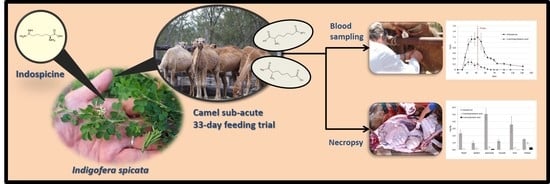
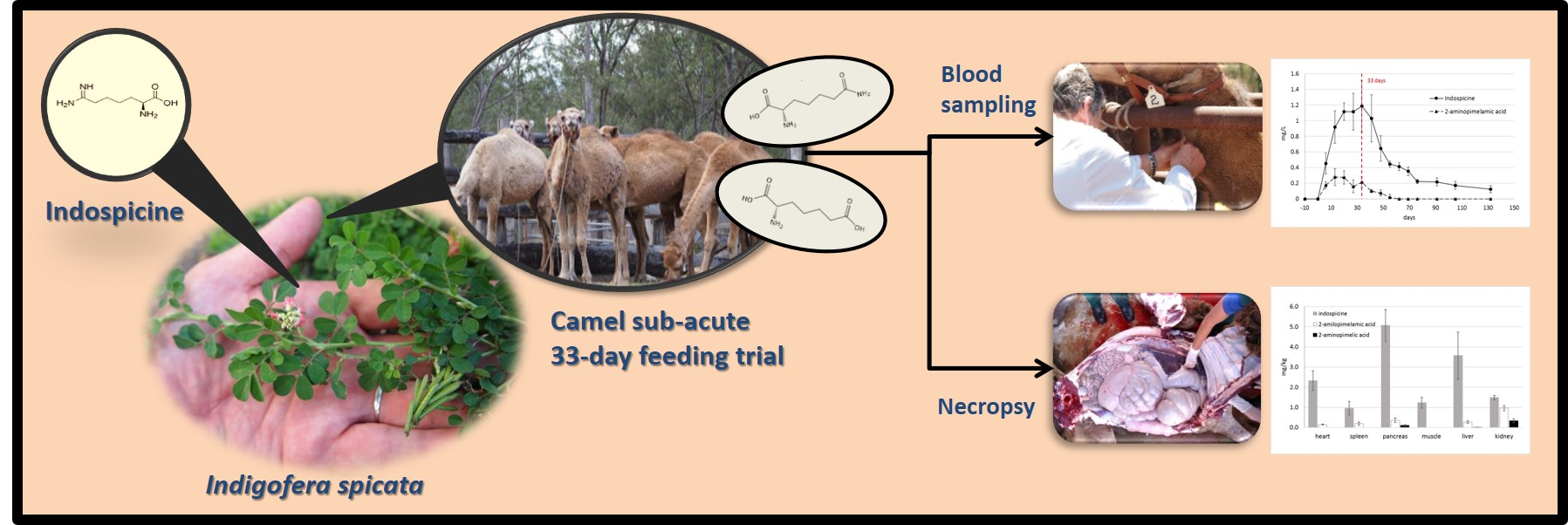
Bioaccumulation and Distribution of Indospicine and Its Foregut Metabolites in Camels Fed Indigofera spicata



Abstract
:
1. Introduction
2. Results and Discussion
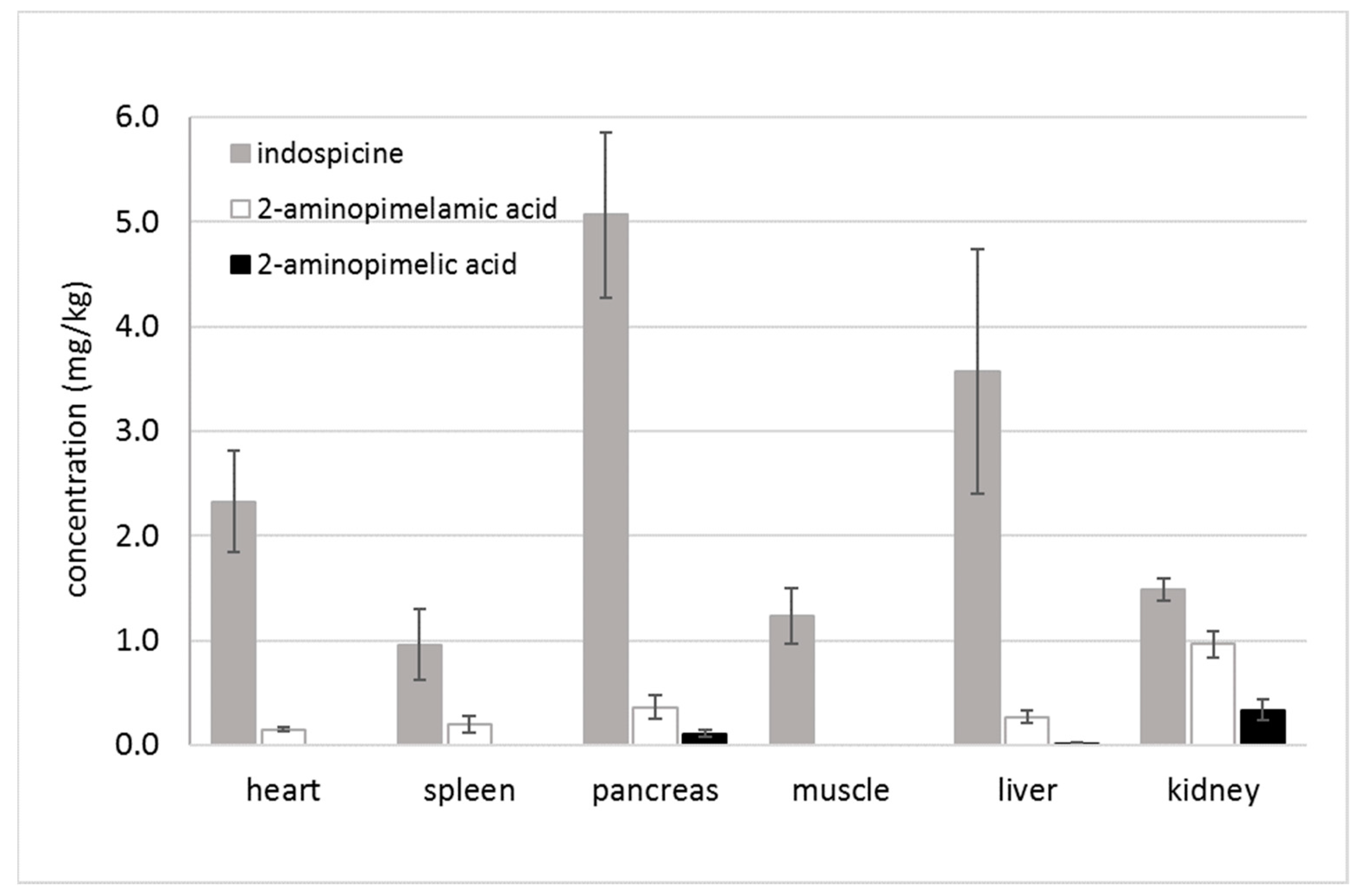
2.1. Indospicine and Foregut Metabolites in Tissue Samples
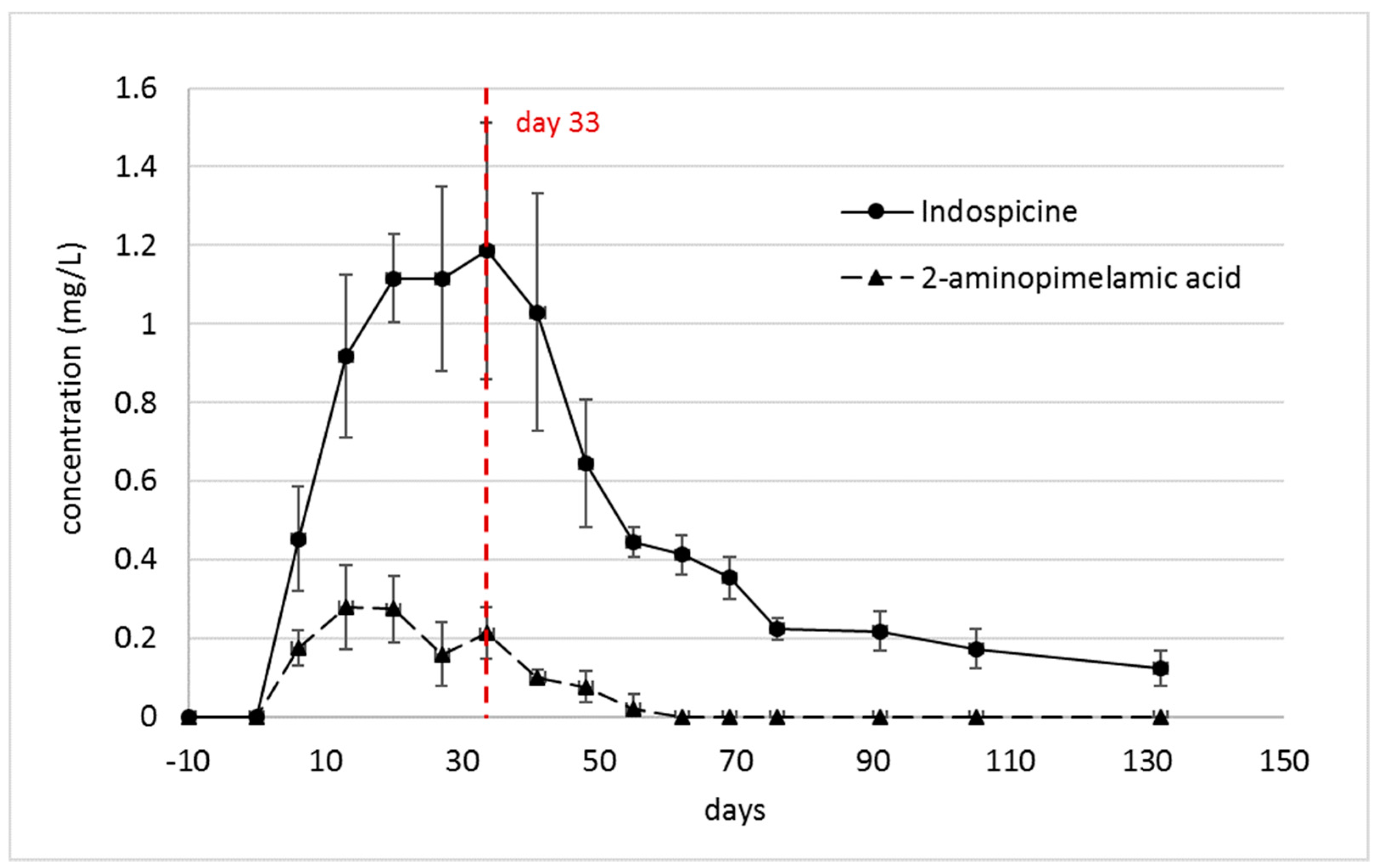
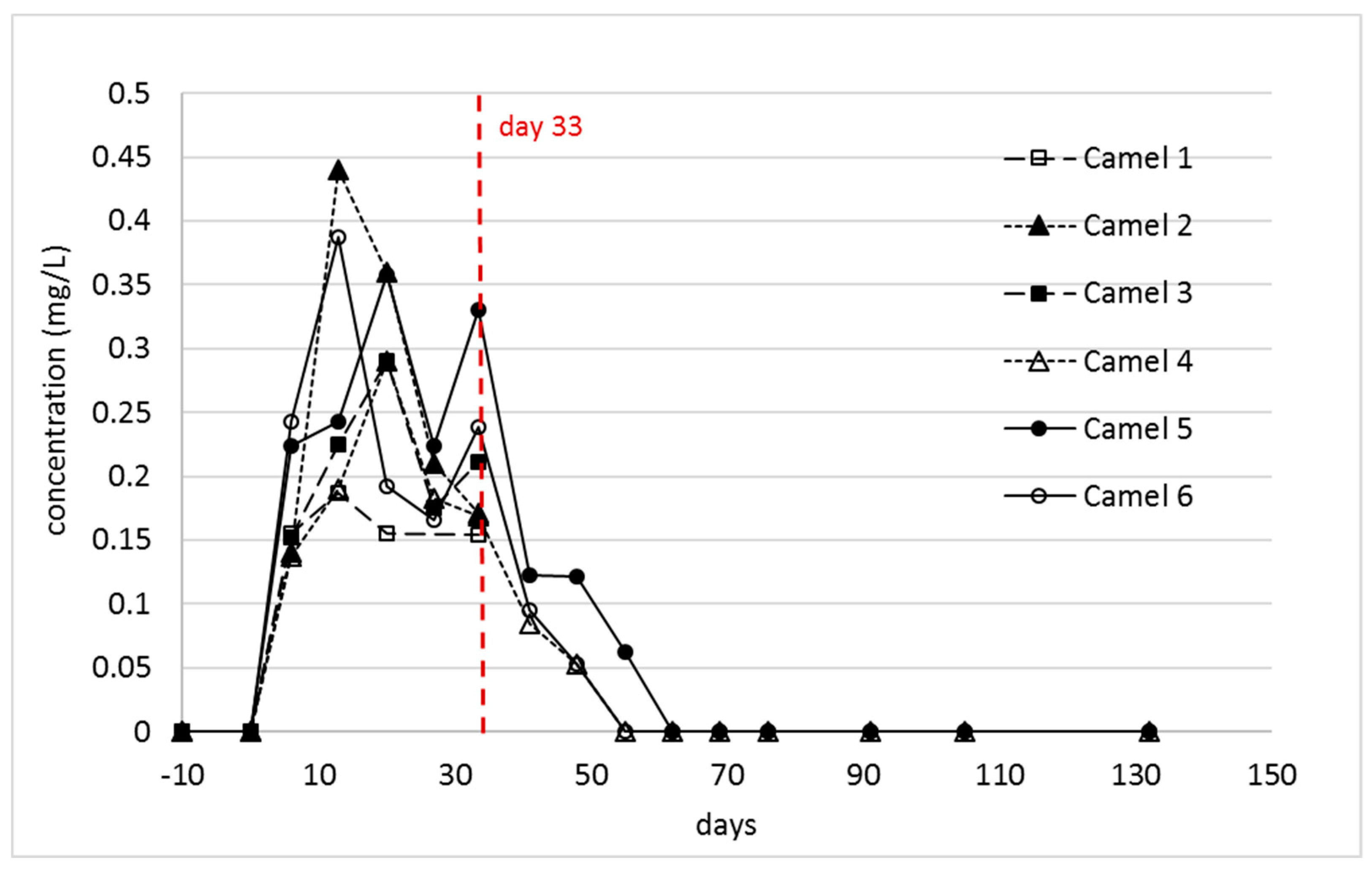
2.2. Indospicine and Foregut Metabolites in Plasma
3. Conclusions
4. Materials and Methods
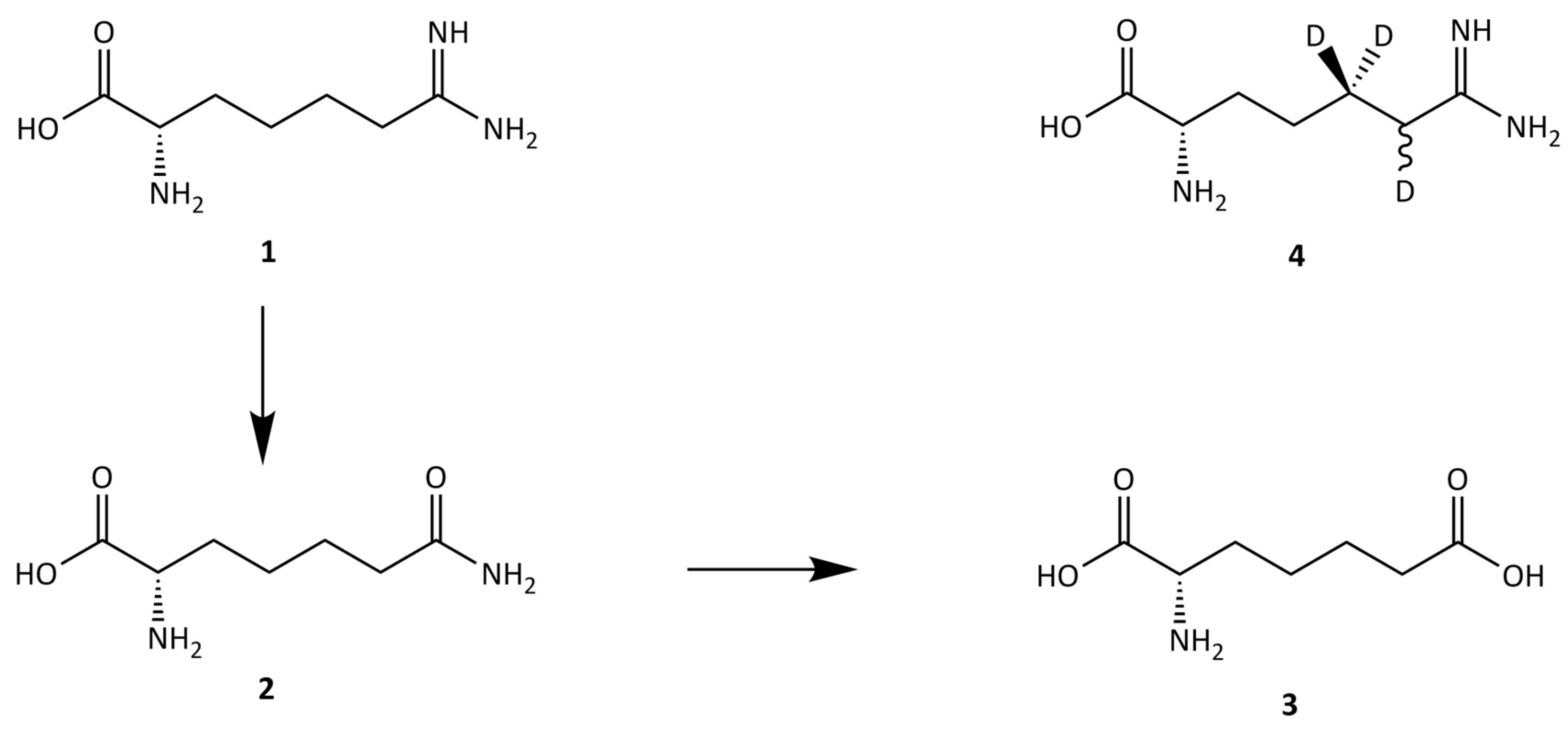
4.1. Standards and Reagents
4.2. Tissue Collection
4.3. Indospicine Extraction
4.4. UHPLC-MS/MS Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Tan, E.T.T.; Materne, C.M.; Silcock, R.G.; D’Arcy, B.R.; Al Jassim, R.; Fletcher, M.T. Seasonal and species variation of the hepatotoxin indospicine in Australian Indigofera legumes as measured by UPLC-MS/MS. J. Agric. Food. Chem. 2016, 64, 6613–6621. [Google Scholar] [CrossRef]
- Wilson, P.G.; Rowe, R. A revision of the Indigofereae (Fabaceae) in Australia. 1. Indigastrum and the simple or unifoliolate species of Indigofera. Telopea 2004, 10, 651–682. [Google Scholar]
- Wilson, P.G.; Rowe, R. A revision of the Indigofereae (Fabaceae) in Australia. 2. Indigofera species with trifoliolate and alternately pinnate leaves. Telopea 2008, 12, 293–307. [Google Scholar] [CrossRef]
- Wilson, P.G.; Rowe, R. Three new species of Indigofera (Fabaceae: Faboideae) from Cape York Peninsula. Telopea 2008, 12, 285–292. [Google Scholar] [CrossRef]
- FitzGerald, L.M.; Fletcher, M.T.; Paul, A.E.H.; Mansfield, C.S.; O’Hara, A.J. Hepatotoxicosis in dogs consuming a diet of camel meat contaminated with indospicine. Aust. Vet. J. 2011, 89, 95–100. [Google Scholar] [CrossRef]
- Fletcher, M.T.; Al Jassim, R.A.M.; Cawtdell-Smith, A.J. The occurrence and toxicity of indospicine to grazing animals. Agriculture 2015, 5, 427–440. [Google Scholar] [CrossRef]
- Hegarty, M.P.; Kelly, W.R.; McEwan, D.; Williams, O.J.; Cameron, R. Hepatotoxicity to dogs of horse meat contaminated with indospicine. Aust. Vet. J. 1988, 65, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Hutton, E.M.; Windrum, G.M.; Kratzing, C.C. Studies on the toxicity of Indigofera endecaphylla: I. Toxicity for rabbits. J. Nutr. 1958, 64, 321–337. [Google Scholar] [CrossRef] [PubMed]
- Nordfeldt, S.; Henke, L.A.; Morita, K.; Matsumoto, H.; Takahash, M.; Younge, O.R.; Willers, E.H.; Cross, R.F. Feeding tests with Indigofera endecaphylla Jacq. (Creeping indigo) and some observations on its poisonous effects on domestic animals. Hawaii Agric. Exp. Station Coll. Agric. Univ. Hawaii Tech. Bull. 1952, 15, 5–23. [Google Scholar]
- Hegarty, M.P. Toxic amino acids in foods of animals and man. Proc. Nutr. Soc. Australia 1986, 11, 73–81. [Google Scholar]
- Fowler, M.E. Plant poisoning in free-living wild animals: A review. J. Wildl. Dis. 1983, 19, 34–43. [Google Scholar] [CrossRef]
- Tan, E.T.T.; Al Jassim, R.; D’Arcy, B.R.; Fletcher, M.T. Level of natural hepatotoxin (Indospicine) contamination in Australian camel meat. Food Addit. Contam. Part A 2016, 33, 1587–1595. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.T.T.; Al Jassim, R.; Cawdell-Smith, A.J.; Ossedryver, S.M.; D’Arcy, B.R.; Fletcher, M.T. Accumulation, persistence, and effects of indospicine residues in camels fed Indigofera plant. J. Agric. Food. Chem. 2016, 64, 6622–6629. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.T.T.; Al Jassim, R.; D’Arcy, B.R.; Fletcher, M.T. In vitro biodegradation of hepatotoxic indospicine in Indigofera spicata and its degradation derivatives by camel foregut and cattle rumen fluids. J. Agric. Food. Chem. 2017, 65, 7528–7534. [Google Scholar] [CrossRef] [PubMed]
- Sultan, S.; Giles, C.; Netzel, G.; Osborne, S.A.; Netzel, M.E.; Fletcher, M.T. Release of indospicine from contaminated camel meat following cooking and simulated gastrointestinal digestion: Implications for human consumption. Toxins 2018, 10, 356. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.T.T.; Yong, K.W.L.; Wong, S.H.; D’Arcy, B.R.; Al Jassim, R.; De Voss, J.J.; Fletcher, M.T. Thermo-alkaline treatment as a practical degradation strategy to reduce indospicine contamination in camel meat. J. Agric. Food. Chem. 2016, 64, 8447–8453. [Google Scholar] [CrossRef]
- Hegarty, M.P.; Pound, A.W. Indospicine, a hepatotoxic amino acid from Indigofera spicata: Isolation, structure, and biological studies. Aust. J. Biol. Sci 1970, 23, 831–842. [Google Scholar] [CrossRef]
- Fletcher, M.T.; Reichmann, K.G.; Ossedryver, S.M.; McKenzie, R.A.; Carter, P.D.; Blaney, B.J. Accumulation and depletion of indospicine in calves (Bos taurus) fed creeping indigo (Indigofera spicata). Anim. Prod. Sci. 2018, 58, 568–576. [Google Scholar] [CrossRef]
- Madsen, N.P.; Hegarty, M.P. Inhibition of rat liver homogenate arginase activity in vitro by the hepatotoxic amino acid indospicine. Biochem. Pharmacol. 1970, 19, 2391–2393. [Google Scholar] [CrossRef]
- Biczó, G.; Hegyi, P.; Berczi, S.; Dósa, S.; Hracskó, Z.; Varga, I.S.; Iványi, B.; Venglovecz, V.; Wittmann, T.; Takács, T.; et al. Inhibition of arginase activity ameliorates L-arginine-induced acute pancreatitis in rats. Pancreas 2010, 39, 868–874. [Google Scholar]
- Emmanuel, B. Urea cycle enzymes in tissues (liver, rumen epithelium, heart, kidney, lung and spleen) of sheep (Ovis aries). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1980, 65, 693–697. [Google Scholar] [CrossRef]
- Morris, S.M. Regulation of enzymes of urea and arginine synthesis. Annu. Rev. Nutr. 1992, 12, 81–101. [Google Scholar] [CrossRef]
- Hegarty, M.P.; Court, R.D. Indigofera spicata. In Tropical Crops and Pastures Division of CSIRO 1975-1976 Annual Report; Commonwealth Scientific and Industrial Research Organisation: Brisbane, QLD, Australia, 1976; p. 70. [Google Scholar]
- Sultan, S.; Osborne, S.A.; Addepalli, R.; Netzel, G.; Netzel, M.E.; Fletcher, M.T. Indospicine cytotoxicity and transport in human cell lines. Food Chem. 2018, 267, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Lang, C.S.; Wong, S.H.; Chow, S.; Challinor, V.L.; Yong, K.W.L.; Fletcher, M.; Arthur, D.M.; Ng, J.C.; De Voss, J.J. Synthesis of L-indospicine, [5,5,6-2H3]-l-indospicine and L-norindospicine. Org. Biomol. Chem. 2016, 14, 6826–6832. [Google Scholar] [CrossRef]
- Tan, E.T.; Fletcher, M.T.; Yong, K.W.; D’Arcy, B.R.; Al Jassim, R. Determination of hepatotoxic indospicine in Australian camel meat by ultra-performance liquid chromatography-tandem mass spectrometry. J. Agric. Food. Chem. 2014, 62, 1974–1979. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Collison Energy (eV) | |
|---|---|---|
| Quantifier SRM | Verifier SRM | |
| Indospicine (1) | −23.0 | −17.0 |
| D3-l-indospicine (4) | −17.0 | −16.5 |
| 2-aminopimelamic acid (2) | −15.0 | −28.0 |
| 2-aminopimelic acid (3) | −15.0 | −20.0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Netzel, G.; Tan, E.T.T.; Yin, M.; Giles, C.; Yong, K.W.L.; Al Jassim, R.; Fletcher, M.T. Bioaccumulation and Distribution of Indospicine and Its Foregut Metabolites in Camels Fed Indigofera spicata. Toxins 2019, 11, 169. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11030169
Netzel G, Tan ETT, Yin M, Giles C, Yong KWL, Al Jassim R, Fletcher MT. Bioaccumulation and Distribution of Indospicine and Its Foregut Metabolites in Camels Fed Indigofera spicata. Toxins. 2019; 11(3):169. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11030169
Chicago/Turabian StyleNetzel, Gabriele, Eddie T. T. Tan, Mukan Yin, Cindy Giles, Ken W. L. Yong, Rafat Al Jassim, and Mary T. Fletcher. 2019. "Bioaccumulation and Distribution of Indospicine and Its Foregut Metabolites in Camels Fed Indigofera spicata" Toxins 11, no. 3: 169. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11030169