Morphological and Molecular Identification of Microcystin-Producing Cyanobacteria in Nine Shallow Bulgarian Water Bodies

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Phytoplankton Species Composition Obtained by Light Microscopy (LM)
2.2. Results from PCR Analysis for Microcystin and Nodularin-Producing Strains
3. Discussion
- (1)
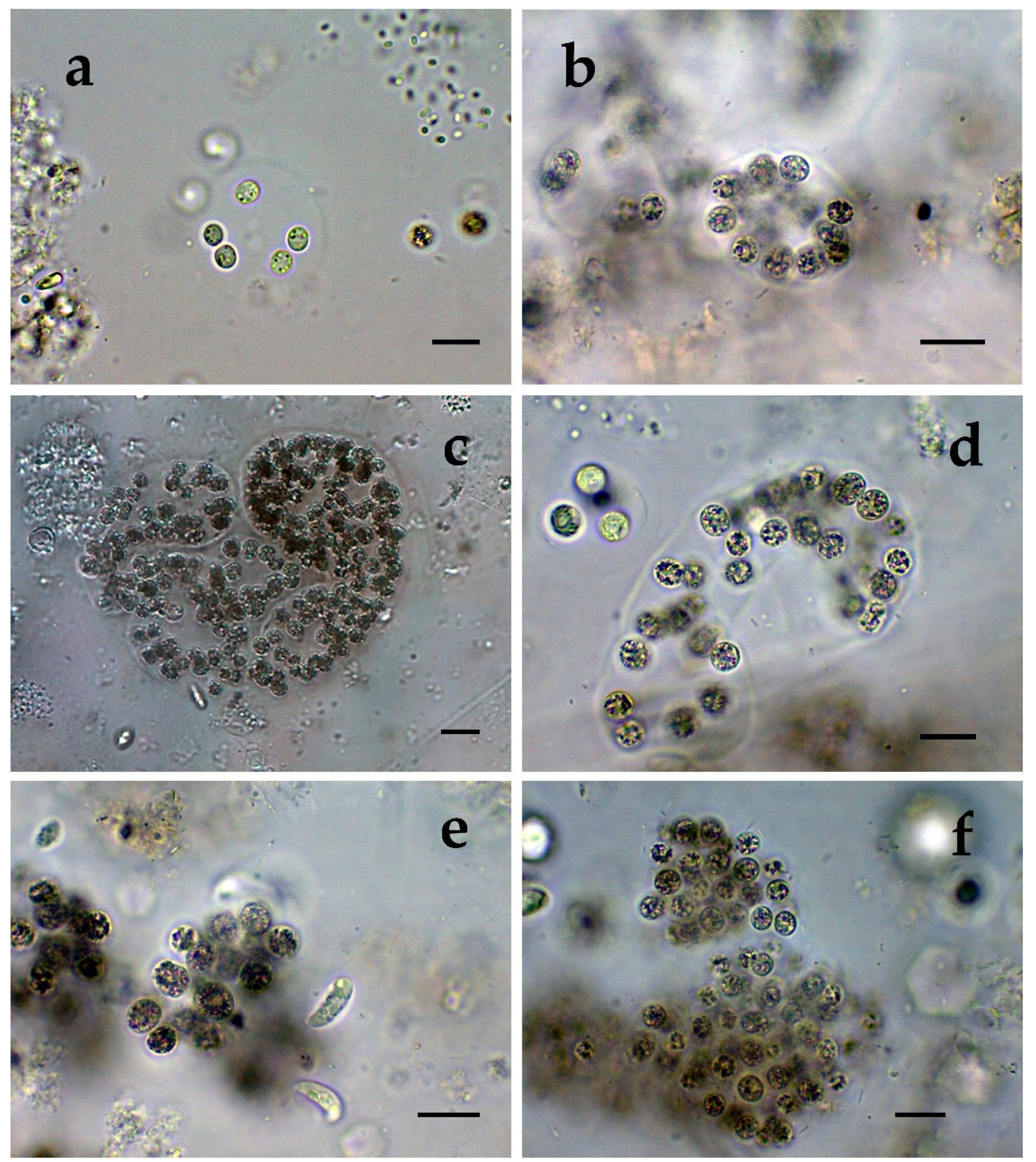
- Cluster I contains mainly strains identical to Microcystis sp. Kot12/08-3 (NCBI:txid1402958), which are similar in LM to Microcystis novacekii, from which the best morphologically expressed features of the colonies were found in the reservoir Mandra (Figure 5c–f);
- (2)
- Because of the finding of Microcystis natans only in Durankulak site 2 (Figure 3d) cluster II most probably comprises its two strains which are close to Microcystis sp. Brat 12/07-7 (NCBI:txid1402954) and uncultured cyanobacterium (AB638245.1);
- (3)
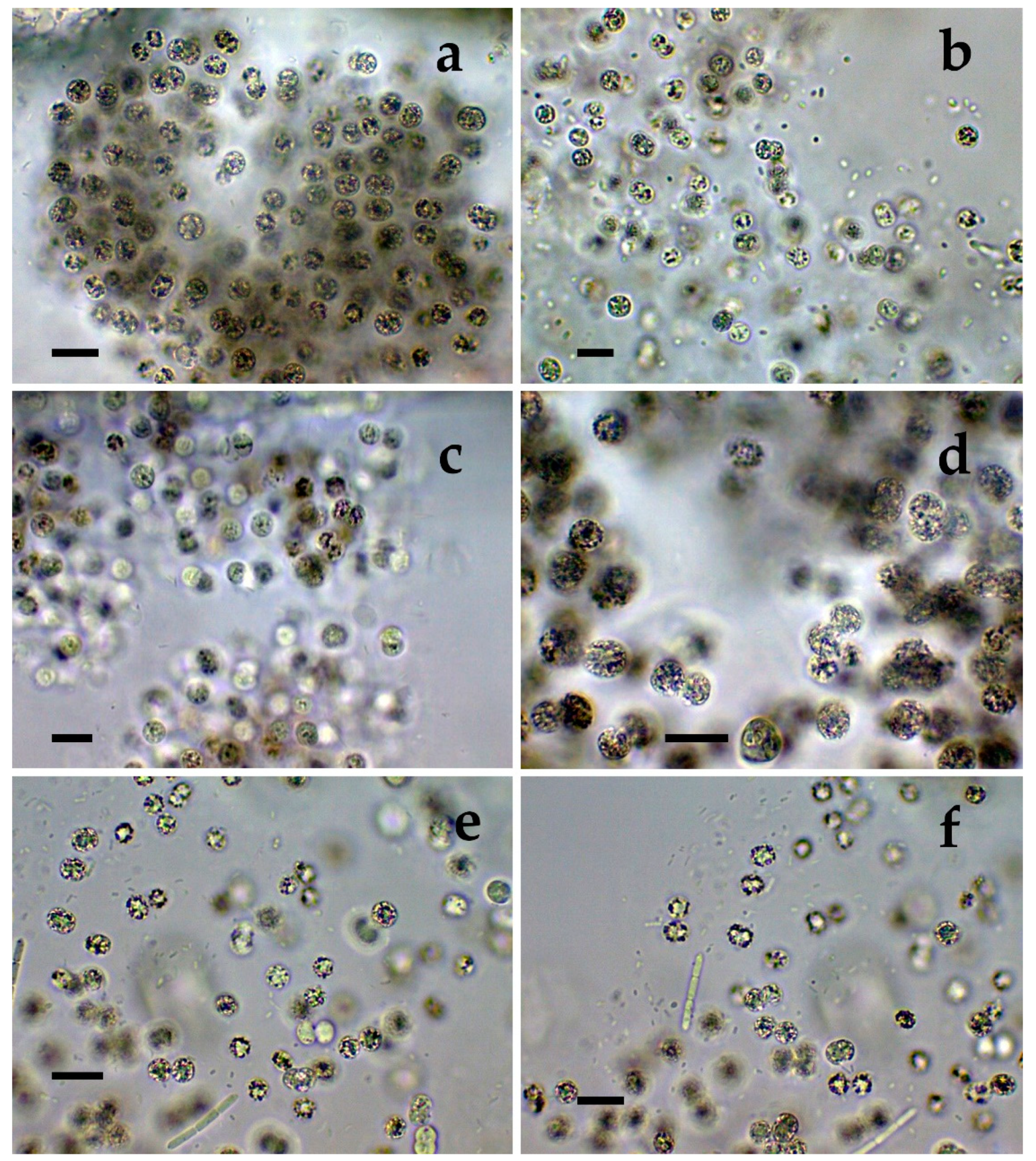
- Cluster III contained a group of seven strains of typical Microcystis wesenbergii (identical with Microcystis sp. Brat12/07-2, NCBI:txid1402949), some of which are capable of easy dissolving to separate cells accompanied with some morphological transitions to Microcystis aeruginosa during the disintegration of the colonies (Figure 4e,f) and four other groups of strains with disputable from genetic point of view affiliation despite the fact that by LM similar strains of M. wesenbergii were seen in the reservoir Sinyata Reka and in the lakes Durankulak (site 2) and Vaya (sites 1 and 2);
- (4)
- Cluster IV contained two groups of strains: (a) a strain of Microcystis wesenbergii (100% identical to Microcystis wesenbergii NIES-107, NCBI:txid315483) with sharp, hard and well-pronounced margin of the colonies, which easily defragment to small spherical initial colonies in the lake Vaya (Figure 4d); (b) strains from Vaya site 3 and Durankulak site 2 (the last similar to Uncultured cyanobacterium AB638231), for which it is possible to suppose close affiliation to Microcystis aeruginosa from cluster VI according to their genetic distances but which could not be clearly separated by LM;
- (5)
- Cluster V comprises generally of morphologically different strains of Microcystis aeruginosa with a sub-cluster of clones distributed in Vaya 1–2 and Poroy, the colonies of which are easily disintegrating in separate cells, often dividing in twos, which strongly resemble Synechocystis aquatilis and are impossible to be reliably ascertained to M. wesenbergii or M. aeruginosa but obviously are genetically close to uncultured Microcystis sp. clone BS12/06-10; another close sub-cluster is formed by strains found in Mandra 1, Durankulak 2, and Vaya 3 and identical with to Microcystis aeruginosa (AB032549.2) and uncultured Microcystis sp. clone Vi12/07-2, which could not be clearly separated by LM;
- (6)
- Cluster VI contains the most typical but strongly variable Microcystis aeruginosa strains, which were found mainly in the lake Durankulak (Figure 1a–f) and were identical to Microcystis aeruginosa FCY-26 (NCBI:txid1150859).
4. Conclusions
5. Materials and Methods
5.1. Sites and Sampling
5.2. Phytoplankton Identification by Conventional Light Microscopy (LM)
5.3. Molecular Studies
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Svirčev, Z.; Lalić, D.; Savić, G.B.; Tokodi, N.; Backović, D.D.; Chen, L.; Meriluoto, J.; Codd, G.A. Global geographical and historical overview of cyanotoxin distribution and cyanobacterial poisonings. Arch. Toxicol. 2019, 93, 2429–2481. [Google Scholar] [CrossRef] [PubMed]
- Stoyneva-Gärtner, M.P.; Descy, J.-P.; Latli, A.; Uzunov, B.; Pavlova, V.; Bratanova, Z.L.; Babica, P.; Maršálek, B.; Meriluoto, J.; Spoof, L. Assessment of cyanoprokaryote blooms and of cyanotoxins in Bulgaria in a 15-years period (2000–2015). Adv. Oceanogr. Limnol. 2017, 8, 131–152. [Google Scholar] [CrossRef] [Green Version]
- Descy, J.-P.; Stoyneva-Gärtner, M.P.; Uzunov, B.A.; Dimitrova, P.H.; Pavlova, V.T.; Gärtner, G. Studies on cyanoprokaryotes of the water bodies along the Bulgarian Black Sea Coast (1890–2017): A review, with special reference to new, rare and harmful taxa. Acta Zool. Bulg. 2018, 11, 43–52. [Google Scholar]
- Stoyneva-Gärtner, M.P.; Uzunov, B.A.; Descy, J.-P.; Gärtner, G.; Draganova, P.H.; Borisova, C.I.; Pavlova, V.; Mitreva, M. Pilot application of drone-observations and pigment marker detection by HPLC in the studies of CyanoHABs in Bulgarian inland waters. Mar. Freshw. Res. 2019. [Google Scholar] [CrossRef]
- Gkelis, S.; Rajaniemi, P.; Vardaka, E.; Moustaka-Gouni, M.; Lanaras, T.; Sivonen, K. Limnothrix redekei (Van Goor) Meffert (Cyanobacteria) strains from Lake Kastoria, Greece form a separate phylogenetic group. Microb. Ecol. 2005, 49, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Rajaniemi-Wacklin, P.; Rantala, A.; Mugnai, M.A.; Turicchia, S.; Ventura, S.; Komárková, J.; Lepistö, L.; Sivonen, K. Correspodence between phylogeny and morphology of Snowella spp. and Woronichinia naegeliana, cyanobacteria commonly occurring in lakes. J. Phycol. 2006, 42, 226–232. [Google Scholar] [CrossRef]
- Moustaka-Gouni, M.; Kormas, K.A.; Vardaka, E.; Katsiapi, M.; Gkelis, S. Raphidiopsis mediterranea Skuja represents non-heterocytous life-cycle stages of Cylindrospermopsis raciborskii (Woloszynska) Seenayya et Subba Raju in Lake Kastoria (Greece), its type locality: Evidence by morphological and phylogenetic analysis. Harmful Algae 2009, 8, 864–872. [Google Scholar] [CrossRef]
- Xiao, M.; Li, M.; Reynolds, C.S. Colony formation in the cyanobacterium Microcystis. Biol. Rev. 2018, 93, 1399–1420. [Google Scholar] [CrossRef] [Green Version]
- Komárek, J.; Kaštovsky, J.; Mareš, J.; Johansen, J.R. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Komárek, J. A polyphasic approach for the taxonomy of cyanobacteria: Principles and applications. Eur. J. Phycol. 2016, 51, 346–353. [Google Scholar] [CrossRef]
- Komárek, J. Several problems of the polyphasic approach in the modern cyanobacterial system. Hydrobiologia 2018, 811, 7–17. [Google Scholar] [CrossRef]
- Mankiewicz-Boczek, J.; Izydorczyk, K.; Romanowska-Duda, Z.; Jurczak, T.; Stefaniak, K.; Kokocinski, M. Detection and monitoring toxigenicity of cyanobacteria by application of molecular methods. Environ. Toxicol. 2006, 21, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Saker, M.L.; Vale, M.; Kramer, D.; Vasconcelos, V.M. Molecular techniques for the early warning of toxic cyanobacteria blooms in fresh water lakes and rivers. Appl. Microbiol. Biotechnol. 2007, 75, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wilhelm, S.W.; Carmichael, W.W.; Watanabe, M.M. Polyphasic characterization of water bloom forming Raphidiopsis species (Cyanobacteria) from central China. Harmful Algae 2008, 7, 146–153. [Google Scholar] [CrossRef]
- Valério, E.; Chambel, L.; Paulino, S.; Faria, N.; Pereira, P.; Tenreiro, R. Molecular identification, typing and traceability of cyanobacteria from freshwater reservoirs. Microbiology 2009, 155, 642–656. [Google Scholar] [CrossRef] [Green Version]
- Kormas, K.A.; Gkelis, S.; Vardaka, E.; Moustaka-Gouni, M. Morphological and molecular analysis of bloom-forming Cyanobacteria in two eutrophic, shallow Mediterranean lakes. Limnologica 2010, 41, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Lymperopoulou, D.S.; Kormas, K.A.; Moustaka-Gouni, M.; Karagouni, A.D. Diversity of cyanobacterial phylotypes in a Mediterranean drinking water reservoir (Marathonas, Greece). Environ. Monit. Assess. 2011, 173, 155–165. [Google Scholar] [CrossRef]
- Kobos, J.; Błaszczyk, A.; Hohlfeld, N.; Toruńska-Sitarz, A.; Krakowiak, A.; Hebel, A.; Sutryk, K.; Grabowska, M.; Toporowska, M.; Kokociński, M.; et al. Cyanobacteria and cyanotoxins in Polish freshwater bodies. Oceanol. Hydrobiol. Stud. 2013, 42, 358–378. [Google Scholar] [CrossRef] [Green Version]
- Rott, E.; Pentecost, A.; Mareš, J. Introduction: Recent developments in cyanobacterial research with special reference to aquatic habitats, molecular ecology and phylogenetic taxonomy. Hydrobiologia 2018, 811, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Ramos, V.M.C.; Castelo-Branco, R.; Leão, P.N.; Martins, J.; Carvalhal-Gomes, S.; Sobrinho da Silva, F.; Mendonça Filho, J.G.; Vasconcelos, V.M. Cyanobacterial diversity in microbial mats from the hypersaline lagoon system of Araruama, Brazil: An In-depth polyphasic study. Front. Microbiol. 2017, 8, 1233. [Google Scholar] [CrossRef] [Green Version]
- Humbert, J.-F. Molecular Tools for the Detection of Toxigenic Cyanobacteria in Natural Ecosystems. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, J., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 280–283. [Google Scholar] [CrossRef]
- Catherine, A.; Bernard, C.; Spoof, L.; Bruno, M. Microcystins and nodularins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, J., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 109–126. [Google Scholar] [CrossRef]
- Moffitt, M.C.; Neilan, B.A. Characterization of the nodularin synthetase gene cluster and proposed theory of the evolution of cyanobacterial hepatotoxins. Appl. Environ. Microb. 2004, 70, 6353–6362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, L.; Mihali, T.; Moffitt, M.; Kellmann, R.; Neilan, B. On the chemistry, toxicology and genetics of the cyanobacterial toxins, microcystin, nodularin, saxitoxin and cylindrospermopsin. Mar. Drugs 2010, 8, 1650–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouellette, A.J.A.; Wilhelm, S.W. Toxic cyanobacteria: The evolving molecular toolbox. Front. Ecol. Environ. 2003, 1, 359–366. [Google Scholar] [CrossRef]
- Mbukwa, E.A.; Boussiba, S.; Wepener, V.; Leu, S.; Kaye, Y.; Msagati, T.A.M.; Mamba, B.B. PCR amplification and DNA sequence of mcyA gene: The distribution profile of a toxigenic Microcystis aeruginosa in the Hartbeespoort Dam, South Africa. J. Water Health 2013, 11, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Meriluoto, J.; Blaha, L.; Bojadzija, G.; Bormans, M.; Brient, L.; Codd, G.A.; Drobac, D.; Faassen, E.J.; Fastner, J.; Anastasia, H.; et al. 2017. Toxic cyanobacteria and cyanotoxins in European waters-recent progress achieved through the CYANOCOST Action and challenges for further research. Adv. Oceanogr. Limnol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Ilieva, V.; Kondeva-Burdina, M.; Pavlova, V.; Bratanova, Z.; Danchev, N.; Georgieva, T.Z. Real-time PCR monitoring of toxic Cyanobacteria in Bulgarian water samples. Proceedings of Annual University Scientific Conference, Vasil Levski National Military University, Veliko Turnovo, Bulgaria, 20–21 October 2016; Publishing Complex of Vasil Levski National Military University: Veliko Turnovo, Bulgaria, 2016; pp. 241–249. [Google Scholar]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baurès, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef]
- Ibelings, B.W.; Backerb, L.C.; Kardinaal, W.E.A.; Chorus, I. Current approaches to cyanotoxin risk assessment and risk management around the globe. Harmful Algae 2015, 49, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Michev, T.; Stoyneva, M. (Eds.) Inventory of Bulgarian Wetlands and Their Biodiversity; Elsi-M: Sofia, Bulgaria, 2007. [Google Scholar]
- Geitler, L. Cyanophyceae. In Kryptogamen-Flora von Deutschland, Österreich und der Schweiz, 2nd ed.; Rabenhorst, L., Ed.; Akademische Verlagsgesellschaft: Leipzig, Germany, 1931; Volume 14, pp. 289–672. [Google Scholar]
- Geitler, L. Schizophyta: Klasse Schizophyceae. In Die Natürlichen Pflanzenfamilien, Sweite Auflage; Engler, A., Prantl, K., Eds.; Wilhelm Engelmann: Leipzig, Germany, 1942; Volume 1b, pp. 1–232. [Google Scholar]
- Gollerbakh, M.M.; Kossinskaya, E.K.; Polyanskiy, V.I. Manual of Freshwater Algae of the USSR. Blue-Green Algae; Sovetskaya Nauka: Moscow, Russia, 1953; Volume 2. [Google Scholar]
- Starmach, K. Cyanophyta-Sinice, Glaucophyta-Glaukofity; Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1966. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota. Teil 1/Part 1: Chroococcales. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Spektrum Akademischer Verlag: Heidelberg, Germany, 2008; Volume 19/1, pp. 1–556. [Google Scholar]
- Komárek, J.; Komárková, J. Review of the European Microcystis morphospecies (Cyanoprokaryotes) from nature. Czech Phycol. 2002, 2, 1–24. [Google Scholar]
- Šejnohová, L.; Maršálek, B. Microcystis. In Ecology of Cyanobacteria II. Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 195–228. [Google Scholar]
- Basic Local Alignment Search Tool (BLAST). Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 24 October 2019).
- National Centre for Biotechnology Information (NCBI). Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/ (accessed on 24 October 2019).
- Tang, X.; Krausfeldt, L.E.; Shao, K.; LeCleir, G.R.; Stough, J.M.A.; Gao, G.; Boyer, G.L.; Zhang, Y.; Paerl, H.W.; Qin, B.; et al. Seasonal Gene Expression and the ecophysiological implications of toxic Microcystis aeruginosa blooms in Lake Taihu. Environ. Sci. Technol. 2018, 52, 11049–11059. [Google Scholar] [CrossRef]
- Vasconcelos, V. Global changes and the new challenges in the research on cyanotoxin risk evaluation. Limnetica 2015, 34, 149–158. [Google Scholar] [CrossRef]
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A review of the global ecology, genomics, and biogeography of the toxic cyanobacterium, Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Manach, S.; Duval, C.; Marie, A.; Djediat, C.; Catherine, A.; Edery, M.; Bernard, C.; Marie, B. Global metabolomic characterizations of Microcystis spp. Highlights clonal diversity in natural bloom-forming populations and expands metabolite structural diversity. Front. Microbiol. 2019, 10, 791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlova, V.; Babica, P.; Todorova, D.; Bratanova, Z.; Maršalek, B. Contamination of some reservoirs and lakes in Republic of Bulgaria by microcystins. Acta Hydrochim. Hydrobiol. 2006, 34, 437–441. [Google Scholar] [CrossRef]
- Stoyneva, M. Contribution to the Studies of the Biodiversity of Hydro- and Aerobiontic Prokaryotic and Eukaryotic Algae in Bulgaria. Ph.D. Thesis, Sofia University “St. Kliment Ohridski”, Sofia, Bulgaria, 2014. [Google Scholar]
- Alvarenga, D.O.; Fiore, M.F.; Varani, A.M. A metagenomic approach to cyanobacterial genomics. Front. Microbiol. 2017, 8, 809. [Google Scholar] [CrossRef] [Green Version]
- Dvořák, P.; Jahodářová, E.; Casamatta, D.A.; Hašler, P.; Poulíčková, A. Difference without distinction? Gaps in cyanobacterial systematics; when more is just too much. Fottea 2018, 18, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Meyer, K.A.; Davis, T.W.; Watson, S.B.; Denef, V.J.; Berry, M.A.; Dick, G.J. Genome sequences of lower Great Lakes Microcystis sp. reveal strain-specific genes that are present and expressed in western Lake Erie blooms. PLoS ONE 2017, 12, e0183859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Tan, W.; Wu, X.; Wu, Z.; Yu, G.; Li, R. First report of microcystin production in Microcystis smithii Komárek and Anagnostidis (Cyanobacteria) from a water bloom in Eastern China. J. Environ. Sci. 2011, 23, 102–107. [Google Scholar] [CrossRef]
- Otsuka, S.; Suda, S.; Li, R.H.; Matsumoto, S.; Watanabe, M.M. Morphological variability of colonies of Microcystis morphospecies in culture. J. Gen. Appl. Microbiol. 2000, 46, 39–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otsuka, S.; Suda, S.; Renhui, L.; Watanabe, M.; Oyaizu, H.; Matsumoto, S.; Watanabe, M.M. 16S rDNA sequences and phylogenetic analyses of Microcystis strains with and without phycoerythrin. FEMS Microbiol. Lett. 1998, 164, 119–124. [Google Scholar] [CrossRef]
- Otsuka, S.; Suda, S.; Shibata, S.; Oyaizu, H.; Matsumoto, S.; Watanabe, M.M. A proposal for the unification of five species of the cyanobacterial genus Microcystis Kützing ex Lemmermann 1907 under the rules of the bacteriological code. Int. J. Syst. Evol. Microbiol. 2001, 51, 873–879. [Google Scholar] [CrossRef] [Green Version]
- Sanchis, D.; Padilla, C.; Del Campo, F.F.; Quesada, A.; Sanz-Alferez, S. Phylogenetic and morphological analyses of Microcystis strains (Cyanophyta/Cyanobacteria) from a Spanish water reservoir. Nova Hedwig. 2005, 81, 431–448. [Google Scholar] [CrossRef]
- Bernard, C.; Ballot, A.; Thomazeau, S.; Maloufi, S.; Furey, A.; Mankiewicz-Boczek, I.; Pawlik-Skowrońska, B.; Capelli, C.; Salmazo, N. Appendix 2. Cyanobacteria associated with the production of cyanotoxins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, J., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 501–525. [Google Scholar] [CrossRef] [Green Version]
- Kurmayer, R.; Dittmann, E.; Fastner, J.; Chorus, I. Diversity of microcystin genes within a population of the toxic cyanobacterium Microcystis spp. in Lake Wannsee (Berlin, Germany). Microb. Ecol. 2002, 43, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Via-Ordorika, L.; Fastner, J.; Kurmayer, R.; Hisbergues, M.; Dittmann, E.; Komárek, J.; Erhard, M.; Chorus, I. Distribution of microcystin-producing and non-microcystin-producing Microcystis sp. in European freshwater bodies: Detection of microcystins and microcystin genes in individual colonies. Syst. Appl. Microbiol. 2004, 27, 592–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlova, V.; Stoyneva, M.; Georgieva, V.; Donchev, D.; Spoof, L.; Meriluoto, J.; Bratanova, Z.; Karadjova, I. New records of microcystins in some Bulgarian water bodies of health and conservational importance. J. Water Res. Protect. 2014, 6, 446–453. [Google Scholar] [CrossRef] [Green Version]
- Komárek, J. Cyanoprokaryota. 3. Heterocytous genera. In Süswasserflora von Mitteleuropa/Freshwater Flora of Central Europe; Büdel, B., Gärtner, G., Krienitz, L., Schagerl, M., Eds.; Springer Spektrum: Berlin, Germany, 2013; pp. 1–1130. [Google Scholar]
- AlgaeBase. Available online: http://www.algaebase.org/ (accessed on 18 September 2019).
- CyanoDB 2.0. Available online: http://www.cyanodb.cz (accessed on 19 September 2019).
- Rott, E. Some results from phytoplankton counting intercalibration. Schweiz. Z. Hydrol. 1981, 43, 34–62. [Google Scholar] [CrossRef]
- Valério, E.; Chambel, L.; Paulino, S.; Faria, N.; Pereira, P.; Tenreiro, R. Multiplex PCR for detection of microcystins-producing cyanobacteria from freshwater samples. Environ. Toxicol. 2010, 25, 251–260. [Google Scholar] [CrossRef]
- Vaitomaa, J.; Rantala, A.; Halinen, K.; Rouhiainen, L.; Tallberg, P.; Mokelke, L.; Sivonen, K. Quantitative real-time PCR for determination of microcystin synthetase E copy numbers for Microcystis and Anabaena in lakes. Appl. Environ. Microbiol. 2003, 69, 7289–7297. [Google Scholar] [CrossRef] [Green Version]
- Jungblut, A.-D.; Neilan, B.A. Molecular identification and evolution of the cyclic peptide hepatotoxins, microcystin and nodularin synthetase genes in three orders of cyanobacteria. Arch. Microbiol. 2006, 185, 107–114. [Google Scholar] [CrossRef]
- Rantala, A.; Rajaniemi-Wacklin, P.; Lyra, C.; Lepistö, L.; Rintala, J.; Mankiewicz-Boczek, J.; Sivonen, K. Detection of microcystin-producing cyanobacteria in Finnish lakes with genus-specific microcystin synthetase gene E (mcyE) PCR and associations with environmental factors. Appl. Environ. Microb. 2006, 72, 6101–6110. [Google Scholar] [CrossRef] [Green Version]
- Weller, D.I. Detection, identification and toxigenicity of cyanobacteria in New Zealand lakes using PCR-based methods. N. Z. J. Mar. Freshw. Res. 2011, 45, 651–664. [Google Scholar] [CrossRef]
- Conradie, K.R.; Barnard, S. The dynamics of toxic Microcystis strains and microcystin production in two hypertrofic South African reservoirs. Harmful Algae 2012, 20, 1–10. [Google Scholar] [CrossRef]
- Ngwa, F.; Madramootoo, C.A.; Jabaj, S. Comparison of cyanobacterial microcystin synthetase (mcy) E gene transcript levels, mcy E gene copies, and biomass as indicators of microcystin risk under laboratory and field conditions. Microbiol. Open 2014, 3, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Rantala, A. Evolution and Detection of Cyanobacterial Hepatotoxin Synthetase Genes. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 27 November 2007. [Google Scholar]
- Rantala, A.; Rizzi, E.; Castiglioni, B.; de Bellis, G.; Sivonen, K. Identification of hepatotoxin-producing cyanobacteria by DNA-chip. Environ. Microbiol. 2008, 10, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.K.; Das, P.K.; Bagchi, S.N. PCR-based microcystin-producing cyanobacterial blooms from Central India. Indian J. Exp. Biol. 2008, 46, 66–70. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WBN and IBW | SAN | MA | MB | MF | MNs | MNv | MS | MW | SS | SL | TCs |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Reservoir Sinyata Reka—SR (IBW1890) | SR1 | d | x | x | |||||||
| SR2 | d | x | x | ||||||||
| Vaya Lake—VA (IBW0191) | VA1 | x | r | x | x | x | |||||
| VA2 | x | r | r | x | x | x | |||||
| VA3 | x | r | x | ||||||||
| Reservoir Mandra—MN (IBW1720) | MN1 | x | |||||||||
| MN3 | x | x | |||||||||
| Reservoir Poroy (IBW3038) | x | x | r | ||||||||
| Durankulak Lake—DR (IBW0216) | DR1 | x | x | x | x | ||||||
| DR2 | x | r | r | r | r | x | |||||
| DR3 | x | r | r | x | |||||||
| DR4 | x | r | x |
| WBN and IBW | SAN | Date | Alt | Latitude | Longitude | WT | pH | SD | CND | TDS | DO | TP | TN | CT |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Res. Sinyata Reka (IBW1890) | SR1 | 21.06.18 | 317 | 42°28.1480’ | 24°42.217 | 27.4 | 9.72 | 0.5 | 470 | 305 | 9.36 | 25 | 4.8 | MCs |
| SR2 | 21.06.18 | 317 | 42°28.1473’ | 24°42.2175 | 26.7 | 9.36 | 0.6 | 468 | 306 | 9,21 | 27 | 4.3 | ||
| L. Vaya (IBW0191) | VA1 | 22.06.18 | −2 | 42°30.5940’ | 27°22.075 | 26.9 | 9.65 | 0.25 | 2588 | 1682 | 12.51 | 13 | 5,4 | CYN |
| VA2 | 22.06.18 | 0 | 42°28.4540’ | 27°25.482 | 28.28 | 8.86 | 0.25 | 1183 | 768 | 11.94 | 11 | 3.7 | ||
| VA3 | 23.06.18 | 6 | 42°29.1850’ | 27°26.531 | 23.7 | 9.5 | 0.25 | 1024 | 665 | 7.01 | 12 | 4.6 | ||
| Res. Mandra (IBW1720) | MN1 | 23.06.18 | 12 | 42°24.0463’ | 27°26.1120’ | 25.88 | 8.28 | 0.4 | 649 | 421 | 6.81 | 3 | 3.0 | |
| MN2 | 23.06.18 | 13 | 42°24.0670’ | 27°19.1310’ | 26.2 | 8.2 | 0.2 | 663 | 461 | 5.89 | 6 | 4.0 | CYN | |
| MN3 | 23.06.18 | 9 | 42 26.1420’ | 27°26.5860’ | 24.9 | 8.48 | 0.3 | 639 | 415 | 7.91 | 4 | 3.3 | ||
| L. Uzungeren (IBW0710) | UZ | 23.06.18 | 7 | 42°26.1782’ | 27°27.1998’ | 25.9 | 8.06 | 0.4 | 14.38 | 9351 | 7.83 | 5 | 2.8 | |
| Res. Poroy (IBW3038) | PR | 24.06.18 | 41 | 42°43.0190’ | 27°37.3160’ | 25.10 | 8.33 | 1.2 | 762 | 495 | 9.45 | 1 | 2.8 | |
| Res. Aheloy (IBW3032) | AH | 24.06.18 | 144 | 42°42.8230’ | 27°30.9740’ | 25.4 | 8.51 | 1.10 | 614 | 399 | 8.92 | 1 | 4.1 | |
| L. Ezerets (IBW0233) | EZ | 25.06.18 | −2 | 43°35.2770’ | 28°33.2290’ | 26.4 | 8.35 | TTB | 1084 | 0 | 9.94 | 0.5 | 5.3 | |
| L. Shabla (IBW0219) | SH | 25.06.18 | −2 | 43°33.8180’ | 28°34.1860’ | 27.1 | 8.46 | TTB | 1087 | 0706 | 9.97 | 0.1 | 5.1 | |
| L. Durankulak (IBW0216) | DR1 | 25.06.18 | 6 | 43°40.3240’ | 28°32.0470’ | 24.03 | 8.54 | 1 | 1111 | 722 | 7.35 | 21 | 2.8 | |
| DR2 | 25.06.18 | 6 | 43°40.3340’ | 28°32.0220’ | 24.7 | 8.21 | 1 | 1094 | 711 | 7.79 | 20 | 4.0 | MCs | |
| DR3 | 25.06.18 | 4 | 43°40.5300’ | 28°32.9930’ | 24.6 | 8.49 | 1 | 1075 | 698 | 6.19 | 24 | 3.9 | MCs, SXT | |
| DR4 | 25.06.18 | 3 | 43°40.6950’ | 28°32.6000’ | 26.5 | 8.53 | 1 | 1087 | 706 | 9.6 | 20 | 3.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radkova, M.; Stefanova, K.; Uzunov, B.; Gärtner, G.; Stoyneva-Gärtner, M. Morphological and Molecular Identification of Microcystin-Producing Cyanobacteria in Nine Shallow Bulgarian Water Bodies. Toxins 2020, 12, 39. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12010039
Radkova M, Stefanova K, Uzunov B, Gärtner G, Stoyneva-Gärtner M. Morphological and Molecular Identification of Microcystin-Producing Cyanobacteria in Nine Shallow Bulgarian Water Bodies. Toxins. 2020; 12(1):39. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12010039
Chicago/Turabian StyleRadkova, Mariana, Katerina Stefanova, Blagoy Uzunov, Georg Gärtner, and Maya Stoyneva-Gärtner. 2020. "Morphological and Molecular Identification of Microcystin-Producing Cyanobacteria in Nine Shallow Bulgarian Water Bodies" Toxins 12, no. 1: 39. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12010039