Biological System Responses of Dairy Cows to Aflatoxin B1 Exposure Revealed with Metabolomic Changes in Multiple Biofluids

 , ,
, ,
Abstract
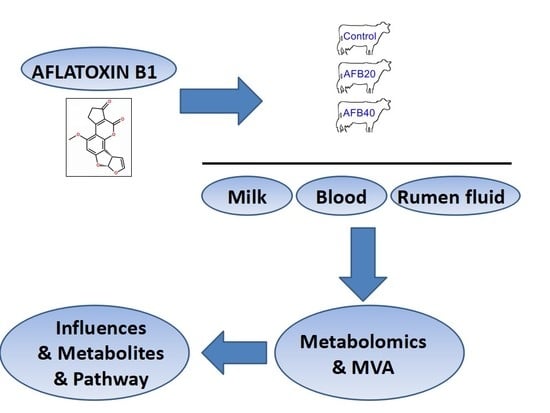
:
1. Introduction
2. Results
2.1. Feed Intake, Milk Yield, and Milk Composition
2.2. Aflatoxin M1 Concentrations in Milk
2.3. Serum Biochemical Parameters
2.4. Rumen Function
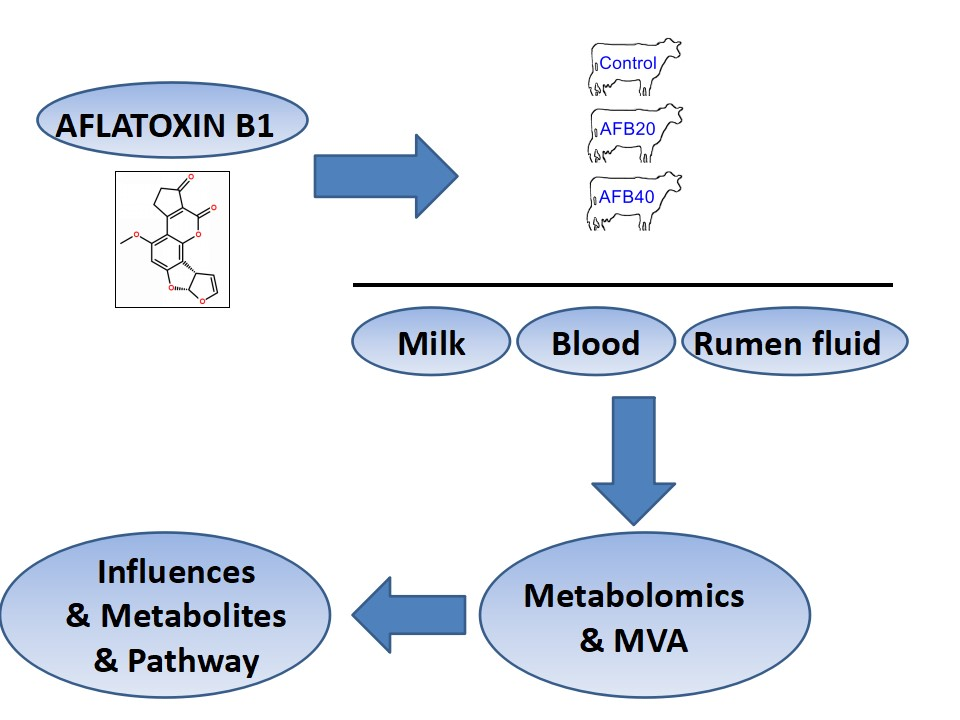
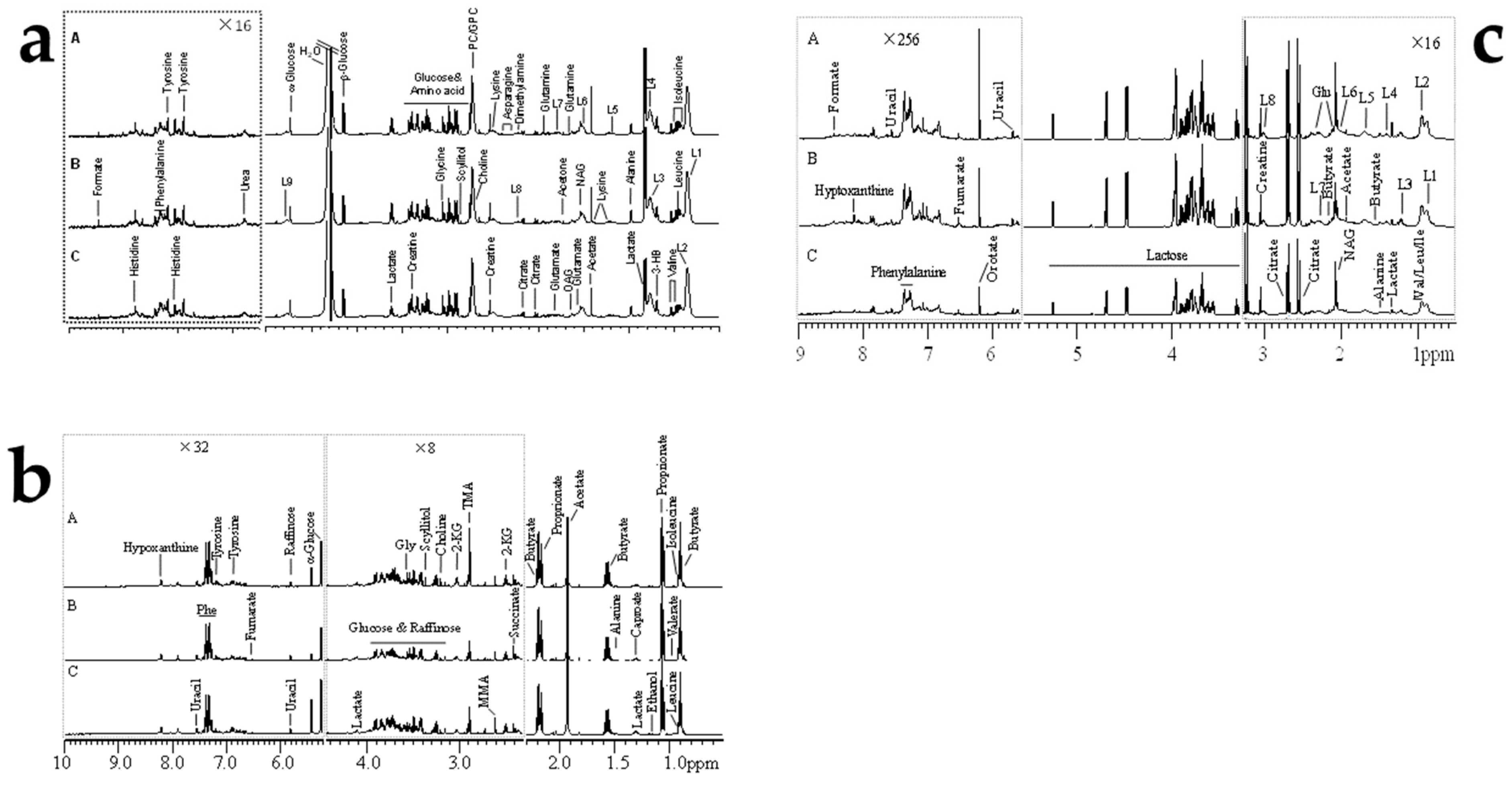
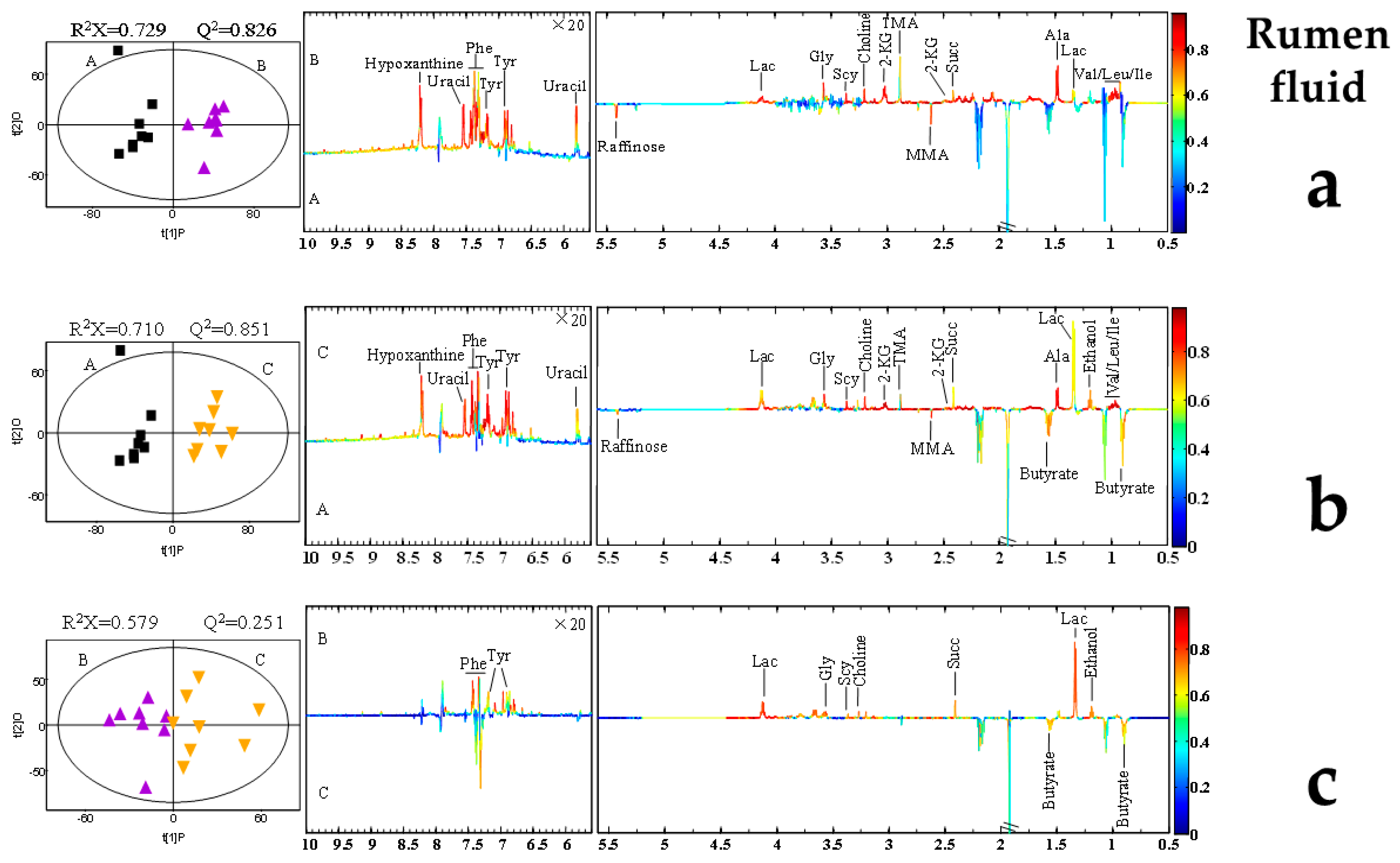
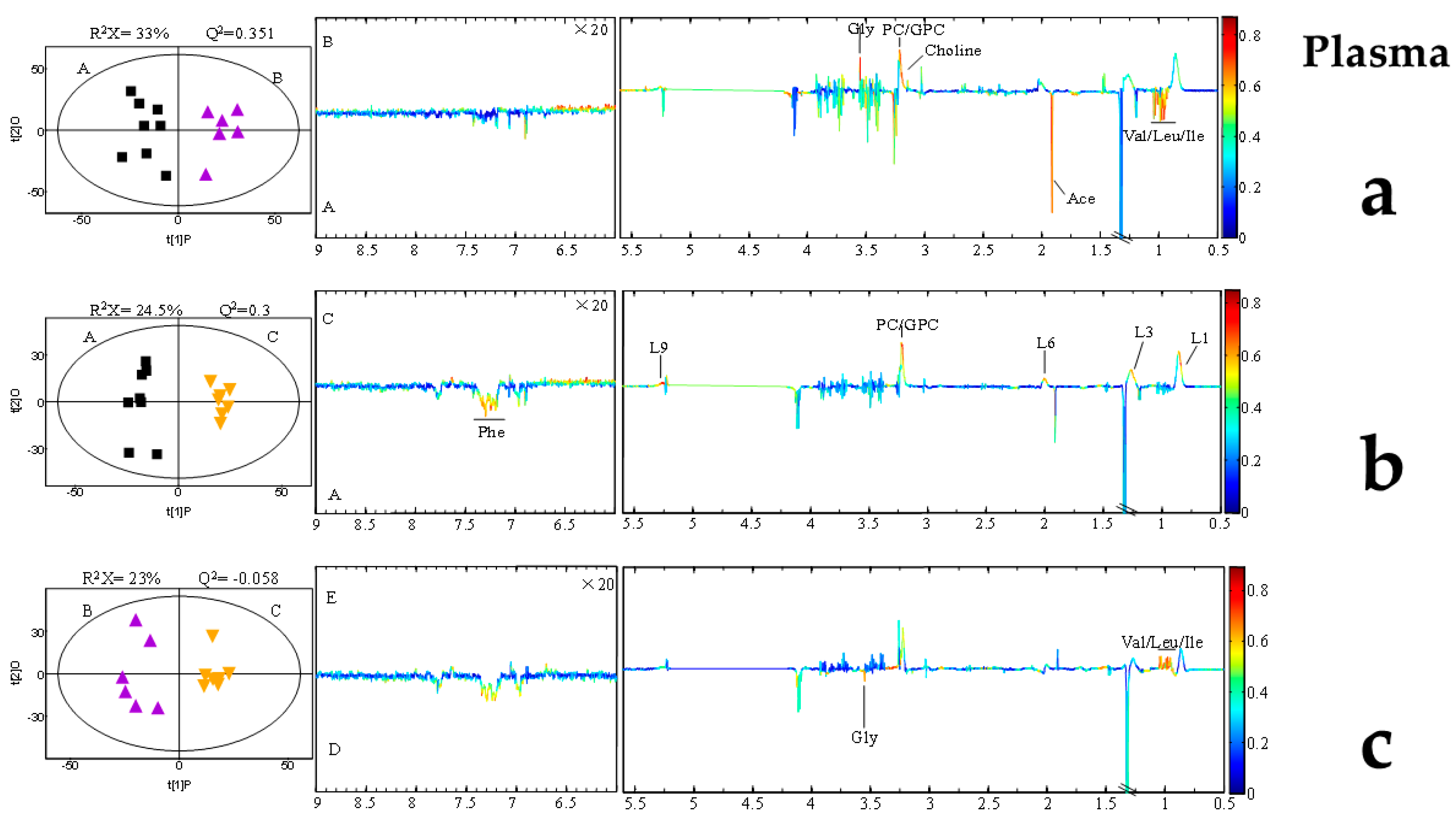
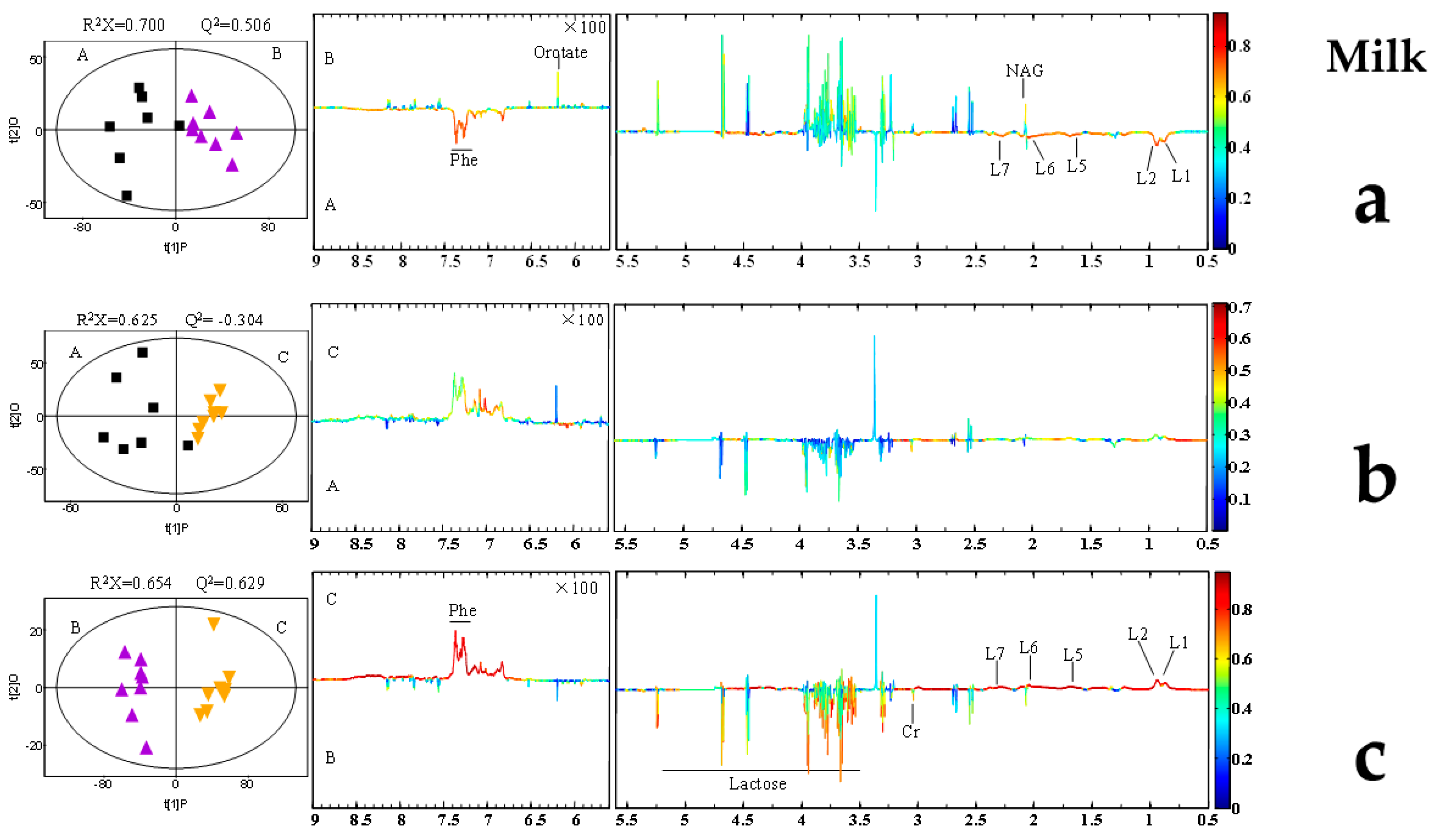
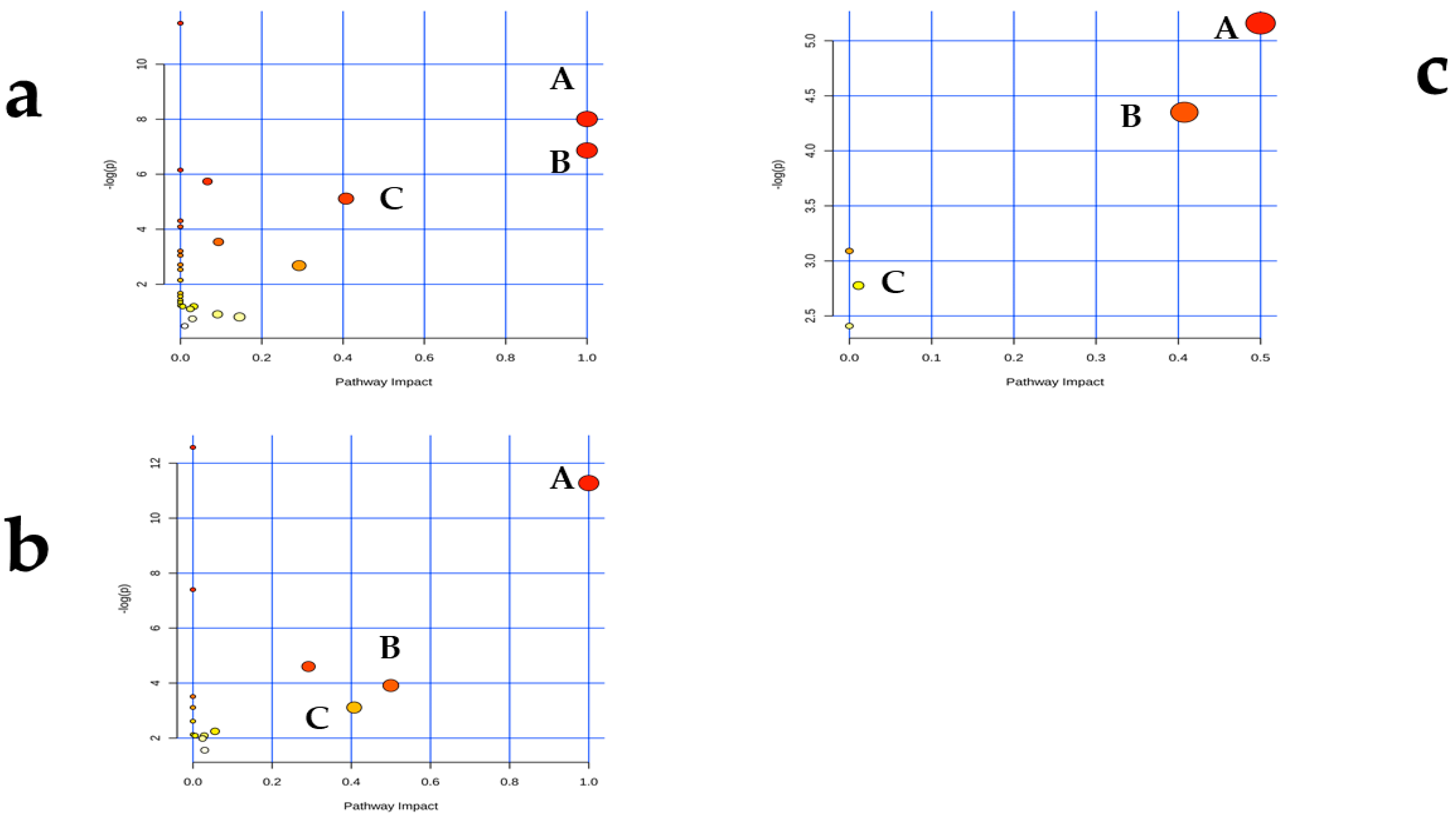
2.5. AFB1-Induced Metabolomic Changes
3. Discussion
3.1. Effect of AFB1 on Milking Performance
3.2. Aflatoxin M1 Concentrations in Milk
3.3. Effects of AFB1 on Serum Biochemical Parameters
3.4. AFB1-Induced Rumen VFAs and NH3-N Changes
3.5. AFB1-Induced Metabolomic Changes
3.5.1. AFB1 Affects Amino Acid Metabolism
3.5.2. AFB1 Affects Lipid and Nucleic Acid Metabolism
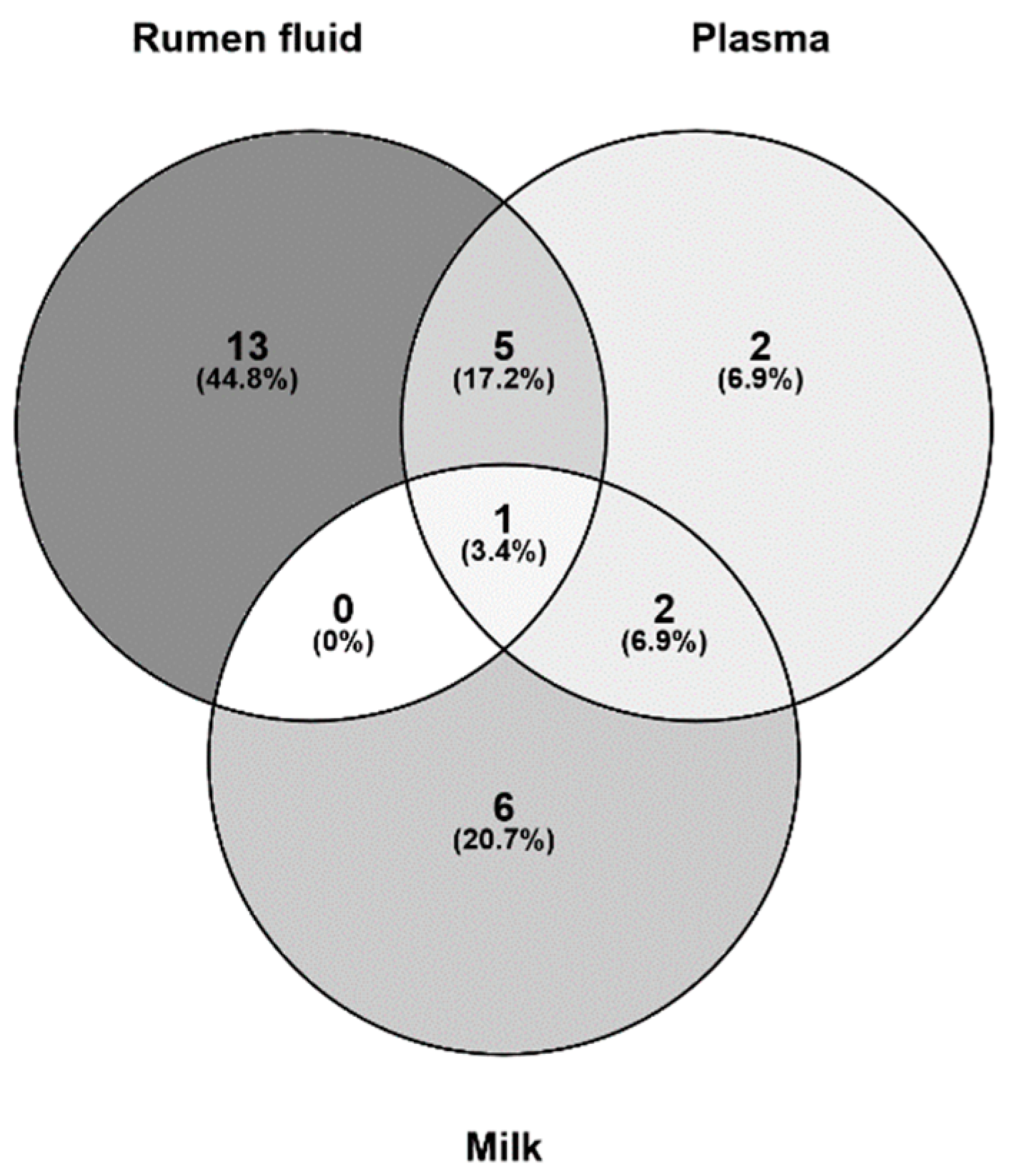
3.6. Putative Analysis of Metabolites from Rumen Fluid, Plasma, and Milk
4. Conclusions
5. Materials and Methods
5.1. Animal Handling and Sample Preparation
5.2. Experimental Methods and Sample Collection
5.3. 1H NMR Spectroscopic Analysis of Milk
5.4. 1H NMR Spectroscopic Analysis of Plasma
5.5. 1H NMR Spectroscopic Analysis of Rumen Fluid
5.6. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Streit, E.; Schatzmayr, G.; Tassis, P.; Tzika, E.; Marin, D.; Taranu, I.; Tabuc, C.; Nicolau, A.; Aprodu, I.; et al. Current Situation of Mycotoxin Contamination and Co-occurrence in Animal Feed—Focus on Europe. Toxins 2012, 4, 788–809. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.R.; Fink-Gremmels, J. Mycotoxin syndrome in dairy cattle: Characterisation and intervention results. World Mycotoxin J. 2014, 7, 357–366. [Google Scholar] [CrossRef]
- Smith, M.C.; Madec, S.; Coton, E.; Hymery, N. Natural co-occurrence of mycotoxins in foods and feeds and their in vitro combined toxicological effects. Toxins 2016, 8, 94. [Google Scholar] [CrossRef]
- Sargeant, K.; Sheridan, A.; O’Kelly, J.; Carnaghan, R.B.A. Toxicity associated with certain samples of groundnuts. Nature 1961, 192, 1096–1097. [Google Scholar] [CrossRef]
- Surai, P.; Mezes, M.; Fotina, T.I.; Denev, S.D. Mycotoxins in Human Diet: A Hidden Danger. In Modern Dietary Fat Intakes in Disease Promotion; De Meester, F., Zibadi, S., Watson, R., Eds.; Humana Press: Totowa, NJ, USA, 2010. [Google Scholar]
- Zhang, L.; Ye, Y.; An, Y.; Tian, Y.; Wang, Y.; Tang, H. Systems responses of rats to aflatoxin B1 exposure revealed with metabonomic changes in multiple biological matrices. J. Proteome Res. 2011, 10, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Zheng, N.; Fan, C.; Cheng, M.; Wang, S.; Jabar, A.; Wang, J.Q.; Cheng, J.B. Effects of aflatoxin B1 combined with ochratoxin A and/or zearalenone on metabolism, immune function, and antioxidant status in lactating dairy goats. Asian-Australas J. Anim. Sci. 2018, 31, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.L.; Wang, Y.M.; Nennich, T.D.; Li, Y.; Liu, J.X. Transfer of dietary aflatoxin B1, to milk aflatoxin M1, and effect of inclusion of adsorbent in the diet of dairy cows. J. Dairy Sci. 2015, 98, 2545–2554. [Google Scholar] [CrossRef]
- Battacone, G.; Nudda, A.; Cannas, A.; Cappio, B.A.; Bomboi, G.; Pulina, G. Excretion of aflatoxin M1 in milk of dairy ewes treated with different doses of aflatoxin B1. J. Dairy Sci. 2003, 86, 2667–2675. [Google Scholar] [CrossRef]
- Battacone, G.; Nudda, A.; Palomba, M.; Pascale, M.; Nicolussi, P.; Pulina, G. Transfer of aflatoxin B1 from feed to milk and from milk to curd and whey in dairy sheep fed artificially contaminated concentrates. J. Dairy Sci. 2005, 88, 3063–3069. [Google Scholar] [CrossRef]
- Fink-Gremmels, J. Mycotoxins in cattle feeds and carry-over to dairy milk: A review. Food Addit. Contam. 2008, 25, 172–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovati, L.; Magliani, W.; Ciociola, T.; Santinoli, C.; Conti, S.; Polonelli, L. AFM1 in milk: Physical, biological, and prophylactic methods to mitigate contamination. Toxins 2015, 7, 4330–4349. [Google Scholar] [CrossRef] [PubMed]
- Battacone, G.; Nudda, A.; Palomba, M.; Mazzette, A.; Pulina, G. The transfer of aflatoxin M1 in milk of ewes fed diet naturally contaminated by aflatoxins and effect of inclusion of dried yeast culture in the diet. J. Dairy Sci. 2009, 92, 4997–5004. [Google Scholar] [CrossRef] [PubMed]
- Dänicke, S.; Matthaus, K.; Lebzien, P.; Valenta, H.; Stemme, K.; Ueberschar, K.H.; Razzazi-Fazeli, E.; Bohm, J.; Flachowsky, G. Effects of Fusarium toxin contaminated wheat grain on nutrient turnover, microbial protein synthesis and metabolism of deoxynivalenol and zearalenone in the rumen of dairy cows. J. Anim. Physiol. Anim. Nutr. 2005, 89, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.H.; Xu, Z.R.; Feng, J.L.; Wang, C.Z. Efficacy of modified montmorillonite nanocomposite to reduce the toxicity of aflatoxin in broiler chicks. Anim. Feed Sci. Technol. 2006, 129, 138–148. [Google Scholar] [CrossRef]
- Cheng, J.; Huang, S.; Fan, C.; Zheng, N.; Zhang, Y.; Li, S.; Wang, J. Metabolomic analysis of alterations in lipid oxidation, carbohydrate and amino acid metabolism in dairy goats caused by exposure to Aflotoxin B1. J. Dairy Res. 2017, 84, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.K.; Wilson, I.D. Understanding ‘global’ systems biology: Metabonomics and the continuum of metabolism. Nat. Rev. Drug Discov. 2003, 2, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.S.; Buttchereit, N.; Miemczyk, S.P.; Immervoll, A.K.; Louis, C.; Wiedemann, S.; Junge, W.; Thaller, G.; Oefner, P.J.; Gronwald, W. NMR metabolomic analysis of dairy cows reveals milk glycerophosphocholine to phosphocholine ratio as prognostic biomarker for risk of ketosis. J. Proteome Res. 2012, 11, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Wang, W.Y.; Zheng, N.; Cheng, J.B.; Li, S.L.; Zhang, Y.D.; Wang, J.Q. Identification of diagnostic biomarkers and metabolic pathway shifts ofheat-stressed lactating dairy cows. J. Proteomics 2015, 125, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Toda, K.; Kokushi, E.; Uno, S.; Shiiba, A.; Hasunuma, H.; Fushimi, Y.; Wijayagunawardane, M.P.B.; Zhang, C.; Yamato, O.; Taniguchi, M.; et al. Gas chromatography-mass spectrometry for metabolite profiling of Japanese black cattle naturally contaminated with Zearalenone and Sterigmatocystin. Toxins 2017, 9, E294. [Google Scholar] [CrossRef] [PubMed]
- Kiessling, K.H. Biochemical mechanism of action of mycotoxins. Pure Appl. Chem. 1986, 58, 327–338. [Google Scholar] [CrossRef]
- Mishra, H.N.; Das, C. A review on biological control and metabolism of aflatoxin. Crit. Rev. Food Sci. 2003, 43, 245–264. [Google Scholar] [CrossRef] [PubMed]
- Mcallan, A.B.; Smith, R.H. Degradation of nucleic acids in the rumen. Br. J. Nutr. 1973, 29, 331–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, D.P.H. Potential human health hazards of mycotoxins. In Mycotoxins and Phycotoxins; Natori, S., Hashimoto, H., Ueno, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1989; pp. 69–80. [Google Scholar]
- De Pascali, S.A.; Gambacorta, L.; Oswald, I.P.; Del Coco, L.; Solfrizzo, M.; Fanizzi, F.P. 1H NMR and MVA metabolomic profiles of urines from piglets fed with boluses contaminated with a mixture of five mycotoxins. Biochem. Biophys. Rep. 2017, 11, 9–18. [Google Scholar] [PubMed]
- Xia, K.; He, X.; Dai, Q.; Cheng, W.H.; Qi, X.; Guo, M.; Luo, Y.; Huang, K.; Zhao, C.; Xu, W. Discovery of systematic responses and potential biomarkers induced by ochratoxin A using metabolomics. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2014, 31, 1904–1913. [Google Scholar] [CrossRef] [PubMed]
- Hopton, R.P.; Turner, E.; Burley, V.J.; Turner, P.C.; Fisher, J. Urine metabolite analysis as a function of deoxynivalenol exposure: An NMR-based metabolomics investigation. Food Addit. Contam. 2010, 27, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Waters, N.J.; Holmes, E.; Williams, A.; Waterfield, C.J.; Farrant, R.D.; Nicholson, J.K. NMR and pattern recognition studies on the time-related metabolic effects of alpha-naphthylisothiocyanate on liver, urine, and plasma in the rat: An integrative metabonomic approach. Chem. Res. Toxicol. 2001, 14, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- General Administration of Quality Supervision, Inspection and Quarantine tPsRoC (GAQSIQ). GB/T 13078-2017, Hygienical Standard for Feeds; Standards Press of China: Beijing, China, 2017.
- Hall, M.B.; Nennich, T.D.; Doane, P.H.; Brink, G.E. Total volatile fatty acid concentrations are unreliable estimators of treatment effects on ruminal fermentation in vivo. J. Dairy Sci. 2015, 98, 3988–3999. [Google Scholar]
- Korosteleva, S.N.; Smith, T.K.; Boermans, H.J. Effects of feedborne Fusarium mycotoxins on the performance, metabolism, and immunity of dairy cows. J. Dairy Sci. 2007, 90, 3867–3873. [Google Scholar] [CrossRef]
- Krishna, D.R.; Klotz, U. Extrahepatic metabolism of drugs in humans. Clin. Pharmacokinet. 1994, 26, 144–160. [Google Scholar] [CrossRef]
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Queiroz, O.C.M.; Han, J.H.; Staples, C.R.; Adesogan, A.T. Effect of adding a mycotoxin-sequestering agent on milk aflatoxin M1 concentration and the performance and immune response of dairy cattle fed an aflatoxin B1-contaminated diet. J. Dairy Sci. 2012, 95, 5901–5908. [Google Scholar] [CrossRef] [PubMed]
- Milićević, D.; Nikšić, M.; Baltić, T.; Vranić, D. Isolation, characterization and evaluation of significant mycoflora and mycotoxins in pig feed from serbian farms. World J. Microbiol. Biotechnol. 2010, 26, 1715–1720. [Google Scholar] [CrossRef]
- Choudhary, P.L.; Sharma, R.S.; Borkhataria, V.N.; Desai, M.C. Effect of feeding aflatoxin B1 on feed consumption through naturally contaminated feeds. Indian J. Anim. Sci. 1998, 68, 400–401. [Google Scholar]
- Wu, G. Functional amino acids in nutrition and health. Amino Acids 2013, 45, 407–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, D.E.; Hagler, W.M.; Blackwelder, J.T.; Eve, J.A.; Hopkins, B.A.; Anderson, K.L.; Jones, F.T.; Whitlow, L.W. Aflatoxin binders II: Reduction of aflatoxin M1 in milk by sequestering agents of cows consuming aflatoxin in feed. Mycopathologia 2004, 157, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Masoero, F.; Gallo, A.; Moschini, M.; Piva, G.; Diaz, D. Carryover of aflatoxin from feed to milk in dairy cows with low or high somatic cell counts. Animal 2007, 1, 1344–1350. [Google Scholar] [CrossRef] [PubMed]
- Kuilman, M.E.M.; Maas, R.F.M.; Fink-Gremmels, J. Cytochrome P450-mediated metabolism and cytotoxicity of aflatoxin B1 in bovine hepatocytes. Toxicol. In Vitro 2000, 14, 321–327. [Google Scholar] [CrossRef]
- Battacone, G.; Nudda, A.; Rassu, S.P.; Decandia, M.; Pulina, G. Excretion pattern of aflatoxin M1 in milk of goats fed a single dose of aflatoxin B1. J. Dairy Sci. 2012, 95, 2656–2661. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration (FDA). Guidance for Industry: Action Levels for Poisonous or Deleterious Substances in Human Food and Animal Feed. Available online: http://www.fda.gov/Food/GuidanceComplianceRegulatoryInformation/GuidanceDocuments/ChemicalContaminantsandPesticides/ucm077969.htm (accessed on 20 May 2011).
- European Food Safety Authority (EFSA). Opinion of the scientific panel on contaminants in the food chain on a request from the Commission related to aflatoxin B1 as undesirable substance in animal feed. EFSA J. 2004, 39, 1–27. [Google Scholar]
- Murphy, K.; Travers, P.; Walport, M. Janeway’s Immunology, 7th rev. ed.; Garland Sci.: Abingdon, UK, 2008. [Google Scholar]
- Hiss, S.; Mielenz, M.; Bruckmaier, R.M.; Sauerwein, H. Haptoglobin concentrations in blood and milk after endotoxin challenge and quantification of mammary Hp mRNA expression. J. Dairy Sci. 2004, 87, 3778–3784. [Google Scholar] [CrossRef]
- Godson, D.L.; Campos, M.; Attah-Poku, S.K.; Redmond, M.J.; Cordeiro, D.M.; Sethi, M.S.; Harland, R.J.; Babiuk, L.A. Serum haptoglobin as an indicator of the acute phase response in bovine respiratory disease. Vet. Immunol. Immunop. 1996, 51, 277–292. [Google Scholar] [CrossRef]
- Spooner, R.L.; Miller, J.K. The measurement of haemoglobin reactive protein in ruminants as an aid to the diagnosis of acute inflammation. Vet. Rec. 1971, 88, 2–4. [Google Scholar] [CrossRef] [PubMed]
- Reverberi, M.; Zjalic, S.; Ricelli, A.; Fabbri, A.A.; Fanelli, C. Oxidant/antioxidant balance in Aspergillus parasiticus affects aflatoxin biosynthesis. Mycotoxin Res. 2006, 22, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Soyöz, M.; Özçelik, N.; Kihnç, I.; Altuntaş, I. The effects of ochratoxin A on lipid peroxidation and antioxidant enzymes: A protective role of melatonin. Cell Biol. Toxicol. 2004, 20, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.P. Cellular defenses against damage from reactive oxygen species. Physiol. Rev. 1994, 74, 139–162. [Google Scholar] [CrossRef]
- Armstrong, D.; Browne, R. The analysis of free radicals, lipid peroxides, antioxidant enzymes and compounds related to oxidative stress as applied to the clinical chemistry laboratory. Adv. Exp. Med. Biol. 1994, 366, 43–58. [Google Scholar]
- Wan, Q.F.; He, Q.H.; Deng, X.B.; Hao, F.H.; Tang, H.R.; Wang, Y.L. Systemic metabolic responses of broiler chickens and piglets to acute T-2 toxin intravenous exposure. J. Agric. Food Chem. 2016, 64, 714–723. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, W. Aflatoxin B1 impairs mitochondrial functions, activates ROS generation, induces apoptosis and involves Nrf2 signal pathway in primary broiler hepatocytes. Anim. Sci. J. 2016, 87, 1490–1500. [Google Scholar] [CrossRef]
- Yarru, L.P.; Settivari, R.S.; Antoniou, E.; Ledoux, D.R.; Rottinghaus, G.E. Toxicological and gene expression analysis of the impact of aflatoxin B1 on hepatic function of male broiler chicks. Poult. Sci. 2009, 88, 360–371. [Google Scholar] [Green Version]
- Gao, H.N.; Hu, H.; Zheng, N.; Wang, J.Q. Leucine and histidine independently regulate milk protein synthesis in bovine mammary epithelial cells via mTOR signaling pathway. J. Zhejiang Univ. Sci. B 2015, 16, 560–572. [Google Scholar] [CrossRef]
- Luo, C.; Zhao, S.; Zhang, M.; Gao, Y.; Wang, J.; Hanigan, M.D.; Zheng, N. SESN2 negatively regulates cell proliferation and casein synthesis by inhibition the amino acid-mediated mTORC1 pathway in cow mammary epithelial cells. Sci. Rep. 2018, 8, 3912. [Google Scholar] [CrossRef] [PubMed]
- Allison, M.J. Phenylalanine biosynthesis from phenylacetic acid by anaerobic bacteria from the rumen. Biochem. Biophys. Res. Commun. 1965, 18, 30–35. [Google Scholar] [CrossRef]
- Khan, R.I.; Onodera, R.; Amin, M.R.; Mohammed, N. Production of tyrosine and other aromatic compounds from phenylalanine by rumen microorganisms. Amino Acids 1999, 17, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Liou, A.P.; Lu, X.; Zhao, X.; Feng, J.; Pechhold, S.B.; Raybould, H.; Wank, S.A. The extracellular calcium sensing receptor is required for cholecystokinin secretion in response to L-phenylalanine in acutely isolated murine intestinal I cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.P.; Xu, M.; Yao, J.H.; Liu, K.; Li, F.; Liu, Y.; Wang, F.; Sun, F.F.; Liu, N.N. Regulation of pancreatic exocrine secretion in goats: Differential effects of short- and long-term duodenal phenylalanine treatment. J. Anim. Physiol. Anim. Nutr. 2013, 97, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.C.; Yang, X.J.; Guo, L.; Zheng, C.; Wang, D.D.; Cai, C.J.; Liu, S.M.; Yao, J.H. Effects of dietary leucine and phenylalanine on pancreas development, enzyme activity, and relative gene expression in milk-fed holstein dairy calves. J. Dairy Sci. 2018, 101, 4235–4244. [Google Scholar] [CrossRef] [PubMed]
- Houweling, M.; Drift, S.G.A.V.D.; Jorritsma, R.; Tielens, A.G.M. Technical note: Quantification of plasma 1- and 3-methylhistidine in dairy cows by high-performance liquid chromatography–tandem mass spectrometry. J. Dairy Sci. 2012, 95, 3125–3130. [Google Scholar] [CrossRef] [Green Version]
- Klepacki, J.; Klawitter, J.; Klawitter, J.; Karimpour-Fard, A.; Thurman, J.; Ingle, G.; Patel, D.; Christians, U. Amino Acids in a Targeted Versus a Non-Targeted Metabolomics LC-MS/MS Assay. Are the Results Consistent? Clin. Biochem. 2016, 49, 955–961. [Google Scholar] [CrossRef]
- Aranibar, N.; Vassallo, J.D.; Rathmacher, J.; Stryker, S.; Zhang, Y.; Dai, J.; Janovitz, E.B.; Robertson, D.; Reily, M.; Lowe-Krentz, L.; et al. Identification of 1- and 3-methylhistidine as biomarkers of skeletal muscle toxicity by nuclear magnetic resonance-based metabolic profiling. Anal. Biochem. 2011, 410, 84–91. [Google Scholar] [CrossRef]
- Izui, K.; Matsumura, H.; Furumoto, T.; Kai, Y. Phosphoenolpyruvate carboxylase: A new era of structural biology. Annu. Rev. Plant Biol. 2004, 55, 69–84. [Google Scholar] [CrossRef]
- Tessari, E.N.; Kobashigawa, E.; Cardoso, A.L.; Ledoux, D.R.; Rottinghaus, G.E.; Oliveira, C.A. Effects of aflatoxin B (1) and fumonisin B(1) on blood biochemical parameters in broilers. Toxins 2010, 2, 453–460. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhang, K.Y.; Chen, D.W.; Ding, X.M.; Feng, G.D.; Ao, X. Effects of maize naturally contaminated with aflatoxin B1, on growth performance, blood profiles and hepatic histopathology in ducks. Livest. Sci. 2013, 152, 192–199. [Google Scholar] [CrossRef]
- Leighton, F.; Bergseth, S.; Rortveit, T.; Christiansen, E.N.; Bremer, J. Free acetate production by rat hepatocytes during peroxisomalfatty acid and dicarboxylic acid oxidation. J. Biol. Chem. 1989, 264, 10347–10350. [Google Scholar] [PubMed]
- Gesing, A.; Karbownik-Lewinska, M. Protective effects of melatoninand N-acetylserotonin on aflatoxin B1-induced lipid peroxidationin rats. Cell Biochem. Funct. 2008, 26, 314–319. [Google Scholar] [CrossRef]
- Manson, M.M.; Hudson, E.A.; Ball, H.W.; Barrett, M.C.; Clark, H.L.; Judah, D.J.; Verschoyle, R.D.; Neal, G.E. Chemoprevention of aflatoxin B1-induced carcinogenesis by indole-3-carbinol in rat liver–predicting the outcome using early biomarkers. Carcinogenesis 1998, 19, 1829–1836. [Google Scholar] [CrossRef]
- Ellinger-Ziegelbauer, H.; Ahr, H.J.; Gmuender, H.; Pallez, C.; Koenig, J. Characteristic expression profiles induced by carcinogens in rat liver. Toxicol. Sci. 2004, 77, 19–34. [Google Scholar] [CrossRef]
- Chen, X.B.; Mejia, A.T.; Kyle, D.J.; Orskov, E.R. Evaluation of the use of the purine derivative: Creatinine ratio in spot urine and plasma samples as an index of microbial protein supply in ruminants: Studies in sheep. J. Agric. Sci. 1995, 125, 137–143. [Google Scholar] [CrossRef]
- Reynal, S.M.; Broderick, G.A.; Bearzi, C. Comparison of four markers for quantifying microbial protein flow from the rumen of lactating dairy cows. J. Dairy Sci. 2005, 88, 4065–4082. [Google Scholar] [CrossRef]
- Saleem, F.; Bouatra, S.; Guo, A.C.; Psychogios, N.; Mandal, R.; Dunn, S.M.; Ametaj, B.N.; Wishart, D.S. The bovine ruminal fluid metabolome. Metabolomics 2013, 9, 360–378. [Google Scholar] [CrossRef]
- Saleem, F.; Ametaj, B.N.; Bouatra, S.; Mandal, R.; Zebeli, Q.; Dunn, S.M.; Wishart, D.S. A metabolomics approach to uncover the effects of grain diets on rumen health in dairy cows. J. Dairy Sci. 2012, 95, 6606–6623. [Google Scholar] [CrossRef]
- Ametaj, B.N.; Zebeli, Q.; Saleem, F.; Psychogios, N.; Lewis, M.J.; Dunn, S.M.; Xia, J.G.; Wishart, D.S. Metabolomics reveals unhealthy alterations in rumen metabolism with increased proportion of cereal grain in the diet of dairy cows. Metabolomics 2010, 6, 583–594. [Google Scholar] [CrossRef]
- Bauman, D.E.; Mather, I.H.; Wall, R.J.; Lock, A.L. Major advances associated with the biosynthesis of milk. J. Dairy Sci. 2006, 89, 1235–1243. [Google Scholar] [CrossRef]
- Wu, G. Dietary requirements of synthesizable amino acids by animals: A paradigm shift in protein nutrition. J. Anim. Sci. Biotechnol. 2015, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Stroup, B.M.; Nair, N.; Murali, S.G.; Broniowska, K.; Rohr, F.; Levy, H.L.; Ney, D.M. Metabolomic markers of essential fatty acids, carnitine, and cholesterol metabolism in adults and adolescents with phenylketonuria. J. Nutr. 2018, 148, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Snigdha, K.; Singh, B.K.; Mehta, A.S.; Tewari, R.P.; Dutta, P.K. Self-assembling N-(9-Fluorenylmethoxycarbonyl)-l-Phenylalanine hydrogel as novel drug carrier. Int. J. Biol. Macromol. 2016, 93, 1639–1646. [Google Scholar] [CrossRef] [PubMed]
- Polekhina, G.; Board, P.G.; Blackburn, A.C.; Parker, M.W. Crystal structure of maleylacetoacetate isomerase/glutathione transferase zeta reveals the molecular basis for its remarkable catalytic promiscuity. Biochemistry 2001, 40, 1567–1576. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Agriculture of China (MOA). Feeding Standard of Dairy Cattle, NY/T 34-2004; MOA: Beijing, China, 2004.
- National Research Council (NRC). Nutrient Requirements of Dairy Cattle; National Academy Press: Washington, DC, USA, 2001. [Google Scholar]
- Shen, J.S.; Chai, Z.; Song, L.J.; Liu, J.X.; Wu, Y.M. Insertion depth of oral stomach tubes may affect the fermentation parameters of ruminal fluid collected in dairy cows. J. Dairy Sci. 2012, 95, 5978–5984. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Control | AFB20 | AFB40 | SEM | p Value |
|---|---|---|---|---|---|
| Daily milk yield (kg/day) | 17.23 | 16.94 | 17.9 | 5.33 | 0.29 |
| Milk fat (%) | 4.93 | 4.50 | 4.69 | 0.11 | 0.32 |
| Milk protein (%) | 4.01 | 4.13 | 4.17 | 0.08 | 0.48 |
| Milk lactose (%) | 5.08 | 5.06 | 5.05 | 0.06 | 0.98 |
| Total solids (%) | 15.17 | 14.73 | 14.97 | 1.17 | 0.58 |
| Urea (mg/dl) | 33.04 | 25.79 | 23.2 | 5.01 | 0.73 |
| SCC (cells/mL) 2 | 0.27 | 0.21 | 0.22 | 0.05 | 0.28 |
| FCM (kg) 2 | 16.19 | 17.03 | 16.54 | 0.58 | 0.59 |
| ECM (kg) 2 | 7.36 | 7.67 | 7.42 | 0.41 | 0.85 |
| Item 2 | Control | AFB20 | AFB40 | SEM | p Value 3 |
|---|---|---|---|---|---|
| ALT (U/L) | 29.75 | 26.00 | 26.38 | 1.02 | 0.26 |
| AST (U/L) | 72.75 | 79.14 | 75.88 | 3.65 | 0.80 |
| GGT (U/L) | 43.09 | 38.06 | 37.84 | 2.39 | 0.61 |
| ALP (U/L) | 90.31 | 96.56 | 68.33 | 15.83 | 0.76 |
| TP (g/L) | 37.53 | 41.64 | 39.30 | 1.15 | 0.54 |
| ALB (g/L) | 36.44 | 35.46 | 36.95 | 0.62 | 0.64 |
| GLOB (g/L) | 3.10 | 3.03 | 3.18 | 1.22 | 0.42 |
| Urea (mmol/mL) | 71.13 | 69.29 | 65.50 | 0.10 | 0.84 |
| CR (μmol/L) | 25.85 | 40.86 | 34.01 | 1.62 | 0.35 |
| UA (μmol/L) | 11.05 | 9.58 | 10.98 | 3.37 | 0.20 |
| TBil (μmol/L) | 2.28 | 2.03 | 2.39 | 0.57 | 0.53 |
| DBil (μmol/L) | 8.78 | 7.54 | 8.59 | 0.09 | 0.32 |
| IBiL (μmol/L) | 0.05 | 0.05 | 0.05 | 0.50 | 0.59 |
| TG (mmol/mL) | 6.03 | 6.60 | 6.57 | 0.00 | 0.83 |
| TC (mmol/mL) | 3.10 | 3.03 | 3.18 | 0.24 | 0.56 |
| GSH-PX (U/mL) | 760.5 | 714.0 | 683.25 | 37.8 | 0.71 |
| MDA (nmol/mL) | 6.61 a | 10.74 a | 13.17 b | 1.01 | 0.02 |
| SOD (U/mL) | 113.03 a | 109.02 a | 106.17 b | 1.04 | 0.01 |
| SOD/MDA | 18.49 a | 11.43 b | 9.91 b,c | 1.3 | 0.01 |
| T-AOC (U/mL) | 0.74 a | 2.96 b | 4.15 b,c | 0.39 | <0.01 |
| IgG (μg/mL) | 13.12 | 11.91 | 12.7 | 1.03 | 0.90 |
| IgA (μg/mL) | 59.43 | 51.86 | 52.32 | 2.87 | 0.50 |
| IgM (μg/mL) | 22.4 | 23.64 | 22.24 | 1.24 | 0.89 |
| Item (μg/mL) | Control | AFB20 | AFB40 | SEM | p Value |
|---|---|---|---|---|---|
| Acetate | 65.55 a | 62.54 a,b | 53.62 c | 1.63 | <0.01 |
| Propionate | 22.49 a | 22.75 a,b | 19.09 c | 0.59 | 0.01 |
| Acetate/Propionate | 2.93 | 2.77 | 2.81 | 0.05 | 0.44 |
| Isobutyrate | 12.73 a | 12.35 b | 9.73 c | 0.48 | <0.01 |
| Butyrate | 1.27 a | 1.44 a,b | 1.08 c | 0.05 | 0.01 |
| Isovalerate | 1.39 a | 1.29 a | 0.99 a,b | 0.05 | <0.01 |
| Valerate | 65.55 a | 62.53 a,b | 53.62 b,c | 1.62 | <0.01 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Zhang, Y.; Zheng, N.; Guo, L.; Song, X.; Zhao, S.; Wang, J. Biological System Responses of Dairy Cows to Aflatoxin B1 Exposure Revealed with Metabolomic Changes in Multiple Biofluids. Toxins 2019, 11, 77. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11020077
Wang Q, Zhang Y, Zheng N, Guo L, Song X, Zhao S, Wang J. Biological System Responses of Dairy Cows to Aflatoxin B1 Exposure Revealed with Metabolomic Changes in Multiple Biofluids. Toxins. 2019; 11(2):77. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11020077
Chicago/Turabian StyleWang, Qian, Yangdong Zhang, Nan Zheng, Liya Guo, Xiaoming Song, Shengguo Zhao, and Jiaqi Wang. 2019. "Biological System Responses of Dairy Cows to Aflatoxin B1 Exposure Revealed with Metabolomic Changes in Multiple Biofluids" Toxins 11, no. 2: 77. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11020077