Cloning and Immunosuppressive Properties of an Acyl-Activating Enzyme from the Venom Apparatus of Tetrastichus brontispae (Hymenoptera: Eulophidae)


Abstract
:1. Introduction
2. Results
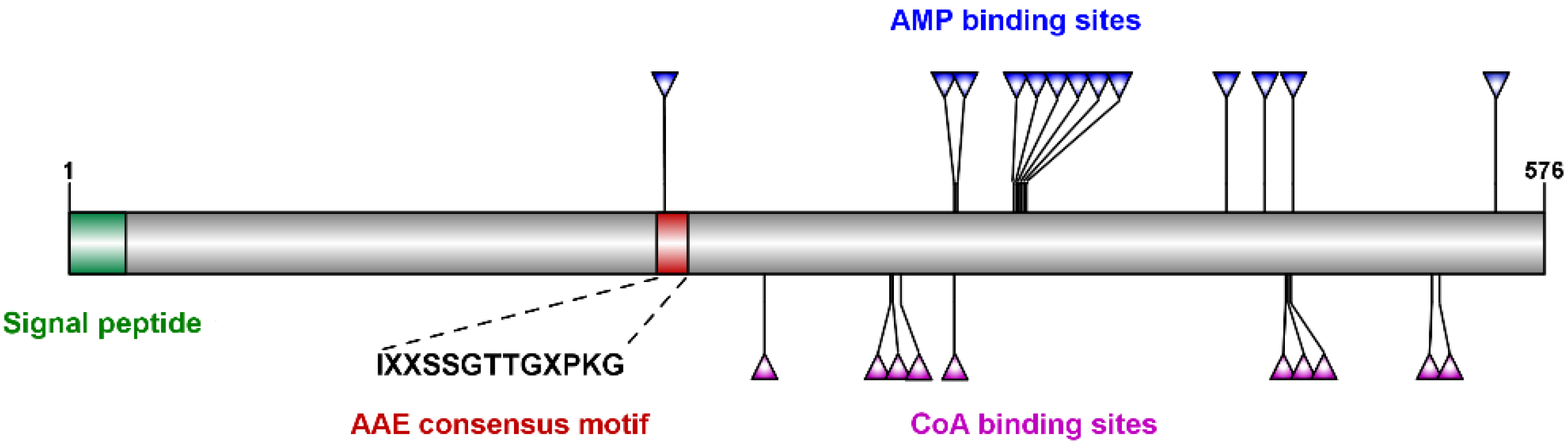
2.1. Characterization and Sequence, Phylogenetic and Motif Analyses of Tb4CL4-Like
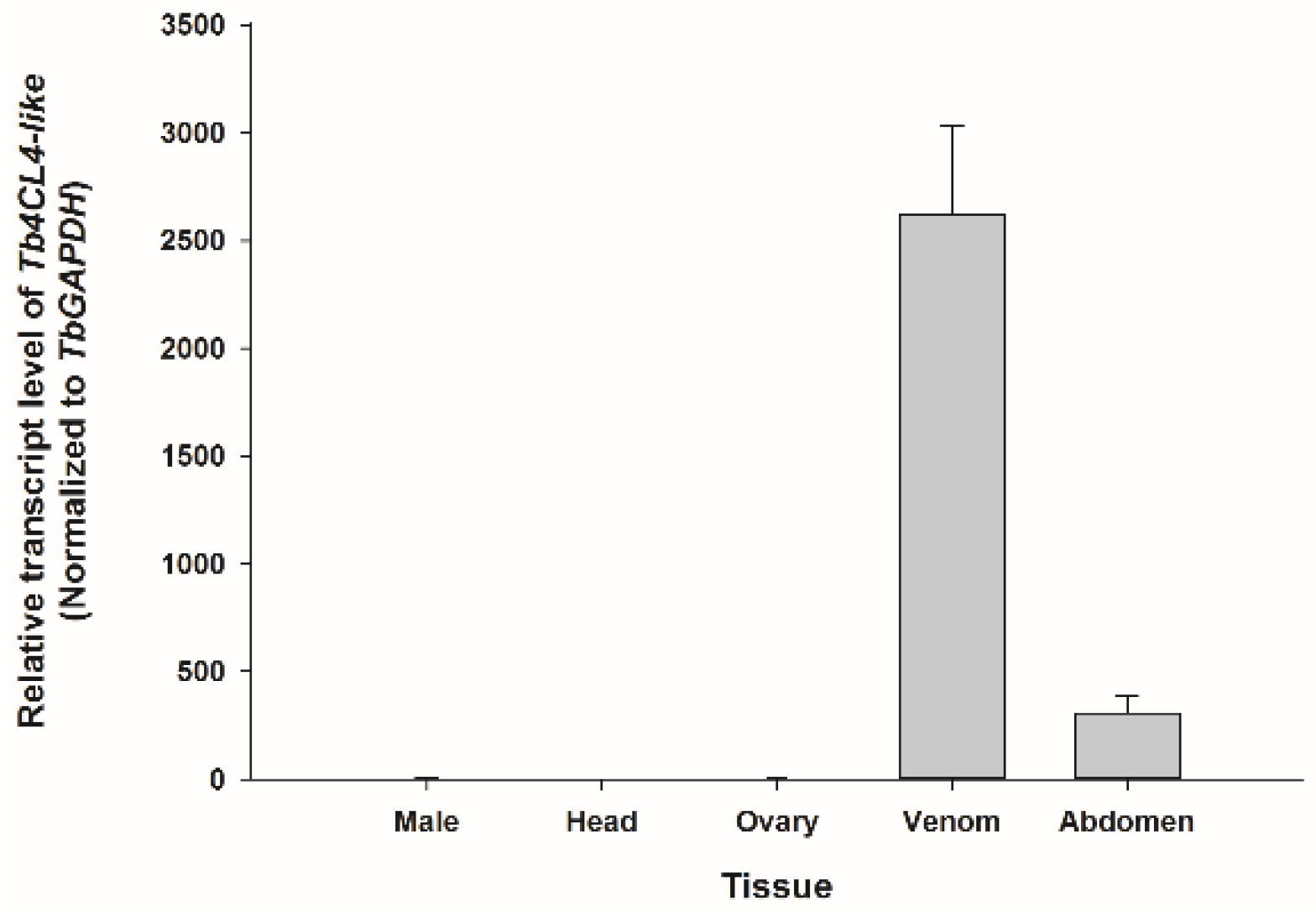
2.2. Transcription Profiles of Tb4CL4-Like in Different Tissues
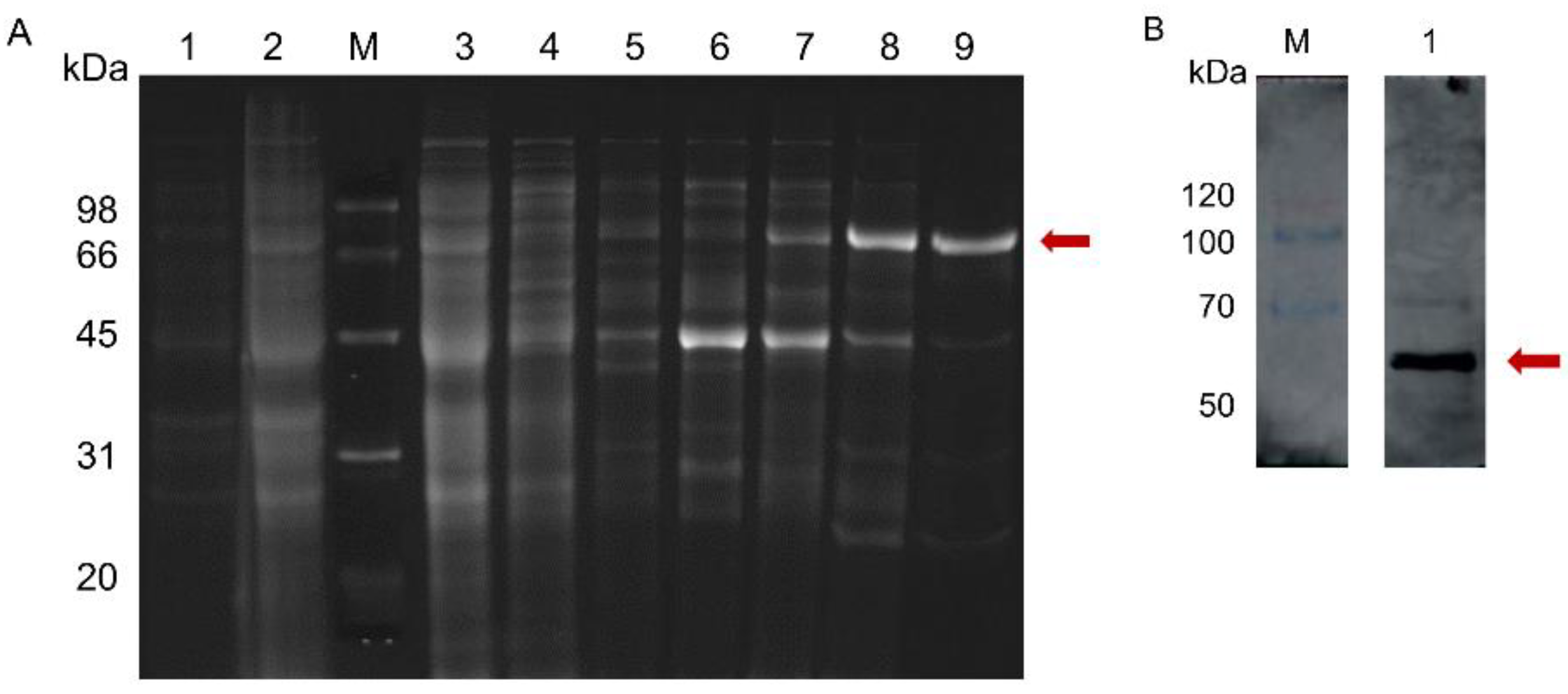
2.3. Tb4CL4-Like Expression, Purification and Western Blot Analysis
2.4. Effect of Tb4CL4-Like on Host Cell Encapsulation Response
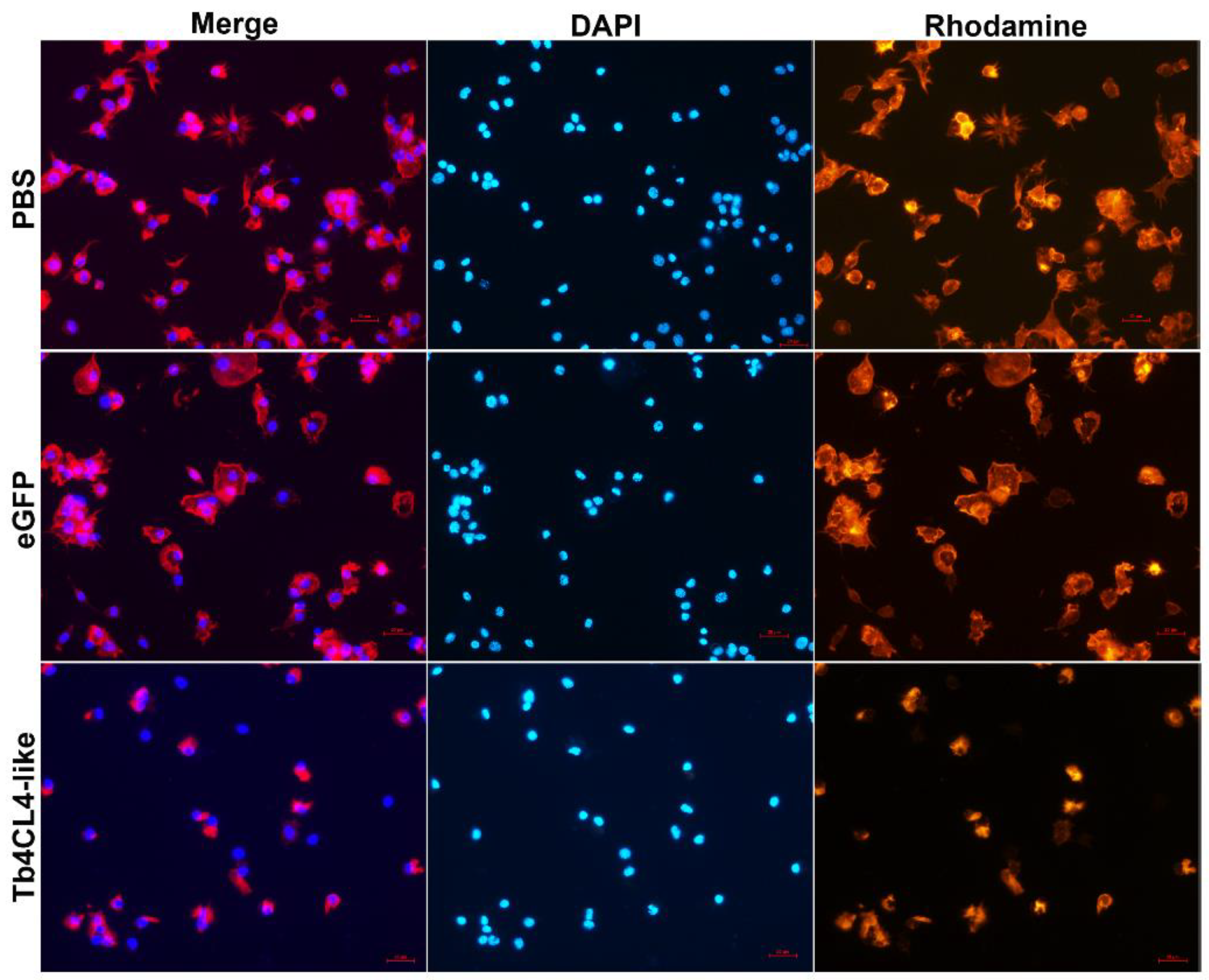
2.5. Effect of Tb4CL4-Like on Host Hemocyte Cytoskeleton, F-Actin Content and Spreading Ability
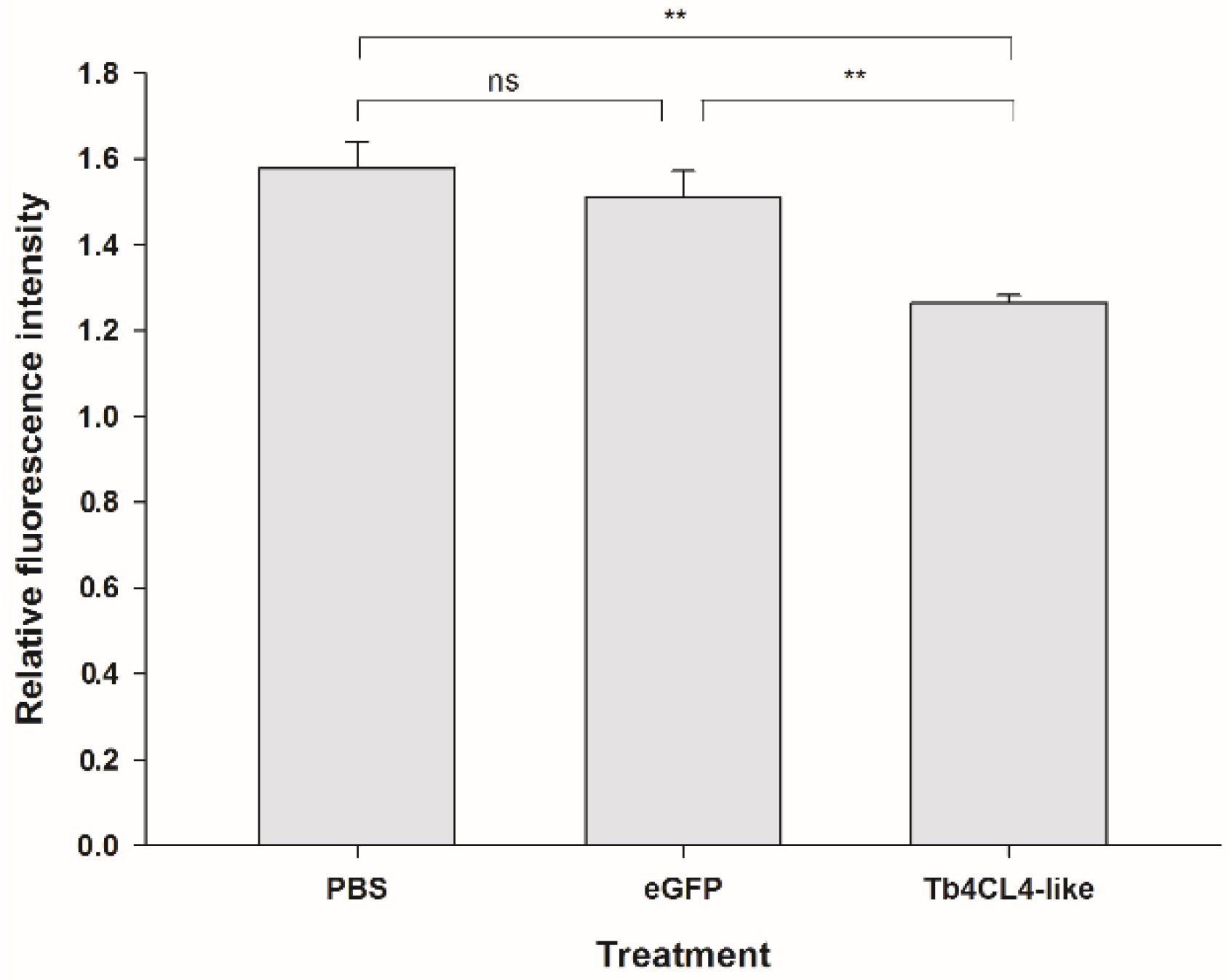
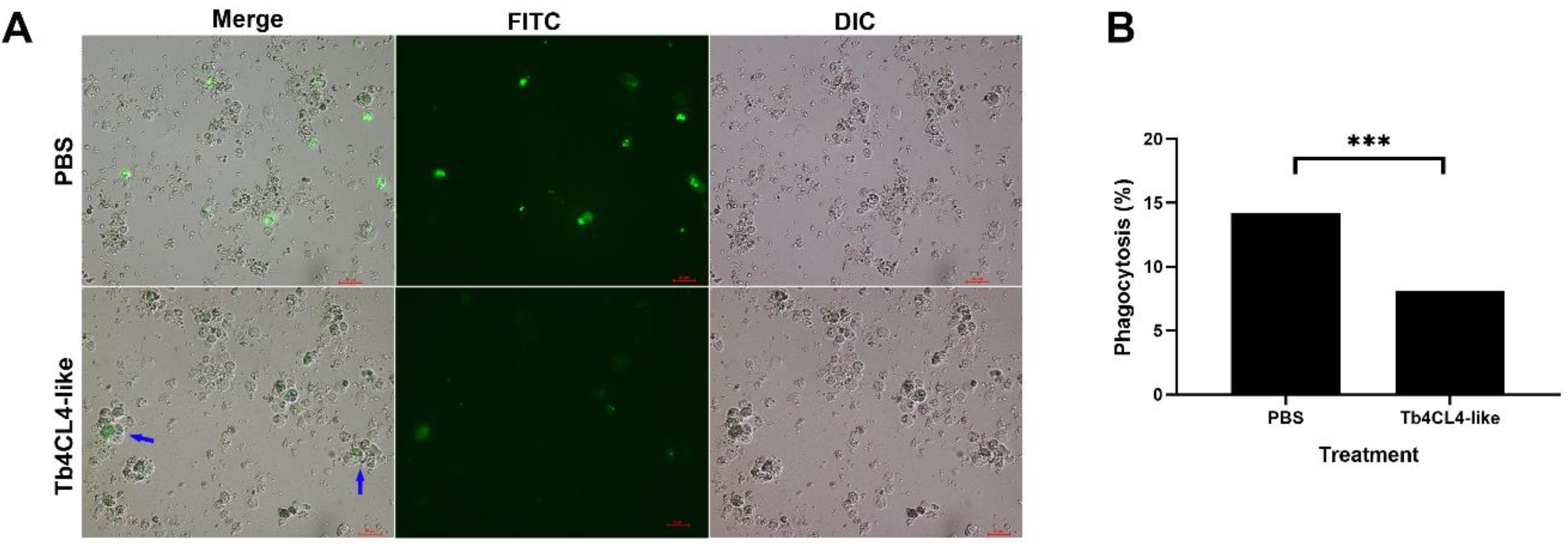
2.6. Effect of Tb4CL4-Like on Phagocytosis
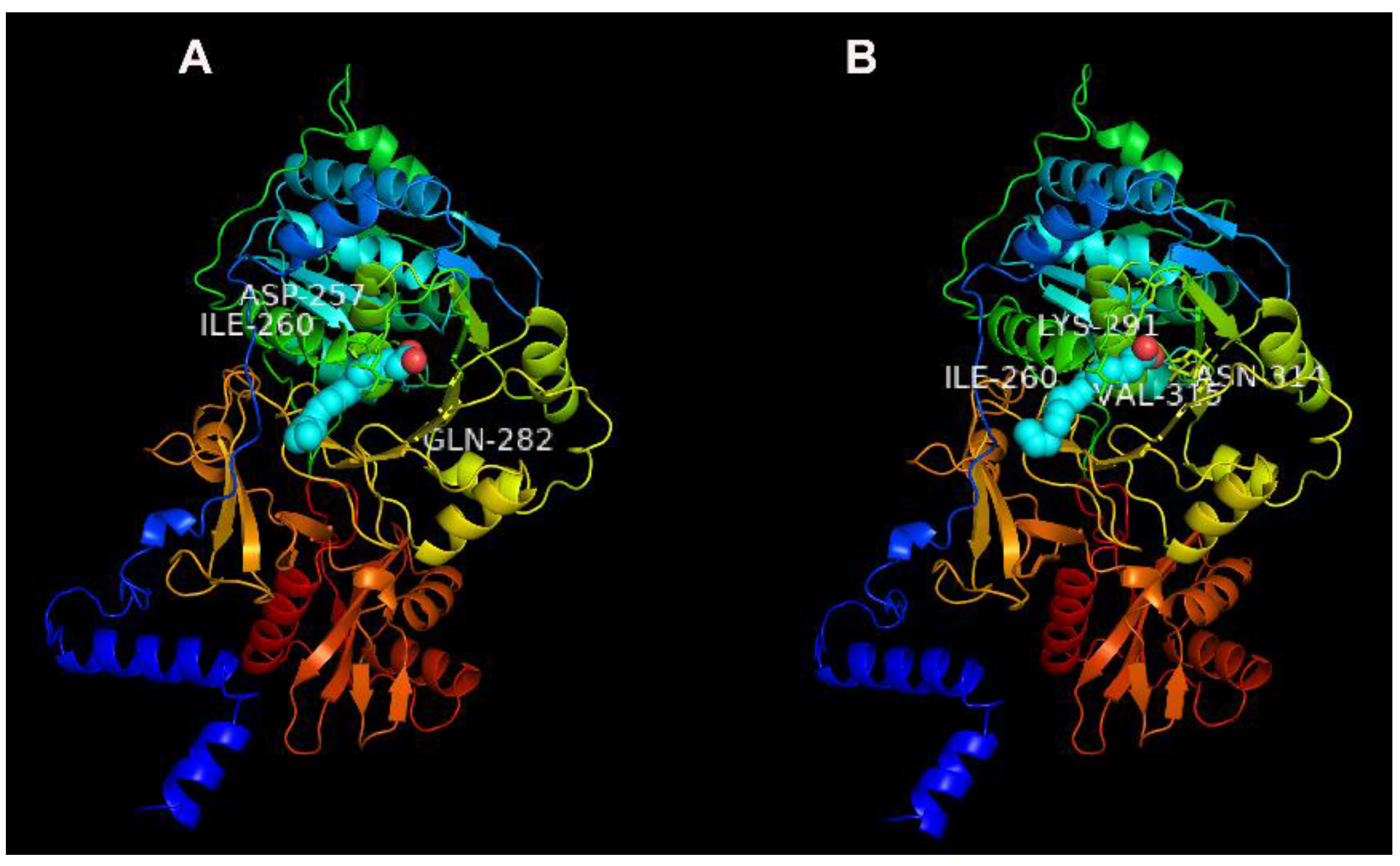
2.7. Potential Binding Characteristics of Tb4CL4-Like
3. Discussion
4. Materials and Methods
4.1. Insect Rearing
4.2. Cloning the Full-Length of the Tb4CL4-Like Gene
4.3. Sequence Analysis
4.4. Quantitative Real-Time PCR (qRT-PCR) Analysis of the Tb4CL4-Like Tissue Expression Profile
4.5. Expression and Purification of Recombinant Tb4CL4-Like
4.6. SDS-PAGE and Immunoblot Analysis
4.7. In Vitro Encapsulation Assay
4.8. Effect of Tb4CL4-Like on F-Actin of Hemocytes
4.9. Hemocyte Spreading Assay
4.10. In Vivo Phagocytosis Assay
4.11. Molecular Docking of Tb4CL4-Like
4.12. Statistical Analysis
Supplementary Materials
Data and Materials Availability
Author Contributions
Funding
Conflicts of Interest
References
- Moreau, S.J.; Asgari, S. Venom proteins from parasitoid wasps and their biological functions. Toxins 2015, 7, 2385–2412. [Google Scholar] [CrossRef]
- Strand, M.R. The insect cellular immune response. Insect Sci. 2008, 15, 1–14. [Google Scholar] [CrossRef]
- Asgari, S.; Rivers, D.B. Venom proteins from endoparasitoid wasps and their role in host-parasite interactions. Annu. Rev. Entomol. 2011, 56, 313–335. [Google Scholar] [CrossRef]
- Poirié, M.; Colinet, D.; Gatti, J.-L. Insights into function and evolution of parasitoid wasp venoms. Curr. Opin. Insect Sci. 2014, 6, 52–60. [Google Scholar] [CrossRef]
- Colinet, D.; Mathe-Hubert, H.; Allemand, R.; Gatti, J.L.; Poirie, M. Variability of venom components in immune suppressive parasitoid wasps: From a phylogenetic to a population approach. J. Insect Physiol. 2013, 59, 205–212. [Google Scholar] [CrossRef]
- Kaeslin, M.; Reinhard, M.; Buhler, D.; Roth, T.; Pfister-Wilhelm, R.; Lanzrein, B. Venom of the egg-larval parasitoid Chelonus inanitus is a complex mixture and has multiple biological effects. J. Insect Physiol. 2010, 56, 686–694. [Google Scholar] [CrossRef]
- Asgari, S. Venoms from endoparasitoids. In Parasitoid Viruses Symbionts and Pathogens; Beckage, N.E., Drezen, J.-M., Eds.; Academic Press: London, UK, 2012; pp. 217–231. [Google Scholar]
- Casewell, N.R.; Wuster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef]
- Dani, M.P.; Richards, E.H. Cloning and expression of the gene for an insect haemocyte anti-aggregation protein (VPr3), from the venom of the endoparasitic wasp, Pimpla hypochondriaca. Arch. Insect Biochem. Physiol. 2009, 71, 191–204. [Google Scholar] [CrossRef]
- Richards, E.H.; Dani, M.P.; Bradish, H. Immunosuppressive properties of a protein (rVPr1) from the venom of the endoparasitic wasp, Pimpla hypochondriaca: Mechanism of action and potential use for improving biological control strategies. J. Insect Physiol. 2013, 59, 213–222. [Google Scholar] [CrossRef]
- Dani, M.P.; Richards, E.H. Identification, cloning and expression of a second gene (vpr1) from the venom of the endoparasitic wasp, Pimpla hypochondriaca that displays immunosuppressive activity. J. Insect Physiol. 2010, 56, 195–203. [Google Scholar] [CrossRef]
- Colinet, D.; Schmitz, A.; Depoix, D.; Crochard, D.; Poirie, M. Convergent use of RhoGAP toxins by eukaryotic parasites and bacterial pathogens. PLoS Pathog. 2007, 3, e203. [Google Scholar] [CrossRef]
- Wang, L.; Fang, Q.; Qian, C.; Wang, F.; Yu, X.Q.; Ye, G. Inhibition of host cell encapsulation through inhibiting immune gene expression by the parasitic wasp venom calreticulin. Insect Biochem. Mol. Biol. 2013, 43, 936–946. [Google Scholar] [CrossRef]
- Yan, Z.C.; Fang, Q.; Liu, Y.; Xiao, S.; Yang, L.; Wang, F.; An, C.J.; Werren, J.H.; Ye, G.Y. A venom serpin splicing isoform of the endoparasitoid wasp Pteromalus puparum suppresses host prophenoloxidase cascade by forming complexes with host hemolymph proteinase. J. Biol. Chem. 2017, 292, 1038–1051. [Google Scholar] [CrossRef]
- Colinet, D.; Cazes, D.; Belghazi, M.; Gatti, J.L.; Poirie, M. Extracellular superoxide dismutase in insects: Characterization, function, and interspecific variation in parasitoid wasp venom. J. Biol. Chem. 2011, 286, 40110–40121. [Google Scholar] [CrossRef]
- Coulette, Q.; Lemauf, S.; Colinet, D.; Prevost, G.; Anselme, C.; Poirie, M.; Gatti, J.L. Biochemical characterization and comparison of aspartylglucosaminidases secreted in venom of the parasitoid wasps Asobara tabida and Leptopilina heterotoma. PLoS ONE 2017, 12, e0181940. [Google Scholar] [CrossRef]
- Chen, Q.; Peng, Z.; Xu, C.; Tang, C.; Lu, B.; Jin, Q.; Wen, H.; Wan, F. Biological assessment of Tetrastichus brontispae, a pupal parasitoid of coconut leaf beetle Brontispa longissima. Biocontrol Sci. Technol. 2010, 20, 283–295. [Google Scholar] [CrossRef]
- Tang, B.Z.; Xu, L.; Hou, Y.M. Effects of rearing conditions on the parasitism of Tetrastichus brontispae on its pupal host Octodonta nipae. BioControl 2014, 59, 647–657. [Google Scholar] [CrossRef]
- Meng, E.; Qiao, T.; Tang, B.Z.; Hou, Y.M.; Yu, W.Z.; Chen, Z.M. Effects of ovarian fluid, venom and egg surface characteristics of Tetrastichus brontispae (Hymenoptera: Eulophidae) on the immune response of Octodonta nipae (Coleoptera: Chrysomelidae). J. Insect Physiol. 2018, 109, 125–137. [Google Scholar] [CrossRef]
- Tang, B.Z.; Meng, E.; Zhang, H.J.; Zhang, X.M.; Asgari, S.; Lin, Y.P.; Lin, Y.Y.; Peng, Z.Q.; Qiao, T.; Zhang, X.F.; et al. Combination of label-free quantitative proteomics and transcriptomics reveals intraspecific venom variation between the two strains of Tetrastichus brontispae, a parasitoid of two invasive beetles. J. Proteomics 2019, 192, 37–53. [Google Scholar] [CrossRef]
- Shockey, J.M.; Fulda, M.S.; Browse, J. Arabidopsis contains a large superfamily of acyl-activating enzymes. Phylogenetic and biochemical analysis reveals a new class of acyl-coenzyme a synthetases. Plant. Physiol. 2003, 132, 1065–1076. [Google Scholar] [CrossRef]
- Schmelz, S.; Naismith, J.H. Adenylate-forming enzymes. Curr. Opin. Struct. Biol. 2009, 19, 666–671. [Google Scholar] [CrossRef]
- Clark, L.; Leatherby, D.; Krilich, E.; Ropelewski, A.J.; Perozich, J. In silico analysis of class I adenylate-forming enzymes reveals family and group-specific conservations. PLoS ONE 2018, 13, e0203218. [Google Scholar] [CrossRef]
- Hamberger, B.; Hahlbrock, K. The 4-coumarate: CoA ligase gene family in Arabidopsis thaliana comprises one rare, sinapate-activating and three commonly occurring isoenzymes. Proc. Natl. Acad. Sci. USA 2004, 101, 2209–2214. [Google Scholar] [CrossRef]
- Oba, Y.; Ojika, M.; Inouye, S. Firefly luciferase is a bifunctional enzyme: ATP-dependent monooxygenase and a long chain fatty acyl-CoA synthetase. FEBS Lett. 2003, 540, 251–254. [Google Scholar] [CrossRef]
- Qian, C.; Fang, Q.; Wang, L.; Ye, G.Y. Molecular cloning and functional studies of two Kazal-type serine protease inhibitors specifically expressed by Nasonia vitripennis venom apparatus. Toxins 2015, 7, 2888–2905. [Google Scholar] [CrossRef]
- Qian, C.; Liang, D.; Liu, Y.; Wang, P.; Kausar, S.; Wei, G.Q.; Zhu, B.J.; Wang, L.; Liu, C.L. Identification of a small pacifastin protease inhibitor from Nasonia vitripennis venom that inhibits humoral immunity of host (Musca domestica). Toxicon 2017, 131, 54–62. [Google Scholar] [CrossRef]
- Tsakas, S.; Marmaras, V.J. Insect immunity and its signalling: An overview. Invertebr. Surviv. J. 2010, 7, 228–238. [Google Scholar]
- Boutros, M.; Agaisse, H.; Perrimon, N. Sequential activation of signaling pathways during innate immune responses in Drosophila. Dev. Cell 2002, 3, 711–722. [Google Scholar] [CrossRef]
- Meng, E.; Tang, B.Z.; Hou, Y.M.; Chen, X.X.; Chen, J.T.; Yu, X.Q. Altered immune function of Octodonta nipae (Maulik) to its pupal endoparasitoid, Tetrastichus brontispae Ferriere. Comp. Biochem. Physiol. Part. B 2016, 198, 100–109. [Google Scholar] [CrossRef]
- Amaya, K.E.; Asgari, S.; Jung, R.; Hongskula, M.; Beckage, N.E. Parasitization of Manduca sexta larvae by the parasitoid wasp Cotesia congregata induces an impaired host immune response. J. Insect Physiol. 2005, 51, 505–512. [Google Scholar] [CrossRef]
- Asgari, S.; Hellers, M.; Schmidt, O. Host haemocyte inactivation by an insect parasitoid: Transient expression of a polydnavirus gene. J. Gen. Virol. 1996, 77, 2653–2662. [Google Scholar] [CrossRef] [PubMed]
- Nalini, M.; Ibrahim, A.M.A.; Hwang, I.; Kim, Y. Altered actin polymerization of Plutella xylostella (L.) in response to ovarian calyx components of an endoparasitoid Cotesia plutellae (Kurdjumov). Physiol. Entomol. 2009, 34, 110–118. [Google Scholar] [CrossRef]
- Wu, M.L.; Ye, G.Y.; Zhu, J.Y.; Chen, X.X.; Hu, C. Isolation and characterization of an immunosuppressive protein from venom of the pupa-specific endoparasitoid Pteromalus puparum. J. Invertebr. Pathol. 2008, 99, 186–191. [Google Scholar] [CrossRef]
- Cai, J.; Ye, G.Y.; Hu, C. Parasitism of Pieris rapae (Lepidoptera: Pieridae) by a pupal endoparasitoid, Pteromalus puparum (Hymenoptera: Pteromalidae): Effects of parasitization and venom on host hemocytes. J. Insect Physiol. 2004, 50, 315–322. [Google Scholar] [CrossRef]
- Kim, Y.; Ahmed, S.; Stanley, D.; An, C. Eicosanoid-mediated immunity in insects. Dev. Comp. Immunol. 2018, 83, 130–143. [Google Scholar] [CrossRef]
- Stanley, D. Prostaglandins and other eicosanoids in insects: Biological significance. Annu. Rev. Entomol. 2006, 51, 25–44. [Google Scholar] [CrossRef]
- Stanley, D.; Kim, Y. Prostaglandins and other eicosanoids in insects: Biosynthesis and biological actions. Front. Physiol. 2019, 9, 1927. [Google Scholar] [CrossRef] [Green Version]
- Li, J.L.; Zhang, X.; Hou, Y.M.; Tang, B.Z. Effects of multiple mating on the fecundity of an invasive pest (Octodonta nipae): The existence of an intermediate optimal female mating rate. Physiol. Entomol. 2014, 39, 348–354. [Google Scholar] [CrossRef]
- Tang, B.Z.; Chen, J.; Hou, Y.M.; Meng, E. Transcriptome immune analysis of the invasive beetle Octodonta nipae (Maulik) (Coleoptera: Chrysomelidae) parasitized by Tetrastichus brontispae Ferrière (Hymenoptera: Eulophidae). PLoS ONE 2014, 9, e91482. [Google Scholar] [CrossRef]
- Hu, J.; Xu, Q.; Hu, S.; Yu, X.; Liang, Z.; Zhang, W. Hemomucin, an O-glycosylated protein on embryos of the wasp Macrocentrus cingulum that protects it against encapsulation by hemocytes of the host Ostrinia furnacalis. J. Innate Immun. 2014, 6, 663–675. [Google Scholar] [CrossRef]
- Zhao, H.; Liu, J.; Hu, J. An improved method on encapsulation assays by larval hemocytes from Ostrinia furnacalis in vitro. Zool. Res. 2007, 28, 675–680. [Google Scholar]
- Teng, Z.W.; Xu, G.; Gan, S.Y.; Chen, X.; Fang, Q.; Ye, G.Y. Effects of the endoparasitoid Cotesia chilonis (Hymenoptera: Braconidae) parasitism, venom, and calyx fluid on cellular and humoral immunity of its host Chilo suppressalis (Lepidoptera: Crambidae) larvae. J. Insect Physiol. 2016, 85, 46–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.J.; Lin, Y.P.; Liu, M.; Liang, X.Y.; Ji, Y.N.; Tang, B.Z.; Hou, Y.M. Functional conservation and division of two single-carbohydrate-recognition domain C-type lectins from the nipa palm hispid beetle Octodonta nipae (Maulik). Dev. Comp. Immunol. 2019, 100, 103416. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligands | Tb4CL4-Like | |
|---|---|---|
| Binding Energy (kcal/mol) | Ki (μM) | |
| Arachidonic acid | −8.06 | 1.23 |
| Caffeic acid | −6.40 | 20.27 |
| Ferulic acid | −6.47 | 18.03 |
| Linoleic acid | −6.37 | 21.39 |
| Linolenic acid | −8.35 | 0.72 |
| Palmitic acid | −8.69 | 0.43 |
| p-Coumarate | −6.41 | 19.92 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.-M.; Zhang, H.-J.; Liu, M.; Liu, B.; Zhang, X.-F.; Ma, C.-J.; Fu, T.-T.; Hou, Y.-M.; Tang, B.-Z. Cloning and Immunosuppressive Properties of an Acyl-Activating Enzyme from the Venom Apparatus of Tetrastichus brontispae (Hymenoptera: Eulophidae). Toxins 2019, 11, 672. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110672
Zhang X-M, Zhang H-J, Liu M, Liu B, Zhang X-F, Ma C-J, Fu T-T, Hou Y-M, Tang B-Z. Cloning and Immunosuppressive Properties of an Acyl-Activating Enzyme from the Venom Apparatus of Tetrastichus brontispae (Hymenoptera: Eulophidae). Toxins. 2019; 11(11):672. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110672
Chicago/Turabian StyleZhang, Xiao-Mei, Hua-Jian Zhang, Min Liu, Bin Liu, Xia-Fang Zhang, Cheng-Jun Ma, Ting-Ting Fu, You-Ming Hou, and Bao-Zhen Tang. 2019. "Cloning and Immunosuppressive Properties of an Acyl-Activating Enzyme from the Venom Apparatus of Tetrastichus brontispae (Hymenoptera: Eulophidae)" Toxins 11, no. 11: 672. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110672