Interannual Variability of Dinophysis acuminata and Protoceratium reticulatum in a Chilean Fjord: Insights from the Realized Niche Analysis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
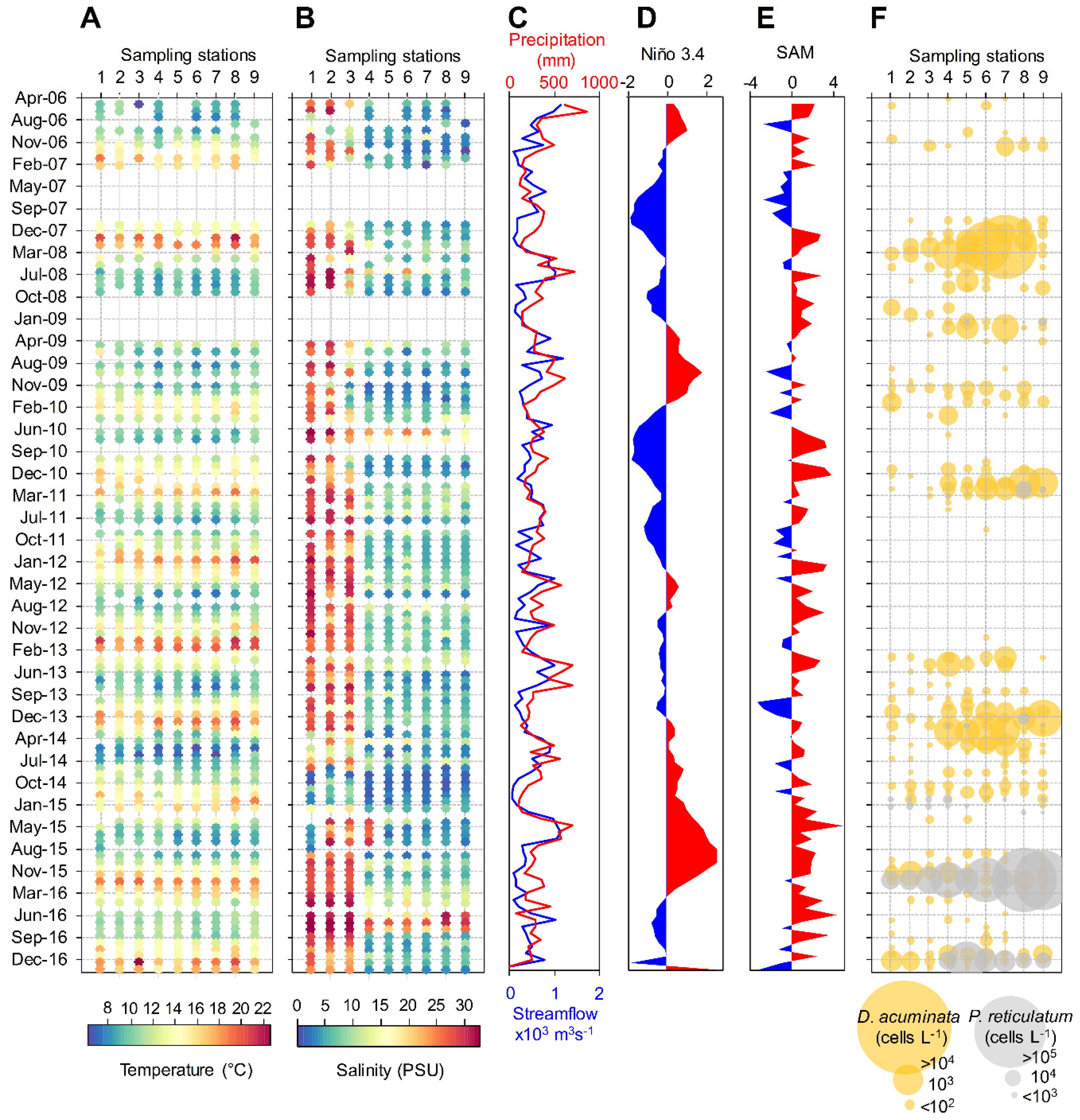
2.1. Physical and Meteorological Conditions
2.2. Spatio-Temporal Distribution of Dinophysis spp. and P. reticulatum
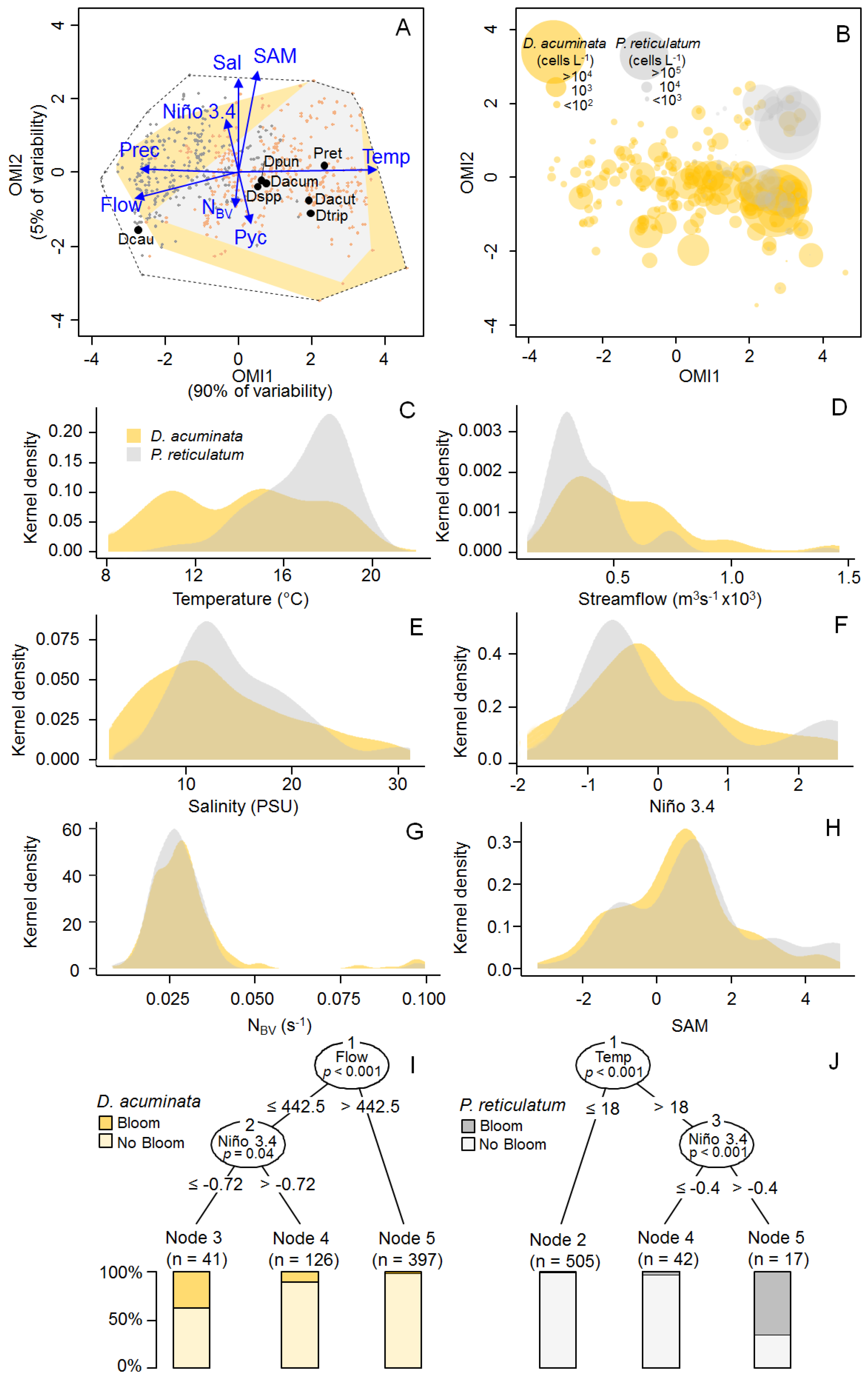
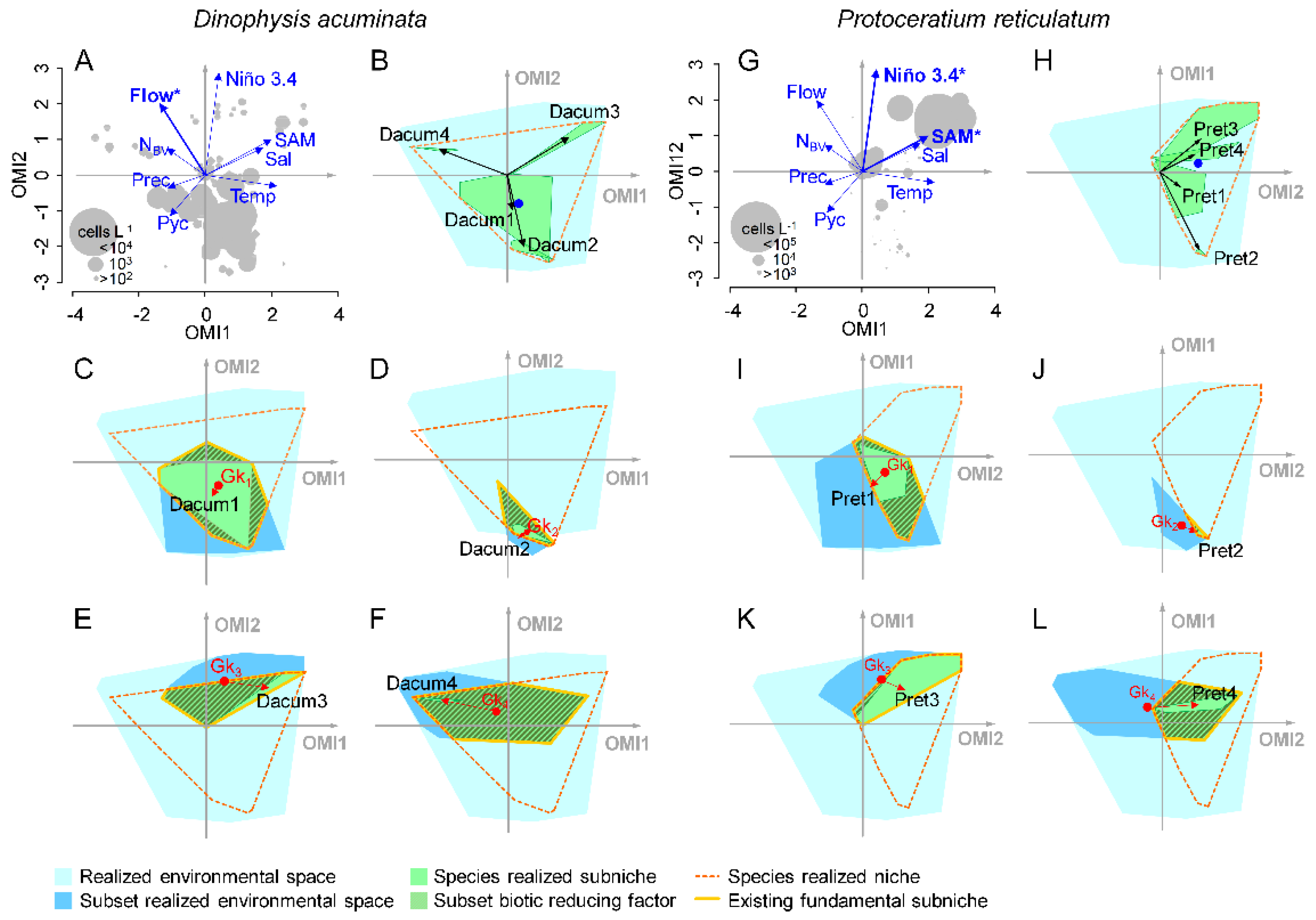
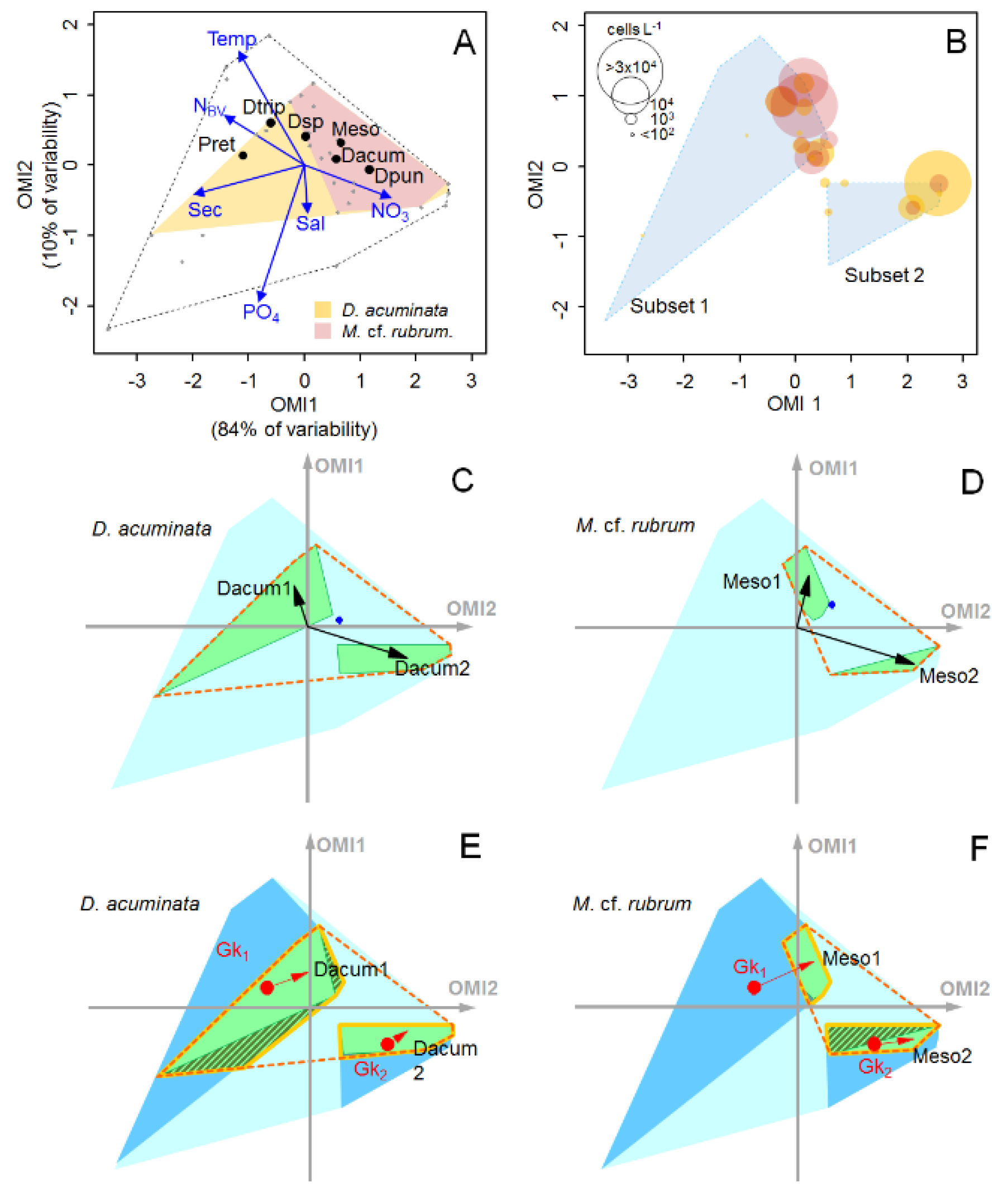
2.3. Niche Analysis
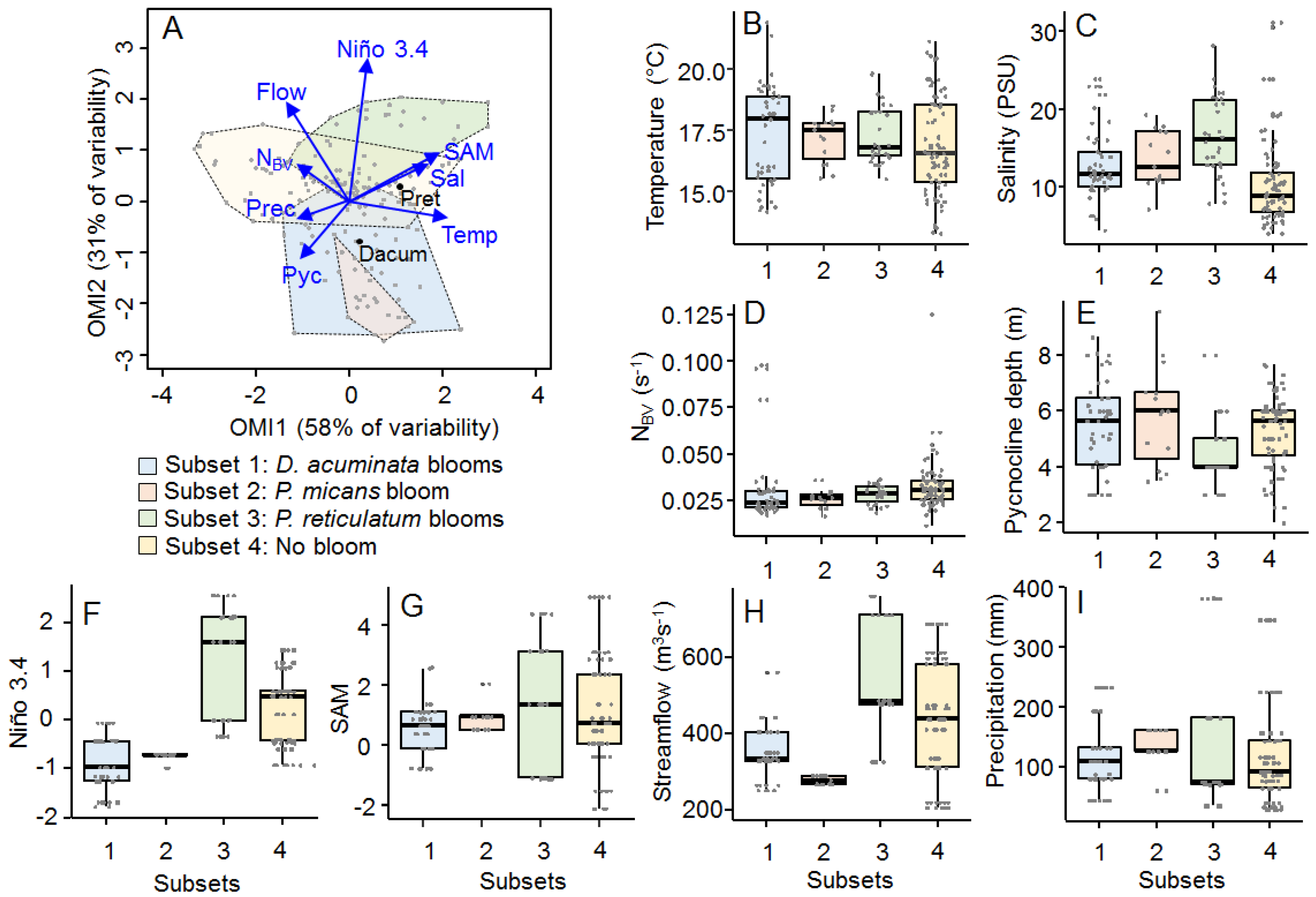
2.4. Subniche Analysis
2.4.1. Subsets
2.4.2. Subniches
3. Discussion
3.1. Seasonal and Interannual Variability
3.2. Summer Subsets
3.3. Subniches and Reducing Biotic Factors
3.4. Additional Aspects Affecting D. acuminata
3.5. Concluding Remarks
4. Materials and Methods
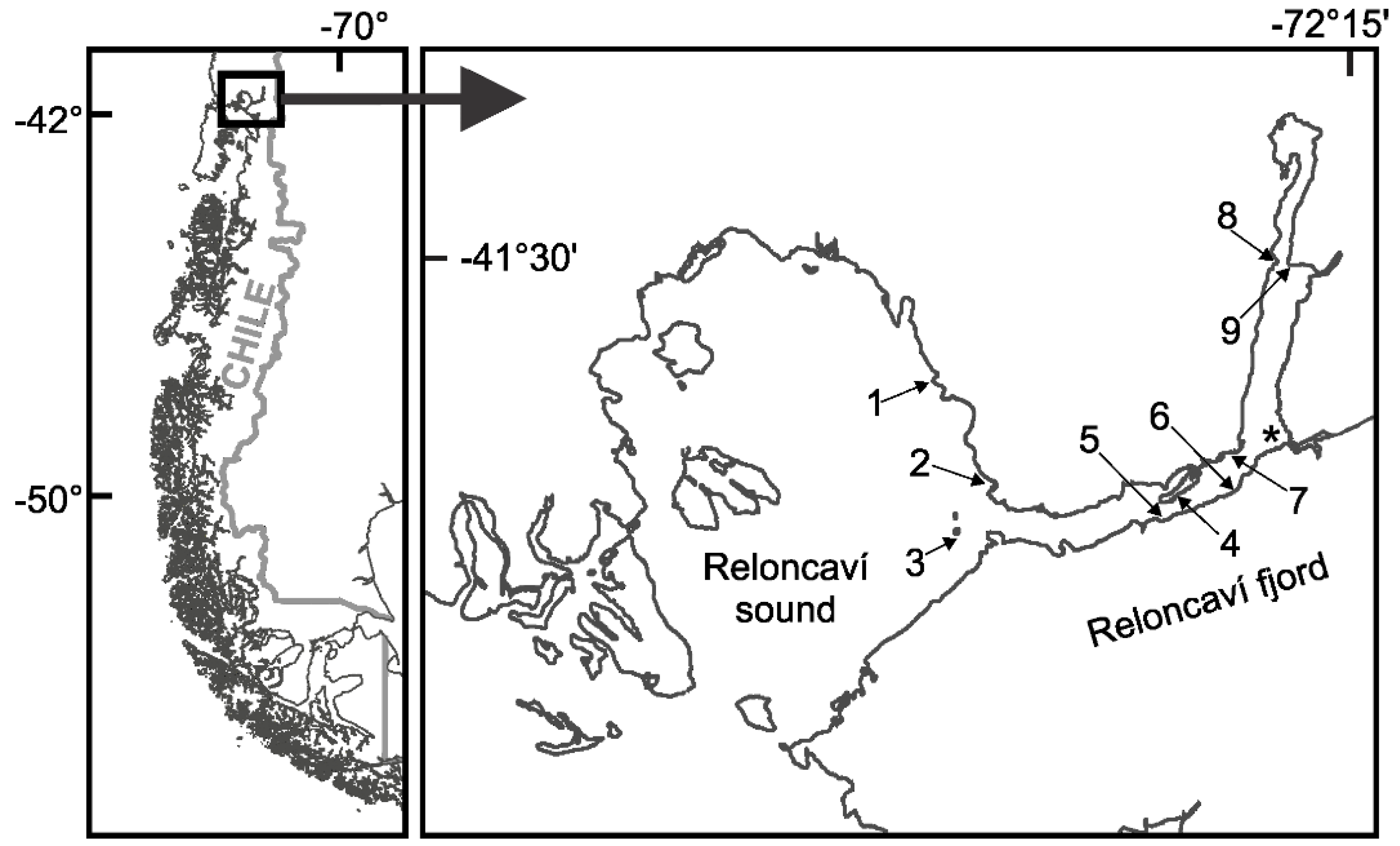
4.1. Study Area and Datasets
4.2. Statistical Analysis
4.2.1. Niche Analysis
4.2.2. Subniche analysis
4.2.3. Relevance of Environmental Variables
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- van Dolah, F. Marine algal toxins: Origins, health effects, and their increased occurrence. Environ. Health Perspect. 2000, 108, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Reguera, B.; Riobó, P.; Rodríguez, F.; Díaz, P.A.; Pizarro, G.; Paz, B.; Franco, J.M.; Blanco, J. Dinophysis toxins: Causative organisms, distribution and fate in shellfish. Mar. Drugs 2014, 12, 394–461. [Google Scholar] [CrossRef] [PubMed]
- Hamano, Y.; Kinoshita, Y.; Yasumoto, T. Enteropathogenicity of diarrhetic shellfish toxins in intestinal models. J. Food Hyg. Soc. Jpn. 1986, 27, 375–379. [Google Scholar] [CrossRef]
- Blanco, J.; Moroño, A.; Fernández, M.L. Toxic episodes in shellfish, produced by lipophilic phycotoxins: An overview. Rev. Gal. Rec. Mar. (Monog.) 2005, 1, 1–70. [Google Scholar]
- Tubaro, A.; Dell’Ovo, V.; Sosa, S.; Florio, C. Yessotoxins: A toxicological overview. Toxicon 2010, 56, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, G.; Uribe, E.; Regueiro, J.; Blanco, J.; Fraga, S. Gonyaulax taylorii, a new yessotoxins-producer dinoflagellate species from Chilean waters. Harmful Algae 2016, 58, 8–15. [Google Scholar] [CrossRef]
- Tillmann, U.; Elbrächter, M.; Krock, B.; John, U.; Cembella, A. Azadinium spinosum gen. et sp. nov. (Dinophyceae) identified as a primary producer of azaspiracid toxins. Eur. J. Phycol. 2009, 44, 63–79. [Google Scholar] [CrossRef]
- Muñoz, F.; Avaria, S.; Sieveiï, H.; Prado, R. Presencia de dinoflagelados toxicos del genero Dinophysis en el seno Aysén, Chile. Rev. Biol. Mar. 1992, 27, 187–212. [Google Scholar]
- Uribe, J.C.; García, C.; Rivas, M.; Lagos, N. First report of diarrhetic shellfish toxins in Magellanic Fjords, southern Chile. J. Shellfish Res. 2001, 20, 69–74. [Google Scholar]
- Cassis, D.; Muñoz, P.; Avaria, S. Variación temporal del fitoplancton entre 1993 y 1998 en una estación fija del seno Aysén, Chile (45°26′S 73°00′W). Rev. Biol. Mar. Oceanogr. 2002, 37, 43–65. [Google Scholar] [CrossRef]
- Seguel, M.; Tocornal, M.A.; Sfeir, A. Floraciones algales nocivas en los canales y fiordos del sur de Chile. Cienc. Tecnol. Mar. 2005, 28, 5–13. [Google Scholar]
- Pizarro, G.; Paz, B.; Alarcón, C.; Toro, C.; Frangópulos, M.; Salgado, P.; Olave, C.; Zamora, C.; Pacheco, H.; Guzmán, L. Winter distribution of toxic, potentially toxic phytoplankton, and shellfish toxins in fjords and channels of the Aysén region, Chile. Lat. Am. J. Aquat. Res. 2018, 46, 120–139. [Google Scholar] [CrossRef]
- Moreno-Pino, M.; Krock, B.; De la Iglesia, R.; Echenique-Subiabre, I.; Pizarro, G.; Vásquez, M.; Trefault, N. Next Generation Sequencing and mass spectrometry reveal high taxonomic diversity and complex phytoplankton-phycotoxins patterns in Southeastern Pacific fjords. Toxicon 2018. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, L.; Campodónico, E. Mareas rojas en Chile. Interciencia 1978, 3, 144–149. [Google Scholar]
- Zhao, J.; Lembeye, G.; Cenci, G.; Wall, B.; Yasumoto, T. Determination of okadaic acid and dinophysistoxin-1 in mussels from Chile, Italy and Ireland. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 587–592. [Google Scholar]
- García, C.; González, V.; Cornejo, C.; Palma-Fleming, H.; Lagos, N. First evidence of dinophisistoxin-1 and carcinogenic polycyclic aromatic hydrocarbons in smoked bivalves collected in the patagonic fjords. Toxicon 2004, 43, 121–131. [Google Scholar] [CrossRef]
- García, C.; Pruzzo, N.; Rodríguez-Unda, N.; Contreras, C.; Lagos, N. First evidence of Okadaic acid acyl-derivative and dinophysistoxin-3 in mussel samples collected in Chiloe Island, southern Chile. J. Toxicol. Sci. 2010, 35, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Lembeye, G.; Yasumoto, T.; Zhao, J.; Fernández, R. DSP outbreak in Chilean Fjords. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 525–529. [Google Scholar]
- Garcia, C.; Rodriguez-Unda, N.; Contreras, C.; Barriga, A.; Lagos, N. Lipophilic toxin profiles detected in farmed and benthic mussels populations from the most relevant production zones in southern Chile. Food Addit. Contam. A 2012, 29, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Trefault, N.; Krock, B.; Delherbe, N.; Cembella, A.; Vásquez, M. Latitudinal transects in the southeastern Pacific Ocean reveal a diverse but patchy distribution of phycotoxins. Toxicon 2011, 58, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Alves-de-Souza, C.; Varela, D.; Contreras, C.; de La Iglesia, P.; Fernández, P.; Hipp, B.; Hernández, C.; Riobó, P.; Reguera, B.; Franco, J.M. Seasonal variability of Dinophysis spp. and Protoceratium reticulatum associated to lipophilic shellfish toxins in a strongly stratified Chilean fjord. Deep Sea Res. II Top. Stud. Oceanogr. 2014, 101, 152–162. [Google Scholar] [CrossRef]
- Goto, H.; Igarashi, T.; Watai, M.; Yasumoto, T.; Gomez, O.V.; Valdivia, G.L.; Noren, E.; Gisselson, L.A.; Graneli, E. Worldwide occurrence of pectenotoxins and yessotoxins in shellfish and phytoplankton. In Harmful Algal Blooms 2000, Proceedings of the IX International Conference on Harmful Alga Blooms, Hobart, Australia, 7–11 February 2000; Hallegraeff, G., Blackburn, S.I., Bolch, C.J., Lewis, R.J., Eds.; Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 2001; p. 49. [Google Scholar]
- Pizarro, G.; Alarcón, C.; Franco, J.M.; Escalera, L.; Reguera, B.; Vidal, G.; Palma, M.; Guzmán, L. Distribución espacial de Dinophysis spp. y detección de toxinas DSP en el agua mediante resinas DIAION (verano 2006, X región de Chile). Cienc. Tecnol. Mar. 2011, 34, 31–48. [Google Scholar]
- Fux, E.; Smith, J.L.; Tong, M.; Guzmán, L.; Anderson, D.M. Toxin profiles of five geographical isolates of Dinophysis spp. from North and South America. Toxicon 2011, 57, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Villarroel, O. Detección de toxina paralizante, diarreica y amnésica en mariscos de la XI región por cromatografía de alta resolución (HPLC) y bioensayo de ratón. Cienc. Tecnol. Mar. 2004, 27, 33–42. [Google Scholar]
- Yasumoto, T.; Takizawa, A. Fluorometric measurement of yessotoxins in shellfish by highpressure liquid chromatography. Biosci. Biotechnol. Biochem. 1997, 61, 1775–1777. [Google Scholar] [CrossRef] [PubMed]
- López-Rivera, A.; O’callaghan, K.; Moriarty, M.; O’driscoll, D.; Hamilton, B.; Lehane, M.; James, K.; Furey, A. First evidence of azaspiracids (AZAs): A family of lipophilic polyether marine toxins in scallops (Argopecten purpuratus) and mussels (Mytilus chilensis) collected in two regions of Chile. Toxicon 2010, 55, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Díaz, P.; Molinet, C.; Caceres, M.A.; Valle-Levinson, A. Seasonal and intratidal distribution of Dinophysis spp. in a Chilean fjord. Harmful Algae 2011, 10, 155–164. [Google Scholar] [CrossRef]
- Dolédec, S.; Chessel, D.; Gimaret-Carpentier, C. Niche separation in community analysis: A new method. Ecology 2000, 81, 2914–2927. [Google Scholar] [CrossRef]
- Karasiewicz, S.; Dolédec, S.; Lefebvre, S. Within outlying mean indexes: Refining the OMI analysis for the realized niche decomposition. PeerJ 2017, 5, e3364. [Google Scholar] [CrossRef] [PubMed]
- Karasiewicz, S.; Breton, E.; Lefebvre, A.; Fariñas, T.H.; Lefebvre, S. Realized niche analysis of phytoplankton communities involving HAB: Phaeocystis spp. as a case study. Harmful Algae 2018, 72, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Park, M.G.; Kim, S.; Kim, H.S.; Myung, G.; Kang, Y.G.; Yih, W. First successful culture of the marine dinoflagellate Dinophysis acuminata. Aquat. Microb. Ecol. 2006, 45, 101–106. [Google Scholar] [CrossRef]
- Jennings, E.; Jones, S.; Arvola, L.; Staehr, P.A.; Gaiser, E.; Jones, I.D.; Weathers, K.; Weyhenmeyer, G.A.; Chiu, C.-Y.; de Eyto, E. Effects of weather-related episodic events in lakes: An analysis based on high frequency data. Freshw. Biol. 2012, 57, 589–601. [Google Scholar] [CrossRef]
- Reguera, B.; Velo-Suárez, L.; Raine, R.; Park, M.G. Harmful Dinophysis species: A review. Harmful Algae 2012, 14, 87–106. [Google Scholar] [CrossRef]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Henry, M.; Stevens, H. The vegan package. Commun. Ecol. Package 2007, 10, 631–637. [Google Scholar]
- Karasiewicz, S. Within Outlying Mean Indexes: Refining the OMI Analysis. R Package Version 0.9.7. Available online: https://cran.r-project.org/web/packages/subniche/subniche.pdf (accessed on 2 January 2018).
- Alves-de-Souza, C.; Varela, D.; Iriarte, J.L.; González, H.E.; Guillou, L. Infection dynamics of Amoebophryidae parasitoids on harmful dinoflagellates in a southern Chilean fjord dominated by diatoms. Aquat. Microb. Ecol. 2012, 66, 183–187. [Google Scholar] [CrossRef]
- Rodríguez, J.J.G.; Miron, A.S.; García, M.d.C.C.; Belarbi, E.H.; Camacho, F.G.; Chisti, Y.; Grima, E.M. Macronutrients requirements of the dinoflagellate Protoceratium reticulatum. Harmful Algae 2009, 8, 239–246. [Google Scholar] [CrossRef]
- Clément, A.; Lembeye, G.; Lassus, P.; Le Baut, C. Bloom superficial no tóxico de Dinophysis cf. acuminata en el fiordo Reloncaví. XIV Jornadas de Ciencias del Mar y I Jornada chilena de Salmonicultura. In Proceedings of the XIV Jornadas de Ciencias del Mar & I Jornada Chilena de Salmonicultura, Puerto Montt, Chile, 23–25 May 1994; p. 83. [Google Scholar]
- Lembeye, G.; Molinet, C.; Marcos, N.; Sfeir, A.; Clément, A.; Rojas, X. Seguimiento de la Toxicidad en Recursos Pesqueros de Importancia Comercial en la X y XI Región (Proyecto FIP-IT/97-49); Universidad Austral de Chile: Puerto Montt, Chile, 1997; p. 86. [Google Scholar]
- Uribe, J.C.; Guzmán, L.; Jara, S. Monitoreo Mensual de la Marea Roja en la XI y XII Regiones (Proyecto FIP-IT/93-16); Universidad de Magallanes: Punta Arenas, Chile, 1995; p. 282. [Google Scholar]
- Castillo, M.I.; Cifuentes, U.; Pizarro, O.; Djurfeldt, L.; Caceres, M. Seasonal hydrography and surface outflow in a fjord with a deep sill: The Reloncaví Fjord, Chile. Ocean Sci. 2016, 12, 533–544. [Google Scholar] [CrossRef]
- León-Muñoz, J.; Marcé, R.; Iriarte, J. Influence of hydrological regime of an Andean river on salinity, temperature and oxygen in a Patagonia fjord, Chile. N. Z. J. Mar. Freshw. 2013, 47, 515–528. [Google Scholar] [CrossRef] [Green Version]
- Paz, B.; Vázquez, J.A.; Riobó, P.; Franco, J.M. Study of the effect of temperature, irradiance and salinity on growth and yessotoxin production by the dinoflagellate Protoceratium reticulatum in culture by using a kinetic and factorial approach. Mar. Environ. Res. 2006, 62, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Röder, K.; Hantzsche, F.M.; Gebühr, C.; Miene, C.; Helbig, T.; Krock, B.; Hoppenrath, M.; Luckas, B.; Gerdts, G. Effects of salinity, temperature and nutrients on growth, cellular characteristics and yessotoxin production of Protoceratium reticulatum. Harmful Algae 2012, 15, 59–70. [Google Scholar] [CrossRef]
- Hatosy, S.M.; Martiny, J.B.; Sachdeva, R.; Steele, J.; Fuhrman, J.A.; Martiny, A.C. Beta diversity of marine bacteria depends on temporal scale. Ecology 2013, 94, 1898–1904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, C. Temporal scales of variability in pelagic environments and the response of phytoplankton. Freshw. Biol. 1990, 23, 25–53. [Google Scholar] [CrossRef]
- Alves-de-Souza, C.; Benevides, T.S.; Santos, J.B.; Von Dassow, P.; Guillou, L.; Menezes, M. Does environmental heterogeneity explain temporal β diversity of small eukaryotic phytoplankton? Example from a tropical eutrophic coastal lagoon. J. Plankton Res. 2017, 39, 698–714. [Google Scholar] [CrossRef]
- Alves-de-Souza, C.; Benevides, T.; Menezes, M.; Jeanthon, C.; Guillou, L. First report of vampyrellid predator-prey dynamics in a marine system. ISME J. 2019, in press. [Google Scholar] [CrossRef] [PubMed]
- Garreaud, R. The Andes climate and weather. Adv. Geosci. 2009, 22, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Montecinos, A.; Aceituno, P. Seasonality of the ENSO-related rainfall variability in central Chile and associated circulation anomalies. J. Clim. 2003, 16, 281–296. [Google Scholar] [CrossRef]
- Garreaud, R. Record-breaking climate anomalies lead to severe drought and environmental disruption in western Patagonia in 2016. Clim. Res. 2018, 74, 217–229. [Google Scholar] [CrossRef]
- Seeyave, S.; Probyn, T.; Pitcher, G.; Lucas, M.; Purdie, D. Nitrogen nutrition in assemblages dominated by Pseudo-nitzschia spp., Alexandrium catenella and Dinophysis acuminata off the west coast of South Africa. Mar. Ecol. Prog. Ser. 2009, 379, 91–107. [Google Scholar] [CrossRef]
- Hernández-Urcera, J.; Rial, P.; García-Portela, M.; Lourés, P.; Kilcoyne, J.; Rodríguez, F.; Fernández-Villamarín, A.; Reguera, B. Notes on the cultivation of two mixotrophic Dinophysis species and their ciliate prey Mesodinium rubrum. Toxins 2018, 10, 505. [Google Scholar] [CrossRef]
- Crawford, D.W. Mesodinium rubrum: The phytoplankter that wasn’t. Mar. Ecol. Prog. Ser. 1989, 58, 161–174. [Google Scholar] [CrossRef]
- Valle-Levinson, A.; Sarkar, N.; Sanay, R.; Soto, D.; León, J. Spatial structure of hydrography and flow in a Chilean fjord, Estuario Reloncaví. Estuar. Coasts 2007, 30, 113–126. [Google Scholar] [CrossRef]
- Peterson, A.T. Ecological niche conservatism: A time-structured review of evidence. J. Biogeogr. 2011, 38, 817–827. [Google Scholar] [CrossRef]
- Riisgaard, K.; Hansen, P.J. Role of food uptake for photosynthesis, growth and survival of the mixotrophic dinoflagellate Dinophysis acuminata. Mar. Ecol. Prog. Ser. 2009, 381, 51–62. [Google Scholar] [CrossRef]
- Hansen, P.J.; Nielsen, L.T.; Johnson, M.; Berge, T.; Flynn, K.J. Acquired phototrophy in Mesodinium and Dinophysis—A review of cellular organization, prey selectivity, nutrient uptake and bioenergetics. Harmful Algae 2013, 28, 126–139. [Google Scholar] [CrossRef]
- Jiang, H.; Kulis, D.M.; Brosnahan, M.L.; Anderson, D.M. Behavioral and mechanistic characteristics of the predator-prey interaction between the dinoflagellate Dinophysis acuminata and the ciliate Mesodinium rubrum. Harmful Algae 2018, 77, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Gustafson Jr, D.E.; Stoecker, D.K.; Johnson, M.D.; Van Heukelem, W.F.; Sneider, K. Cryptophyte algae are robbed of their organelles by the marine ciliate Mesodinium rubrum. Nature 2000, 405, 1049. [Google Scholar] [CrossRef]
- Velo-Suárez, L.; González-Gil, S.; Pazos, Y.; Reguera, B. The growth season of Dinophysis acuminata in an upwelling system embayment: A conceptual model based on in situ measurements. Deep Sea Res. II Top. Stud. Oceanogr. 2014, 101, 141–151. [Google Scholar] [CrossRef]
- Moita, M.T.; Pazos, Y.; Rocha, C.; Nolasco, R.; Oliveira, P.B. Toward predicting Dinophysis blooms off NW Iberia: A decade of events. Harmful Algae 2016, 53, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Harred, L.B.; Campbell, L. Predicting harmful algal blooms: A case study with Dinophysis ovum in the Gulf of Mexico. J. Plankton Res. 2014, 36, 1434–1445. [Google Scholar] [CrossRef]
- Maestrini, S.Y. Bloom dynamics and ecophysiology of Dinophysis spp. In Physiological Ecology of Harmful Algal Blooms; Anderson, D., Cembella, A., Hallegraeff, G., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany; New York, NY, USA, 1998; pp. 243–266. [Google Scholar]
- Xie, H.; Lazure, P.; Gentien, P. Small scale retentive structures and Dinophysis. J. Mar. Syst. 2007, 64, 173–188. [Google Scholar] [CrossRef]
- Soudant, D.; Beliaeff, B.; Thomas, G. Explaining Dinophysis cf. acuminata abundance in Antifer (Normandy, France) using dynamic linear regression. Mar. Ecol. Prog. Ser. 1997, 156, 67–74. [Google Scholar] [CrossRef]
- Koukaras, K.; Nikolaidis, G. Dinophysis blooms in Greek coastal waters (Thermaikos Gulf, NW Aegean Sea). J. Plankton Res. 2004, 26, 445–457. [Google Scholar] [CrossRef]
- Velo-Suárez, L.; Reguera, B.; González-Gil, S.; Lunven, M.; Lazure, P.; Nézan, E.; Gentien, P. Application of a 3D Lagrangian model to explain the decline of a Dinophysis acuminata bloom in the Bay of Biscay. J. Mar. Syst. 2010, 83, 242–252. [Google Scholar] [CrossRef]
- Delmas, D.; Herbland, A.; Maestrini, S.Y. Do Dinophysis spp. come from the open sea along the French Atlantic coast? In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 489–494. [Google Scholar]
- Batifoulier, F.; Lazure, P.; Velo-Suarez, L.; Maurer, D.; Bonneton, P.; Charria, G.; Dupuy, C.; Gentien, P. Distribution of Dinophysis species in the Bay of Biscay and possible transport pathways to Arcachon Bay. J. Mar. Syst. 2013, 109, S273–S283. [Google Scholar] [CrossRef]
- Raine, R. A review of the biophysical interactions relevant to the promotion of HABs in stratified systems: The case study of Ireland. Deep Sea Res. II Top. Stud. Oceanogr. 2014, 101, 21–31. [Google Scholar] [CrossRef]
- Raine, R.; Farrell, H.; Gentien, P.; Fernand, L.; Lunven, M.; Reguera, B.; Gill, S.G. Transport of toxin producing dinoflagellate populations along the coast of Ireland within a seasonal coastal jet. ICES CM2010/N:05. Available online: http://www.ices.dk/sites/pub/CM%20Doccuments/CM-2010/N/N0510.pdf (accessed on 27 November 2018).
- Velo-Suarez, L.; Gonzalez-Gil, S.; Gentien, P.; Lunven, M.; Bechemin, C.; Fernand, L.; Raine, R.; Reguera, B. Thin layers of Pseudo-nitzschia spp. and the fate of Dinophysis acuminata during an upwelling-downwelling cycle in a Galician Ria. Limnol. Oceanogr. 2008, 53, 1816. [Google Scholar] [CrossRef]
- Díaz, P.A.; Reguera, B.; Ruiz-Villarreal, M.; Pazos, Y.; Velo-Suárez, L.; Berger, H.; Sourisseau, M. Climate variability and oceanographic settings associated with interannual variability in the initiation of Dinophysis acuminata blooms. Mar. Drugs 2013, 11, 2964–2981. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, O.; Lundve, B.; Johansen, M. Toxicity of Dinophysis spp. in relation to population density and environmental conditions on the Swedish west coast. Harmful Algae 2007, 6, 218–231. [Google Scholar] [CrossRef]
- Berdalet, E.; McManus, M.; Ross, O.; Burchard, H.; Chavez, F.; Jaffe, J.; Jenkinson, I.; Kudela, R.; Lips, I.; Lips, U. Understanding harmful algae in stratified systems: Review of progress and future directions. Deep Sea Res. II Top. Stud. Oceanogr. 2014, 101, 4–20. [Google Scholar] [CrossRef]
- Belgrano, A.; Lindahl, O.; Hernroth, B. North Atlantic Oscillation primary productivity and toxic phytoplankton in the Gullmar Fjord, Sweden (1985–1996). Proc. R. Soc. Lond. B Biol. Sci. 1999, 266, 425–430. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, O. Hydrodynamical processes: A trigger and source for flagellate blooms along the Skagerrak coasts? In Ecology of Fjords and Coastal Waters; Smayda, T.J., Shimizu, Y., Eds.; Elsevier Science Publishers BV: Amsterdam, The Netherlands, 1993; pp. 105–112. [Google Scholar]
- Meire, L.; Mortensen, J.; Rysgaard, S.; Bendtsen, J.; Boone, W.; Meire, P.; Meysman, F.J. Spring bloom dynamics in a subarctic fjord influenced by tidewater outlet glaciers (Godthåbsfjord, SW Greenland). J. Geophys. Res. Biogeosci. 2016, 121, 1581–1592. [Google Scholar] [CrossRef] [Green Version]
- Nishitani, G.; Nagai, S.; Takano, Y.; Sakiyama, S.; Baba, K.; Kamiyama, T. Growth characteristics and phylogenetic analysis of the marine dinoflagellate Dinophysis infundibulus (Dinophyceae). Aquat. Microb. Ecol. 2008, 52, 209–221. [Google Scholar] [CrossRef]
- Rodríguez, L. Revisión del fenómeno de Marea Roja en Chile. Rev. Biol. Mar. 1985, 21, 173–197. [Google Scholar]
- Toro, J.E.; Paredes, P.I.; Villagra, D.J.; Senn, C.M. Seasonal variation in the phytoplanktonic community, seston and environmental variables during a 2-year period and oyster growth at two mariculture sites, southern Chile. Mar. Ecol. 1999, 20, 63–89. [Google Scholar] [CrossRef]
- Campodónico, I.; Guzmán, L.; Lembeye, G. Una discoloración causada por el ciliado Mesodinium rubrum (Lohmann) en Ensenada Wilson, Magallanes. Ans. Inst. Pat. 1975, 6, 225–239. [Google Scholar]
- Jara, S. Observations on a red tide caused by Mesodinium rubrum (Lohmann) in the Aysén Fjord (Chile). Cienc. Tecnol. Mar. 1985, 9, 53–63. [Google Scholar]
- Lara, A.; Villalba, R.; Urrutia, R. A 400-year tree-ring record of the Puelo River summer-fall streamflow in the Valdivian Rainforest eco-region, Chile. Clim. Chang. 2008, 86, 331–356. [Google Scholar] [CrossRef]
- Utermöhl, H. Zur vendlhommung der quantitativen Phytoplankton-Methodik. Mitt. Int. Ver. Theor. Angew. Limnol. 1958, 9, 1–38. [Google Scholar]
- Climate Explorer. Available online: http://explorador.cr2.cl (accessed on 24 May 2018).
- NOAA: Global Climate Observing System (GCOS) Working Group on Surface Pressure (WG-SP). Available online: https://www.esrl.noaa.gov/psd/gcos_wgsp/Timeseries (accessed on 25 May 2018).
- CRAN: The Comprehensive R Archive Network. Available online: https://cran.r-project.org/ (accessed on 25 May 2018).
- Dray, S.; Dufour, A.-B. The ade4 package: Implementing the duality diagram for ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef]
- Xie, Y. A General-Purpose Package for Dynamic Report Generation in R. R Package Version 1.21. Available online: https://cran.r-project.org/web/packages/knitr/knitr.pdf (accessed on 14 April 2018).
- Kassambara, A.; Mundt, F. Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 1.0.5. Available online: https://cran.r-project.org/web/packages/factoextra/factoextra.pdf (accessed on 2 January 2018).
- Karasiewicz, S. Subniche Documentation for the Within Outlying Mean Indexes calculations (WitOMI). Available online: https://github.com/KarasiewiczStephane/WitOMI (accessed on 2 January 2018).
- Wickham, H. ggplot2—Elegant graphics for data analysis. J. Stat. Softw. 2010, 35, 65–88. [Google Scholar]
- Hothorn, T.; Zeileis, A. partykit: A modular toolkit for recursive partitioning in R. J. Mach. Learn. Res. 2015, 16, 3905–3909. [Google Scholar]
- Álvarez, G.; Uribe, E.; Díaz, R.; Braun, M.; Mariño, C.; Blanco, J. Bloom of the Yessotoxin producing dinoflagellate Protoceratium reticulatum (Dinophyceae) in Northern Chile. J. Sea Res. 2011, 65, 427–434. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alves-de-Souza, C.; Iriarte, J.L.; Mardones, J.I. Interannual Variability of Dinophysis acuminata and Protoceratium reticulatum in a Chilean Fjord: Insights from the Realized Niche Analysis. Toxins 2019, 11, 19. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11010019
Alves-de-Souza C, Iriarte JL, Mardones JI. Interannual Variability of Dinophysis acuminata and Protoceratium reticulatum in a Chilean Fjord: Insights from the Realized Niche Analysis. Toxins. 2019; 11(1):19. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11010019
Chicago/Turabian StyleAlves-de-Souza, Catharina, José Luis Iriarte, and Jorge I. Mardones. 2019. "Interannual Variability of Dinophysis acuminata and Protoceratium reticulatum in a Chilean Fjord: Insights from the Realized Niche Analysis" Toxins 11, no. 1: 19. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11010019