. hannah) | 5 | B6RLX2 | 13 | 11 |
| Kunitz-type serine protease inhibitor Vur-Kin (V. renardi) | P0DKL8 | 2 | 2 | ||
| Kunitz trypsin inhibitor protein 2 (V. renardi) | S4S375 | 2 | 2 | ||
| Venom Kunitz trypsin inhibitor protein 2 (V. renardi) | S4S374 | 2 | 2 | ||
| Uncharacterized protein (O. hannah)—Peptidase inhibitor activity | V8NAQ0 | 2 | 2 | ||
| Neprilysin | Neprilysin (B. irregularis) | 1 | A0A0B8RU83 | 5 | 2 |
| NGF | Venom nerve growth factor (W. aegyptia) | 2 | V9I1F9 | 2 | 2 |
| Uncharacterized protein (O. hannah) | V8N4D8 | 2 | 2 | ||
| Ohanin/Vespryn | Ohanin (O. hannah) | 1 | P83234 | 5 | 5 |
| PDE | Ectonucleotide pyrophosphatase/phosphodiesterase 3 (M. fulvius) | 3 | U3FAB3 | 9 | 7 |
| Phosphodiesterase (M. fulvius) | A0A0F7YYZ8 | 9 | 7 | ||
| Phosphodiesterase (M. tener) | A0A194ARD7 | 9 | 7 | ||
| PDGF/VEGF | Uncharacterized protein (O. hannah) | 1 | V8P6V3 | 5 | 5 |
| PLA2 | Acidic phospholipase A2 2 (O. hannah) | 2 | Q9DF33 | 4 | 4 |
| Basic phospholipase A2 (C. rhodostoma) | Q9PVF4 | 2 | 2 | ||
| SVGF | Hepatocyte growth factor activator (O. hannah) | 1 | V8NIY2 | 2 | 2 |
| SVMP | Metalloproteinase (type III) 2 (Hypsiglena sp.) | 21 | A0A098M156 | 3 | 2 |
| Metalloproteinase type III 14a (Hypsiglena sp.) | A0A098M221 | 3 | 2 | ||
| Metalloproteinase type III 14b (Hypsiglena sp.) | A0A098M219 | 3 | 2 | ||
| Metalloproteinase (type III) 14c (Hypsiglena sp.) | A0A098M223 | 3 | 2 | ||
| Metalloproteinase type III 2 (M. tener) | A0A194AS47 | 8 | 2 | ||
| Scutellatease-1 (O. scutellatus) | B5KFV5 | 4 | 3 | ||
| Snake venom metalloproteinase BaP1 (B. asper) | P83512 | 3 | 2 | ||
| Snake venom metalloproteinase BjussuMP-2 (B. jararacussu) | Q7T1T4 | 3 | 2 | ||
| Snake venom metalloproteinase BmooMPalpha-I (B. moojeni) | P85314 | 3 | 2 | ||
| Snake venom metalloproteinase bothrojaractivase (B. jararaca) | P0C7A9 | 3 | 2 | ||
| Snake venom metalloproteinase kistomin (C. rhodostoma) | P0CB14 | 14 | 14 | ||
| Snake venom metalloproteinase leucurolysin-A (B. leucurus) | P84907 | 3 | 2 | ||
| Zinc metalloproteinase/disintegrin (B. asper) | Q072L5 | 3 | 2 | ||
| Zinc metalloproteinase/disintegrin (B. insularis) | Q5XUW8 | 3 | 2 | ||
| Zinc metalloproteinase/disintegrin (C. rhodostoma) | P30403 | 5 | 5 | ||
| Zinc metalloproteinase-disintegrin-like atrase-A (N. atra) | D5LMJ3 | 4 | 2 | ||
| Zinc metalloproteinase-disintegrin-like atrase-B (N. atra) | D6PXE8 | 5 | 2 | ||
| Zinc metalloproteinase-disintegrin-like BjussuMP-1 (B. jararacussu) | Q1PHZ4 | 3 | 2 | ||
| Zinc metalloproteinase-disintegrin-like kaouthiagin (N. atra) | D3TTC1 | 8 | 4 | ||
| Zinc metalloproteinase-disintegrin-like MTP9 (D. coronoides) | F8RKV9 | 6 | 2 | ||
| Zinc metalloproteinase-disintegrin-like ohanin (O. hannah) | A3R0T9 | 5 | 5 | ||
| SVSP | Alpha and beta fibrinogenase OhS1 (O. hannah) | 1 | A8QL56 | 2 | 2 |
| TOTAL PROTEINS | 76 |
| Protein Family | Protein Description | No. of Proteins | Protein Accession No. | No. of Peptides | No. of Unique Peptides |
|---|---|---|---|---|---|
| 5′-NTD | Snake venom 5′-nucleotidase (B. irregularis) | 4 | A0A0B8RXZ9 | 12 | 3 |
| 5′-nucleotidase (P. flavoviridis) | T2HRS9 | 4 | 4 | ||
| 5′-nucleotidase (P. elegans) | A0A077L6M5 | 5 | 4 | ||
| 5′-nucleotidase (P. flavoviridis) | A0A077L7M9 | 4 | 4 | ||
| Aminopeptidase | Aminopeptidase (C. horridus) | 2 | T1DNX8 | 4 | 4 |
| Aminopeptidase (C. horridus) | A0A0K8S2L4 | 4 | 4 | ||
| CRiSP | Cysteine-rich secretory protein LCCL … 2 (C. adamanteus) | 1 | A0A0F7ZEB6 | 9 | 9 |
| CTL/snaclec | Snaclec alboaggregin-D subunit alpha (T. albolabris) | 8 | P0DM38 | 4 | 3 |
| Snaclec rhodocetin subunit alpha (C. rhodostoma) | P81397 | 58 | 57 | ||
| Snaclec rhodocetin subunit beta (C. rhodostoma) | P81398 | 54 | 53 | ||
| Snaclec rhodocetin subunit gamma (C. rhodostoma) | D2YW39 | 25 | 23 | ||
| Snaclec rhodocetin subunit delta (C. rhodostoma) | D2YW40 | 22 | 22 | ||
| Snaclec rhodocytin subunit alpha (C. rhodostoma) | Q9I841 | 12 | 11 | ||
| Snaclec rhodocytin subunit beta (C. rhodostoma) | Q9I840 | 10 | 8 | ||
| C-type lectin 12a (A. contortrix contortrix) | A0A1W7RJZ7 | 7 | 7 | ||
| Endonuclease | Endonuclease domain-containing protein (O. hannah) | 3 | U3FCT9 | 2 | 2 |
| Endonuclease domain-containing protein (O. hannah) | V8NCE3 | 2 | 2 | ||
| Deoxyribonuclease-2-alpha-like (C. adamanteus)—DNase | J3RZ14 | 4 | 4 | ||
| FMO | Amine oxidase (O. okinavensis) | 8 | T2HQ57 | 40 | 4 |
| Amine oxidase (P. regius) | A0A098LWS4 | 12 | 3 | ||
| Amine oxidase (P. guttatus) | A0A098LX00 | 11 | 2 | ||
| L-amino-acid oxidase (C. rhodostoma) | P81382 | 147 | 99 | ||
| L-amino acid oxidase (B. schlegelii) | A0A024BTN9 | 46 | 10 | ||
| L-amino-acid oxidase (T. stejnegeri) | Q6WP39 | 19 | 2 | ||
| L-amino-acid oxidase (L. muta) | J7H670 | 15 | 2 | ||
| L-amino-acid oxidase (B. fasciatus) | A8QL52 | 10 | 2 | ||
| NGF | Venom nerve growth factor (B. jararacussu) | 4 | Q90W38 | 3 | 2 |
| Nerve growth factor (P. flavoviridis) | B1Q3K2 | 5 | 5 | ||
| Nerve growth factor (P. flavoviridis) | A0A077L854 | 5 | 5 | ||
| BATXNGF1 (B. atrox) | A0A1L8D608 | 3 | 2 | ||
| PDE | Cadam 10_PDE-1 (C. adamanteus) | 18 | A0A1W7RBB0 | 12 | 2 |
| Ectonucleotide pyrophosphatase/phosphodiesterase 3 (M. fulvius) | U3FAB3 | 5 | 2 | ||
| Ectonucleotide pyrophosphatase/phosphodiesterase 3 (C. horridus) | T1DJT5 | 12 | 3 | ||
| Phosphodiesterase (P. flavoviridis) | T2HQA0 | 17 | 2 | ||
| Phosphodiesterase (P. flavoviridis) | T2HRT4 | 17 | 2 | ||
| Phosphodiesterase (P. flavoviridis) | T2HPD6 | 17 | 2 | ||
| Phosphodiesterase (P. flavoviridis) | T2HP62 | 17 | 2 | ||
| Phosphodiesterase (M. fulvius) | A0A0F7YYZ8 | 5 | 2 | ||
| Phosphodiesterase (M. tener) | A0A194ARD7 | 5 | 2 | ||
| Phosphodiesterase (M. fulvius) | A0A0F7YYZ8 | 3 | 2 | ||
| Phosphodiesterase (C. adamanteus) | A0A0F7Z2Q3 | 12 | 3 | ||
| Phosphodiesterase (C. horridus) | T1D6P7 | 12 | 3 | ||
| Phosphodiesterase (S. miliarius barbouri) | A0A194AS02 | 12 | 3 | ||
| Phosphodiesterase (M. tener) | A0A194ARD7 | 3 | 2 | ||
| Phosphodiesterase (M. rudis) | A0A141DWM1 | 4 | 3 | ||
| Phosphodiesterase (M. lebetina) | W8E7D1 | 4 | 2 | ||
| Venom phosphodiesterase 1 (C. adamanteus) | J3SEZ3 | 12 | 2 | ||
| Venom phosphodiesterase 2 (C. adamanteus) | J3SBP3 | 12 | 2 | ||
| PLA2 | Acidic phospholipase A2 (C. rhodostoma) | 10 | Q9PVF0 | 4 | 3 |
| Acidic phospholipase A2 (C. rhodostoma) | Q9PVF1 | 16 | 4 | ||
| Acidic phospholipase A2 (C. rhodostoma) | Q9PVF2 | 57 | 54 | ||
| Acidic phospholipase A2 (C. rhodostoma) | Q9PVE9 | 6 | 2 | ||
| Acidic phospholipase A2 (O. hannah) | Q9DF33 | 5 | 5 | ||
| Acidic phospholipase A2 (T. stejnegeri) | Q6H3C7 | 34 | 32 | ||
| Basic phospholipase A2 W6D49 (C. rhodostoma) | Q9PVF4 | 25 | 11 | ||
| Phospholipase A2 (C. rhodostoma) | A0A0H3U266 | 41 | 19 | ||
| Phospholipase A2 (C. atrox) | A0A1J0R081 | 5 | 2 | ||
| Phospholipase A2 1c (C. horridus) | A0A0K8RZ17 | 5 | 2 | ||
| PLB | Phospholipase B (C. adamanteus) | 6 | F8S101 | 16 | 5 |
| Phospholipase B (C. adamanteus) | A0A0F7Z632 | 16 | 5 | ||
| Phospholipase B (P. regius) | A0A098LY74 | 8 | 2 | ||
| Phospholipase B (P. guttatus) | A0A098LWY9 | 4 | 2 | ||
| Phospholipase B (P. flavoviridis) | T2HP68 | 7 | 5 | ||
| Phospholipase B (P. elegans) | A0A077L7E7 | 7 | 5 | ||
| SVMP | AAV1 protein (D. acutus) | 23 | A0A0M4MEY5 | 4 | 2 |
| BATXSVMPIII (B. atrox) | A0A1L8D5X9 | 3 | 2 | ||
| Disintegrin triflavin (P. flavoviridis) | P21859 | 8 | 6 | ||
| Metalloprotease, mRNA (G. intermedius) | A0A0C4ZNF1 | 8 | 2 | ||
| Metalloproteinase type III 2 (M. tener) | A0A194AS47 | 3 | 2 | ||
| Metalloproteinase type III 2 (M. tener) | A0A194AR91 | 7 | 3 | ||
| Metalloproteinase type III 2 (Hypsiglena sp.) | A0A098M156 | 2 | 2 | ||
| Metalloproteinase type III 14c (Hypsiglena sp.) | A0A098M223 | 2 | 2 | ||
| Metalloproteinase type III 5 (Hypsiglena sp.) | A0A098M215 | 2 | 2 | ||
| p-III snake venom metalloprotease (M. ikaheca) | A0A024AXX7 | 5 | 2 | ||
| Snake venom metalloproteinase kistomin (C. rhodostoma) | P0CB14 | 198 | 197 | ||
| SVMP-disintegrin-like mocarhagin (N. mossambica) | Q10749 | 8 | 4 | ||
| SVMP-Hop-15 (H. bungaroides) | R4FJZ4 | 5 | 2 | ||
| Zinc metalloproteinase/disintegrin (C. rhodostoma) | P30403 | 58 | 58 | ||
| Zinc metalloproteinase-disintegrin-like agkihagin (D. acutus) | Q1PS45 | 8 | 2 | ||
| Zinc metalloproteinase-disintegrin-like atragin (N. atra) | D3TTC2 | 4 | 2 | ||
| Zinc metalloproteinase-disintegrin-like atrase-A (N. atra) | D5LMJ3 | 4 | 2 | ||
| Zinc metalloproteinase-disintegrin-like BfMP (B. fasciatus) | A8QL48 | 7 | 7 | ||
| Zinc metalloproteinase-disintegrin-like BmMP (B. multicinctus) | A8QL49 | 7 | 7 | ||
| Zinc metalloproteinase-disintegrin-like cobrin (N. kaouthia) | Q9PVK7 | 5 | 2 | ||
| Zinc metalloproteinase-disintegrin-like halysase (G. halys) | Q8AWI5 | 8 | 2 | ||
| Zinc metalloproteinase-disintegrin-like HV1 (P. flavoviridis) | Q90ZI3 | 9 | 2 | ||
| Zinc metalloproteinase-disintegrin-like ohanin (O. hannah) | A3R0T9 | 12 | 12 | ||
| SVSP | Snake venom serine protease catroxase (C. atrox) | 12 | Q8QHK2 | 3 | 2 |
| Snake venom serine proteinase (C. adamanteus) | F8S116 | 3 | 2 | ||
| Serine protease (P. flavoviridis) | A0A077L6P3 | 12 | 2 | ||
| Serine proteinase 3 (A. piscivorus) | A0A194APA8 | 3 | 2 | ||
| Serine proteinase 3a (A. contortrix contortrix) | A0A1W7RJX1 | 3 | 2 | ||
| Serine proteinase 4 (A. contortrix contortrix) | A0A1W7RJU2 | 3 | 2 | ||
| Serine proteinase 9 (A. contortrix contortrix) | A0A1W7RJU0 | 3 | 2 | ||
| Serine proteinase 10 (C. horridus) | T1DH10 | 5 | 2 | ||
| Serine proteinase 12a (A. piscivorus) | A0A194APD3 | 3 | 2 | ||
| Thrombin-like enzyme ancrod (C. rhodostoma) | P26324 | 64 | 59 | ||
| Thrombin-like enzyme ancrod-2 (C. rhodostoma) | P47797 | 39 | 37 | ||
| Thrombin-like protein (A. piscivorus leucostoma) | E9NX14 | 3 | 2 | ||
| TOTAL PROTEINS | 99 |
| Protein Family | Protein Description | No. of Proteins | Protein Accession No. | No. of Peptides | No. of Unique Peptides |
|---|---|---|---|---|---|
| 3FTx | Alpha-cobratoxin (N. kaouthia) | 51 | P01391 | 4 | 2 |
| Alpha-elapitoxin-Dpp2d (D. polylepis polylepis) | C0HJD7 | 3 | 2 | ||
| Alpha-elapitoxin-Dv2a (D. viridis) | P01395 | 3 | 2 | ||
| Alpha-elapitoxin-Nno2a (N. oxiana) | P01382 | 5 | 4 | ||
| Alpha-elapitoxin-Oh2b (O. hannah) | P82662 | 15 | 4 | ||
| Haditoxin (O. hannah) | A8N286 | 16 | 16 | ||
| Kappa-6-bungarotoxin (B. multicinctus) | Q9W729 | 3 | 2 | ||
| Long neurotoxin 1 (A. superbus) | A8S6A8 | 3 | 2 | ||
| Long neurotoxin 1 (N. annulata annulata) | P34074 | 9 | 2 | ||
| Long neurotoxin 1 (N. haje haje) | P25674 | 9 | 4 | ||
| Long neurotoxin 1 (N. nivea) | P01390 | 7 | 4 | ||
| Long neurotoxin 1 (O. hannah) | P01387 | 26 | 6 | ||
| Long neurotoxin 2 (O. hannah) | P01386 | 5 | 5 | ||
| Long neurotoxin 2 (N. melanoleuca) | P01388 | 8 | 3 | ||
| Long neurotoxin 3 (O. hannah) | P07526 | 5 | 2 | ||
| Long neurotoxin 4 (O. hannah) | P80156 | 14 | 3 | ||
| Long neurotoxin LlLong (L. laticaudata) | Q7T2I3 | 3 | 2 | ||
| Long neurotoxin OH-17 (O. hannah) | Q53B54 | 8 | 2 | ||
| Long neurotoxin OH-37 (O. hannah) | Q53B59 | 20 | 9 | ||
| Long neurotoxin OH-55 (O. hannah) | Q53B58 | 44 | 21 | ||
| Long neurotoxin OH-56 (O. hannah) | Q53B57 | 18 | 15 | ||
| Long neurotoxin OH-57 (O. hannah) | Q53B56 | 7 | 5 | ||
| Long neurotoxin LNTX1 (O. hannah) | Q2VBP8 | 13 | 5 | ||
| Long neurotoxin LNTX-2 (O. hannah) | A8N285 | 14 | 3 | ||
| Long neurotoxin LNTX8 (O. hannah) | Q2VBP6 | 21 | 3 | ||
| Long neurotoxin LNTX22 (O. hannah) | Q2VBP5 | 5 | 3 | ||
| Non-conventional three finger toxin isoform (B. flaviceps) | D5J9P9 | 4 | 2 | ||
| Oxiana weak toxin (N. oxiana) | P85520 | 3 | 3 | ||
| Short neurotoxin OH-35 (O. hannah) | Q53B49 | 5 | 5 | ||
| Three-finger toxin (B. multicinctus) | E2IU12 | 2 | 2 | ||
| Three-finger toxin (B. multicinctus) | E2IU14 | 2 | 2 | ||
| Weak neurotoxin (N. N.) | P29181 | 2 | 2 | ||
| Weak neurotoxin OH-72 (O. hannah) | Q53B61 | 13 | 10 | ||
| Weak neurotoxin WNTX33 (O. hannah) | Q2VBN3 | 65 | 55 | ||
| Weak neurotoxin WNTX34 (O. hannah) | Q2VBN2 | 13 | 10 | ||
| Weak neurotoxin 7 (N. N.) | P29181 | 4 | 3 | ||
| Weak neurotoxin 10 (N. sputatrix) | Q802B2 | 2 | 2 | ||
| Weak toxin CM-11 (N. haje haje) | P01401 | 3 | 3 | ||
| Weak toxin DE-1 (O. hannah) | P01412 | 117 | 110 | ||
| Weak toxin DE-1 homolog 1 (O. hannah) | Q69CJ8 | 12 | 12 | ||
| Weak toxin S4C11 (N. melanoleuca) | P01400 | 3 | 3 | ||
| Putative long neurotoxin (A. labialis) | B2BRQ8 | 3 | 2 | ||
| Putative long neurotoxin (A. labialis) | B2BRQ9 | 3 | 2 | ||
| Putative long neurotoxin (A. labialis) | B2BRR0 | 3 | 2 | ||
| Putative long neurotoxin (A. labialis) | B2BRR1 | 3 | 2 | ||
| Putative long neurotoxin (A. labialis) | B2BRR2 | 3 | 2 | ||
| Putative long neurotoxin (A. labialis) | B2BRR3 | 3 | 2 | ||
| Putative long neurotoxin (A. labialis) | B2BRR6 | 3 | 2 | ||
| Putative long neurotoxin (A. labialis) | B2BRR8 | 3 | 2 | ||
| Uncharacterized protein (O. hannah)—Toxin activity | V8N389 | 7 | 5 | ||
| Uncharacterized protein (O. hannah)—Toxin activity | V8N3G9 | 5 | 5 | ||
| AChE | Carboxylic ester hydrolase (S. fasciata) | 1 | R4FKE6 | 2 | 2 |
| CC3H | Complement-depleting factor (O. hannah) | 7 | A8QL55 | 8 | 8 |
| O. venom factor (O. hannah) | I2C090 | 34 | 23 | ||
| Venom factor (C. horridus) | T1E3W8 | 2 | 2 | ||
| Venom factor (Sistrurus catenatus tergeminus) | A0A194APJ4 | 2 | 2 | ||
| Venom factor (Agkistrodon piscivorus) | A0A194ARG1 | 2 | 2 | ||
| Venom factor (C. adamanteus) | A0A0F7Z1I7 | 2 | 2 | ||
| Venom factor (C. adamanteus) | J3S836 | 2 | 2 | ||
| CRiSP | Cysteine-rich venom protein (P. olfersii) | 13 | Q09GJ9 | 3 | 2 |
| Cysteine-rich venom protein mossambin (N. mossambica) | P0DL16 | 9 | 2 | ||
| Cysteine-rich venom protein ophanin (O. hannah) | Q7ZT98 | 44 | 14 | ||
| Cysteine-rich venom protein TRI1 (T. biscutatus) | Q2XXP4 | 4 | 2 | ||
| Cysteine-rich secretory protein (O. okinavensis) | T2HPR8 | 7 | 2 | ||
| Cysteine-rich secretory protein (P. chamissonis) | A0A0B4SXI8 | 2 | 2 | ||
| Cysteine-rich secretory protein Az-CRP (A. feae) | F2Q6E3 | 6 | 2 | ||
| Cysteine-rich secretory protein Bc-CRP (B. schlegelii) | F2Q6E4 | 7 | 2 | ||
| Cysteine-rich secretory protein Bc-CRPa (B. candidus) | F2Q6G3 | 3 | 3 | ||
| Cysteine-rich secretory protein Bc-CRPb (B. candidus) | F2Q6G2 | 2 | 2 | ||
| Cysteine-rich secretory protein 1b (B. irregularis) | A0A0B8RZW7 | 3 | 2 | ||
| Cysteine-rich secretory protein 1c (B. irregularis) | A0A0B8RYV8 | 3 | 2 | ||
| Uncharacterized protein (O. hannah) | V8N8B4 | 35 | 5 | ||
| CTL/snaclec | Snaclec rhodocetin subunit alpha (C. rhodostoma) | 1 | P81397 | 2 | 2 |
| Endonuclease | Endonuclease domain-containing protein (O. hannah) | 2 | V8NCE3 | 3 | 2 |
| Endonuclease domain-containing protein (O. hannah) | V8N4Y2 | 3 | 2 | ||
| FMO | Amine oxidase (O. hannah) | 2 | V8N3Q9 | 52 | 8 |
| L-amino-acid oxidase (O. hannah) | P81383 | 59 | 15 | ||
| Insulin | Insulin-like growth factor 1 (P. guttatus) | 5 | I6TIH6 | 19 | 15 |
| Insulin-like growth factor I (O. hannah) | V8NHJ8 | 7 | 5 | ||
| Insulin-like growth factor 1 (C. helena) | I6S9K4 | 7 | 2 | ||
| Insulin-like growth factor 1 (T. elegans) | I6T666 | 9 | 9 | ||
| Uncharacterized protein (O. hannah)—Hormone activity | V8N2J2 | 4 | 4 | ||
| Kunitz-type | Kunitz-type serine protease inhibitor (O. hannah) | 6 | P82966 | 7 | 3 |
| Kunitz-type serine protease inhibitor TCI (O. hannah) | B6RLX2 | 31 | 27 | ||
| Kunitz-type serine protease inhibitor Vur-KIn (V. renardi) | P0DKL8 | 4 | 3 | ||
| Kunitz trypsin inhibitor protein 2 (V. renardi) | S4S375 | 4 | 3 | ||
| Venom Kunitz trypsin inhibitor protein 2 (V. renardi) | S4S374 | 4 | 3 | ||
| Uncharacterized protein (O. hannah)—Peptidase inhibitor activity | V8NAQ0 | 4 | 4 | ||
| NGF | Uncharacterized protein (O. hannah) | 1 | V8N4D8 | 2 | 2 |
| Ohanin/Vespryn | Ohanin (O. hannah) | 2 | P83234 | 22 | 17 |
| Thaicobrin (N. kaouthia) | P82885 | 2 | 2 | ||
| PDE | Cadam 10_PDE-1 (C. adamanteus) | 11 | A0A1W7RBB0 | 3 | 2 |
| Ectonucleotide pyrophosphatase/phosphodiesterase 3 (M. fulvius) | U3FAB3 | 11 | 11 | ||
| Ectonucleotide pyrophosphatase/phosphodiesterase 3 (C. horridus) | T1DJT5 | 3 | 2 | ||
| Phosphodiesterase (M. tener) | A0A194ARD7 | 11 | 11 | ||
| Phosphodiesterase (M. fulvius) | A0A0F7YYZ8 | 11 | 11 | ||
| Phosphodiesterase (C. horridus) | T1D6P7 | 3 | 2 | ||
| Phosphodiesterase (C. adamanteus) | A0A0F7Z2Q3 | 3 | 2 | ||
| Phosphodiesterase (O. okinavensis) | U3TBJ5 | 3 | 2 | ||
| Phosphodiesterase (O. okinavensis) | U3TAI7 | 3 | 2 | ||
| Venom phosphodiesterase 1 (C. adamanteus) | J3SEZ3 | 3 | 2 | ||
| Venom phosphodiesterase 2 (C. adamanteus) | J3SBP3 | 3 | 2 | ||
| PDGF/VEGF | Uncharacterized protein (O. hannah) | 1 | V8P6V3 | 7 | 7 |
| PLA2 | Acidic phospholipase A2 2 (O. hannah) | 8 | Q9DF33 | 54 | 49 |
| Acidic phospholipase A2 (O. hannah) | Q9DF56 | 20 | 17 | ||
| Acidic phospholipase A2 KBf-grIB (B. fasciatus) | P0C551 | 2 | 2 | ||
| Phospholipase A2 12 (M. fulvius) | U3FVG1 | 5 | 2 | ||
| Phospholipase A2 23 (M. fulvius) | U3FVF5 | 5 | 2 | ||
| PLA2 (IB)-Tri1 (T. biscutatus) | A7X418 | 5 | 3 | ||
| Secretory phospholipase A2 (Group 10) (O. hannah) | V8PDP0 | 2 | 2 | ||
| Uncharacterized protein (O. hannah) | V8N1D8 | 2 | 2 | ||
| PLB | Phospholipase B-like 1 (O. hannah) | 1 | V8ND68 | 9 | 9 |
| SVGF | Hepatocyte growth factor activator (O. hannah) | 3 | V8NIY2 | 2 | 2 |
| Macrophage colony-stimulating factor 1 (O. hannah) | V8NW35 | 3 | 3 | ||
| Tumor necrosis factor receptor (O. hannah) | V8P0T5 | 5 | 5 | ||
| SVMP | Asrin (A. superbus) | 33 | A6XJS7 | 11 | 6 |
| BATXSVMPII2 (B. atrox) | A0A1L8D6A9 | 4 | 2 | ||
| BATXSVMPII1 (B. atrox) | A0A1L8D600 | 4 | 2 | ||
| BATXSVMPII3 (B. atrox) | A0A1L8D5Z6 | 4 | 2 | ||
| BATXSVMPIII9 (B. atrox) | A0A1L8D641 | 4 | 2 | ||
| Metalloproteinase (A. andersonii) | Q9PT47 | 6 | 2 | ||
| Metalloproteinase (D. vestigiata) | B5G6F7 | 6 | 2 | ||
| Metalloprotease BOJUMET III (B. jararacussu) | Q7T1T3 | 4 | 2 | ||
| Metalloproteinase PIII (T. gracilis) | V5IWF4 | 2 | 2 | ||
| Metalloproteinase type III 2 (Hypsiglena sp.) | A0A098M156 | 6 | 2 | ||
| Metalloproteinase type III 14b (Hypsiglena sp.) | A0A098M219 | 7 | 4 | ||
| Metalloproteinase type III 14c (Hypsiglena sp.) | A0A098M223 | 5 | 2 | ||
| Metalloproteinase type III 1 (M. fulvius) | U3EPC7 | 5 | 5 | ||
| Metalloproteinase type III 2 (M. tener) | A0A194AS47 | 16 | 4 | ||
| Metalloproteinase type III 2b (M. tener) | A0A0F7YYV1 | 5 | 2 | ||
| Scutellatease-1 (O. scutellatus) | B5KFV5 | 13 | 7 | ||
| SVMP-disintegrin-like mocarhagin (N. mossambica) | Q10749 | 9 | 5 | ||
| Snake venom metalloproteinase kistomin (C. rhodostoma) | P0CB14 | 4 | 4 | ||
| Snake venom metalloprotease (P.olfersii) | C6JUN3 | 4 | 2 | ||
| Snake venom metalloprotease (P. olfersii) | C6JUN4 | 4 | 2 | ||
| Stephensease-1 (H. stephensii) | B5KFV4 | 8 | 2 | ||
| SVMP-Aca-4 (A. wellsi) | R4G2D3 | 11 | 4 | ||
| SVMP-Den-9 (D. devisi) | R4FIC4 | 15 | 2 | ||
| SVMP-Ech-32 (E. curta) | R4FJM6 | 6 | 2 | ||
| SVMP-Fur-1 (F. ornata) | R4G2G3 | 4 | 2 | ||
| SVMP-Hem-2 (H. signata) | R4G2W9 | 8 | 3 | ||
| SVMP-Hop-14 (H. bungaroides) | R4FIM1 | 5 | 3 | ||
| Zinc metalloproteinase-disintegrin-like atrase-A (N. atra) | D5LMJ3 | 14 | 3 | ||
| Zinc metalloproteinase-disintegrin-like atrase-B (N. atra) | D6PXE8 | 9 | 3 | ||
| Zinc metalloproteinase-disintegrin-like cobrin (N. kaouthia) | Q9PVK7 | 7 | 2 | ||
| Zinc metalloproteinase-disintegrin-like kaouthiagin-like (N. atra) | D3TTC1 | 9 | 3 | ||
| Zinc metalloproteinase-disintegrin-like MTP4 (D. coronoides) | F8RKW1 | 11 | 3 | ||
| Zinc metalloproteinase-disintegrin-like ohanin (O. hannah) | A3R0T9 | 19 | 10 | ||
| SVSP | Alpha and beta fibrinogenase OhS1 (O. hannah) | 2 | A8QL56 | 6 | 4 |
| Neuroendocrine convertase 1 (O. hannah) | V8P1Y2 | 3 | 3 | ||
| TOTAL PROTEINS | 150 |
| Snake Venom Protein Families | Calloselasma rhodostoma | Ophiophagus hannah |
|---|---|---|
| 3FTx | ND | 31 (55) |
| Complement C3 Homolog | ND | 4 (7) |
| Kunitz-type | ND | 3 (6) |
| Insulin | ND | 3 (5) |
| SVGF | ND | 2 (3) |
| AChE | ND | 1 (2) |
| Ohanin/Vespryn | ND | 1 (2) |
| Neprilysin | ND | 1 (1) |
| PDGF/VEGF | ND | 1 (1) |
| SVMP | 20 (23) | 25 (44) |
| PDE | 16 (18) | 6 (11) |
| SVSP | 12 (14) | 1 (2) |
| CTL/snaclec | 12 (14) | 1 (2) |
| PLA2 | 10 (11) | 5 (9) |
| FMO | 7 (8) | 2 (3) |
| PLB | 6 (7) | 1 (1) |
| 5′-NTD | 5 (6) | 2 (3) |
| NGF | 4 (4) | 1 (2) |
| Endonuclease | 3 (3) | 1 (2) |
| CRiSP | 1 (1) | 9 (15) |
| Aminopeptidase | 2 (2) | ND |
| QPCT | 1 (1) | ND |
| Trypsinogen | 1 (1) | ND |
| Others (ANK) | 1 (1) | ND |
| TOTAL | 100 (114) | 100 (176) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kunalan, S.; Othman, I.; Syed Hassan, S.; Hodgson, W.C. Proteomic Characterization of Two Medically Important Malaysian Snake Venoms, Calloselasma rhodostoma (Malayan Pit Viper) and Ophiophagus hannah (King Cobra). Toxins 2018, 10, 434. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10110434
Kunalan S, Othman I, Syed Hassan S, Hodgson WC. Proteomic Characterization of Two Medically Important Malaysian Snake Venoms, Calloselasma rhodostoma (Malayan Pit Viper) and Ophiophagus hannah (King Cobra). Toxins. 2018; 10(11):434. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10110434
Chicago/Turabian StyleKunalan, Sugita, Iekhsan Othman, Sharifah Syed Hassan, and Wayne C. Hodgson. 2018. "Proteomic Characterization of Two Medically Important Malaysian Snake Venoms, Calloselasma rhodostoma (Malayan Pit Viper) and Ophiophagus hannah (King Cobra)" Toxins 10, no. 11: 434. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10110434