Development of an Immunoassay for Detection of Staphylococcal Enterotoxin-Like J, A Non-Characterized Toxin

Abstract
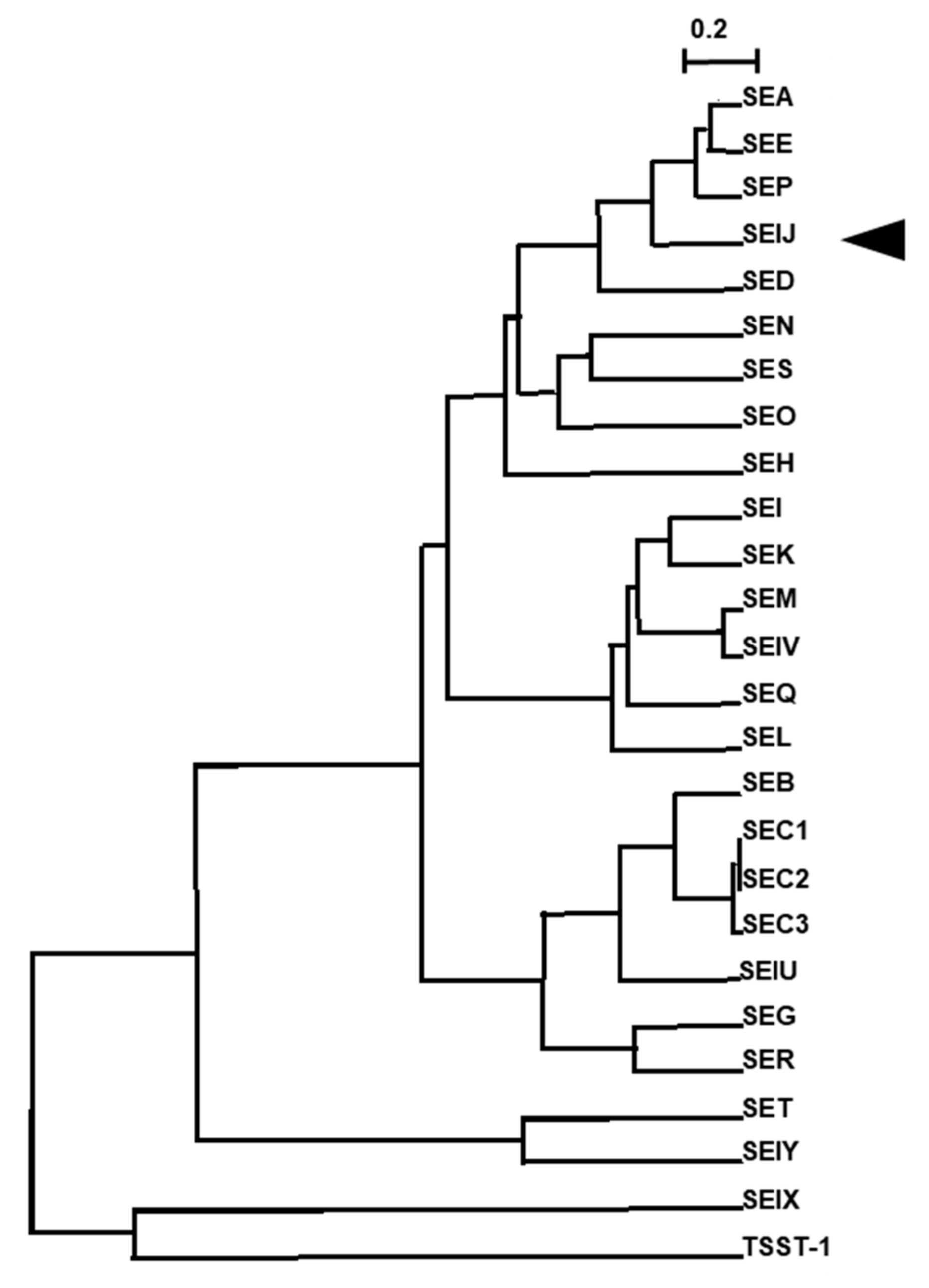
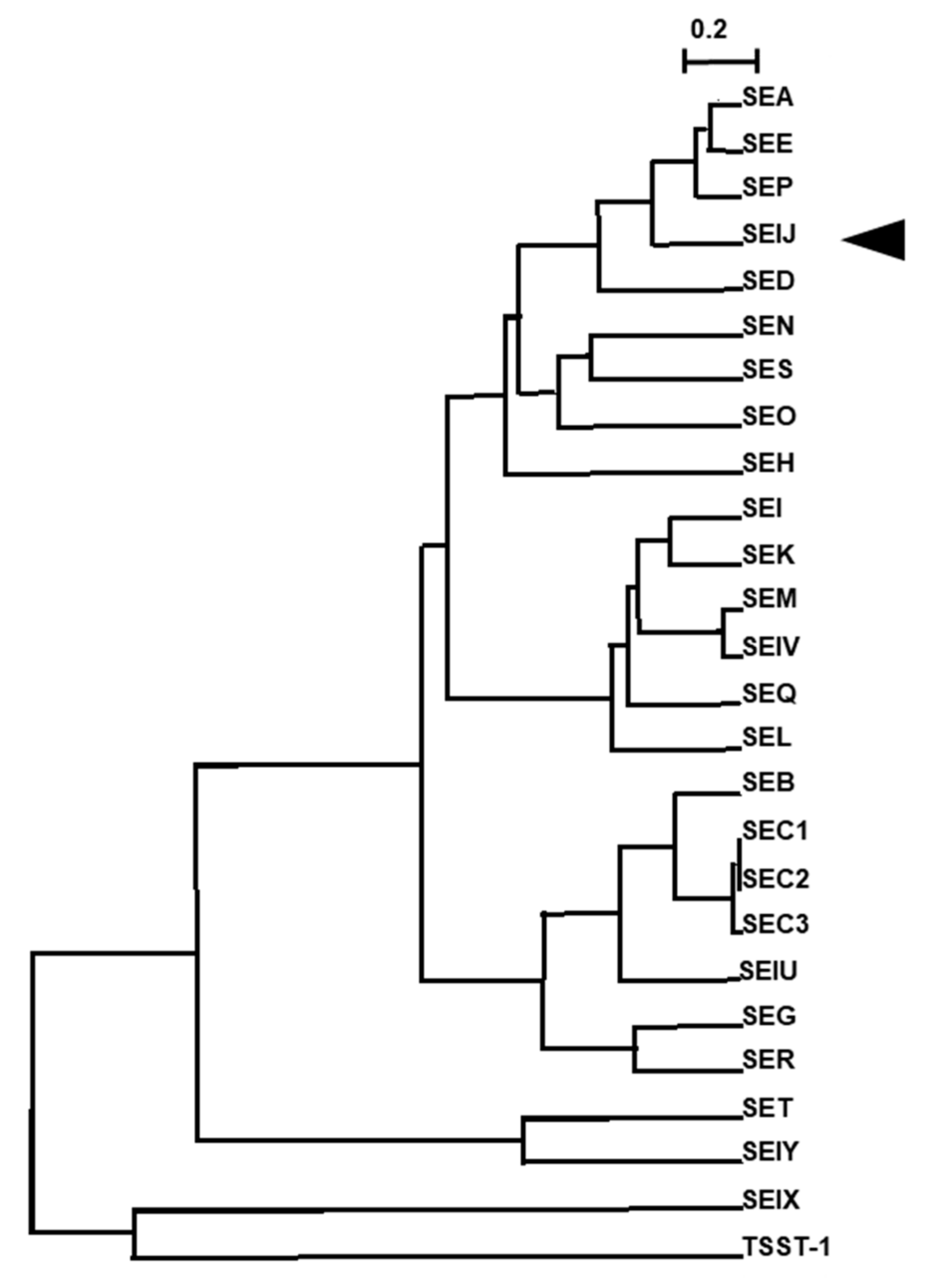
:1. Introduction
2. Results
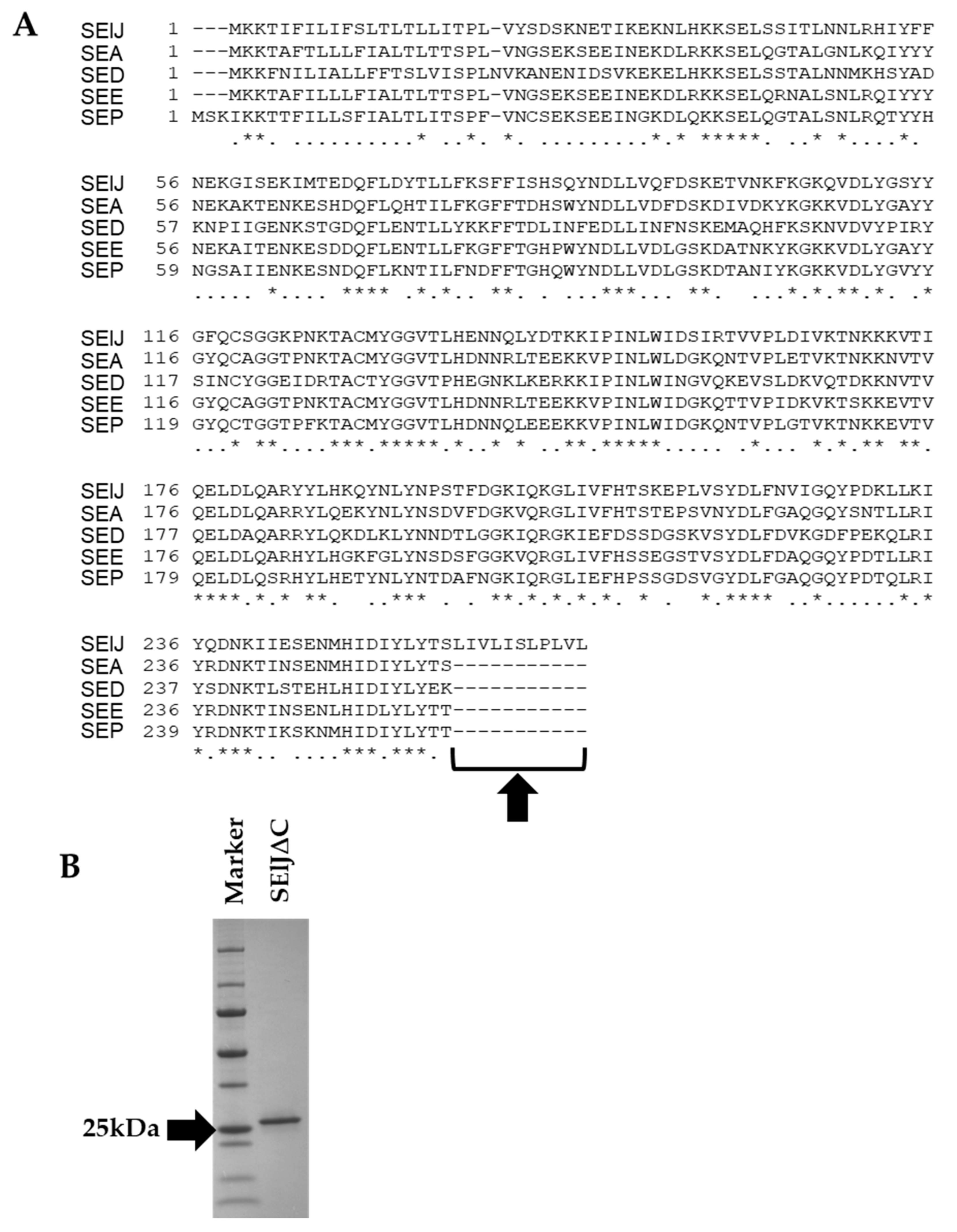
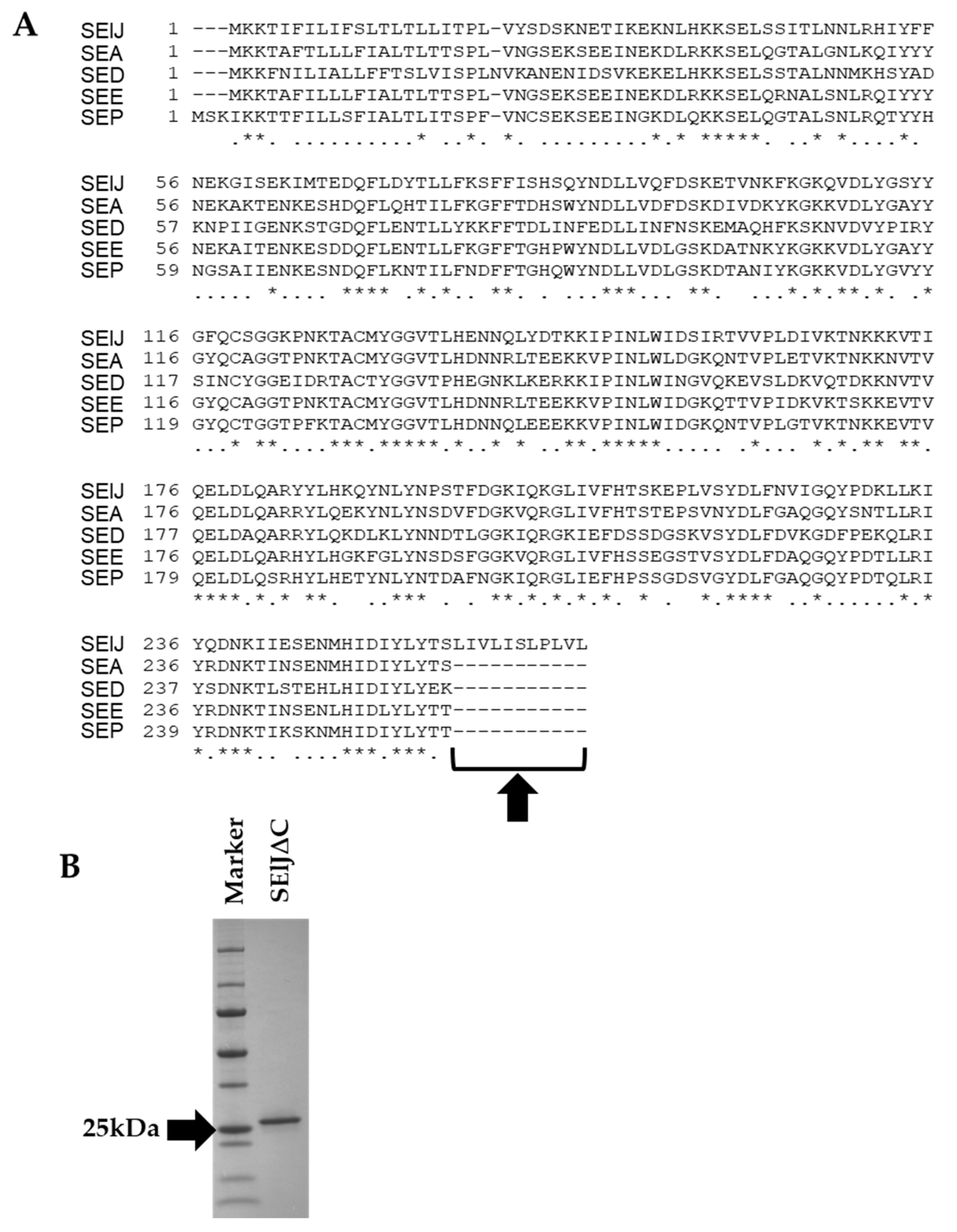
2.1. C-Terminus-Depleted SElJ Was Expressed and Purified
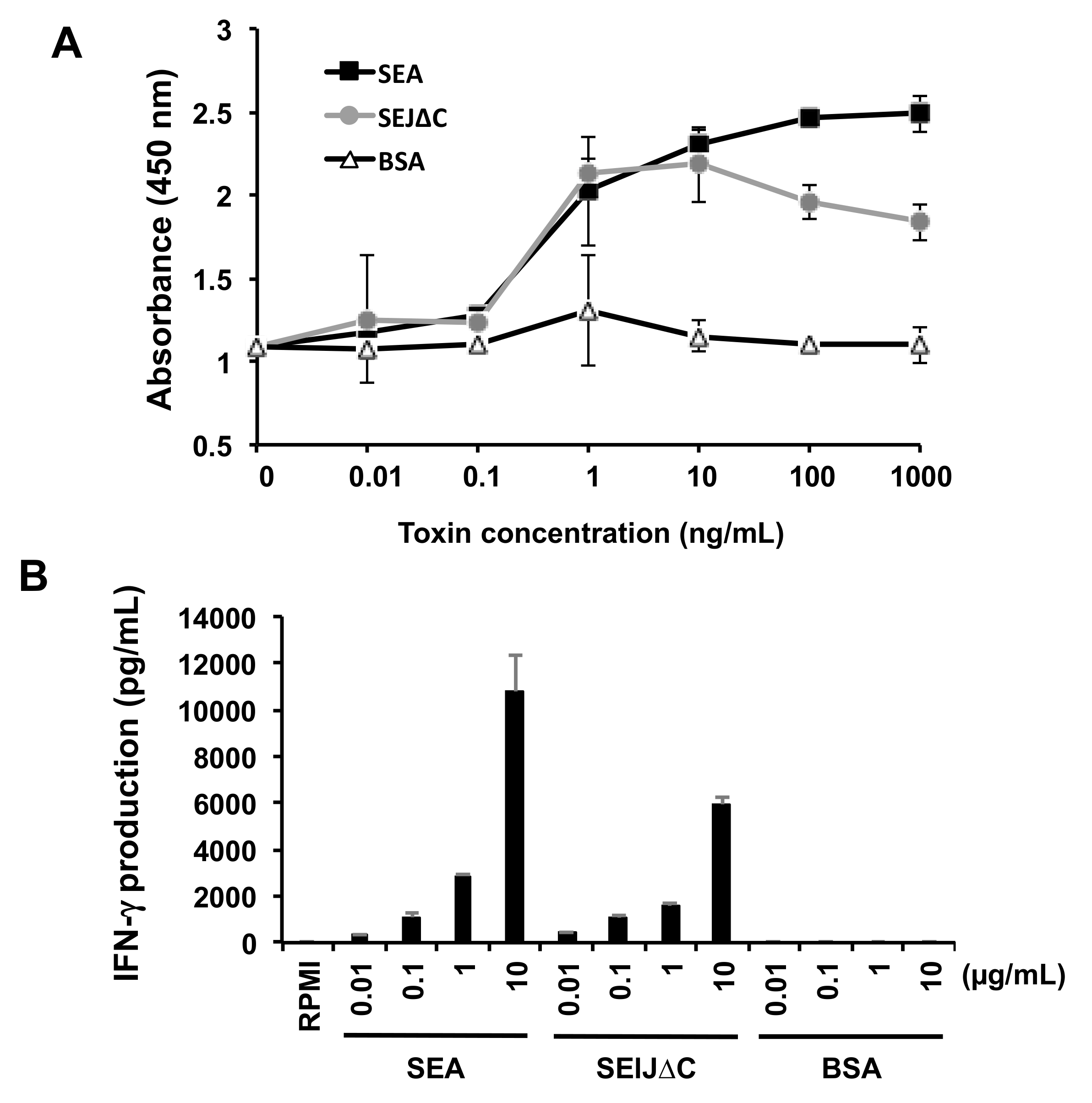
2.2. SElJ∆C Has Superantigenic Activity in Mouse Splenocytes
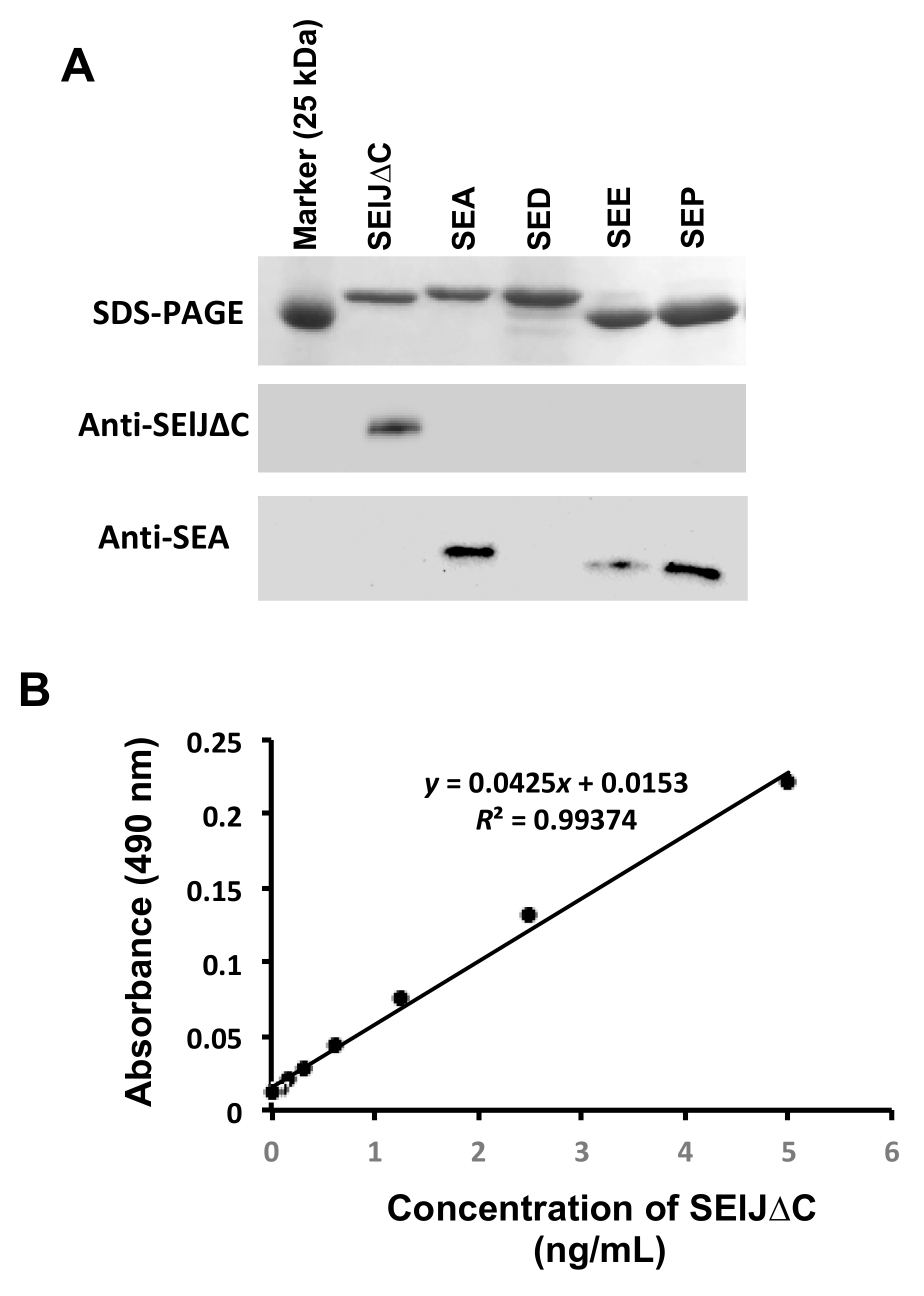
2.3. Development of a Sensitive and Specific Immunoassay for Detection of SElJ
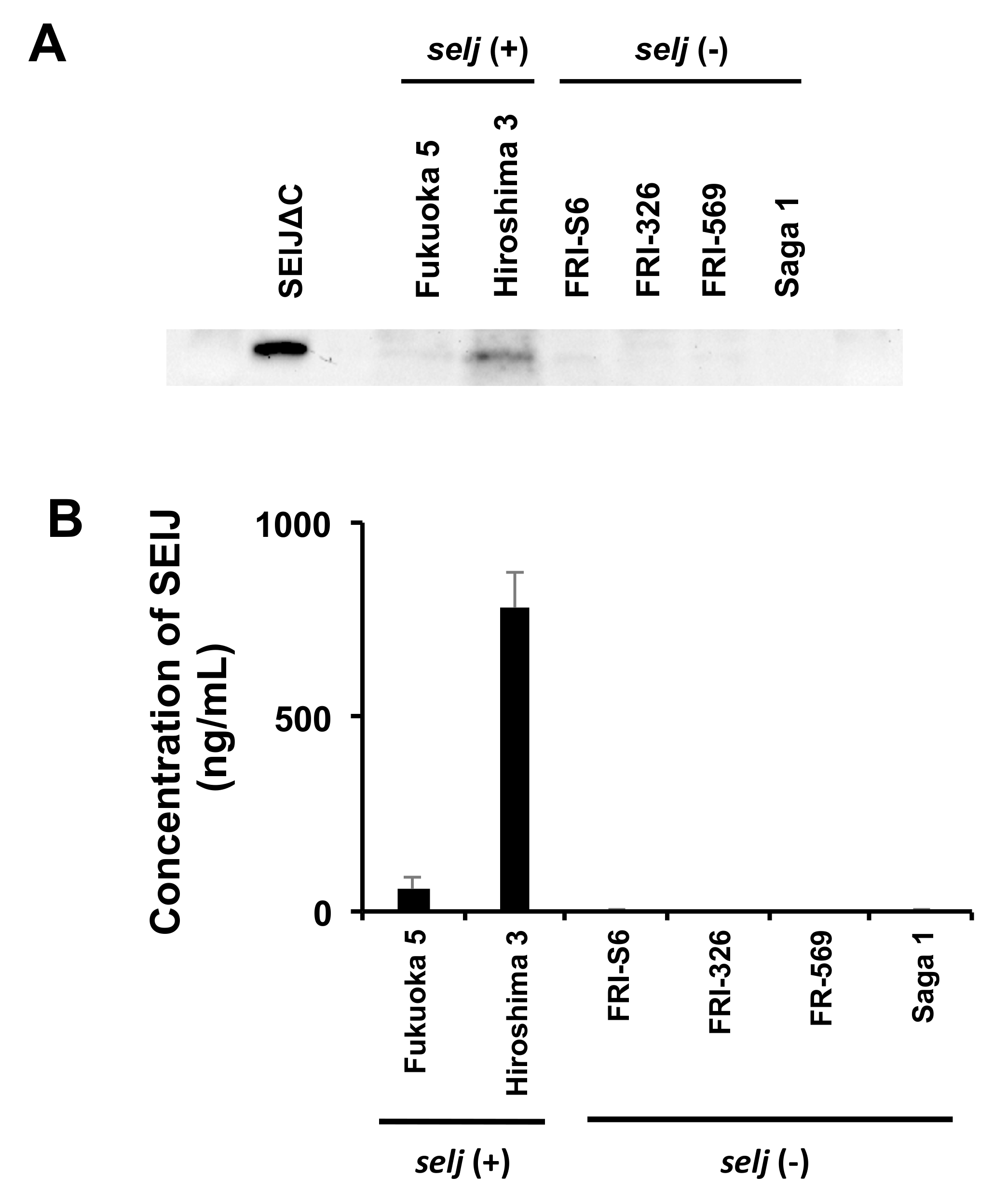
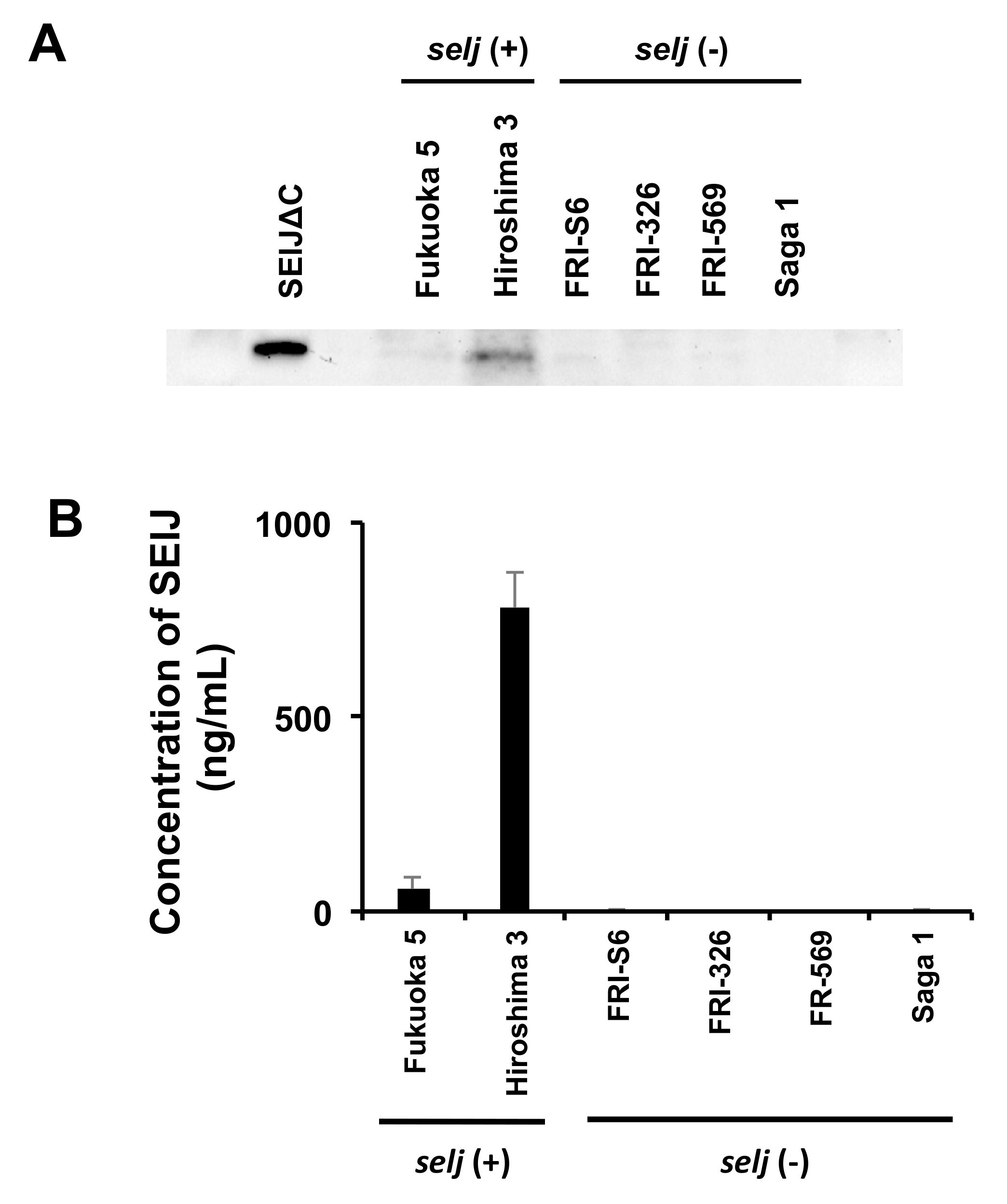
2.4. Production of SElJ in S. aureus Strains Isolated from Food Poisoning
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Cloning and Preparation of SElJ
4.3. Analysis of Superantigenic Activity of SElJ∆C
4.4. Preparation of Anti-SElJ Antibody
4.5. Specificity of Anti-SElJ Antibody by Western Blot Analysis
4.6. Development of Sandwich ELISA
4.7. Detection of SElJ Production in S. aureus Isolates
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McCormick, J.K.; Yarwood, J.M.; Schlievert, P.M. Toxic shock syndrome and bacterial superantigens: An update. Annu. Rev. Microbiol. 2001, 55, 77–104. [Google Scholar] [CrossRef] [PubMed]
- Dinges, M.M.; Orwin, P.M.; Schlievert, P.M. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 2000, 13, 16–34. [Google Scholar] [CrossRef] [PubMed]
- Omoe, K.; Hu, D.L.; Takahashi-Omoe, H.; Nakane, A.; Shinagawa, K. Identification and characterization of a new staphylococcal enterotoxin-related putative toxin encoded by two kinds of plasmids. Infect. Immun. 2003, 71, 6088–6094. [Google Scholar] [CrossRef] [PubMed]
- Ono, H.K.; Omoe, K.; Imanishi, K.; Iwakabe, Y.; Hu, D.L.; Kato, H.; Saito, N.; Nakane, A.; Uchiyama, T.; Shinagawa, K. Identification and characterization of two novel Staphylococcal enterotoxins, types S and T. Infect. Immun. 2008, 76, 4999–5005. [Google Scholar] [CrossRef] [PubMed]
- Ono, H.K.; Sato’o, Y.; Narita, K.; Naito, I.; Hirose, S.; Hisatsune, J.; Asano, K.; Hu, D.L.; Omoe, K.; Sugai, M.; et al. Identification and characterization of a novel staphylococcal emetic toxin. Appl. Environ. Microbiol. 2015, 81, 7034–7040. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.Y.; Jarraud, S.; Lemercier, B.; Cozon, G.; Echasserieau, K.; Etienne, J.; Gougeon, M.L.; Lina, G.; Vandenesch, F. Staphylococcal enterotoxin-like toxins U2 and V, two new staphylococcal superantigens arising from recombination within the enterotoxin gene cluster. Infect. Immun. 2006, 74, 4724–4734. [Google Scholar] [CrossRef] [PubMed]
- Wilson, G.J.; Seo, K.S.; Cartwright, R.A.; Connelley, T.; Chuang-Smith, O.N.; Merriman, J.A.; Guinane, C.M.; Park, J.Y.; Bohach, G.A.; Schlievert, P.M.; et al. A novel core genome-encoded superantigen contributes to lethality of community-associated MRSA necrotizing pneumonia. PLoS Pathog. 2011, 7, e1002271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omoe, K.; Hu, D.L.; Takahashi-Omoe, H.; Nakane, A.; Shinagawa, K. Comprehensive analysis of classical and newly described staphylococcal superantigenic toxin genes in Staphylococcus aureus isolates. FEMS Microbiol. Lett. 2005, 246, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Omoe, K.; Hu, D.L.; Ono, H.K.; Shimizu, S.; Takahashi-Omoe, H.; Nakane, A.; Uchiyama, T.; Shinagawa, K.; Imanishi, K. Emetic potentials of newly identified staphylococcal enterotoxin-like toxins. Infect. Immun. 2013, 81, 3627–3631. [Google Scholar] [CrossRef] [PubMed]
- Ono, H.K.; Hirose, S.; Naito, I.; Sato’o, Y.; Asano, K.; Hu, D.L.; Omoe, K.; Nakane, A. The emetic activity of staphylococcal enterotoxins, SEK, SEL, SEM, SEN and SEO in a small emetic animal model, the house musk shrew. Microbiol. Immunol. 2017, 61, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Iandolo, J.J.; Stewart, G.C. The enterotoxin D plasmid of Staphylococcus aureus encodes a second enterotoxin determinant (sej). FEMS Microbiol. Lett. 1998, 168, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Lv, G.; Xu, B.; Wei, P.; Song, J.; Zhang, H.; Zhao, C.; Qin, L.; Zhao, B. Molecular characterization of foodborne-associated Staphylococcus aureus isolated in Shijiazhuang, China, from 2010 to 2012. Diagn. Microbiol. Infect. Dis. 2014, 78, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Evenson, M.L.; Hinds, M.W.; Bernstein, R.S.; Bergdoll, M.S. Estimation of human dose of staphylococcal enterotoxin A from a large outbreak of staphylococcal food. Int. J. Food Microbiol. 1998, 31, 311–316. [Google Scholar]
- Ikeda, T.; Tamate, N.; Yamaguchi, K.; Makino, S. Mass outbreak of food poisoning disease caused by small amounts of staphylococcal enterotoxins A and H. Appl. Environ. Microbiol. 2005, 71, 2793–2795. [Google Scholar] [CrossRef] [PubMed]
- Bergdoll, M.S. Staphylococcus aureus. In Foodborne Bacterial Pathogens; Doyle, M.P., Ed.; Marcel Dekker Inc.: New York, NY, USA, 1989; pp. 463–523. [Google Scholar]
- Omoe, K.; Ishikawa, M.; Shimoda, Y.; Hu, D.L.; Ueda, S.; Shinagawa, K. Detection of seg, seh, sei genes in Staphylococcus aureus isolates and determination of the enterotoxin productivities of S. aureus isolates Harboring seg, seh, or sei genes. J. Clin. Microbiol. 2002, 40, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.J.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.L.; Omoe, K.; Shimoda, Y.; Nakane, A.; Shinagawa, K. Induction of emetic response to staphylococcal enterotoxins in the house musk shrew (Suncus murinus). Infect. Immun. 2003, 71, 567–570. [Google Scholar] [CrossRef] [PubMed]
- Nakane, A.; Numata, A.; Minagawa, T. Endogenous tumor necrosis factor, interleukin-6, and gamma interferon levels during Listeria monocytogenes infection in mice. Infect. Immun. 1992, 60, 523–528. [Google Scholar] [PubMed]
- Shinagawa, K.; Ishibashi, M.; Yamamoto, H.; Kunita, N.; Hisa, K. A consideration to immune doses of staphylococcal enterotoxin B to rabbits. Jpn. J. Med. Sci. Biol. 1974, 27, 309–314. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. aureus Strains | se Genotype | References |
|---|---|---|
| Fukuoka 5 | selj, ser, ses, set | [3] |
| Hiroshima 3 | seg, sei, sem, sen, seo, selj, ser, ses | this study |
| S6 | sea, seb, selk, seq | [16] |
| FRI-326 | sea, seb, selk, seq | [16] |
| FRI-569 | sea, seb, selk, seq | [16] |
| Saga 1 | seg, sei, sem, sen, seo, sep | [16] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ono, H.K.; Hachiya, N.; Suzuki, Y.; Naito, I.; Hirose, S.; Asano, K.; Omoe, K.; Nakane, A.; Hu, D.-L. Development of an Immunoassay for Detection of Staphylococcal Enterotoxin-Like J, A Non-Characterized Toxin. Toxins 2018, 10, 458. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10110458
Ono HK, Hachiya N, Suzuki Y, Naito I, Hirose S, Asano K, Omoe K, Nakane A, Hu D-L. Development of an Immunoassay for Detection of Staphylococcal Enterotoxin-Like J, A Non-Characterized Toxin. Toxins. 2018; 10(11):458. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10110458
Chicago/Turabian StyleOno, Hisaya K., Nobuaki Hachiya, Yasunori Suzuki, Ikunori Naito, Shouhei Hirose, Krisana Asano, Katsuhiko Omoe, Akio Nakane, and Dong-Liang Hu. 2018. "Development of an Immunoassay for Detection of Staphylococcal Enterotoxin-Like J, A Non-Characterized Toxin" Toxins 10, no. 11: 458. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10110458