A Natural Variation of Fumonisin Gene Cluster Associated with Fumonisin Production Difference in Fusarium fujikuroi

Abstract
:1. Introduction
2. Results
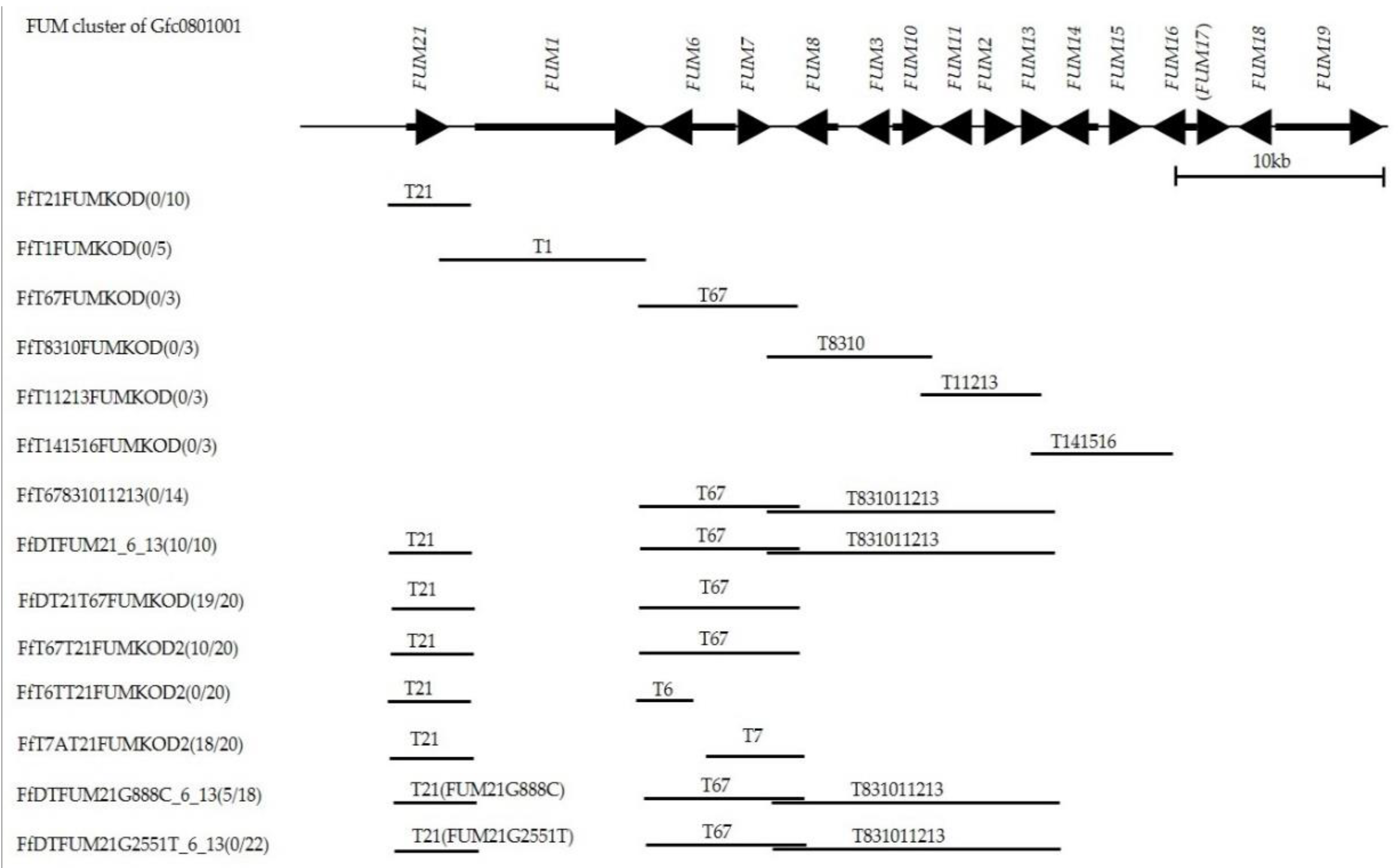
2.1. Linkage Analysis of Fumonisin Nonproduction
2.2. Sequence Comparison of the FUM Cluster
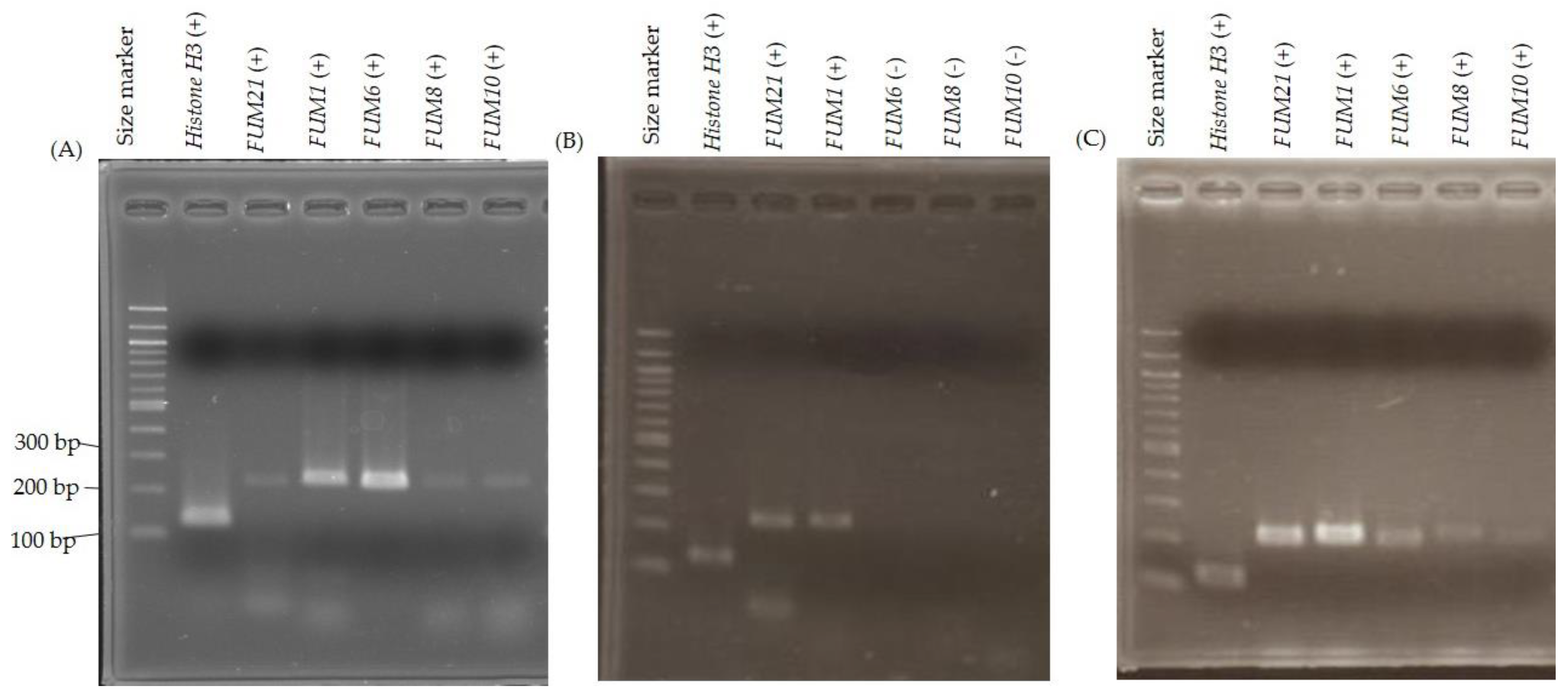
2.3. FUM Gene Expression
2.4. Fumonisin Production Recovery by Gene Complementation
2.5. Causative Mutation in FUM21
3. Discussion
4. Materials and Methods
4.1. Fungal Strains
4.2. Crossing
4.3. DNA Extraction
4.4. SNP Analysis by Luminex and MAT Typing
4.5. PCR and Sequencing
4.6. Construction of the Transformation Vector
4.7. Fungal Transformation
4.8. RT-PCR
4.9. Fumonisin Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fotso, J.; Leslie, J.F.; Smith, J.S. Production of beauvericin, moniliform, fusaproliferin and fumonisins B1, B2 and B3 by fifteen ex-type strains of Fusarium species. Appl. Environ. Microbiol. 2002, 68, 5195–5197. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E.; Plattner, R.D.; Nelson, P.E. Production of fumonisin B1 and moniliformin by Gibberella fujikuroi rice from various geographic areas. Appl. Environ. Microbiol. 1997, 63, 1838–1842. [Google Scholar]
- Nelson, P.E.; Plattner, R.D.; Shackelford, D.D.; Desjardins, A.E. Fumonisin B1 production by Fusarium species other than F. moniliforme in section Liseola and by some related species. Appl. Environ. Microbiol. 1992, 58, 984–989. [Google Scholar] [PubMed]
- Rheeder, J.P.; Marasas, W.F.O.; Vismer, H.F. Production of fumonisin analogs by Fusarium species. Appl. Environ. Microbiol. 2002, 68, 2101–2105. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhu, T. Mycoflora analysis in staple corn from Linxian Jinggtoucun with high incidence of esophageal cancer. J. China Agric. Univ. 2002, 7, 5–8. [Google Scholar]
- Frisvad, J.C.; Smedsgaard, J.; Samson, R.A.; Larsen, T.O.; Thrane, U. Fumonisin B2 production by Aspergillus niger. J. Agric. Food Chem. 2007, 55, 9727–9732. [Google Scholar] [CrossRef] [PubMed]
- Marasas, W.F.O. Fumonisins: History, worldwide occurrence and the impact. In Fumonisins in Food; Jackson, L.S., De Varies, J.W., Bullerman, L.B., Eds.; Plenum Publishing Co.: New York, NY, USA, 1996; pp. 1–17. [Google Scholar]
- Harrison, L.R.; Colvin, B.M.; Green, J.T.; Newman, L.E.; Cole, J.R. Pulmonary edema and hydrothorax in swine produced by fumonisin B1, a toxic metabolite of Fusarium moniliforme. J. Vet. Diagn. Investig. 1990, 2, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Marasas, W.F.O.; Miller, J.D.; Riley, R.T.; Visconti, A. Fumonisins occurrence, toxicology, metabolism and risk assessment. In Fusarium; Summerell, B.A., Lesilie, J.F., Backhouse, D., Bryden, W.L., Burgess, L., Eds.; The American Phytopathological Society Press: St. Paul, MN, USA, 2001; pp. 332–359. [Google Scholar]
- Marasas, W.F.O.; Riley, R.T.; Hendricks, K.A.; Stevens, V.L.; Sadler, T.W.; Gelineau-van Waes, J.; Missmer, S.A.; Cabrera, J.; Torres, O.; Gelderblom, W.C.A.; et al. Fumonisins disrupt sphingolipid metabolism, folate transport, and neural tube development in embryo culture and in vivo: A potential risk factor for human neural tube defects among populations consuming fumonisin-contaminated maize. J. Nutr. 2004, 134, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Abbas, H.K.; Tanaka, T.; Duke, S.O.; Porter, J.K.; Wray, E.M.; Hodges, L.; Sessions, A.E.; Wang, E.; Merril, A.H., Jr.; Riley, R.T. Fumonisin-and AAL-toxin-induced disruption of sphingolipid metabolism with accumulation of free sphingoid bases. J. Plant Physiol. 1994, 106, 1085–1093. [Google Scholar] [CrossRef]
- Ross, P.F.; Rice, R.D.; Plattner, G.D.; Osweiler, T.M.; Wilson, D.L.; Owens, H.A.; Nelson; Richard, J.L. Concentrations of fumonisin B1 in feeds associated with animal health problems. Mycopathologia 1991, 114, 129–135. [Google Scholar] [CrossRef]
- Cruz, A.; Marín, P.; González-Jaén, M.T.; Aguilar, K.G.I.; Cumagun, C.J.R. Phylogenetic analysis, fumonisin production and pathogenicity of Fusarium fujikuroi strains isolated from rice in the Philippines. J. Sci. Food Agric. 2013, 93, 3032–3039. [Google Scholar] [CrossRef]
- Leslie, J.F.; Zeller, K.A.; Wohler, M.; Summerell, B.A. Interfertility of two mating populations in the Gibberella fujikuroi species complex. Eur. J. Plant Pathol. 2004, 110, 611–618. [Google Scholar] [CrossRef]
- Leslie, J.F.; Anderson, L.L.; Bowden, R.L.; Lee, Y.-W. Inter- and intra-specific genetic variation in Fusarium. Int. J. Food. Microbiol. 2007, 119, 25–32. [Google Scholar] [CrossRef]
- Lee, S.-H.; Kim, J.-H.; Son, S.-W.; Lee, T.; Yun, S.-H. Fumonisin production by field isolates of the Gibberella fujikuroi species complex from rice and corn in Korea. Res. Plant Dis. 2012, 18, 310–316. [Google Scholar] [CrossRef]
- Suga, H.; Kitajima, M.; Nagumo, R.; Tsukiboshi, T.; Uegaki, R.; Nakajima, T.; Kushiro, M.; Nakagawa, H.; Shimizu, M.; Kageyama, K. A single nucleotide polymorphism in the translation elongation factor 1α gene correlates with the ability to produce fumonisin in Japanese Fusarium fujikuroi. Fungal Biol. 2014, 118, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Rosler, S.M.; Sieber, M.K.C.; Humpf, H.-U.; Tudzynki, B. Interplay between pathway-specific and global regulation of the fumonisin gene cluster in the rice pathogen Fusarium fujikuroi. Appl. Microbiol. Biotechnol. 2016, 100, 5869–5882. [Google Scholar] [CrossRef] [PubMed]
- Suga, H.; Arai, M.; Fukasawa, E.; Motohashi, K.; Nakagawa, H.; Tateishi, H.; Fuji, S.; Shimizu, M.; Kageyama, K.; Hyakumachi, M. Genetic differentiation associated with fumonisin and gibberellin production in Japanese Fusarium fujikuroi. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef]
- Niehaus, E.-M.; Kim, H.-K.; Münsterkötter, M.; Janevska, S.; Arndt, B.; Kalinina, S.A.; Houterman, P.M.; Ahn, I.-P.; Alberti, I.; Tonti, S.; et al. Comparative genomics of geographically distant Fusarium fujikuroi isolates revealed two distinct pathotypes correlating with secondary metabolite profiles. PLoS Pathog. 2017, 13, e1006670. [Google Scholar] [CrossRef]
- Jeon, Y.-A.; Yu, S.-H.; Lee, Y.Y.; Park, H.-J.; Lee, S.; Sung, J.S.; Kim, Y.-G.; Lee, H.-S. Incidence molecular characteristics and pathogenicity of Gibberella fujikuroi species complex associated with rice seeds from Asian countries. Mycobiology 2013, 41, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Sydenham, E.W.; Shepherd, G.S.; Thiel, P.G.; Marasas, W.F.O.; Stockenstrom, S. Fumonisin contamination of commercial corn-based human foodstuffs. J. Agric. Food Chem. 1991, 3, 2014–2018. [Google Scholar] [CrossRef]
- Theil, P.G.; Marasas, W.F.O.; Sydenham, E.W.; Shephard, G.S.; Gelderblom, W.C.A. The implications of naturally occurring levels of fumonisins in corn for human and animal health. Mycopathologia 1992, 117, 3–9. [Google Scholar] [CrossRef]
- Visconti, A.; Doko, M.B. Survey of fumonisin production by Fusarium isolated from cereals in Europe. J. AOAC Int. 1994, 77, 546–550. [Google Scholar]
- Shephard, G.S.; Theil, P.G.; Stokenstrom, S.; Sydenham, E.W. Worldwide survey of fumonisin contamination of corn and corn-based products. J. AOAC Int. 1996, 79, 671–687. [Google Scholar] [PubMed]
- Bolton, S.L.; Brannen, P.M.; Glenn, A.E. A novel population of Fusarium fujikuroi isolated from southeastern U.S. winegrapes reveals the need to re-evaluate the species’ fumonisin production. Toxins 2016, 8, 254. [Google Scholar] [CrossRef]
- Proctor, R.H.; Brown, D.W.; Plattner, R.D.; Desjardins, A.E. Co-expression of 15 contiguous genes delineates a fumonisin biosynthetic gene cluster in Gibberella moniliformis. Fungal Genet. Biol. 2003, 38, 237–249. [Google Scholar] [CrossRef]
- Butchko, R.A.E.; Plattner, R.D.; Proctor, R.H. Deletion analysis of FUM genes involved in tricarballylic ester formation during fumonisin during fumonisin biosynthesis. J. Agric. Food Chem. 2006, 54, 9398–9404. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E. Fumonisins. In Fusarium Mycotoxins Chemistry, Genetics, and Biology; Desjardins, A.E., Ed.; The American Phytopathology Society Press: St. Paul, MN, USA, 2006; pp. 79–108. [Google Scholar]
- Brown, D.W.; Butchko, R.A.; Busman, M.; Proctor, R.H. The Fusarium verticillioides fum gene cluster encodes a Zn(II)2Cys6 protein that affects FUM gene expression and fumonisin production. Eukaryot. Cell 2007, 6, 1210–1218. [Google Scholar] [CrossRef] [PubMed]
- Glenn, A.E.; Zitomer, N.C.; Zimeri, A.M.; Williams, L.D.; Riley, R.T.; Proctor, R.H. Transformation-mediated complementation of a FUM gene cluster deletion in Fusarium verticillioides of restores both fumonisin and pathogenicity on maize seedlings. Mol. Plant Microbe Interact. 2007, 21, 87–97. [Google Scholar] [CrossRef]
- Chiara, M.; Fanelli, F.; Mulè, G.; Logrieco, A.F.; Pesole, G.; Leslie, J.F.; Horner, D.S.; Toomajian, C. Genome sequencing of multiple isolates highlights subtelomeric genomic diversity within Fusarium fujikuroi. Genome Biol. Evol. 2015, 7, 3062–3069. [Google Scholar] [CrossRef]
- Wiemann, P.; Sieber, C.M.K.; von Bargen, K.W.; Studt, L.; Niehaus, E.-M.; Espino, J.J.; Huß, K.; Michielse, C.B.; Albermann, S.; Wagner, D.; et al. Deciphering the cryptic genome: Genome-wide analyses of the rice pathogen Fusarium fujikuroi reveal complex regulation of secondary metabolism and novel metabolites. PLoS. Pathog. 2013, 9, e1003475. [Google Scholar] [CrossRef]
- Tominaga, M.; Lee, Y.-H.; Hayashi, R.; Suzuki, Y.; Yamada, O.; Sakamoto, K.; Gotoh, K.; Akita, O. Molecular analysis of an inactive aflatoxin biosynthesis gene cluster in Aspergillus oryzae RIB strains. Appl. Environ. Microbiol. 2006, 72, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.-K.; Matsushima, K.; Takahashi, T.; Yu, J.; Abe, D.B.; Yuan, G.-F.; Koyama, Y.; Cleveland, T.E. Understanding nonaflatoxigenicity of Aspergillus sojae: A windfall of aflatoxin biosynthesis research. Appl. Microbiol. Biotechnol. 2007, 76, 977–984. [Google Scholar] [CrossRef]
- Ma, J.; Ptashne, M. Deletion analysis of GAL4 defines two transcriptional activating segments. Cell 1987, 48, 847–853. [Google Scholar] [CrossRef]
- Malonek, S.; Rojas, M.C.; Hedden, P.; Gaskin, P.; Hopkins, P.; Tudzynski, B. Functional characterization of two cytochrome P450 monooxygenase genes, P450-1 and P450-4, of the gibberellic acid gene cluster in Fusarium proliferatum (Gibberella fujikuroi MP-D). Appl. Environ. Microbiol. 2005, 3, 1462–1472. [Google Scholar] [CrossRef] [PubMed]
- Malonek, S.; Rojas, M.C.; Hedden, P.; Hopkins, P.; Tudzynski, B. Restoration of gibberellin production in Fusarium proliferatum by functional complementation of enzymatic blocks. Appl. Environ. Microbiol. 2005, 71, 6014–6025. [Google Scholar] [CrossRef]
- Bowden, R.L.; Leslie, J.F. Sexual recombination in Gibberella zeae. Phytopathology 1999, 89, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Suga, H.; Karugia, G.W.; Ward, T.; Gale, L.R.; Tomimura, K.; Nakajima, T.; Miyasaka, A.; Koizumi, S.; Kageyama, K.; Hyakumachi, M. Molecular characterization of Fusarium graminearum species complex in Japan. Phytopathology 2008, 98, 159–166. [Google Scholar] [CrossRef]
- Malonek, S.; Rojas, M.C.; Hedden, P.; Gaskin, P.; Hopkins, P. The NADPH-cytochrome p450 reductase gene from Gibberella fujikuroi is essential for gibberellin biosynthesis. J. Biol. Chem. 2004, 279, 25075–25084. [Google Scholar] [CrossRef] [PubMed]
- Kerenyi, Z.; Moretti, A.; Walwijk, C.; Olah, B.; Hornok, L. Mating type sequences in asexually reproducing Fusarium species. Appl. Environ. Microbiol. 2004, 70, 4419–4423. [Google Scholar] [CrossRef]
- Lian, C.; Hogetsu, T. A protocol for efficient development of microsatellite markers. J. Jpn. For. Soc. 2004, 86, 191–198. (In Japanese) [Google Scholar]
- Carroll, A.M.; Sweigard, J.A.; Valent, B. Improved vectors for selecting resistance to hygromycin. Fungal Genet. Newsl. 1994, 41, 20–21. [Google Scholar] [CrossRef]
- Kück, U.; Hoff, B. Application of the nourseothricin acetyltransferase gene (nat1) as dominant marker for the transformation of filamentous fungi. Fungal Genet. Rep. 2006, 53, 9–11. [Google Scholar] [CrossRef]
- Namiki, F.; Matsunaga, M.; Okuda, M.; Inoue, I.; Nishi, K.; Fujita, Y.; Tsuge, T. Mutation of an arginine biosynthesis gene causes reduced pathogenicity in Fusarium oxysporum f. sp. melonis. Mol. Plant Microbe Interact. 2001, 14, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Suga, H.; Kageyama, K.; Shimizu, M.; Hyakumachi, M. A natural mutation involving both pathogenicity and perithecium formation in the Fusarium graminearum species complex. G3 2016, 6, 3883–3892. [Google Scholar] [CrossRef] [PubMed]
- Clouse, S.D.; Martensen, A.N.; Gilchrist, D.G. Rapid purification of host-specific pathotoxins from Alternaria alternata f. sp. lycopersici by solid-phase adsorption on octadecylsilane. J. Chromatogr. 1985, 350, 255–263. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | Gene ID a | Predictive Function b | Homology (%) | Length of Amino Acid c | The Number of Different Amino Acid d | |
|---|---|---|---|---|---|---|
| Nucleic Acid | Amino Acid | |||||
| FUM21 | FFUJ_09240 e | Zn(II)2Cys6 type-transcriptional regulator | 99.6 | 99.8 | 688 e | 1 substitution, 11 deletion |
| FUM1 | FFUJ_09241 | Polyketide synthase | 99.6 | 99.6 | 2580 | 8 substitutions |
| FUM6 | FFUJ_09242 | Fumonisin oxygenase | 99.3 | 99.2 | 1115 | 8 substitutions |
| FUM7 | FFUJ_09243 | Alcohol dehydrogenase | 98.8 | 99.2 | 420 | 3 substitutions |
| FUM8 | FFUJ_09244 | Aminotransferase | 98.2 | 98.3 | 830 | 14 substitutions, 2 insertion |
| FUM3 | FFUJ_09245 | Fumonisin 5-oxygenase | 99.3 | 99.6 | 300 | 1 substitution |
| FUM10 | FFUJ_09246 | Tricarballylic esterification | 99.5 | 99.4 | 561 | 3 substitutions |
| FUM11 | FFUJ_09247 | Tricarballylic esterification | 98.7 | 98.3 | 300 | 5 substitutions |
| FUM2 | FFUJ_09248 | Fumonisin 10-oxygenase | 98.8 | 99.0 | 502 | 5 substitutions |
| FUM13 | FFUJ_09249 | C3 carbonyl reductase | 99.5 | 99.5 | 367 | 2 substitutions |
| FUM14 | FFUJ_09250 | Tricarballylic esterification | 99.1 | 99.1 | 556 | 5 substitutions, 5 insertion |
| FUM15 | FFUJ_09251 | Cytochromosome P450 monooxygenase | 98.7 | 98.6 | 599 | 8 substitutions |
| FUM16 | FFUJ_09252 | Fatty acyl-coenzyme A synthetase | 98.4 | 98.7 | 682 | 9 substitutions, 10 deletion |
| FUM17 | - f | Similarity to tomato longevity assurance factor (ASC-1) of Alternaria alternata f. sp. lycopersici | - f | - f | - f | - f |
| FUM18 | FFUJ_09253 | Similarity to tomato longevity assurance factor (ASC-1) of Alternaria alternata f. sp. lycopersici | 99.1 | 97.5 | 413 | 10 substitutions |
| FUM19 | FFUJ_09254 | Similarity to ATP-binding cassette (ABC) multidrug resistant transporter | 99.1 | 99.0 | 1505 | 15 substitutions |
| Transformant b | First Time Culture | Second Time Culture | Third Time Culture | ||||||
|---|---|---|---|---|---|---|---|---|---|
| FB1 | FB2 | FB3 | FB1 | FB2 | FB3 | FB1 | FB2 | FB3 | |
| FfDTFUM21_6_13(#1) | 43.63 | 1.12 | >10.00 | >50.00 | 7.57 | 7.96 | 4.62 | 0.85 | 1.29 |
| FfDTFUM21_6_13(#2) | 46.56 | 1.86 | >10.00 | >50.00 | 5.76 | >10.00 | >50.00 | 3.73 | 7.78 |
| FfDTFUM21_6_13(#3) | 24.80 | 1.01 | 8.35 | 0.41 | ND | 0.12 | 0.20 | ND | 0.12 |
| FfDTFUM21G888C_6_13(#5) | 3.70 | 0.20 | >1.00 | 47.03 | 4.09 | >10.00 | 27.15 | 2.45 | 5.22 |
| FfDTFUM21G888C_6_13(#6) | 37.23 | 1.28 | >10.00 | ND | ND | ND | ND | ND | ND |
| FfDTFUM21G888C_6_13(#7) | >50.00 | 2.39 | >10.00 | 0.19 | ND | 0.08 | ND | ND | ND |
| FfDT21T67FUMKOD(#11) | 1.57 | 0.26 | 0.98 | 0.95 | 0.19 | 0.30 | 1.33 | 0.24 | 0.36 |
| FfDT21T67FUMKOD(#16) | 3.62 | 0.38 | 0.71 | 2.88 | 0.93 | 0.94 | 1.72 | 0.67 | 0.79 |
| FfDT21T67FUMKOD(#17) | 0.57 | 0.17 | 0.09 | 2.59 | 1.42 | 0.36 | 0.14 | ND | ND |
| FfT67T21FUMKOD2(#7) | 0.20 | ND | 0.06 | ND | ND | ND | ND | ND | ND |
| FfT67T21FUMKOD2(#11) | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| FfT67T21FUMKOD2(#16) | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| FfT7AT21FUMKOD2(#23) | 0.19 | ND | 0.15 | ND | ND | ND | ND | ND | ND |
| FfT7AT21FUMKOD2(#34) | 0.11 | ND | 0.10 | ND | ND | ND | ND | ND | ND |
| FfT7AT21FUMKOD2(#18) | 0.13 | ND | 0.08 | 1.06 | ND | 0.25 | 0.48 | ND | 0.21 |
| Gfc0825009(Donor strain) | >50.00 | >50.00 | 3.70 | 44.40 | 35.70 | 2.13 | 42.60 | 36.66 | 2.59 |
| Gfc0801001(Original strain) | ND | ND | ND | ND | ND | ND | ND | ND | ND |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sultana, S.; Kitajima, M.; Kobayashi, H.; Nakagawa, H.; Shimizu, M.; Kageyama, K.; Suga, H. A Natural Variation of Fumonisin Gene Cluster Associated with Fumonisin Production Difference in Fusarium fujikuroi. Toxins 2019, 11, 200. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11040200
Sultana S, Kitajima M, Kobayashi H, Nakagawa H, Shimizu M, Kageyama K, Suga H. A Natural Variation of Fumonisin Gene Cluster Associated with Fumonisin Production Difference in Fusarium fujikuroi. Toxins. 2019; 11(4):200. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11040200
Chicago/Turabian StyleSultana, Sharmin, Miha Kitajima, Hironori Kobayashi, Hiroyuki Nakagawa, Masafumi Shimizu, Koji Kageyama, and Haruhisa Suga. 2019. "A Natural Variation of Fumonisin Gene Cluster Associated with Fumonisin Production Difference in Fusarium fujikuroi" Toxins 11, no. 4: 200. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11040200