AflSte20 Regulates Morphogenesis, Stress Response, and Aflatoxin Biosynthesis of Aspergillus flavus

Abstract
:1. Introduction
2. Results
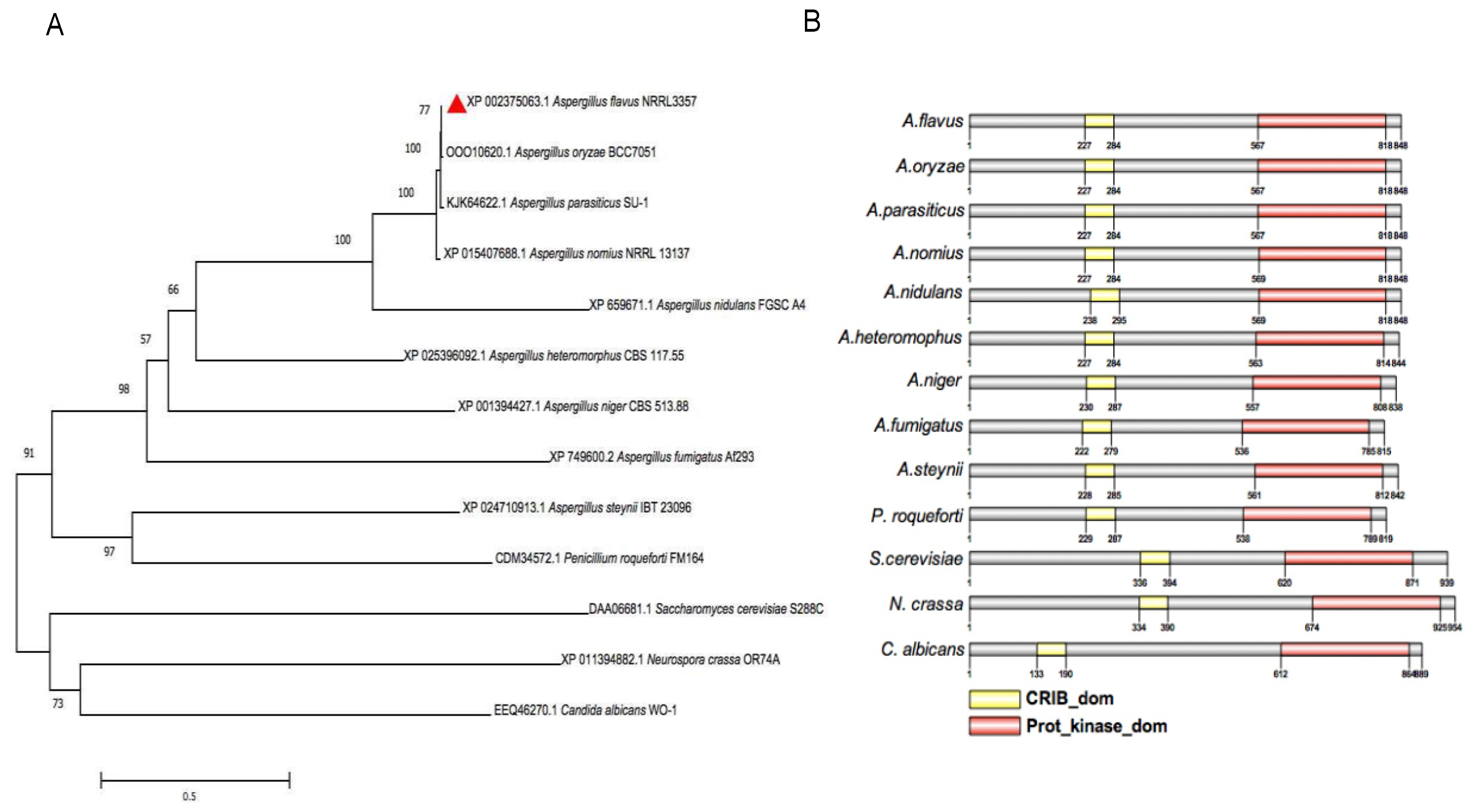
2.1. Sequences Analysis of AflSte20 in A. flavus
2.2. AflSte20 is Involved in Vegetative Growth and Conidial Production
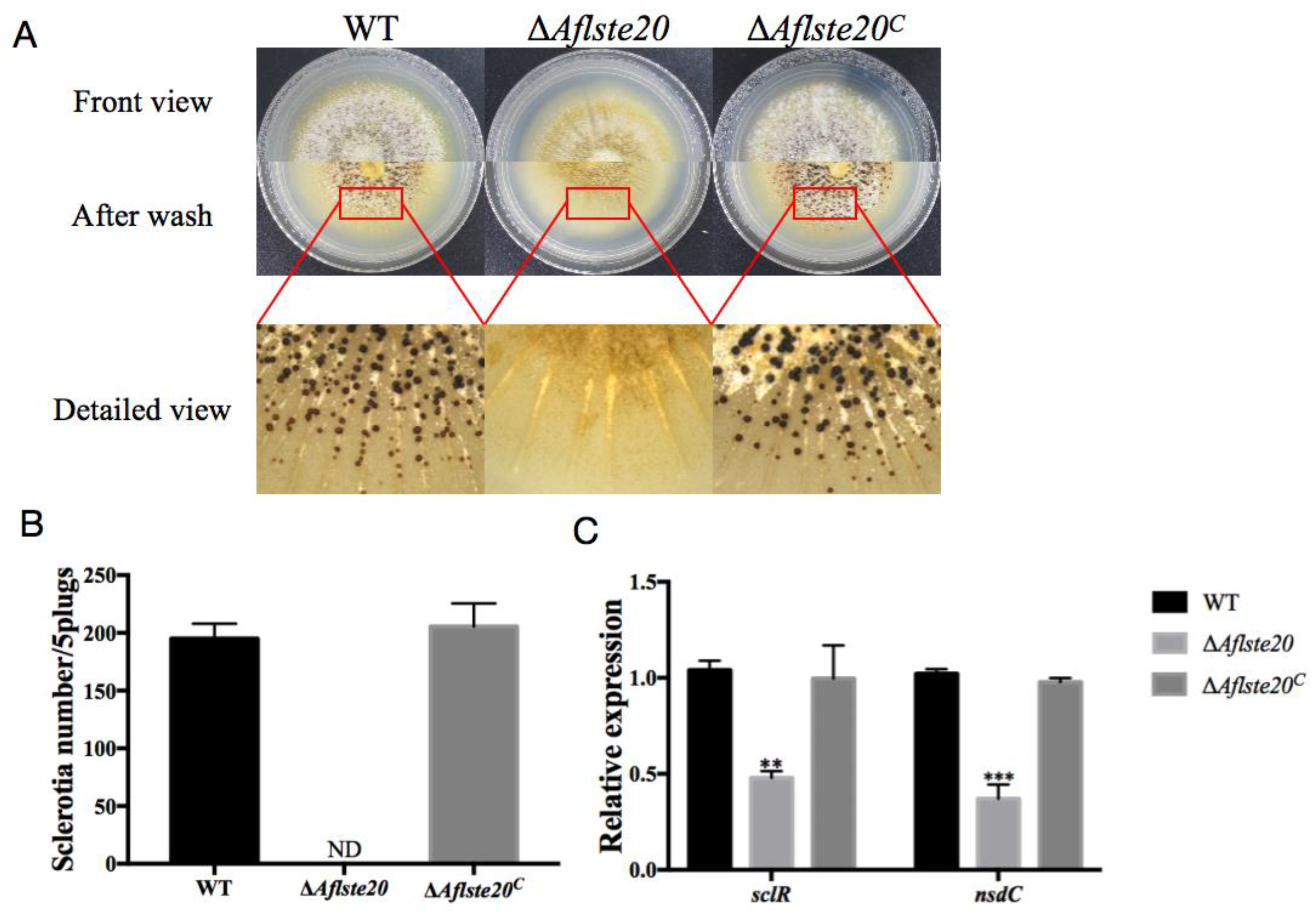
2.3. AflSte20 Mediates Sclerotia Formation in A. flavus
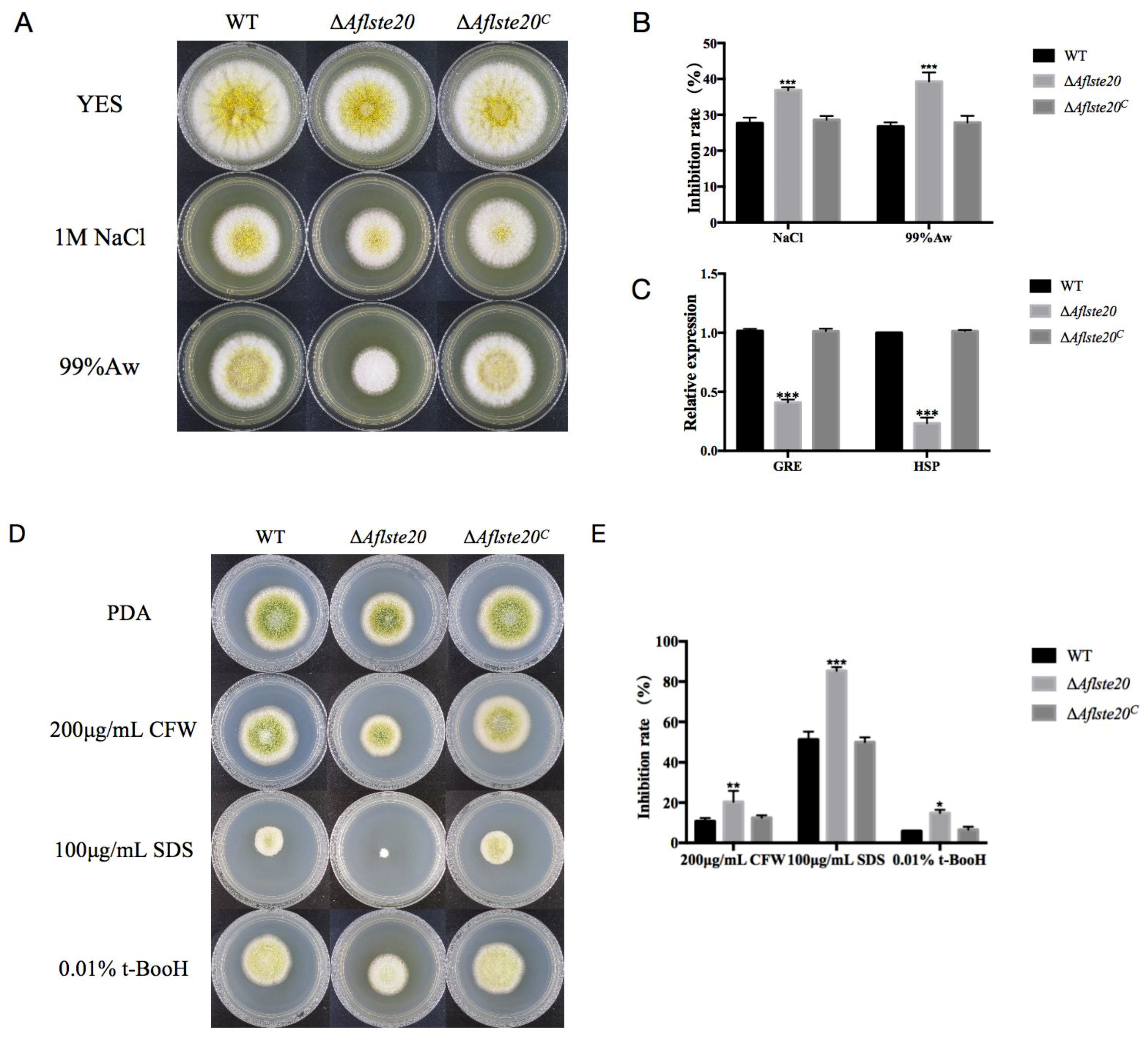
2.4. AflSte20 Participates in the Response to Different Stresses in A. flavus
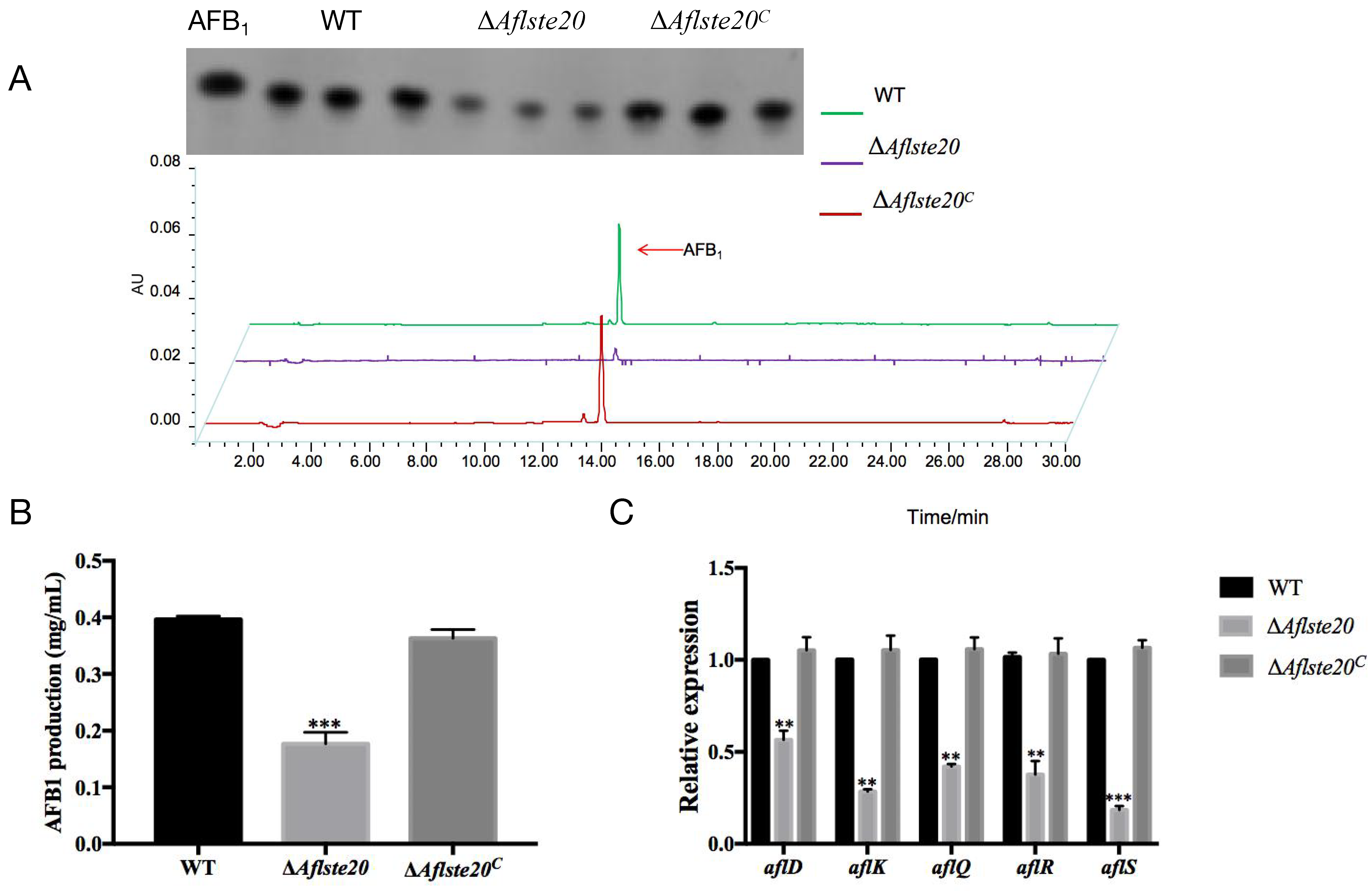
2.5. AflSte20 Positively Regulates Aflatoxin Biosynthesis
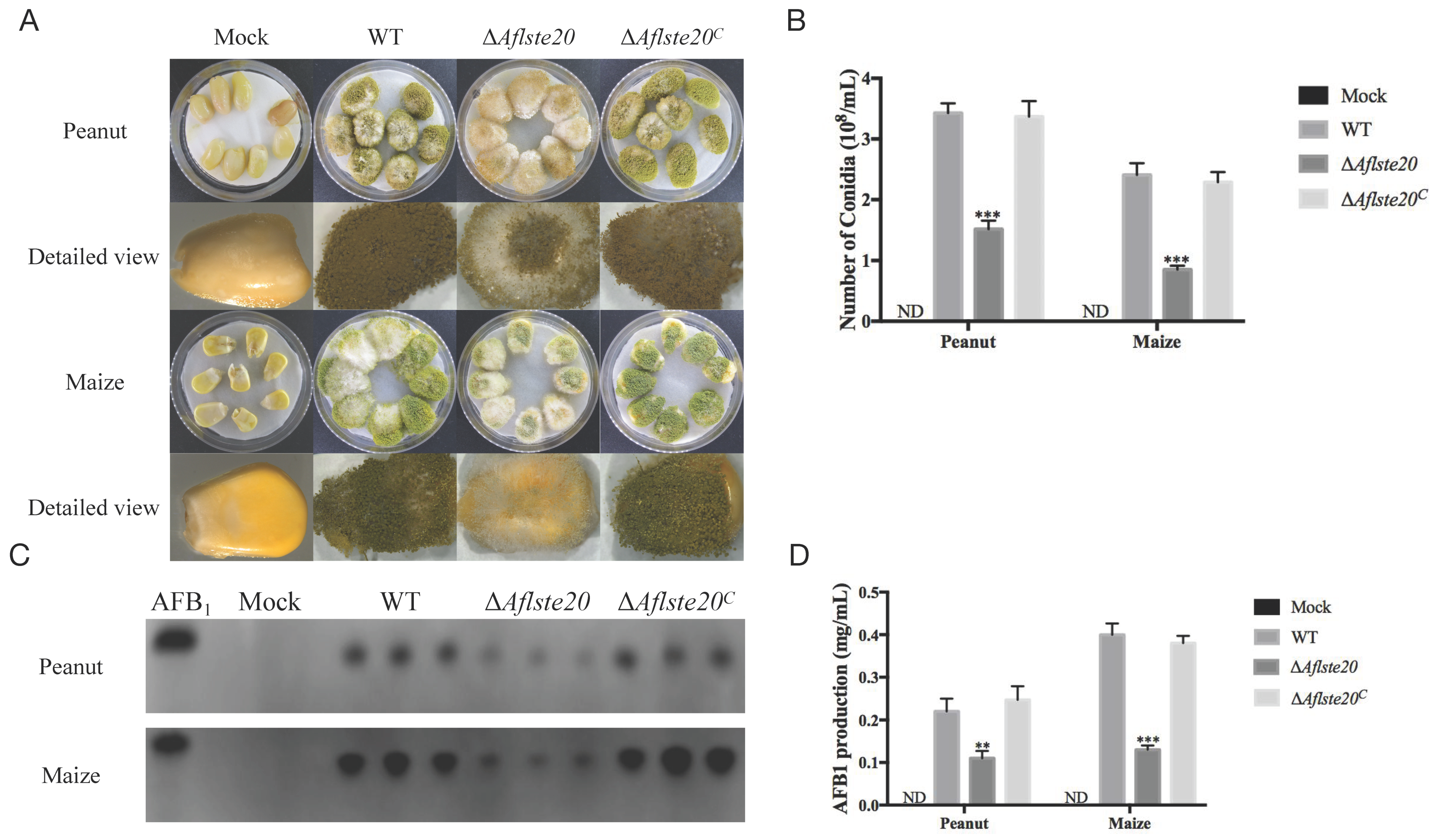
2.6. AflSte20 Has an Influence on Conidiation and Aflatoxin Biosynthesis to Crop Seeds
3. Discussion
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. Bioinformatics Analysis
4.3. Gene Deletion and Complementation
4.4. Measurement of Conidia, Conidiophores, and Sclerotia
4.5. Stress Assay
4.6. Determination of AFB1 Production
4.7. Conidiation and Aflatoxin Biosynthesis of A. flavus in Crop Seeds
4.8. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Klich, M.A. Aspergillus flavus: The major producer of aflatoxin. Mol. Plant Pathol. 2010, 8, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Hedayati, M.T.; Pasqualotto, A.C.; Warn, P.A.; Bowyer, P.; Denning, D.W. Aspergillus flavus: Human pathogen, allergen and mycotoxin producer. Microbiology 2007, 153, 1677–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, S.; Manavathu, E.K.; Chandrasekar, P.H. Aspergillus flavus: An emerging non-fumigatus aspergillus species of significance. Mycoses 2010, 52, 206–222. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, A.; Sen, M.M.; Gupta-Bhattacharya, S.; Chanda, S. Volumetric assessment of airborne fungi in two sections of a rural indoor dairy cattle shed. Environ. Int. 2004, 29, 1071–1078. [Google Scholar] [CrossRef]
- Jiujiang, Y.; Cleveland, T.E.; Nierman, W.C.; Bennett, J.W. Aspergillus flavus genomics: Gateway to human and animal health, food safety, and crop resistance to diseases. Rev. Iberoam. Micol. 2005, 22, 194–202. [Google Scholar]
- Kokalisburelle, N. Compendium of peanut diseases. J. Periodontol. 1997, 75, 1196–1202. [Google Scholar]
- Georgianna, D.R.; Fedorova, N.D.; Burroughs, J.L.; Dolezal, A.L.; Bok, J.W.; Horowitzbrown, S.; Woloshuk, C.P.; Yu, J.J.; Keller, N.P.; Payne, G.A. Beyond aflatoxin: Four distinct expression patterns and functional roles associated with Aspergillus flavus secondary metabolism gene clusters. Mol. Plant Pathol. 2010, 11, 213–226. [Google Scholar] [CrossRef]
- Shuguang, Z.; Monahan, B.J.; Tkacz, J.S.; Barry, S. Indole-diterpene gene cluster from Aspergillus flavus. Appl. Environ. Microbiol. 2004, 70, 6875–6883. [Google Scholar]
- Trucksess, M.W.; Dombrink-Kurtzman, M.A.; Tournas, V.H.; White, K.D. Occurrence of aflatoxins and fumonisins in incaparina from Guatemala. Food Addit. Contam. 2002, 19, 671–675. [Google Scholar] [CrossRef]
- Gustin, M.C.; Albertyn, J.; Alexander, M.; Davenport, K. MAP kinase pathways in the yeast saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 1998, 62, 1264–1300. [Google Scholar]
- Kentaro, F.; Yukiko, H.; Tatsuya, M.; Tasuku, N.; Keietsu, A. Aspergillus nidulans HOG pathway is activated only by two-component signalling pathway in response to osmotic stress. Mol. Microbiol. 2010, 56, 1246–1261. [Google Scholar]
- Raitt, D.C.; Posas, F.; Saito, H. Yeast Cdc42 GTPase and Ste20 PAK-like kinase regulate Sho1-dependent activation of the Hog1 MAPK pathway. EMBO J. 2000, 19, 4623. [Google Scholar] [CrossRef] [PubMed]
- Herskowitz, I. MAP kinase pathways in yeast: For mating and more. Cell 1995, 80, 187–197. [Google Scholar] [CrossRef] [Green Version]
- Levin, D.E.; Errede, B. The proliferation of MAP kinase signaling pathways in yeast. Curr. Opin. Cell Biol. 1995, 7, 197–202. [Google Scholar] [CrossRef]
- Brewster, J.L.; de Valoir, T.; Dwyer, N.D.; Winter, E.; Gustin, M.C. An osmosensing signal transduction pathway in yeast. Science 1993, 259, 1760–1763. [Google Scholar] [CrossRef]
- Posas, F.; Takekawa, M.; Saito, H. Signal transduction by MAP kinase cascades in budding yeast. Curr. Opin. Microbiol. 1998, 1, 175–182. [Google Scholar] [CrossRef]
- Kosti, I.; Mandel-Gutfreund, Y.; Glaser, F.; Horwitz, B.A. Comparative analysis of fungal protein kinases and associated domains. BMC Genom. 2010, 11, 133. [Google Scholar] [CrossRef] [Green Version]
- Bartholomew, C.R.; Hardy, C.F. P21-activated kinases Cla4 and Ste20 regulate vacuole inheritance in Saccharomyces cerevisiae. Eukaryot. Cell 2009, 8, 560–572. [Google Scholar] [CrossRef] [Green Version]
- Josée, A.; Cunle, W.; Robert, L.; Maleek, J.; Willem, S.; Mike, O.; Thomas, D.Y.; Malcolm, W. Genetic analysis of the interface between Cdc42p and the crib domain of Ste20p in Saccharomyces cerevisiae. Genetics 2003, 163, 9. [Google Scholar]
- Boyce, K.J.; Andrianopoulos, A. Ste20-related kinases: Effectors of signaling and morphogenesis in fungi. Trends Microbiol. 2011, 19, 400–410. [Google Scholar] [CrossRef]
- Johnson, G.L.; Razvan, L. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, O.; Lee, B.R.; Gomi, K.; Iimura, Y. Cloning and functional analysis of the Aspergillus oryzae conidiation regulator gene brlA by its disruption and misscheduled expression. J. Biosci. Bioeng. 1999, 87, 424–429. [Google Scholar] [CrossRef]
- Park, H.S.; Yu, J.H. Genetic control of asexual sporulation in filamentous fungi. Curr. Opin. Microbiol. 2012, 15, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Horn, B.W.; Sorensen, R.B.; Lamb, M.C.; Sobolev, V.S.; Olarte, R.A.; Worthington, C.J.; Carbone, I. Sexual reproduction in Aspergillus flavus sclerotia naturally produced in corn. Phytopathology 2014, 104, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, F.J.; Takahashi, T.; Matsushima, K.; Hara, S.; Shinohara, Y.; Maruyama, J.; Kitamoto, K.; Koyama, Y. ScIR, a basic helix-loop-helix transcription factor, regulates hyphal morphology and promotes sclerotial formation in Aspergillus oryzae. Eukaryot. Cell 2011, 10, 945–955. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Guo, Z.; Zhong, H.; Wang, S.; Yang, W.; Liu, Y.; Wang, S. RNA-seq-based transcriptome analysis of aflatoxigenic Aspergillus flavus in response to water activity. Toxins 2014, 6, 3187–3207. [Google Scholar] [CrossRef] [Green Version]
- Peter, M.; Neiman, A.M.; Park, H.O.; Van Lohuizen, M.; Herskowitz, I. Functional analysis of the interaction between the small GTP binding protein Cdc42 and the Ste20 protein kinase in yeast. EMBO J. 1996, 15, 7046. [Google Scholar] [CrossRef]
- Lamson, R.E.; Winters, M.J.; Pryciak, P.M. Cdc42 regulation of kinase activity and signaling by the yeast p21-activated kinase Ste20. Mol. Cell. Biol. 2002, 22, 2939–2951. [Google Scholar] [CrossRef] [Green Version]
- Leeuw, T.; Fourest-Lieuvin, A.; Wu, C.; Chenevert, J.; Clark, K.; Whiteway, M.; Thomas, D.Y.; Leberer, E. Pheromone response in yeast: Association of Bem1p with proteins of the MAP kinase cascade and actin. Science 1995, 270, 1210–1213. [Google Scholar] [CrossRef]
- Smith, M.E.; Henkel, T.W.; Rollins, J.A. How many fungi make sclerotia? Fungal Ecol. 2015, 13, 211–220. [Google Scholar] [CrossRef]
- Valiante, V.; Macheleidt, J.; Föge, M.; Brakhage, A.A. The Aspergillus fumigatus cell wall integrity signaling pathway: Drug target, compensatory pathways, and virulence. Front. Microbiol. 2015, 6, 325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Q.; Li, C.; Li, Y.; Shang, J.; Li, D.; Chen, B.; Dong, H. Complexity of roles and regulation of the PMK1-MAPK pathway in mycelium development, conidiation and appressorium formation in Magnaporthe oryzae. Gene Expr. Patterns 2013, 13, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Heller, J.; Ruhnke, N.; Espino, J.J.; Massaroli, M.; Collado, I.G.; Tudzynski, P. The mitogen-activated protein kinase BcSak1 of botrytis cinerea is required for pathogenic development and has broad regulatory functions beyond stress response. Mol. Plant-Microbe Interact. MPMI 2012, 25, 802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, D.; Zhang, S.; Zhou, X.; Wang, C.; Xiang, P.; Zheng, Q.; Xu, J.R. The FgHOG1 pathway regulates hyphal growth, stress responses, and plant infection in Fusarium graminearum. PLoS ONE 2012, 7, e49495. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.E.; Thorner, J. Function and regulation in MAPK signaling pathways: Lessons learned from the yeast Saccharomyces cerevisiae. Biochim. Biophys. Acta 2007, 1773, 1311–1340. [Google Scholar] [CrossRef] [Green Version]
- Ana-Maria, C.; Elaine, B.; Rogers, T.R.; Mariana, C.; Mühlschlegel, F.A.; Ken, H. Candida glabrata Ste20 is involved in maintaining cell wall integrity and adaptation to hypertonic stress, and is required for wild-type levels of virulence. Yeast 2010, 21, 557–568. [Google Scholar]
- Lee, Y.M.; Kim, E.; An, J.; Lee, Y.; Choi, E.; Choi, W.; Moon, E.; Kim, W. Dissection of the HOG pathway activated by hydrogen peroxide in Saccharomyces cerevisiae. Environ. Microbiol. 2017, 19, 584–597. [Google Scholar] [CrossRef]
- Wu, C.; Lee, S.F.; Furmaniak-Kazmierczak, E.; Côté, G.P.; Thomas, D.Y.; Leberer, E. Activation of myosin-I by members of the Ste20p protein kinase family. J. Biol. Chem. 1996, 271, 31787. [Google Scholar] [CrossRef] [Green Version]
- Leberer, E.; Ziegelbauer, K.; Schmidt, A.; Harcus, D.; Dignard, D.; Ash, J.; Johnson, L.; Thomas, D.Y. Virulence and hyphal formation of Candida albicans require the Ste20p-like protein kinase CaCla4p. Curr. Biol. 1997, 7, 539–546. [Google Scholar] [CrossRef] [Green Version]
- Roze, L.V.; Laivenieks, M.; Hong, S.Y.; Wee, J.; Wong, S.S.; Vanos, B.; Awad, D.; Ehrlich, K.C.; Linz, J.E. Aflatoxin biosynthesis is a novel source of reactive oxygen species—A potential redox signal to initiate resistance to oxidative stress? Toxins 2015, 7, 1411–1430. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Geng, L.; Huang, L.; Deng, J.; Fasoyin, O.E.; Yao, G.; Wang, S. Contribution of peroxisomal protein importer AflPex5 to development and pathogenesis in the fungus Aspergillus flavus. Curr. Genet. 2018, 64, 1335–1348. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Yang, G.; Zhang, D.; Liu, Y.; Yu, L.; Lin, G.; Guo, Z.; Wang, S.; Zhuang, Z. The PHD transcription factor Rum1 regulates morphogenesis and aflatoxin biosynthesis in Aspergillus flavus. Toxins 2018, 10, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.; Liang, L.; Ran, F.; Liu, Y.; Li, Z.; Lan, H.; Gao, P.; Zhuang, Z.; Zhang, F.; Nie, X. The Dmta methyl transferase contributes to Aspergillus flavus conidiation, sclerotial production, aflatoxin biosynthesis and virulence. Sci. Rep. 2016, 6, 23259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.; Liu, Y.; Liang, L.; Li, Z.; Qin, Q.; Nie, X.; Wang, S. The high-affinity phosphodiesterase PdeH regulates development and aflatoxin biosynthesis in Aspergillus flavus. Fungal Genet. Biol. 2017, 101, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Yu, S.; Qiu, M.; Wang, X.; Wang, Y.; Bai, Y.; Zhang, F.; Wang, S. Aspergillus flavus sumo contributes to fungal virulence and toxin attributes. J. Agric. Food Chem. 2016, 64, 6772. [Google Scholar] [CrossRef]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative pcr and the 2−ΔΔCT method. Methods 2000, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Genotype Description | Source |
|---|---|---|
| A. flavus CA14 | ∆ku70, ∆pyrG | Purchased from FGSC |
| wild-type (WT) | ∆ku70, ∆pyrG::AfpyrG | This study |
| ∆Aflste20 | ∆ku70, ∆pyrG::AfpyrG, ∆Aflste20 | This study |
| ∆Aflste20C | ∆ku70, ∆Aflste20::Aflste20, ∆pyrG::AfpyrG | This study |
| Primer Name | Sequence (5’–3’) | Amplified Fragment |
|---|---|---|
| ste20-p1 | TGCTTAGAGGATGGGATT | 5’UTR of Aflste20 |
| ste20-p3 | GGGTGAAGAGCATTGTTTGAGGCCGCTAGGCTCAGTGATGG | |
| ste20-p6 | GCATCAGTGCCTCCTCTCAGACCCAGATACCCATTGCTCC | 3’UTR of Aflste20 |
| ste20-p8 | CACCAGCCCACATAGAAT | |
| ste20-p4 | GCCTCAAACAATGCTCTTCACCC | A. fumigatus pyrG |
| ste20-p5 | GTCTGAGAGGAGGCACTGATGC | |
| ste20-p2 | TTTGGCACTCGCTTGTCC | Fusion PCR |
| ste20-p7 | CCGCTCAAGTCTGGGTTA | |
| ste20-p9 | TCGGTTAATCACATCTGTCTC | ORF validates primers |
| ste20-p10 | CATCAATCATCGCCATCTA | |
| pyrG-801 | CAGGAGTTCTCGGGTTGTCG | AP BP |
| pyrG-1020 | CAGAGTATGCGGCAAGTCA | |
| ste20-C-p1 | GCTTCTGGTGGCGTATT | 5’UTR of ∆Aflste20C |
| ste20-C-p2 | GGGTGAAGAGCATTGTTTGAGGCCGCTCAAGTCTGGGTTAT | |
| ste20-C-p3 | GCATCAGTGCCTCCTCTCAGACCTGATGTCTCGCCTGTTA | 3’UTR of ∆Aflste20C |
| ste20-C-p4 | CTCCGCCGCAACTTTAT | |
| ste20-C-p5 | CTCTGGGTGGTGATGGA | Fusion PCR |
| ste20-C-p6 | AGAATACGAGGCTTGTGG |
| Gene | Forward Sequences (5’–3’) | Reverse Sequences (5’–3’) |
|---|---|---|
| Aflste20 | ATCAACGACTCGCACAATAA | GCTCGCCCTCAATCATCT |
| brlA | GCCTCCAGCGTCAACCTTC | TCTCTTCAAATGCTCTTGCCTC |
| abaA | TCTTCGGTTGATGGATGATTTC | CCGTTGGGAGGCTGGGT |
| nsdC | GCCAGACTTGCCAATCAC | CATCCACCTTGCCCTTTA |
| sclR | CAATGAGCCTATGGGAGTGG | ATCTTCGCCCGAGTGGTT |
| aflR | AAAGCACCCTGTCTTCCCTAAC | GAAGAGGTGGGTCAGTGTTTGTAG |
| aflS | CGAGTCGCTCAGGCGCTCAA | GCTCAGACTGACCGCCGCTC |
| aflD | GTGGTGGTTGCCAATGCG | CTGAAACAGTAGGACGGGAGC |
| aflK | GAGCGACAGGAGTAACCGTAAG | CCGATTCCAGACACCATTAGCA |
| aflQ | GTCGCATATGCCCCGGTCGG | GGCAACCAGTCGGGTTCCGG |
| actin | ACGGTGTCGTCACAAACTGG | CGGTTGGACTTAGGGTTGATAG |
| GRE | GCGTATCGTCGTTACCTCATC | CCTTCTCCTTTACCTCCTCGAT |
| HSP | CCGGCATACTATGTCTCGTCT | TAGGGCCTTCGTCGAACA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Qin, L.; Wang, Y.; Xie, Q.; Li, N.; Wang, S.; Yuan, J. AflSte20 Regulates Morphogenesis, Stress Response, and Aflatoxin Biosynthesis of Aspergillus flavus. Toxins 2019, 11, 730. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11120730
Li D, Qin L, Wang Y, Xie Q, Li N, Wang S, Yuan J. AflSte20 Regulates Morphogenesis, Stress Response, and Aflatoxin Biosynthesis of Aspergillus flavus. Toxins. 2019; 11(12):730. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11120730
Chicago/Turabian StyleLi, Ding, Ling Qin, Yinchun Wang, Qingchen Xie, Na Li, Shihua Wang, and Jun Yuan. 2019. "AflSte20 Regulates Morphogenesis, Stress Response, and Aflatoxin Biosynthesis of Aspergillus flavus" Toxins 11, no. 12: 730. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11120730