A Study and Review of Effects of Botulinum Toxins on Mast Cell Dependent and Independent Pruritus

 and
and
Abstract
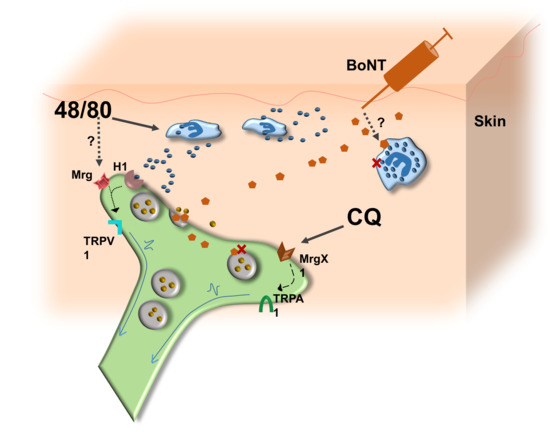
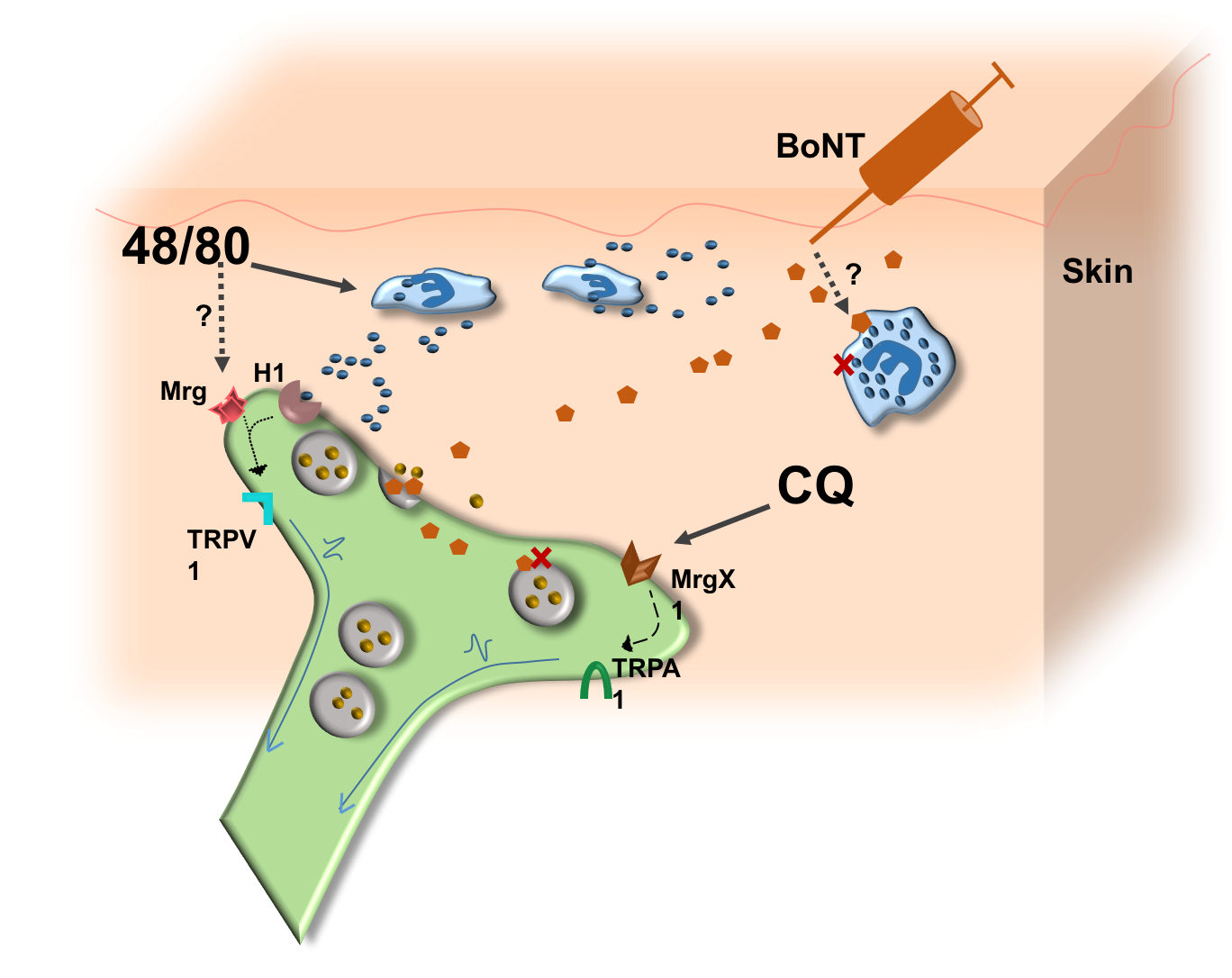
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
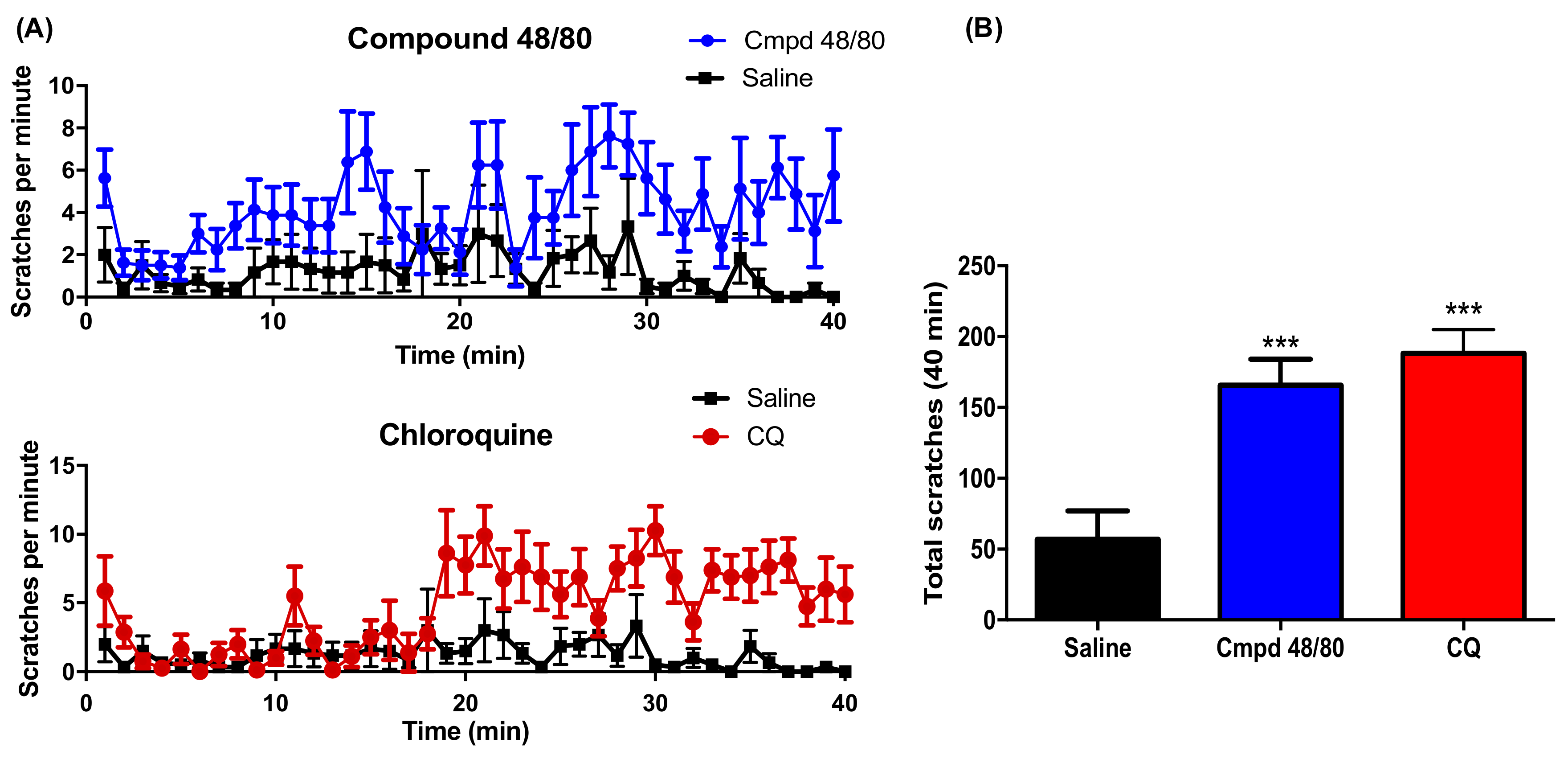
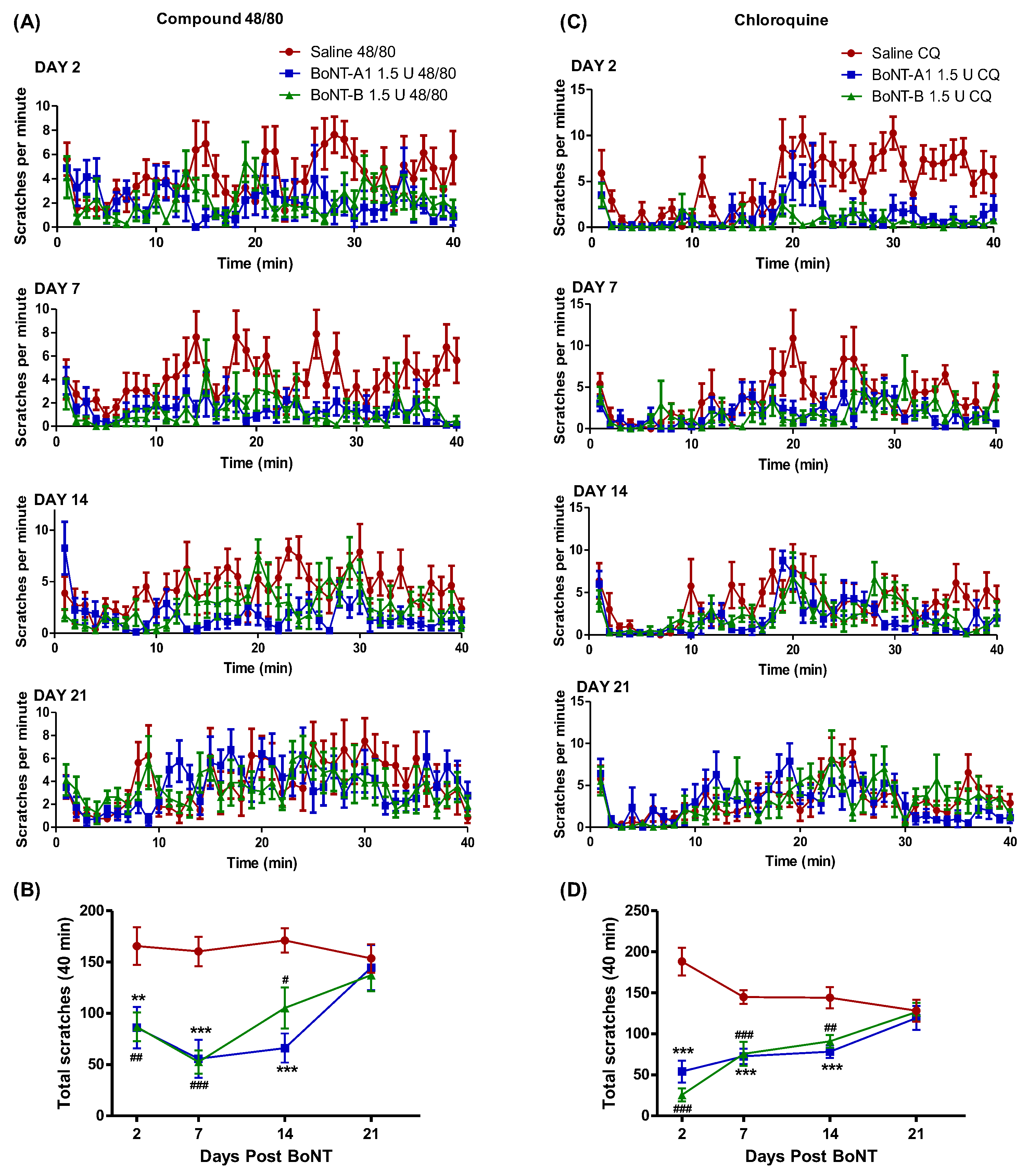
2.1. BoNT/A1 and BoNT/B1 Injection Reduced 48/80 and CQ Induced Scratching
2.2. BoNT/A1 and BoNT/B1 Have a Long Duration of Effect in Reducing Compound 48/80 and CQ Induced Scratching
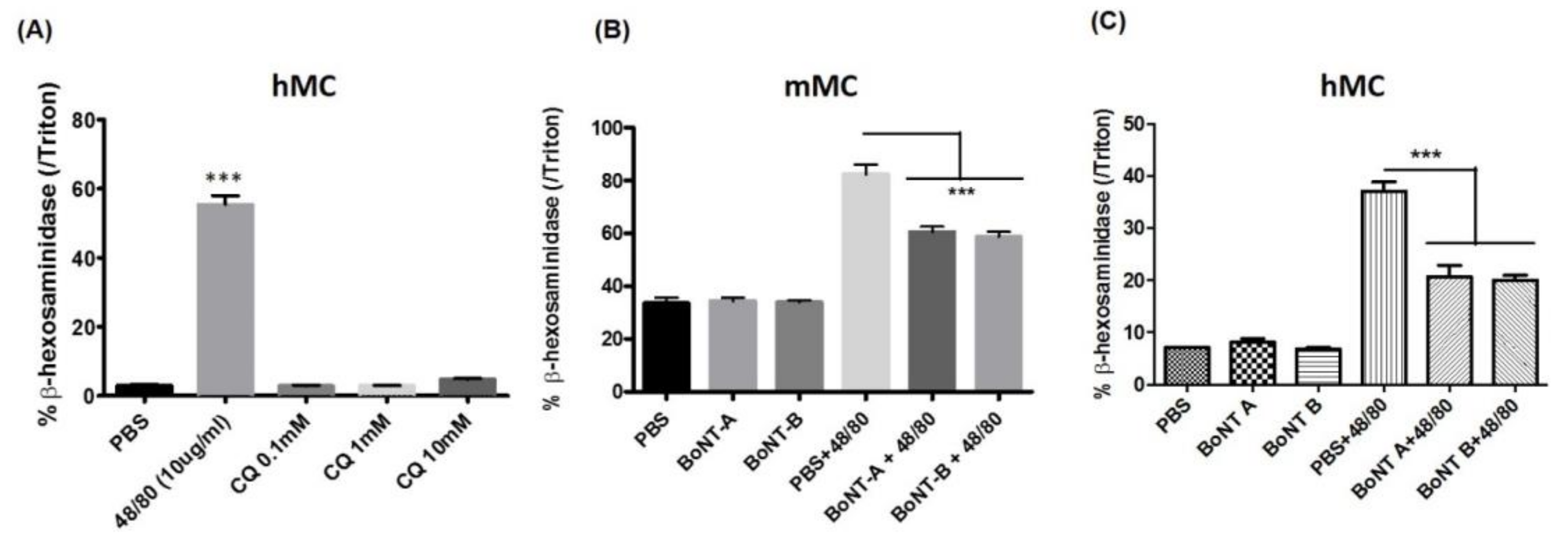
2.3. BoNT/A1 and BoNT/B1 Reduce Compound 48/80 Induced Murine and Human Mast Cell Degranulation
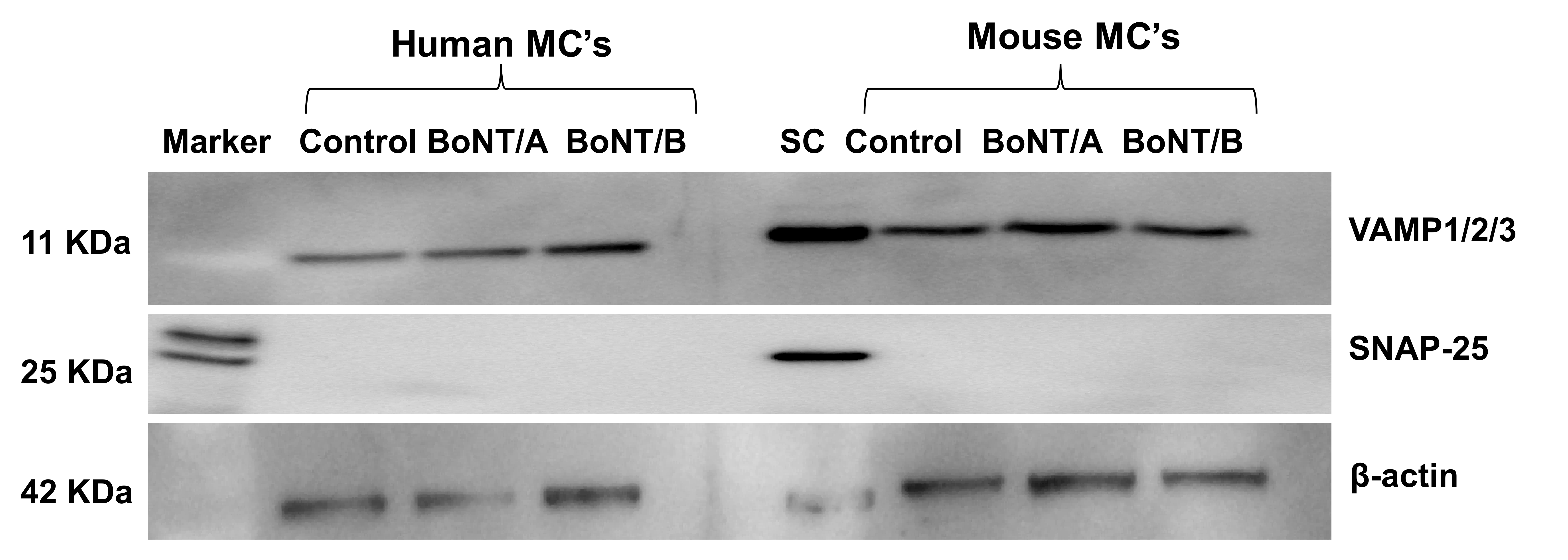
2.4. Western Blot Analysis of Expression and Effect of BoNT/A1 and BoNT/B1 on SNAP-25 and VAMP 1/2/3 in Human and Murine Mast Cells
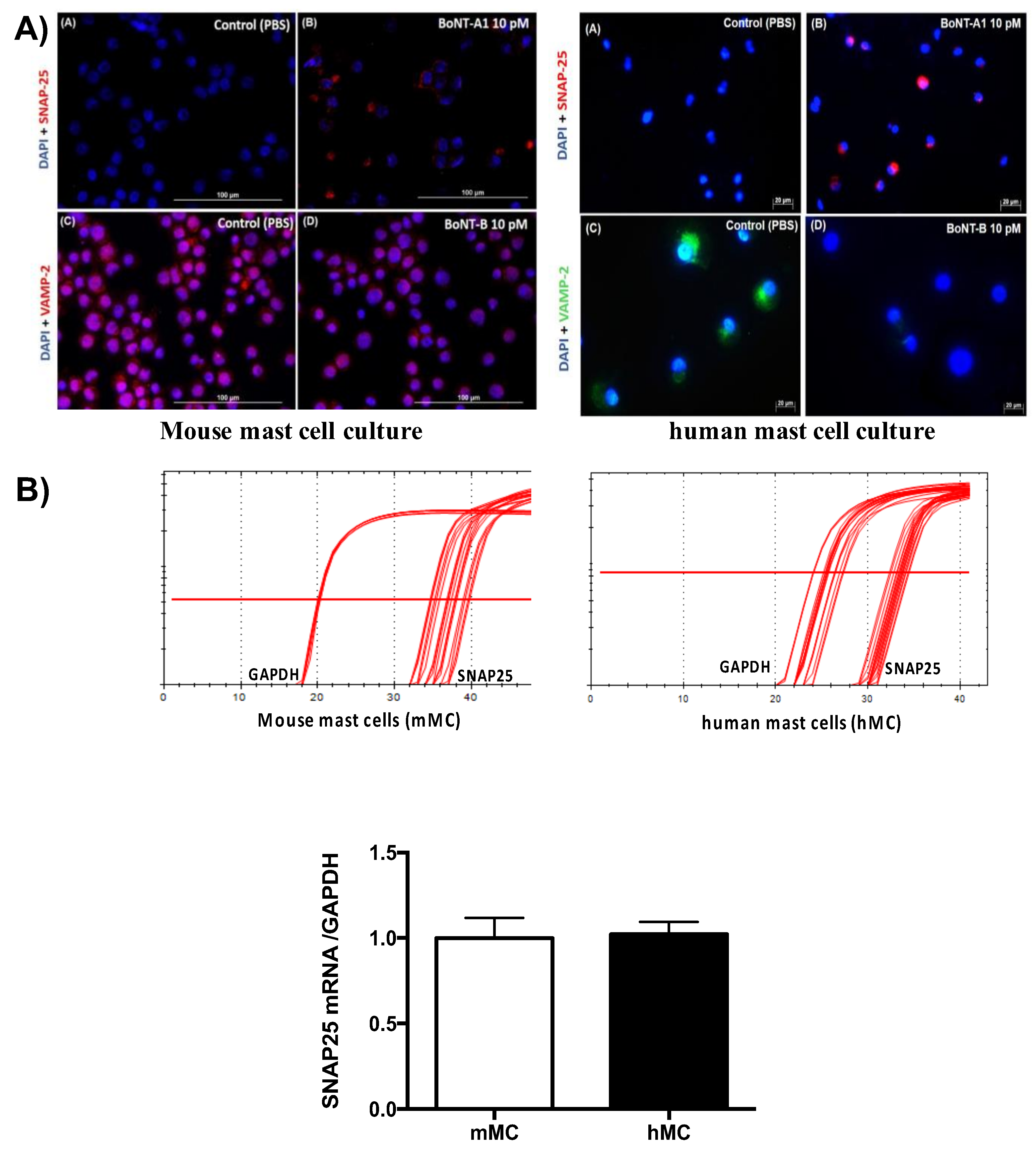
2.5. Expression of SNAP-25 and VAMP 1/2/3 Cleavage with or without BoNT/A1 or B1 Treatment, Respectively, in Human and Murine Mast Cells
3. Discussion
3.1. Mast Cell Dependent and Independent Pruritogens
3.2. Anti-Pruritic Effect of Botulinum Toxin
3.3. Duration of Action of BoNT
3.4. Possible Mechanism of Action of BoNT/A1 and BoNT/B1 in Reducing Induced Itch
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Drug Delivery
4.4. Behavior
4.5. Mast Cell Culture
4.6. Mast Cell Degranulation Assay
4.7. Immunohistochemistry on Mast Cells
4.8. mRNA Isolation and Real-Time Quantitative PCR
4.9. Western Blot Analysis
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Balaskas, E.V.; Chu, M.; Uldall, R.P.; Gupta, A.; Oreopoulos, D.G. Pruritus in continuous ambulatory peritoneal dialysis and hemodialysis patients. Perit. Dial. Int. 1993, 13 (Suppl. 2), S527–S532. [Google Scholar] [PubMed]
- Chia, S.C.; Bergasa, N.V.; Kleiner, D.E.; Goodman, Z.; Hoofnagle, J.H.; Di Bisceglie, A.M. Pruritus as a presenting symptom of chronic hepatitis C. Dig. Dis. Sci. 1998, 43, 2177–2183. [Google Scholar] [CrossRef] [PubMed]
- Yosipovitch, G.; Greaves, M.W.; Schmelz, M. Itch. Lancet 2003, 361, 690–694. [Google Scholar] [CrossRef]
- Oaklander, A.L.; Bowsher, D.; Galer, B.; Haanpää, M.; Jensen, M.P. Herpes zoster itch: Preliminary epidemiologic data. J. Pain 2003, 4, 338–343. [Google Scholar] [CrossRef]
- Ikoma, A.; Steinhoff, M.; Ständer, S.; Yosipovitch, G.; Schmelz, M. The neurobiology of itch. Nat. Rev. Neurosci. 2006, 7, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Schmelz, M.; Schmidt, R.; Weidner, C.; Hilliges, M.; Torebjörk, H.E.; Handwerker, H.O. Chemical response pattern of different classes of C-nociceptors to pruritogens and algogens. J. Neurophysiol. 2003, 89, 2441–2448. [Google Scholar] [CrossRef] [PubMed]
- Schmelz, M.; Schmidt, R.; Bickel, A.; Handwerker, H.O.; Torebjörk, H.E. Specific C-receptors for itch in human skin. J. Neurosci. 1997, 17, 8003–8008. [Google Scholar] [PubMed]
- Shim, W.-S.; Tak, M.-H.; Lee, M.-H.; Kim, M.; Kim, M.; Koo, J.-Y.; Lee, C.-H.; Kim, M.; Oh, U. TRPV1 mediates histamine-induced itching via the activation of phospholipase A2 and 12-lipoxygenase. J. Neurosci. 2007, 27, 2331–2337. [Google Scholar] [CrossRef] [PubMed]
- Imamachi, N.; Park, G.H.; Lee, H.; Anderson, D.J.; Simon, M.I.; Basbaum, A.I.; Han, S.-K. TRPV1-expressing primary afferents generate behavioral responses to pruritogens via multiple mechanisms. Proc. Natl. Acad. Sci. USA 2009, 106, 11330–11335. [Google Scholar] [CrossRef] [PubMed]
- McNaughton, F.L.; Obianwu, H.O.; Isah, A.O.; Arhewoh, I.M. Chloroquine-induced Pruritus. Indian J. Pharm. Sci. 2010, 72, 283–289. [Google Scholar]
- Liu, Q.; Tang, Z.; Surdenikova, L.; Kim, S.; Patel, K.N.; Kim, A.; Ru, F.; Guan, Y.; Weng, H.-J.; Geng, Y.; et al. Sensory neuron-specific GPCR Mrgprs are itch receptors mediating chloroquine-induced pruritus. Cell 2009, 139, 1353–1365. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.R.; Gerhold, K.A.; Bifolck-Fisher, A.; Liu, Q.; Patel, K.N.; Dong, X.; Bautista, D.M. TRPA1 is required for histamine-independent, Mas-related G protein-coupled receptor-mediated itch. Nat. Neurosci. 2011, 14, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Schiavo, G.; Rossetto, O.; Santucci, A.; DasGupta, B.R.; Montecucco, C. Botulinum neurotoxins are zinc proteins. J. Biol. Chem. 1992, 267, 23479–23483. [Google Scholar] [PubMed]
- Montecucco, C.; Schiavo, G. Mechanism of action of tetanus and botulinum neurotoxins. Mol. Microbiol. 1994, 13, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Schiavo, G.; Matteoli, M.; Montecucco, C. Neurotoxins affecting neuroexocytosis. Physiol. Rev. 2000, 80, 717–766. [Google Scholar] [CrossRef] [PubMed]
- Bach-Rojecky, L.; Lacković, Z. Central origin of the antinociceptive action of botulinum toxin type A. Pharmacol. Biochem. Behav. 2009, 94, 234–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marino, M.J.; Terashima, T.; Steinauer, J.J.; Eddinger, K.A.; Yaksh, T.L.; Xu, Q. Botulinum toxin B in the sensory afferent: Transmitter release, spinal activation, and pain behavior. Pain 2014, 155, 674–684. [Google Scholar] [CrossRef] [PubMed]
- Filipović, B.; Matak, I.; Bach-Rojecky, L.; Lacković, Z. Central action of peripherally applied botulinum toxin type A on pain and dural protein extravasation in rat model of trigeminal neuropathy. PLoS ONE 2012, 7, e29803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazerani, P.; Pedersen, N.S.; Drewes, A.M.; Arendt-Nielsen, L. Botulinum toxin type A reduces histamine-induced itch and vasomotor responses in human skin. Br. J. Dermatol. 2009, 161, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Krämer, H.H.; Angerer, C.; Erbguth, F.; Schmelz, M.; Birklein, F. Botulinum Toxin A reduces neurogenic flare but has almost no effect on pain and hyperalgesia in human skin. J. Neurol. 2003, 250, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Tugnoli, V.; Capone, J.G.; Eleopra, R.; Quatrale, R.; Sensi, M.; Gastaldo, E.; Tola, M.R.; Geppetti, P. Botulinum toxin type A reduces capsaicin-evoked pain and neurogenic vasodilatation in human skin. Pain 2007, 130, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, N.M.E.; Dostrovsky, J.O.; Charlton, M.P. Peptide-mediated transdermal delivery of botulinum neurotoxin type A reduces neurogenic inflammation in the skin. Pain 2010, 149, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Wollina, U.; Karamfilov, T. Adjuvant botulinum toxin A in dyshidrotic hand eczema: A controlled prospective pilot study with left-right comparison. J. Eur. Acad. Dermatol. Venereol. 2002, 16, 40–42. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Brooks, P. The use of botulinum toxin in the management of burns itching: Preliminary results. Burns 2012, 38, 1119–1123. [Google Scholar] [CrossRef] [PubMed]
- Heckmann, M.; Heyer, G.; Brunner, B.; Plewig, G. Botulinum toxin type A injection in the treatment of lichen simplex: An open pilot study. J. Am. Acad. Dermatol. 2002, 46, 617–619. [Google Scholar] [CrossRef] [PubMed]
- Pellett, S.; Tepp, W.H.; Whitemarsh, R.C.M.; Bradshaw, M.; Johnson, E.A. In vivo onset and duration of action varies for botulinum neurotoxin A subtypes 1–5. Toxicon 2015, 107, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Moon, T.C.; Befus, A.D.; Kulka, M. Mast cell mediators: Their differential release and the secretory pathways involved. Front. Immunol. 2014, 5, 569. [Google Scholar] [CrossRef] [PubMed]
- Lorentz, A.; Baumann, A.; Vitte, J.; Blank, U. The SNARE Machinery in Mast Cell Secretion. Front. Immunol. 2012, 3, 143. [Google Scholar] [CrossRef] [PubMed]
- Potenzieri, C.; Undem, B.J. Basic mechanisms of itch. Clin. Exp. Allergy 2012, 42, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Kuraishi, Y.; Nagasawa, T.; Hayashi, K.; Satoh, M. Scratching behavior induced by pruritogenic but not algesiogenic agents in mice. Eur. J. Pharmacol. 1995, 275, 229–233. [Google Scholar] [CrossRef]
- Wallengren, J. Neuroanatomy and neurophysiology of itch. Dermatol. Ther. 2005, 18, 292–303. [Google Scholar] [CrossRef] [PubMed]
- Schemann, M.; Kugler, E.M.; Buhner, S.; Eastwood, C.; Donovan, J.; Jiang, W.; Grundy, D. The mast cell degranulator compound 48/80 directly activates neurons. PLoS ONE 2012, 7, e52104. [Google Scholar] [CrossRef] [PubMed]
- De Jonge, F.; De Laet, A.; Van Nassauw, L.; Brown, J.K.; Miller, H.R.P.; van Bogaert, P.P.; Timmermans, J.P.; Kroese, A.B.A. In vitro activation of murine DRG neurons by CGRP-mediated mucosal mast cell degranulation. Am. J. Physiol.-Gastrointest. Liver Physiol. 2004, 287, G178–G191. [Google Scholar] [CrossRef] [PubMed]
- Durham, P.L.; Cady, R.; Cady, R. Regulation of calcitonin gene-related peptide secretion from trigeminal nerve cells by botulinum toxin type A: Implications for migraine therapy. Headache 2004, 44, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Ovsepian, S.V.; Wang, J.; Pickering, M.; Sasse, A.; Aoki, K.R.; Lawrence, G.W.; Dolly, J.O. Activation of TRPV1 mediates calcitonin gene-related peptide release, which excites trigeminal sensory neurons and is attenuated by a retargeted botulinum toxin with anti-nociceptive potential. J. Neurosci. 2009, 29, 4981–4992. [Google Scholar] [CrossRef] [PubMed]
- Heyer, G.; Vogelgsang, M.; Hornstein, O.P. Acetylcholine is an inducer of itching in patients with atopic eczema. J. Dermatol. 1997, 24, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Buddenkotte, J.; Berger, T.G.; Steinhoff, M. Management of itch in atopic dermatitis. Semin. Cutan. Med. Surg. 2011, 30, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Koga, K.; Chen, T.; Li, X.-Y.; Descalzi, G.; Ling, J.; Gu, J.; Zhuo, M. Glutamate acts as a neurotransmitter for gastrin releasing peptide-sensitive and insensitive itch-related synaptic transmission in mammalian spinal cord. Mol. Pain 2011, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.; Nabarro, G.; Baart de la Faille, H.; van Vloten, W.A.; van der Putte, S.C.; Schuurman, H.J. Increased number of immunoreactive nerve fibers in atopic dermatitis. J. Allergy Clin. Immunol. 1992, 90, 613–622. [Google Scholar] [CrossRef]
- Dolly, J.O.; Lawrence, G.W.; Meng, J.; Wang, J.; Ovsepian, S.V. Neuro-exocytosis: Botulinum toxins as inhibitory probes and versatile therapeutics. Curr. Opin. Pharmacol. 2009, 9, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, R.; Lam, C.; Yaksh, T.L. Botulinum toxin in migraine: Role of transport in trigemino-somatic and trigemino-vascular afferents. Neurobiol. Dis. 2015, 79, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Khanijou, S.; Rubino, J.; Aoki, K.R. Subcutaneous administration of botulinum toxin A reduces formalin-induced pain. Pain 2004, 107, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.P.; Khan, I.; Suhail, M.S.A.; Malkmus, S.; Yaksh, T.L. Spinal botulinum neurotoxin B: Effects on afferent transmitter release and nociceptive processing. PLoS ONE 2011, 6, e19126. [Google Scholar]
- Keller, J.E.; Neale, E.A. The role of the synaptic protein SNAP-25 in the potency of botulinum neurotoxin type A. J. Biol. Chem. 2001, 276, 13476–13482. [Google Scholar] [CrossRef] [PubMed]
- Whitemarsh, R.C.M.; Tepp, W.H.; Bradshaw, M.; Lin, G.; Pier, C.L.; Scherf, J.M.; Johnson, E.A.; Pellett, S. Characterization of botulinum neurotoxin A subtypes 1 through 5 by investigation of activities in mice, in neuronal cell cultures, and in vitro. Infect. Immun. 2013, 81, 3894–3902. [Google Scholar] [CrossRef] [PubMed]
- Keller, J.E. Recovery from botulinum neurotoxin poisoning in vivo. Neuroscience 2006, 139, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Park, T.H. The effects of botulinum toxin A on mast cell activity: Preliminary results. Burns 2013, 39, 816–817. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Turner, C.; Castle, D. Relocation of the t-SNARE SNAP-23 from lamellipodia-like cell surface projections regulates compound exocytosis in mast cells. Cell 1998, 94, 537–548. [Google Scholar] [CrossRef]
- Vaidyanathan, V.V.; Puri, N.; Roche, P.A. The last exon of SNAP-23 regulates granule exocytosis from mast cells. J. Biol. Chem. 2001, 276, 25101–25106. [Google Scholar] [CrossRef] [PubMed]
- Woska, J.R.; Gillespie, M.E. SNARE complex-mediated degranulation in mast cells. J. Cell. Mol. Med. 2012, 16, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Shibata, M.; Toriumi, H.; Iwashita, T.; Funakubo, M.; Sato, H.; Kuroi, T.; Ebine, T.; Koizumi, K.; Suzuki, N. Reduction of TRPV1 expression in the trigeminal system by botulinum neurotoxin type-A. Neurobiol. Dis. 2012, 48, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Apostolidis, A.; Popat, R.; Yiangou, Y. Decreased sensory receptors P2X3 and TRPV1 in suburothelial nerve fibers following intradetrusor injections of Botulinum toxin for human detrusor overactivity. J. Urol. 2005, 174, 977–983. [Google Scholar] [CrossRef] [PubMed]
- Cochrane, D.E.; Douglas, W.W. Calcium-induced extrusion of secretory granules (exocytosis) in mast cells exposed to 48-80 or the ionophores A-23187 and X-537A. Proc. Natl. Acad. Sci. USA 1974, 71, 408–412. [Google Scholar] [CrossRef] [PubMed]
- Hirokawa, N.; Heuser, J.E. Structural evidence that botulinum toxin blocks neuromuscular transmission by impairing the calcium influx that normally accompanies nerve depolarization. J. Cell Biol. 1981, 88, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Yaksh, T.L.; Ozaki, G.; McCumber, D.; Rathbun, M.; Svensson, C.; Malkmus, S.; Yaksh, M.C. An automated flinch detecting system for use in the formalin nociceptive bioassay. J. Appl. Physiol. 2001, 90, 2386–2402. [Google Scholar] [CrossRef] [PubMed]
- Marino, M.; Huang, P.; Malkmus, S.; Robertshaw, E.; Mac, E.A.; Shatterman, Y.; Yaksh, T.L. Development and validation of an automated system for detection and assessment of scratching in the rodent. J. Neurosci. Methods 2012, 211, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Lai, Y.; Bernard, J.J.; Macleod, D.T.; Cogen, A.L.; Moss, B.; Di Nardo, A. Skin mast cells protect mice against vaccinia virus by triggering mast cell receptor S1PR2 and releasing antimicrobial peptides. J. Immunol. 2012, 188, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Kirshenbaum, A.S.; Metcalfe, D.D. Growth of human mast cells from bone marrow and peripheral blood-derived CD34+ pluripotent progenitor cells. Methods Mol. Biol. 2006, 315, 105–112. [Google Scholar] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramachandran, R.; Marino, M.J.; Paul, S.; Wang, Z.; Mascarenhas, N.L.; Pellett, S.; Johnson, E.A.; DiNardo, A.; Yaksh, T.L. A Study and Review of Effects of Botulinum Toxins on Mast Cell Dependent and Independent Pruritus. Toxins 2018, 10, 134. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10040134
Ramachandran R, Marino MJ, Paul S, Wang Z, Mascarenhas NL, Pellett S, Johnson EA, DiNardo A, Yaksh TL. A Study and Review of Effects of Botulinum Toxins on Mast Cell Dependent and Independent Pruritus. Toxins. 2018; 10(4):134. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10040134
Chicago/Turabian StyleRamachandran, Roshni, Marc J. Marino, Snighdha Paul, Zhenping Wang, Nicholas L. Mascarenhas, Sabine Pellett, Eric A. Johnson, Anna DiNardo, and Tony L. Yaksh. 2018. "A Study and Review of Effects of Botulinum Toxins on Mast Cell Dependent and Independent Pruritus" Toxins 10, no. 4: 134. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10040134