Proteomic Investigations of Two Pakistani Naja Snake Venoms Species Unravel the Venom Complexity, Posttranslational Modifications, and Presence of Extracellular Vesicles

 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Results
3. Discussion
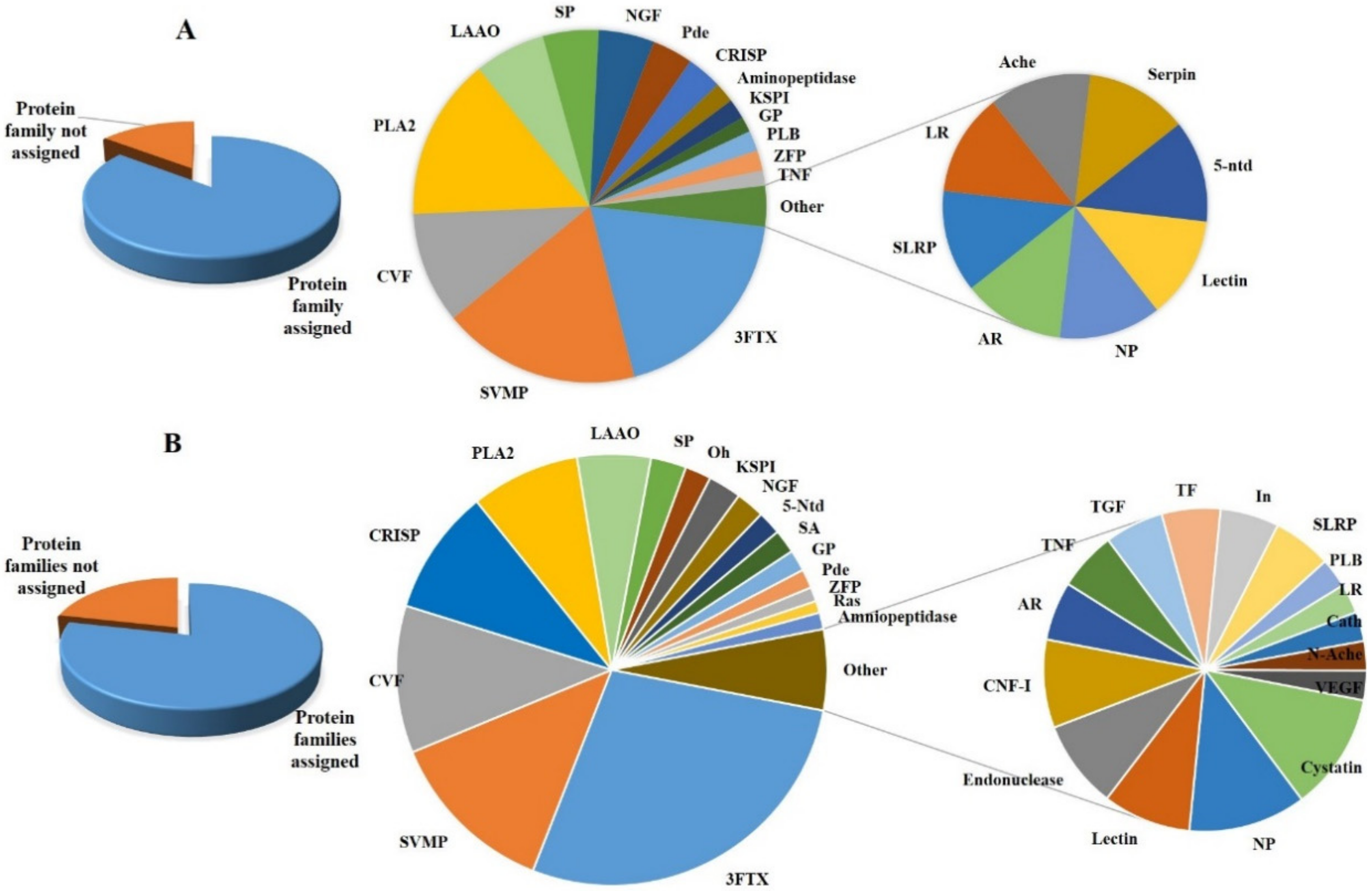
3.1. Major Protein Components (Relative Abundance >2%)
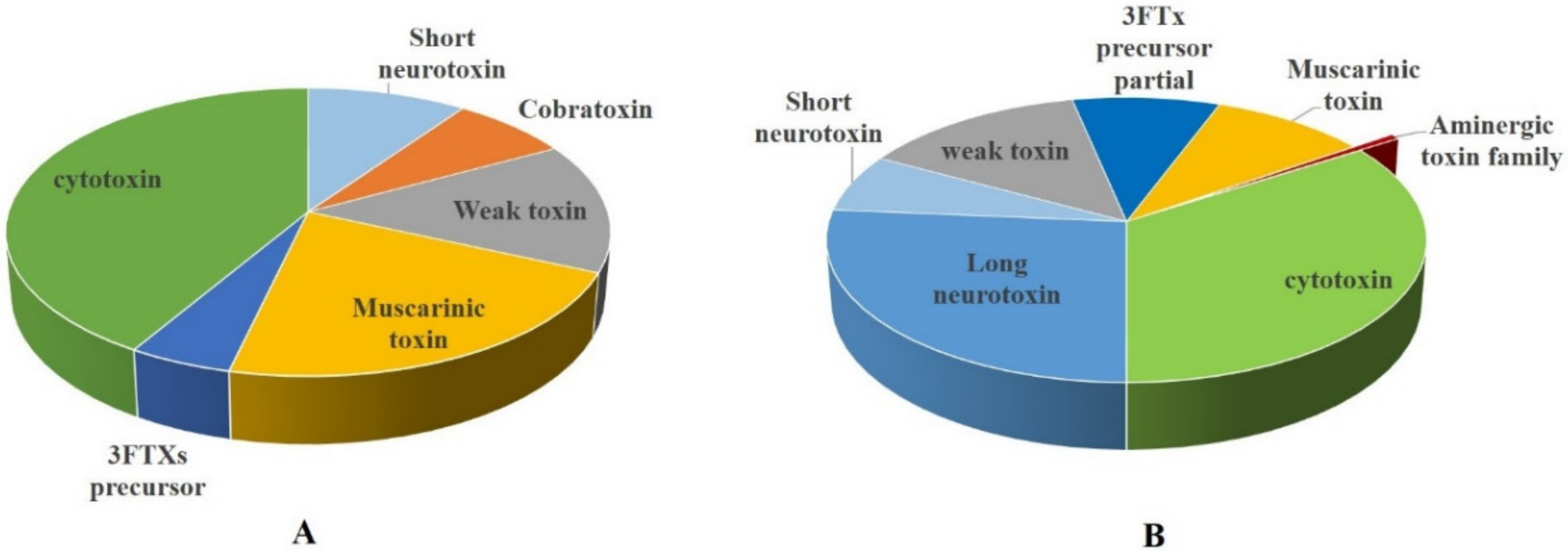
3.1.1. Three-Finger Toxins
3.1.2. Phospholipase A2
3.1.3. Snake Venom Metalloproteinase
3.1.4. L-Amino Acid Oxidase
3.1.5. Cobra Venom Factor
3.1.6. Cysteine-Rich Secretory Protein
3.1.7. Snake Venom Serine Proteinase
3.1.8. Snake Venom Nerve Growth Factor
3.1.9. Snake Venom Phosphodiesterase
3.2. Minor Protein Components (Relative Abundance ≤2%)
3.3. Posttranslational Modifications
4. Conclusions
5. Materials and Methods
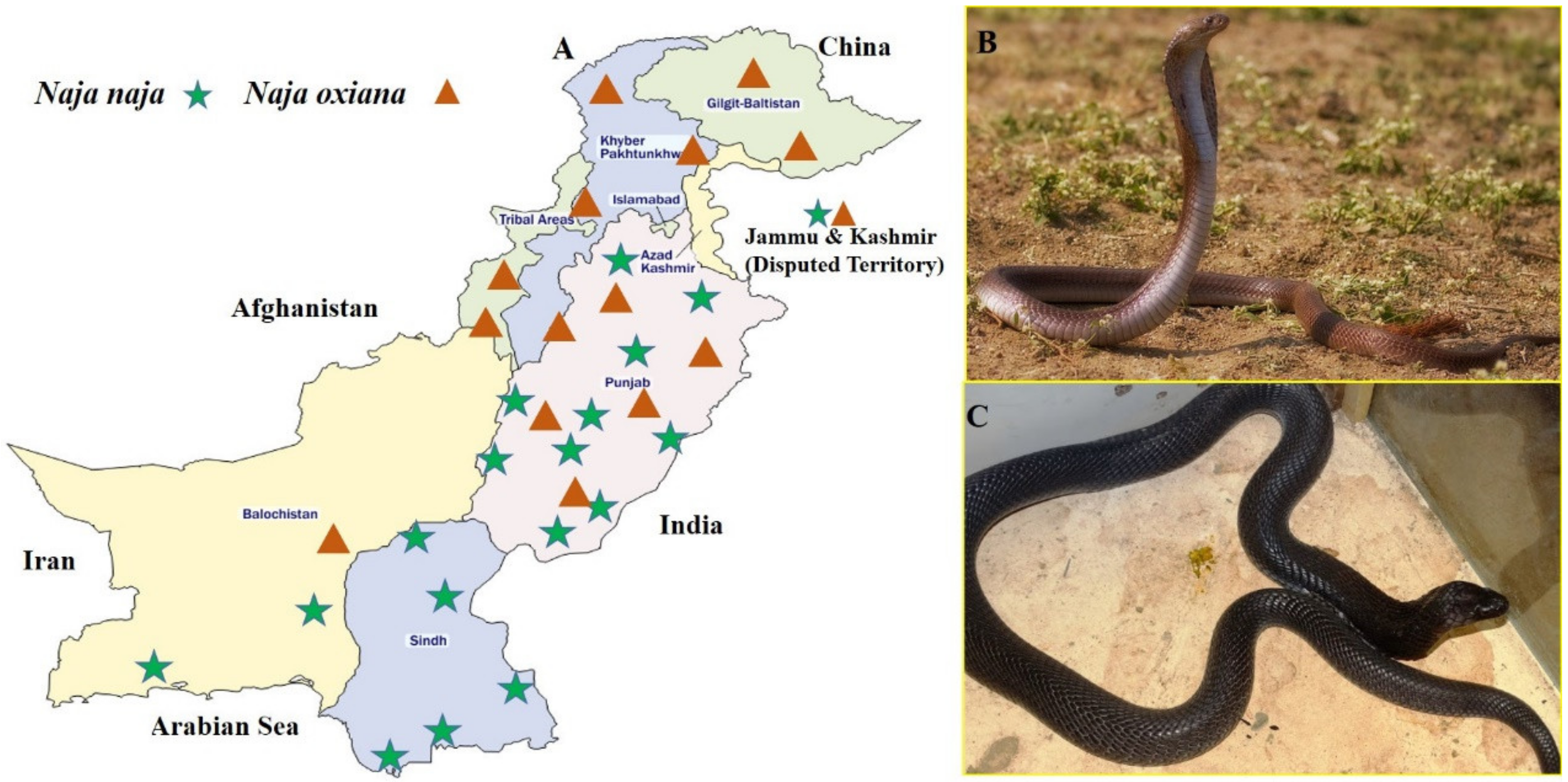
5.1. Venom Collection
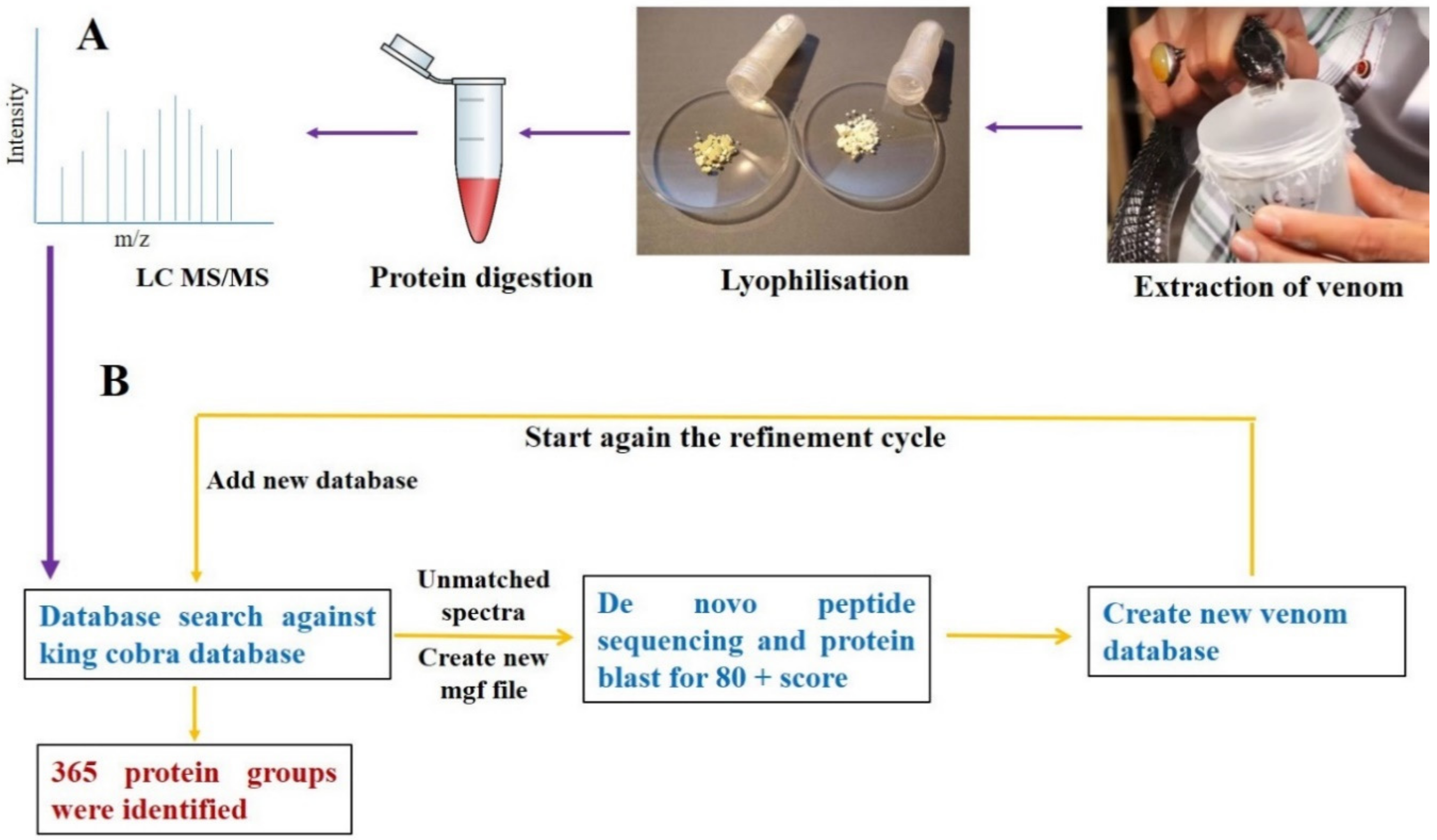
5.2. Sample Preparation for LC-MS/MS
5.3. LC-MS/MS Analysis of the Digested Venom
5.4. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ali, W.; Javaid, A.; Hussain, A.; Bukhari, M.S. Diversity and habitat preferences of amphibians and reptiles in pakistan: A review. J. Asia Pac. Biodivers. 2018, 11, 173–187. [Google Scholar] [CrossRef]
- Khan, M.S. A guide to the snakes of pakistan. In Frankfurt Contributions to Natural History; Chimaira: Frankfurt, Germany, 2002; Volume 16. [Google Scholar]
- Wallach, V.; Kenneth, L.W.; Boundy, J. Snakes of the World. A Catalogue of Living and Extinct Species; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Panagides, N.; Jackson, T.N.; Ikonomopoulou, M.P.; Arbuckle, K.; Pretzler, R.; Yang, D.C.; Ali, S.A.; Koludarov, I.; Dobson, J.; Sanker, B.; et al. How the cobra got its flesh-eating venom: Cytotoxicity as a defensive innovation and its co-evolution with hooding, aposematic marking, and spitting. Toxins 2017, 9, 103. [Google Scholar] [CrossRef] [Green Version]
- Wuster, W. The cobras of the genus naja in india. Hamadryad 1998, 23, 15–32. [Google Scholar]
- Wuster, W.; Thorpe, R.S. Asiatic cobras: Population systematics of the naja naja species complex (serpentes: Elapidae) in india and central asia. Herpetologica 1992, 48, 69–85. [Google Scholar]
- Gutierrez, J.M.; Williams, D.; Fan, H.W.; Warrell, D.A. Snakebite envenoming from a global perspective: Towards an integrated approach. Toxicon 2010, 56, 1223–1235. [Google Scholar] [CrossRef]
- Wong, K.Y.; Tan, C.H.; Tan, N.H. Venom and purified toxins of the spectacled cobra (naja naja) from pakistan: Insights into toxicity and antivenom neutralization. Am. J. Trop Med. Hyg. 2016, 94, 1392–1399. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Rabies and Envenomings-A Neglected Public Health Issue; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Gutierrez, J.M.; Burnouf, T.; Harrison, R.A.; Calvete, J.J.; Kuch, U.; Warrell, D.A.; Williams, D.J.; Global Snakebite, I. A multicomponent strategy to improve the availability of antivenom for treating snakebite envenoming. Bull. World Health Organ. 2014, 92, 526–532. [Google Scholar] [CrossRef]
- Scheske, L.; Ruitenberg, J.; Bissumbhar, B. Needs and availability of snake antivenoms: Relevance and application of international guidelines. Int. J. Health Policy Manag. 2015, 4, 447–457. [Google Scholar] [CrossRef]
- Khan, M.S. The snakebite problem in pakistan. Bull. Chic. Herp. Soc. 2014, 49, 165–167. [Google Scholar]
- Khan, R. Treating snakebites, one snake at a time. Express Trib. 2019. Available online: https://tribune.com.pk/story/1926889/treating-snakebites-one-snake-time (accessed on 11 March 2019).
- Ralph, R.; Sharma, S.K.; Faiz, M.A.; Ribeiro, I.; Rijal, S.; Chappuis, F.; Kuch, U. The timing is right to end snakebite deaths in south asia. BMJ 2019, 364, k5317. [Google Scholar] [CrossRef] [Green Version]
- Faisal, T.; Tan, K.Y.; Sim, S.M.; Quraishi, N.; Tan, N.H.; Tan, C.H. Proteomics, functional characterization and antivenom neutralization of the venom of pakistani russell’s viper (daboia russelii) from the wild. J. Proteom. 2018, 183, 1–13. [Google Scholar] [CrossRef]
- Hashmi, S.U.; Alvi, A.; Munir, I.; Perveen, M.; Fazal, A.; Jackson, T.N.W.; Ali, S.A. Functional venomics of the big-4 snakes of pakistan. Toxicon 2020, 179, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Mackessy, S.P. The field of reptile toxinology: Snakes, lizards, and their venoms. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Taylor and Francis Group, Eds.; CRC Press: Boca Raton, FL, USA, 2009; pp. 3–23. [Google Scholar]
- Daltry, J.C.; Ponnudurai, G.; Shin, C.K.; Tan, N.H.; Thorpe, R.S.; Wuster, W. Electrophoretic profiles and biological activities: Intraspecific variation in the venom of the malayan pit viper (calloselasma rhodostoma). Toxicon 1996, 34, 67–79. [Google Scholar] [CrossRef]
- Alape-Giron, A.; Sanz, L.; Escolano, J.; Flores-Diaz, M.; Madrigal, M.; Sasa, M.; Calvete, J.J. Snake venomics of the lancehead pitviper bothrops asper: Geographic, individual, and ontogenetic variations. J. Proteome Res. 2008, 7, 3556–3571. [Google Scholar] [CrossRef]
- Tan, K.Y.; Tan, C.H.; Fung, S.Y.; Tan, N.H. Venomics, lethality and neutralization of naja kaouthia (monocled cobra) venoms from three different geographical regions of southeast Asia. J. Proteom. 2015, 120, 105–125. [Google Scholar] [CrossRef]
- Amorim, F.G.; Costa, T.R.; Baiwir, D.; De Pauw, E.; Quinton, L.; Sampaio, S.V. Proteopeptidomic, functional and immunoreactivity characterization of bothrops moojeni snake venom: Influence of snake gender on venom composition. Toxins 2018, 10, 177. [Google Scholar] [CrossRef] [Green Version]
- Augusto-de-Oliveira, C.; Stuginski, D.R.; Kitano, E.S.; Andrade-Silva, D.; Liberato, T.; Fukushima, I.; Serrano, S.M.; Zelanis, A. Dynamic rearrangement in snake venom gland proteome: Insights into bothrops jararaca intraspecific venom variation. J. Proteome Res. 2016, 15, 3752–3762. [Google Scholar] [CrossRef] [PubMed]
- Rex, C.J.; Mackessy, S.P. Venom composition of adult western diamondback rattlesnakes (crotalus atrox) maintained under controlled diet and environmental conditions shows only minor changes. Toxicon 2019, 164, 51–60. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Maity, C.R. The composition of naja naja venom samples from three districts of west bengal, india. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 1998, 119, 621–627. [Google Scholar] [CrossRef]
- Quraishi, N.A.; Qureshi, H.I.; Simpson, I.D. A contextual approach to managing snake bite in pakistan: Snake bite treatment with particular reference to neurotoxieity and the ideal hospital snake bite kit. J. Pak. Med. Assoc. 2008, 58, 325–331. [Google Scholar]
- Asad, M.H.; Murtaza, G.; Ubaid, M.; Durre, S.; Sajjad, A.; Mehmood, R.; Mahmood, Q.; Ansari, M.M.; Karim, S.; Mehmood, Z.; et al. Naja naja karachiensis envenomation: Biochemical parameters for cardiac, liver, and renal damage along with their neutralization by medicinal plants. BioMed Res. Int. 2014, 2014, 970540. [Google Scholar] [CrossRef]
- Lomonte, B.; Calvete, J.J. Strategies in ‘snake venomics’ aiming at an integrative view of compositional, functional, and immunological characteristics of venoms. J. Venom. Anim Toxins Incl. Trop. Dis. 2017, 23, 26. [Google Scholar] [CrossRef] [Green Version]
- Vonk, F.J.; Casewell, N.R.; Henkel, C.V.; Heimberg, A.M.; Jansen, H.J.; McCleary, R.J.; Kerkkamp, H.M.; Vos, R.A.; Guerreiro, I.; Calvete, J.J.; et al. The king cobra genome reveals dynamic gene evolution and adaptation in the snake venom system. Proc. Natl. Acad. Sci. USA 2013, 110, 20651–20656. [Google Scholar] [CrossRef] [Green Version]
- Von Reumont, B.M. Studying smaller and neglected organisms in modern evolutionary venomics implementing rnaseq (transcriptomics)-a critical guide. Toxins 2018, 10, 292. [Google Scholar] [CrossRef] [Green Version]
- Fry, B.G. From genome to “venome”: Molecular origin and evolution of the snake venom proteome inferred from phylogenetic analysis of toxin sequences and related body proteins. Genome Res. 2005, 15, 403–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerkkamp, H.M.; Kini, R.M.; Pospelov, A.S.; Vonk, F.J.; Henkel, C.V.; Richardson, M.K. Snake genome sequencing: Results and future prospects. Toxins 2016, 8, 360. [Google Scholar] [CrossRef] [Green Version]
- Wiezel, G.A.; Shibao, P.Y.T.; Cologna, C.T.; Morandi Filho, R.; Ueira-Vieira, C.; De Pauw, E.; Quinton, L.; Arantes, E.C. In-depth venome of the brazilian rattlesnake crotalus durissus terrificus: An integrative approach combining its venom gland transcriptome and venom proteome. J. Proteom. Res. 2018, 17, 3941–3958. [Google Scholar] [CrossRef]
- Rokyta, D.R.; Margres, M.J.; Calvin, K. Post-transcriptional mechanisms contribute little to phenotypic variation in snake venoms. G3 Bethesda 2015, 5, 2375–2382. [Google Scholar] [CrossRef] [Green Version]
- Melani, R.D.; Skinner, O.S.; Fornelli, L.; Domont, G.B.; Compton, P.D.; Kelleher, N.L. Mapping proteoforms and protein complexes from king cobra venom using both denaturing and native top-down proteomics. Mol. Cell Proteom. 2016, 15, 2423–2434. [Google Scholar] [CrossRef] [Green Version]
- Munawar, A.; Ali, S.A.; Akrem, A.; Betzel, C. Snake venom peptides: Tools of biodiscovery. Toxins 2018, 10, 474. [Google Scholar] [CrossRef] [Green Version]
- Simoes-Silva, R.; Alfonso, J.; Gomez, A.; Holanda, R.J.; Sobrinho, J.C.; Zaqueo, K.D.; Moreira-Dill, L.S.; Kayano, A.M.; Grabner, F.P.; da Silva, S.L.; et al. Snake venom, a natural library of new potential therapeutic molecules: Challenges and current perspectives. Curr. Pharm. Biotechnol. 2018, 19, 308–335. [Google Scholar] [CrossRef]
- King, G.F. Venoms as a platform for human drugs: Translating toxins into therapeutics. Expert Opin. Biol. 2011, 11, 1469–1484. [Google Scholar] [CrossRef]
- Koh, D.C.; Armugam, A.; Jeyaseelan, K. Snake venom components and their applications in biomedicine. Cell Mol. Life Sci. 2006, 63, 3030–3041. [Google Scholar] [CrossRef]
- Ciolek, J.; Reinfrank, H.; Quinton, L.; Viengchareun, S.; Stura, E.A.; Vera, L.; Sigismeau, S.; Mouillac, B.; Orcel, H.; Peigneur, S.; et al. Green mamba peptide targets type-2 vasopressin receptor against polycystic kidney disease. Proc. Natl. Acad. Sci. USA 2017, 114, 7154–7159. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Ghani, L.M.; Rahmy, T.R.; Tawfik, M.M.; Kaziri, I.; Al-Obaidi, A.; Rowan, E.G.; Plevin, R.; Abdel-Rahman, M.A. Cytotoxicity of nubein6.8 peptide isolated from the snake venom of naja nubiae on melanoma and ovarian carcinoma cell lines. Toxicon 2019, 168, 22–31. [Google Scholar] [CrossRef]
- Wuster, W. Taxonomic changes and toxinology: Systematic revisions of the asiatic cobras (naja naja species complex). Toxicon 1996, 34, 399–406. [Google Scholar] [CrossRef]
- Asad, M.H.H.B.; McCleary, R.J.R.; Salafutdinov, I.; Alam, F.; Shah, H.S.; Bibi, S.; Ali, A.; Khalid, S.; Hasan, S.M.F.; Sabatier, J.M.; et al. Proteomics study of southern punjab pakistani cobra (naja naja: Formerly naja naja karachiensis) venom. Toxicol. Environ. Chem. 2019. Ahead of Print. [Google Scholar] [CrossRef]
- Ali, S.A.; Yang, D.C.; Jackson, T.N.W.; Undheim, E.A.B.; Koludarov, I.; Wood, K.; Jones, A.; Hodgson, W.C.; McCarthy, S.; Ruder, T.; et al. Venom proteomic characterization and relative antivenom neutralization of two medically important pakistani elapid snakes (bungarus sindanus and naja naja). J. Proteom. 2013, 89, 15–23. [Google Scholar] [CrossRef]
- Wong, K.Y.; Tan, C.H.; Tan, K.Y.; Quraishi, N.H.; Tan, N.H. Elucidating the biogeographical variation of the venom of naja naja (spectacled cobra) from pakistan through a venom-decomplexing proteomic study. J. Proteom. 2018, 175, 156–173. [Google Scholar] [CrossRef]
- Chanda, A.; Patra, A.; Kalita, B.; Mukherjee, A.K. Proteomics analysis to compare the venom composition between naja naja and naja kaouthia from the same geographical location of eastern india: Correlation with pathophysiology of envenomation and immunological cross-reactivity towards commercial polyantivenom. Expert Rev. Proteom. 2018, 15, 949–961. [Google Scholar]
- Chanda, A.; Kalita, B.; Patra, A.; Senevirathne, W.; Mukherjee, A.K. Proteomic analysis and antivenomics study of western india naja naja venom: Correlation between venom composition and clinical manifestations of cobra bite in this region. Expert Rev. Proteom. 2019, 16, 171–184. [Google Scholar] [CrossRef]
- Chanda, A.; Mukherjee, A.K. Quantitative proteomics to reveal the composition of southern india spectacled cobra (naja naja) venom and its immunological cross-reactivity towards commercial antivenom. Int. J. Biol. Macromol. 2020, 160, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.H.; Tan, K.Y.; Fung, S.Y.; Tan, N.H. Venom-gland transcriptome and venom proteome of the malaysian king cobra (ophiophagus hannah). Bmc Genom. 2015, 16, 687. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Xu, Y.; Li, Z.; Fan, J.; Yang, Y. Genome-wide mining of microsatellites in king cobra (ophiophagus hannah) and cross-species development of tetranucleotide ssr markers in chinese cobra (naja atra). Mol. Biol. Rep. 2019, 46, 6087–6098. [Google Scholar] [CrossRef]
- Suryamohan, K.; Krishnankutty, S.P.; Guillory, J.; Jevit, M.; Schroder, M.S.; Wu, M.; Kuriakose, B.; Mathew, O.K.; Perumal, R.C.; Koludarov, I.; et al. The indian cobra reference genome and transcriptome enables comprehensive identification of venom toxins. Nat. Genet. 2020, 52, 106–117. [Google Scholar] [CrossRef] [Green Version]
- Kunalan, S.; Othman, I.; Syed Hassan, S.; Hodgson, W.C. Proteomic characterization of two medically important malaysian snake venoms, calloselasma rhodostoma (malayan pit viper) and ophiophagus hannah (king cobra). Toxins 2018, 10, 434. [Google Scholar] [CrossRef] [Green Version]
- Munawar, A.; Zahid, A.; Negm, A.; Akrem, A.; Spencer, P.; Betzel, C. Isolation and characterization of bradykinin potentiating peptides from agkistrodon bilineatus venom. Proteome Sci. 2016, 14, 1. [Google Scholar] [CrossRef] [Green Version]
- Munawar, A.; Trusch, M.; Georgieva, D.; Hildebrand, D.; Kwiatkowski, M.; Behnken, H.; Harder, S.; Arni, R.; Spencer, P.; Schluter, H.; et al. Elapid snake venom analyses show the specificity of the peptide composition at the level of genera naja and notechis. Toxins 2014, 6, 850–868. [Google Scholar] [CrossRef] [Green Version]
- Munawar, A.; Trusch, M.; Georgieva, D.; Spencer, P.; Frochaux, V.; Harder, S.; Arni, R.K.; Duhalov, D.; Genov, N.; Schluter, H.; et al. Venom peptide analysis of vipera ammodytes meridionalis (viperinae) and bothrops jararacussu (crotalinae) demonstrates subfamily-specificity of the peptidome in the family viperidae. Mol. Biosyst. 2011, 7, 3298–3307. [Google Scholar] [CrossRef]
- Moura-da-Silva, A.M.; Almeida, M.T.; Portes-Junior, J.A.; Nicolau, C.A.; Gomes-Neto, F.; Valente, R.H. Processing of snake venom metalloproteinases: Generation of toxin diversity and enzyme inactivation. Toxins 2016, 8, 183. [Google Scholar] [CrossRef]
- Maiga, A.; Mourier, G.; Quinton, L.; Rouget, C.; Gales, C.; Denis, C.; Lluel, P.; Senard, J.M.; Palea, S.; Servent, D.; et al. G protein-coupled receptors, an unexploited animal toxin targets: Exploration of green mamba venom for novel drug candidates active against adrenoceptors. Toxicon 2012, 59, 487–496. [Google Scholar] [CrossRef]
- Blanchet, G.; Collet, G.; Mourier, G.; Gilles, N.; Fruchart-Gaillard, C.; Marcon, E.; Servent, D. Polypharmacology profiles and phylogenetic analysis of three-finger toxins from mamba venom: Case of aminergic toxins. Biochimie 2014, 103, 109–117. [Google Scholar] [CrossRef]
- Tasoulis, T.; Isbister, G.K. A review and database of snake venom proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef] [Green Version]
- Tsetlin, V.I.; Karlsson, E.; Utkin Yu, N.; Pluzhnikov, K.A.; Arseniev, A.S.; Surin, A.M.; Kondakov, V.V.; Bystrov, V.F.; Ivanov, V.T.; Ovchinnikov Yu, A. Interaction surfaces of neurotoxins and acetylcholine receptor. Toxicon 1982, 20, 83–93. [Google Scholar] [CrossRef]
- Kreienkamp, H.J.; Weise, C.; Raba, R.; Aaviksaar, A.; Hucho, F. Anionic subsites of the catalytic center of acetylcholinesterase from torpedo and from cobra venom. Proc. Natl. Acad Sci. USA 1991, 88, 6117–6121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hucho, F.; Weise, C.; Kreienkamp, H.J.; Tsetlin, V.; Utkin, Y.; Machold, J. Mapping the functional topography of a receptor. Bioorg. Khim 1992, 18, 1319–1329. [Google Scholar] [PubMed]
- Nickitenko, A.V.; Michailov, A.M.; Betzel, C.; Wilson, K.S. Three-dimensional structure of neurotoxin-1 from naja naja oxiana venom at 1.9 a resolution. Febs Lett. 1993, 320, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Kini, R.M.; Koh, C.Y. Snake venom three-finger toxins and their potential in drug development targeting cardiovascular diseases. Biochem. Pharm. 2020, 114105. [Google Scholar] [CrossRef]
- Chang, C.C.; Lee, C.Y. Isolation of neurotoxins from the venom of bungarus multicinctus and their modes of neuromuscular blocking action. Arch. Int. Pharm. 1963, 144, 241–257. [Google Scholar]
- Changeux, J.P.; Kasai, M.; Lee, C.Y. Use of a snake venom toxin to characterize the cholinergic receptor protein. Proc. Natl. Acad. Sci. USA 1970, 67, 1241–1247. [Google Scholar] [CrossRef] [Green Version]
- Jerusalinsky, D.; Kornisiuk, E.; Alfaro, P.; Quillfeldt, J.; Ferreira, A.; Rial, V.E.; Duran, R.; Cervenansky, C. Muscarinic toxins: Novel pharmacological tools for the muscarinic cholinergic system. Toxicon 2000, 38, 747–761. [Google Scholar] [CrossRef]
- Chu, N.S. Contribution of a snake venom toxin to myasthenia gravis: The discovery of alpha-bungarotoxin in taiwan. J. Hist. Neurosci. 2005, 14, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.L.; Kou, J.Q.; Wang, S.Z.; Chen, C.X.; Qin, Z.H. Neurotoxin from naja naja atra venom inhibits skin allograft rejection in rats. Int. Immunopharmacol. 2015, 28, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Kryukova, E.V.; Shelukhina, I.V.; Kolacheva, A.A.; Alieva, A.K.; Shadrina, M.I.; Slominsky, P.A.; Kasheverov, I.E.; Utkin, Y.N.; Ugrumov, M.V.; Tsetlin, V.I. Possible involvement of neuronal nicotinic acetylcholine receptors in compensatory brain mechanisms at early stages of parkinson’s disease. Biomed. Khim 2017, 63, 241–247. [Google Scholar] [CrossRef]
- Tsai, P.C.; Fu, Y.S.; Chang, L.S.; Lin, S.R. Cardiotoxin iii inhibits hepatocyte growth factor-induced epithelial-mesenchymal transition and suppresses invasion of mda-mb-231 cells. J. Biochem. Mol. Toxicol 2016, 30, 12–21. [Google Scholar] [CrossRef]
- Ebrahim, K.; Vatanpour, H.; Zare, A.; Shirazi, F.H.; Nakhjavani, M. Anticancer activity a of caspian cobra (naja naja oxiana) snake venom in human cancer cell lines via induction of apoptosis. Iran J. Pharm Res. 2016, 15, 101–112. [Google Scholar]
- Blanchet, G.; Alili, D.; Protte, A.; Upert, G.; Gilles, N.; Tepshi, L.; Stura, E.A.; Mourier, G.; Servent, D. Ancestral protein resurrection and engineering opportunities of the mamba aminergic toxins. Sci. Rep. 2017, 7, 2701. [Google Scholar] [CrossRef] [Green Version]
- Modahl, C.M.; Roointan, A.; Rogers, J.; Currier, K.; Mackessy, S.P. Interspecific and intraspecific venom enzymatic variation among cobras (naja sp. And ophiophagus hannah). Comp. Biochem Physiol. C Toxicol. Pharm. 2020, 232, 108743. [Google Scholar] [CrossRef]
- Deka, A.; Gogoi, A.; Das, D.; Purkayastha, J.; Doley, R. Proteomics of naja kaouthia venom from north east india and assessment of indian polyvalent antivenom by third generation antivenomics. J. Proteom. 2019, 207, 103463. [Google Scholar] [CrossRef]
- Tan, K.Y.; Wong, K.Y.; Tan, N.H.; Tan, C.H. Quantitative proteomics of naja annulifera (sub-saharan snouted cobra) venom and neutralization activities of two antivenoms in africa. Int. J. Biol. Macromol. 2020, 158, 605–616. [Google Scholar] [CrossRef]
- Tan, C.H.; Wong, K.Y.; Chong, H.P.; Tan, N.H.; Tan, K.Y. Proteomic insights into short neurotoxin-driven, highly neurotoxic venom of philippine cobra (naja philippinensis) and toxicity correlation of cobra envenomation in asia. J. Proteom. 2019, 206, 103418. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.H.; Wong, K.Y.; Tan, N.H.; Ng, T.S.; Tan, K.Y. Distinctive distribution of secretory phospholipases a(2) in the venoms of afro-asian cobras (subgenus: Naja, afronaja, boulengerina and uraeus). Toxins 2019, 11, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodgson, W.C.; Dal Belo, C.A.; Rowan, E.G. The neuromuscular activity of paradoxin: A presynaptic neurotoxin from the venom of the inland taipan (oxyuranus microlepidotus). Neuropharmacology 2007, 52, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Armugam, A.; Gong, N.; Li, X.; Siew, P.Y.; Chai, S.C.; Nair, R.; Jeyaseelan, K. Group ib phospholipase a2 from pseudonaja textilis. Arch. Biochem. Biophys. 2004, 421, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.K.; Luthra, M.G.; Wells, M.A.; Watts, R.P.; Hanahan, D.J. Phospholipase a2 as a probe of phospholipid distribution in erythrocyte membranes. Factors influencing the apparent specificity of the reaction. Biochemistry 1975, 14, 5400–5408. [Google Scholar] [CrossRef]
- Vargas, L.J.; Londono, M.; Quintana, J.C.; Rua, C.; Segura, C.; Lomonte, B.; Nunez, V. An acidic phospholipase a(2) with antibacterial activity from porthidium nasutum snake venom. Comp. Biochem Physiol. B Biochem. Mol. Biol. 2012, 161, 341–347. [Google Scholar] [CrossRef]
- Marques, P.P.; Esteves, A.; Lancellotti, M.; Ponce-Soto, L.A.; Marangoni, S. Novel acidic phospholipase a2 from porthidium hyoprora causes inflammation with mast cell rich infiltrate. Biochem. Biophys. Rep. 2015, 1, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, J.; Gutierrez, J.M.; Angulo, Y.; Sanz, L.; Juarez, P.; Calvete, J.J.; Lomonte, B. Isolation of an acidic phospholipase a2 from the venom of the snake bothrops asper of costa rica: Biochemical and toxicological characterization. Biochimie 2010, 92, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Mora-Obando, D.; Fernandez, J.; Montecucco, C.; Gutierrez, J.M.; Lomonte, B. Synergism between basic asp49 and lys49 phospholipase a2 myotoxins of viperid snake venom in vitro and in vivo. PLoS ONE 2014, 9, e109846. [Google Scholar] [CrossRef] [Green Version]
- Laustsen, A.H. Toxin synergism in snake venoms. J. Toxin Rev. 2016, 35, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Charris, E.; Montealegre-Sanchez, L.; Solano-Redondo, L.; Castro-Herrera, F.; Fierro-Perez, L.; Lomonte, B. Divergent functional profiles of acidic and basic phospholipases a2 in the venom of the snake porthidium lansbergii lansbergii. Toxicon 2016, 119, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M. Excitement ahead: Structure, function and mechanism of snake venom phospholipase a2 enzymes. Toxicon 2003, 42, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, J.P.; Lin, Y.; Lambeau, G.; Ghomashchi, F.; Lazdunski, M.; Gelb, M.H. Localization of structural elements of bee venom phospholipase a2 involved in n-type receptor binding and neurotoxicity. J. Biol. Chem. 1997, 272, 7173–7181. [Google Scholar] [CrossRef] [Green Version]
- Arita, H.; Hanasaki, K.; Nakano, T.; Oka, S.; Teraoka, H.; Matsumoto, K. Novel proliferative effect of phospholipase a2 in swiss 3t3 cells via specific binding site. J. Biol. Chem. 1991, 266, 19139–19141. [Google Scholar] [PubMed]
- Fry, M.R.; Ghosh, S.S.; East, J.M.; Franson, R.C. Role of human sperm phospholipase a2 in fertilization: Effects of a novel inhibitor of phospholipase a2 activity on membrane perturbations and oocyte penetration. Biol. Reprod. 1992, 47, 751–759. [Google Scholar] [CrossRef]
- Nakajima, M.; Hanasaki, K.; Ueda, M.; Arita, H. Effect of pancreatic type phospholipase a2 on isolated porcine cerebral arteries via its specific binding sites. Febs Lett. 1992, 309, 261–264. [Google Scholar] [CrossRef] [Green Version]
- Malih, I.; Ahmad, R.M.R.; Tee, T.Y.; Saile, R.; Ghalim, N.; Othman, I. Proteomic analysis of moroccan cobra naja haje legionis venom using tandem mass spectrometry. J. Proteom. 2014, 96, 240–252. [Google Scholar] [CrossRef]
- Yap, M.K.; Fung, S.Y.; Tan, K.Y.; Tan, N.H. Proteomic characterization of venom of the medically important southeast asian naja sumatrana (equatorial spitting cobra). Acta Trop. 2014, 133, 15–25. [Google Scholar] [CrossRef]
- Huang, H.W.; Liu, B.S.; Chien, K.Y.; Chiang, L.C.; Huang, S.Y.; Sung, W.C.; Wu, W.G. Cobra venom proteome and glycome determined from individual snakes of naja atra reveal medically important dynamic range and systematic geographic variation. J. Proteom. 2015, 128, 92–104. [Google Scholar] [CrossRef]
- Shan, L.L.; Gao, J.F.; Zhang, Y.X.; Shen, S.S.; He, Y.; Wang, J.; Ma, X.M.; Ji, X. Proteomic characterization and comparison of venoms from two elapid snakes (bungarus multicinctus and naja atra) from china. J. Proteom. 2016, 138, 83–94. [Google Scholar] [CrossRef]
- Sintiprungrat, K.; Watcharatanyatip, K.; Senevirathne, W.D.; Chaisuriya, P.; Chokchaichamnankit, D.; Srisomsap, C.; Ratanabanangkoon, K. A comparative study of venomics of naja naja from india and sri lanka, clinical manifestations and antivenomics of an indian polyspecific antivenom. J. Proteom. 2016, 132, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, M.; McCleary, R.J.R.; Kesherwani, M.; Kini, R.M.; Velmurugan, D. Comparison of proteomic profiles of the venoms of two of the ‘big four’ snakes of india, the indian cobra (naja naja) and the common krait (bungarus caeruleus), and analyses of their toxins. Toxicon 2017, 135, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Lauridsen, L.P.; Laustsen, A.H.; Lomonte, B.; Gutierrez, J.M. Exploring the venom of the forest cobra snake: Toxicovenomics and antivenom profiling of naja melanoleuca. J. Proteom. 2017, 150, 98–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hus, K.K.; Buczkowicz, J.; Petrilla, V.; Petrillova, M.; Lyskowski, A.; Legath, J.; Bocian, A. First look at the venom of naja ashei. Molecules 2018, 23, 609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva-de-Franca, F.; Villas-Boas, I.M.; Serrano, S.M.T.; Cogliati, B.; Chudzinski, S.A.A.; Lopes, P.H.; Kitano, E.S.; Okamoto, C.K.; Tambourgi, D.V. Naja annulifera snake: New insights into the venom components and pathogenesis of envenomation. PLoS Negl. Trop. Dis. 2019, 13, e0007017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, J.M.; Rucavado, A. Snake venom metalloproteinases: Their role in the pathogenesis of local tissue damage. Biochimie 2000, 82, 841–850. [Google Scholar] [CrossRef]
- Takeda, S.; Takeya, H.; Iwanaga, S. Snake venom metalloproteinases: Structure, function and relevance to the mammalian adam/adamts family proteins. Biochim. Biophys. Acta 2012, 1824, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Markland, F.S., Jr.; Swenson, S. Snake venom metalloproteinases. Toxicon 2013, 62, 3–18. [Google Scholar] [CrossRef]
- Gutierrez, J.M.; Escalante, T.; Rucavado, A.; Herrera, C. Hemorrhage caused by snake venom metalloproteinases: A journey of discovery and understanding. Toxins 2016, 8, 93. [Google Scholar] [CrossRef] [Green Version]
- Bernardoni, J.L.; Sousa, L.F.; Wermelinger, L.S.; Lopes, A.S.; Prezoto, B.C.; Serrano, S.M.; Zingali, R.B.; Moura-da-Silva, A.M. Functional variability of snake venom metalloproteinases: Adaptive advantages in targeting different prey and implications for human envenomation. PLoS ONE 2014, 9, e109651. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, J.M.; Escalante, T.; Rucavado, A. Experimental pathophysiology of systemic alterations induced by bothrops asper snake venom. Toxicon 2009, 54, 976–987. [Google Scholar] [CrossRef]
- Razi, M.T.; Asad, M.H.; Khan, T.; Chaudhary, M.Z.; Ansari, M.T.; Arshad, M.A.; Saqib, Q.N. Antihaemorrhagic potentials of fagonia cretica against naja naja karachiensis (black pakistan cobra) venom. Nat. Prod. Res. 2011, 25, 1902–1907. [Google Scholar] [CrossRef] [PubMed]
- Costal-Oliveira, F.; Stransky, S.; Guerra-Duarte, C.; Naves de Souza, D.L.; Vivas-Ruiz, D.E.; Yarleque, A.; Sanchez, E.F.; Chavez-Olortegui, C.; Braga, V.M.M. L-amino acid oxidase from bothrops atrox snake venom triggers autophagy, apoptosis and necrosis in normal human keratinocytes. Sci. Rep. 2019, 9, 781. [Google Scholar] [CrossRef]
- Fox, J.W. A brief review of the scientific history of several lesser-known snake venom proteins: L-amino acid oxidases, hyaluronidases and phosphodiesterases. Toxicon 2013, 62, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Izidoro, L.F.; Sobrinho, J.C.; Mendes, M.M.; Costa, T.R.; Grabner, A.N.; Rodrigues, V.M.; da Silva, S.L.; Zanchi, F.B.; Zuliani, J.P.; Fernandes, C.F.; et al. Snake venom l-amino acid oxidases: Trends in pharmacology and biochemistry. BioMed Res. Int. 2014, 2014, 196754. [Google Scholar] [CrossRef] [Green Version]
- Costa, T.R.; Burin, S.M.; Menaldo, D.L.; de Castro, F.A.; Sampaio, S.V. Snake venom l-amino acid oxidases: An overview on their antitumor effects. J. Venom. Anim. Toxins Incl. Trop. Dis. 2014, 20, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suhr, S.M.; Kim, D.S. Identification of the snake venom substance that induces apoptosis. Biochem. Biophys. Res. Commun. 1996, 224, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Ande, S.R.; Kommoju, P.R.; Draxl, S.; Murkovic, M.; Macheroux, P.; Ghisla, S.; Ferrando-May, E. Mechanisms of cell death induction by l-amino acid oxidase, a major component of ophidian venom. Apoptosis 2006, 11, 1439–1451. [Google Scholar] [CrossRef] [PubMed]
- Vogel, C.W.; Muller-Eberhard, H.J. Cobra venom factor: Improved method for purification and biochemical characterization. J. Immunol. Methods 1984, 73, 203–220. [Google Scholar] [CrossRef]
- Laursen, N.S.; Andersen, K.R.; Braren, I.; Spillner, E.; Sottrup-Jensen, L.; Andersen, G.R. Substrate recognition by complement convertases revealed in the c5-cobra venom factor complex. Embo J. 2011, 30, 606–616. [Google Scholar] [CrossRef] [Green Version]
- Vogel, C.W.; Fritzinger, D.C. Cobra venom factor: Structure, function, and humanization for therapeutic complement depletion. Toxicon 2010, 56, 1198–1222. [Google Scholar] [CrossRef]
- Morgan, B.P.; Harris, C.L. Complement therapeutics; history and current progress. Mol. Immunol. 2003, 40, 159–170. [Google Scholar] [CrossRef]
- Wang, B.; Xu, H.; Li, J.; Gao, H.M.; Xing, Y.H.; Lin, Z.; Li, H.J.; Wang, Y.Q.; Cao, S.H. Complement depletion with cobra venom factor alleviates acute hepatic injury induced by ischemiareperfusion. Mol. Med. Rep. 2018, 18, 4523–4529. [Google Scholar] [PubMed]
- Wang, Y.L.; Kuo, J.H.; Lee, S.C.; Liu, J.S.; Hsieh, Y.C.; Shih, Y.T.; Chen, C.J.; Chiu, J.J.; Wu, W.G. Cobra crisp functions as an inflammatory modulator via a novel zn2+ -and heparan sulfate-dependent transcriptional regulation of endothelial cell adhesion molecules. J. Biol. Chem. 2010, 285, 37872–37883. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.S.; Liou, J.C.; Lin, S.R.; Cheng, Y.C. Purification and characterization of taiwan cobra venom proteins with weak toxicity. Toxicon 2005, 45, 21–25. [Google Scholar] [CrossRef]
- Wang, J.; Shen, B.; Guo, M.; Lou, X.; Duan, Y.; Cheng, X.P.; Teng, M.; Niu, L.; Liu, Q.; Huang, Q.; et al. Blocking effect and crystal structure of natrin toxin, a cysteine-rich secretory protein from naja atra venom that targets the bkca channel. Biochemistry 2005, 44, 10145–10152. [Google Scholar] [CrossRef]
- Wang, F.; Li, H.; Liu, M.N.; Song, H.; Han, H.M.; Wang, Q.L.; Yin, C.C.; Zhou, Y.C.; Qi, Z.; Shu, Y.Y.; et al. Structural and functional analysis of natrin, a venom protein that targets various ion channels. Biochem. Biophys. Res. Commun. 2006, 351, 443–448. [Google Scholar] [CrossRef]
- Zhou, Q.; Wang, Q.L.; Meng, X.; Shu, Y.; Jiang, T.; Wagenknecht, T.; Yin, C.C.; Sui, S.F.; Liu, Z. Structural and functional characterization of ryanodine receptor-natrin toxin interaction. Biophys. J. 2008, 95, 4289–4299. [Google Scholar] [CrossRef] [Green Version]
- Osipov, A.V.; Levashov, M.Y.; Tsetlin, V.I.; Utkin, Y.N. Cobra venom contains a pool of cysteine-rich secretory proteins. Biochem. Biophys. Res. Commun. 2005, 328, 177–182. [Google Scholar] [CrossRef]
- Serrano, S.M. The long road of research on snake venom serine proteinases. Toxicon 2013, 62, 19–26. [Google Scholar] [CrossRef]
- Kang, T.S.; Georgieva, D.; Genov, N.; Murakami, M.T.; Sinha, M.; Kumar, R.P.; Kaur, P.; Kumar, S.; Dey, S.; Sharma, S.; et al. Enzymatic toxins from snake venom: Structural characterization and mechanism of catalysis. Febs J. 2011, 278, 4544–4576. [Google Scholar] [CrossRef]
- Katzir, I.; Shani, J.; Goshen, G.; Sela, J.; Ninary, E.; Dogonovski, A.M.; Shabashov, D.; Inoue, S.; Ikeda, K.; Hayashi, K.; et al. Characterization of nerve growth factors (ngfs) from snake venoms by use of a novel, quantitative bioassay utilizing pheochromocytoma (pc12) cells overexpressing human trka receptors. Toxicon 2003, 42, 481–490. [Google Scholar] [CrossRef]
- Kostiza, T.; Meier, J. Nerve growth factors from snake venoms: Chemical properties, mode of action and biological significance. Toxicon 1996, 34, 787–806. [Google Scholar] [CrossRef]
- McCleary, R.J.; Kini, R.M. Non-enzymatic proteins from snake venoms: A gold mine of pharmacological tools and drug leads. Toxicon 2013, 62, 56–74. [Google Scholar] [CrossRef]
- Uzair, B.; Khan, B.A.; Sharif, N.; Shabbir, F.; Menaa, F. Phosphodiesterases (pdes) from snake venoms: Therapeutic applications. Protein Pept. Lett. 2018, 25, 612–618. [Google Scholar] [CrossRef]
- Thirawatananond, P.; McPherson, R.L.; Malhi, J.; Nathan, S.; Lambrecht, M.J.; Brichacek, M.; Hergenrother, P.J.; Leung, A.K.L.; Gabelli, S.B. Structural analyses of nudt16-adp-ribose complexes direct rational design of mutants with improved processing of poly(adp-ribosyl)ated proteins. Sci. Rep. 2019, 9, 5940. [Google Scholar] [CrossRef]
- Yin, J.; Chen, S.; Zhang, N.; Wang, H. Multienzyme cascade bioreactor for a 10 min digestion of genomic DNA into single nucleosides and quantitative detection of structural DNA modifications in cellular genomic DNA. Acs. Appl. Mater. Interfaces 2018, 10, 21883–21890. [Google Scholar] [CrossRef]
- Lai, W.; Lyu, C.; Wang, H. Vertical ultrafiltration-facilitated DNA digestion for rapid and sensitive uhplc-ms/ms detection of DNA modifications. Anal. Chem. 2018, 90, 6859–6866. [Google Scholar] [CrossRef]
- Willmann, L.; Erbes, T.; Krieger, S.; Trafkowski, J.; Rodamer, M.; Kammerer, B. Metabolome analysis via comprehensive two-dimensional liquid chromatography: Identification of modified nucleosides from rna metabolism. Anal. Bioanal. Chem. 2015, 407, 3555–3566. [Google Scholar] [CrossRef]
- Carregari, V.C.; Rosa-Fernandes, L.; Baldasso, P.; Bydlowski, S.P.; Marangoni, S.; Larsen, M.R.; Palmisano, G. Snake venom extracellular vesicles (svevs) reveal wide molecular and functional proteome diversity. Sci. Rep. 2018, 8, 12067. [Google Scholar] [CrossRef]
- Ogawa, Y.; Kanai-Azuma, M.; Akimoto, Y.; Kawakami, H.; Yanoshita, R. Exosome-like vesicles in gloydius blomhoffii blomhoffii venom. Toxicon 2008, 51, 984–993. [Google Scholar] [CrossRef]
- Xu, R.; Greening, D.W.; Zhu, H.J.; Takahashi, N.; Simpson, R.J. Extracellular vesicle isolation and characterization: Toward clinical application. J. Clin. Investig. 2016, 126, 1152–1162. [Google Scholar] [CrossRef] [Green Version]
- Yanez-Mo, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borras, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Raposo, G.; Stahl, P.D. Extracellular vesicles: A new communication paradigm? Nat. Rev. Mol. Cell Biol. 2019, 20, 509–510. [Google Scholar] [CrossRef]
- Maas, S.L.N.; Breakefield, X.O.; Weaver, A.M. Extracellular vesicles: Unique intercellular delivery vehicles. Trends Cell Biol. 2017, 27, 172–188. [Google Scholar] [CrossRef] [Green Version]
- Osier, N.; Motamedi, V.; Edwards, K.; Puccio, A.; Diaz-Arrastia, R.; Kenney, K.; Gill, J. Exosomes in acquired neurological disorders: New insights into pathophysiology and treatment. Mol. Neurobiol. 2018, 55, 9280–9293. [Google Scholar] [CrossRef]
- Chong, S.Y.; Lee, C.K.; Huang, C.; Ou, Y.H.; Charles, C.J.; Richards, A.M.; Neupane, Y.R.; Pavon, M.V.; Zharkova, O.; Pastorin, G.; et al. Extracellular vesicles in cardiovascular diseases: Alternative biomarker sources, therapeutic agents, and drug delivery carriers. Int. J. Mol. Sci. 2019, 20, 3272. [Google Scholar] [CrossRef] [Green Version]
- Klein-Scory, S.; Tehrani, M.M.; Eilert-Micus, C.; Adamczyk, K.A.; Wojtalewicz, N.; Schnolzer, M.; Hahn, S.A.; Schmiegel, W.; Schwarte-Waldhoff, I. New insights in the composition of extracellular vesicles from pancreatic cancer cells: Implications for biomarkers and functions. Proteome Sci. 2014, 12, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luhtala, N.; Aslanian, A.; Yates, J.R., 3rd; Hunter, T. Secreted glioblastoma nanovesicles contain intracellular signaling proteins and active ras incorporated in a farnesylation-dependent manner. J. Biol. Chem. 2017, 292, 611–628. [Google Scholar] [CrossRef] [Green Version]
- Cufaro, M.C.; Pieragostino, D.; Lanuti, P.; Rossi, C.; Cicalini, I.; Federici, L.; De Laurenzi, V.; Del Boccio, P. Extracellular vesicles and their potential use in monitoring cancer progression and therapy: The contribution of proteomics. J. Oncol. 2019, 2019, 1639854. [Google Scholar] [CrossRef]
- Kriebel, P.W.; Majumdar, R.; Jenkins, L.M.; Senoo, H.; Wang, W.; Ammu, S.; Chen, S.; Narayan, K.; Iijima, M.; Parent, C.A. Extracellular vesicles direct migration by synthesizing and releasing chemotactic signals. J. Cell Biol. 2018, 217, 2891–2910. [Google Scholar] [CrossRef] [PubMed]
- Meldolesi, J. Exosomes and ectosomes in intercellular communication. Curr. Biol. 2018, 28, R435–R444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willms, E.; Cabanas, C.; Mager, I.; Wood, M.J.A.; Vader, P. Extracellular vesicle heterogeneity: Subpopulations, isolation techniques, and diverse functions in cancer progression. Front. Immunol. 2018, 9, 738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olamendi-Portugal, T.; Batista, C.V.F.; Pedraza-Escalona, M.; Restano-Cassulini, R.; Zamudio, F.Z.; Benard-Valle, M.; de Roodt, A.R.; Possani, L.D. New insights into the proteomic characterization of the coral snake micrurus pyrrhocryptus venom. Toxicon 2018, 153, 23–31. [Google Scholar] [CrossRef]
- Wolfenden, R.N. On the nature and action of the venom of poisonous snakes: Ii. A note upon the venom of the indian viper (daboia russellii). J. Physiol. 1886, 7, 357–364. [Google Scholar] [CrossRef]
- Shao, J.; Shen, H.; Havsteen, B. Purification, characterization and binding interactions of the chinese-cobra (naja naja atra) serum antitoxic protein csap. Biochem J. 1993, 293, 559–566. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Buck, F.; Havsteen, B. Elucidation of a new biological function of an old protein: Unique structure of the cobra serum albumin controls its specific toxin binding activity. Int. J. Biochem. Cell Biol. 1998, 30, 225–233. [Google Scholar] [CrossRef]
- Ohkura, N.; Inoue, S.; Ikeda, K.; Hayashi, K. The two subunits of a phospholipase a2 inhibitor from the plasma of thailand cobra having structural similarity to urokinase-type plasminogen activator receptor and ly-6 related proteins. Biochem. Biophys Res. Commun 1994, 204, 1212–1218. [Google Scholar] [CrossRef]
- Okumura, K.; Masui, K.; Inoue, S.; Ikeda, K.; Hayashi, K. Purification, characterization and cdna cloning of a phospholipase a2 inhibitor from the serum of the non-venomous snake elaphe quadrivirgata. Biochem. J. 1999, 341, 165–171. [Google Scholar] [CrossRef]
- Ullah, A.; Masood, R. The sequence and three-dimensional structure characterization of snake venom phospholipases b. Front. Mol. Biosci. 2020, 7, 175. [Google Scholar] [CrossRef]
- Georgieva, D.; Seifert, J.; Ohler, M.; von Bergen, M.; Spencer, P.; Arni, R.K.; Genov, N.; Betzel, C. Pseudechis australis venomics: Adaptation for a defense against microbial pathogens and recruitment of body transferrin. J. Proteome Res. 2011, 10, 2440–2464. [Google Scholar] [CrossRef]
- Takahashi, H.; Hattori, S.; Iwamatsu, A.; Takizawa, H.; Shibuya, M. A novel snake venom vascular endothelial growth factor (vegf) predominantly induces vascular permeability through preferential signaling via vegf receptor-1. J. Biol. Chem. 2004, 279, 46304–46314. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, Y.; Matsunaga, Y.; Tokunaga, Y.; Obayashi, S.; Saito, M.; Morita, T. Snake venom vascular endothelial growth factors (vegf-fs) exclusively vary their structures and functions among species. J. Biol. Chem. 2009, 284, 9885–9891. [Google Scholar] [CrossRef] [Green Version]
- Toivanen, P.I.; Nieminen, T.; Laakkonen, J.P.; Heikura, T.; Kaikkonen, M.U.; Yla-Herttuala, S. Snake venom vegf vammin induces a highly efficient angiogenic response in skeletal muscle via vegfr-2/nrp specific signaling. Sci. Rep. 2017, 7, 5525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norton, R.S.; Chandy, K.G. Venom-derived peptide inhibitors of voltage-gated potassium channels. Neuropharmacology 2017, 127, 124–138. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Feng, J.; Wang, B.; Cao, Z.; Li, W.; Wu, Y.; Chen, Z. Bf9, the first functionally characterized snake toxin peptide with kunitz-type protease and potassium channel inhibiting properties. J. Biochem. Mol. Toxicol. 2014, 28, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Pung, Y.F.; Wong, P.T.; Kumar, P.P.; Hodgson, W.C.; Kini, R.M. Ohanin, a novel protein from king cobra venom, induces hypolocomotion and hyperalgesia in mice. J. Biol. Chem. 2005, 280, 13137–13147. [Google Scholar] [CrossRef] [Green Version]
- Aird, S.D. Ophidian envenomation strategies and the role of purines. Toxicon 2002, 40, 335–393. [Google Scholar] [CrossRef]
- Dhananjaya, B.L.; D’Souza, C.J. The pharmacological role of nucleotidases in snake venoms. Cell Biochem. Funct. 2010, 28, 171–177. [Google Scholar] [CrossRef]
- Vaiyapuri, S.; Wagstaff, S.C.; Watson, K.A.; Harrison, R.A.; Gibbins, J.M.; Hutchinson, E.G. Purification and functional characterisation of rhiminopeptidase a, a novel aminopeptidase from the venom of bitis gabonica rhinoceros. PLoS Negl. Trop. Dis. 2010, 4, e796. [Google Scholar] [CrossRef] [Green Version]
- Brillard-Bourdet, M.; Nguyen, V.; Ferrer-di Martino, M.; Gauthier, F.; Moreau, T. Purification and characterization of a new cystatin inhibitor from taiwan cobra (naja naja atra) venom. Biochem. J. 1998, 331 Pt 1, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Fry, B.G.; Vidal, N.; Norman, J.A.; Vonk, F.J.; Scheib, H.; Ramjan, S.F.; Kuruppu, S.; Fung, K.; Hedges, S.B.; Richardson, M.K.; et al. Early evolution of the venom system in lizards and snakes. Nature 2006, 439, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.M.; Lomonte, B.; Leon, G.; Rucavado, A.; Chaves, F.; Angulo, Y. Trends in snakebite envenomation therapy: Scientific, technological and public health considerations. Curr. Pharm. Des. 2007, 13, 2935–2950. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Gan, T.X.; Liu, X.D.; Jin, Y.; Lee, W.H.; Shen, J.H.; Zhang, Y. Identification and characterization of novel reptile cathelicidins from elapid snakes. Peptides 2008, 29, 1685–1691. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.H.; Hyun, J.Y.; Varshavsky, A. Control of hsp90 chaperone and its clients by n-terminal acetylation and the n-end rule pathway. Proc. Natl. Acad. Sci. USA 2017, 114, E4370–E4379. [Google Scholar] [CrossRef] [Green Version]
- Shemorry, A.; Hwang, C.S.; Varshavsky, A. Control of protein quality and stoichiometries by n-terminal acetylation and the n-end rule pathway. Mol. Cell 2013, 50, 540–551. [Google Scholar] [CrossRef] [Green Version]
- Drazic, A.; Myklebust, L.M.; Ree, R.; Arnesen, T. The world of protein acetylation. Biochim Biophys. Acta 2016, 1864, 1372–1401. [Google Scholar] [CrossRef] [Green Version]
- Ree, R.; Varland, S.; Arnesen, T. Spotlight on protein n-terminal acetylation. Exp. Mol. Med. 2018, 50, 90. [Google Scholar] [CrossRef] [Green Version]
- Ma, B. Novor: Real-time peptide de novo sequencing software. J. Am. Soc. Mass Spectrom 2015, 26, 1885–1894. [Google Scholar] [CrossRef] [Green Version]
- Muth, T.; Weilnbock, L.; Rapp, E.; Huber, C.G.; Martens, L.; Vaudel, M.; Barsnes, H. Denovogui: An open source graphical user interface for de novo sequencing of tandem mass spectra. J. Proteome Res. 2014, 13, 1143–1146. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The pride database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Family | First Report in Nn Venom | Abbreviation Used | NN (No of Peptides) | %Age | NO(No. of Peptides) | %Age |
|---|---|---|---|---|---|---|
| Three-Finger toxin | 3FTX | 157 | 21 | 41 | 16 | |
| Snake venom metalloprotease family | SVMP | 72 | 10 | 39 | 15 | |
| Cobra venom factor | CVF | 62 | 9 | 22 | 8.7 | |
| Cysteine-rich secretory protein | CRISP | 53 | 7 | 7 | 2.8 | |
| Phospholipase A2 | PLA2 | 46 | 6 | 32 | 12.6 | |
| Phospholipase B | PLB | 1 | 0.1 | 4 | 1.6 | |
| Phospholipase inhibitor | ✓ | CNF-I | 3 | 0.4 | - | - |
| L-amino-acid oxidase | LAAO | 31 | 4 | 14 | 5.5 | |
| Snake Venom Serine proteinase | SP | 15 | 2 | 11 | 4.3 | |
| Ohanin | Oh | 11 | 1.5 | 2 | 0.8 | |
| Kunitz type serine protease inhibitor | KSPI | 14 | 2 | 4 | 1.6 | |
| Nerve Growth Factor | NGF | 12 | 1.7 | 11 | 4.3 | |
| 5′-nucleotidase | 5-Ntd | 10 | 1.4 | 1 | 0.4 | |
| Serum Albumin | ✓ | SA | 10 | 1.4 | - | |
| Glutathione peroxidase | ✓ | GP | 9 | 1.2 | 3 | 1.2 |
| Phosphodiesterase | Pde | 8 | 1.1 | 8 | 3.1 | |
| Aminopeptidase | - | 7 | 1 | 4 | 1.6 | |
| TNF receptor family | ✓ | TNF | 2 | 0.3 | 3 | 1.2 |
| Lectin | ✓ | - | 3 | 0.4 | 1 | 0.4 |
| Natriuretic peptide family | NP | 4 | 0.54 | 1 | 0.4 | |
| Cystatin | - | 4 | 0.54 | - | ||
| Cathelicidin | ✓ | cath | 1 | 0.1 | - | |
| N-acetylcholinesterase | N-Ache | 1 | 0.1 | 1 | 0.4 | |
| Vascular endothelial growth factor | VEGF | 1 | 0.1 | - | ||
| Transforming growth factor | ✓ | TGF | 2 | 0.3 | - | |
| Zinc finger protein | ✓ | ZFP | 6 | 0.8 | 4 | 1.6 |
| Insulin | ✓ | In | 2 | 0.3 | - | |
| Transferrin | ✓ | TF | 2 | 0.3 | - | |
| Ankyrin repeat | ✓ | AR | 2 | 0.3 | 1 | 0.4 |
| Leucine repeat | ✓ | LR | 1 | 0.1 | 1 | 0.4 |
| Endonuclease | ✓ | - | 3 | 0.4 | - | |
| SLRP family | ✓ | SLRP | 2 | 0.3 | 1 | 0.4 |
| Ras-like protein | ✓ | Ras | 5 | 0.7 | - | |
| Serpin | - | - | 1 | 0.4 | ||
| Others | - | 158 | 37 | |||
| Total | 735 | 254 |
| S. No | Protein Family | Protein | Accession Code | Number of Matched Peptides | Homology with a Protein from the Venom of Snake Species |
|---|---|---|---|---|---|
| 1 | 3FTXs (Neurotoxin) | Long neurotoxin | AHZ08824 | 9 | Micropechis ikaheca |
| 2 | P01389 | 1 | Naja anchietae | ||
| 3 | P01390 | 2 | Naja nivea | ||
| 4 | Long neurotoxin homolog | O93422 | 5 | Naja atra | |
| 5 | Long neurotoxin 1 | P25668 | 4 | Naja naja | |
| 6 | Long neurotoxin 1 | P01380 | 1 | Hydrophis stokesii | |
| 7 | Long neurotoxin 1 | P25674 | 3 | Naja haje haje | |
| 8 | Long neurotoxin 4 | P25672 | 3 | Naja naja | |
| 9 | Long neurotoxin 7 | O42257 | 3 | Naja sputatrix | |
| 10 | putative long neurotoxin | ABX58151 | 1 | Austrelaps labialis | |
| 11 | putative long neurotoxin | ABX58163 | 1 | Austrelaps labialis | |
| 12 | Alpha-neurotoxin NTX-3 | O57326 | 1 | Naja sputatrix | |
| 13 | Short neurotoxin 3 | P01420 | 1 | Naja annulifera | |
| 14 | Short neurotoxin III | P59275 | 1 | Naja kaouthia | |
| 15 | Neurotoxin II | P01427 | 6 | Naja oxiana | |
| 16 | cobrotoxin b | CAA73829 | 3 | Naja atra | |
| 17 | Cobrotoxin-b | P80958 | 4 | Naja atra | |
| 18 | Alpha-cobratoxin | P01391 | 4 | Naja kaouthia | |
| 19 | kappa-cobrotoxin | CAA76846 | 1 | Naja atra | |
| 20 | Weak toxin 2 | Q8AY50 | 2 | Bungarus candidus | |
| 21 | Weak neurotoxin 7 | P29181 | 7 | Naja naja | |
| 22 | Weak neurotoxin 10 | Q802B2 | 1 | Naja sputatrix | |
| 23 | Weak toxin CM-11 | P01401 | 4 | Naja haje haje | |
| 24 | Weak toxin S4C11 | P01400 | 5 | Naja melanoleuca | |
| 25 | three-finger toxin precursor, partial | ADN67572 | 4 | Bungarus multicinctus | |
| 26 | three-finger toxin precursor, partial | ADN67582 | 9 | Naja atra | |
| 27 | three-finger toxin precursor, partial | ADN67583 | 1 | Naja atra | |
| 28 | three-finger toxin precursor | ADN67579 | 1 | Naja atra | |
| 29 | Muscarinic toxin-like protein 3 | P82464 | 3 | Naja kaouthia | |
| 30 | Muscarinic toxin-like protein | Q9W727 | 4 | Bungarus multicinctus | |
| 31 | Muscarinic toxin-like protein 2 | P82463 | 6 | Naja kaouthia | |
| 32 | Muscarinic toxin-like protein 1 | P82462 | 2 | Naja kaouthia | |
| 33 | Muscarinic toxin 38 | Q2VBN0 | 1 | Ophiophagus hannah | |
| 34 | Alpha-elapitoxin-Nk2a | P01391 | 4 | Naja kaouthia | |
| 36 | three finger toxin V | ABX82866 | 1 | Walterinnesia aegyptia | |
| 37 | Three finger toxin W-V | C1IC49 | 3 | Walterinnesia aegyptia | |
| 38 | Chain A, Putative Ancestral Mamba Toxin 1 | 5MG9_A | 1 | Dendroaspis angusticeps | |
| 39 | 3FTXs (cytotoxins) | cytotoxin 17, partial | BAU24676 | 13 | Naja naja |
| 40 | Cytotoxin Vc-5 | Q9PS34 | 6 | Naja oxiana | |
| 41 | Cytotoxin 3a | P86539 | 4 | Naja naja | |
| 42 | Cytotoxin SP15c | P60308 | 13 | Naja atra | |
| 43 | cardiotoxin 7a | AAB36929 | 2 | Naja atra | |
| 44 | cardiotoxin 7a | Q91126 | 3 | Naja atra | |
| 46 | Cytotoxin 8 | P86540 | 2 | Naja naja | |
| 47 | Cytotoxin 1 | P01447 | 1 | Naja naja | |
| 48 | Cytotoxin II | P01441 | 1 | Naja oxiana | |
| 49 | Cytotoxin 5 | P25517 | 2 | Naja mossambica | |
| 50 | Cardiotoxin-6 | Q98965 | 1 | Naja atra | |
| 51 | Cytotoxin 10 | P86541 | 1 | Naja naja | |
| 52 | Cytotoxin homolog 3 | P01473 | 1 | Naja melanoleuca | |
| 53 | Cardiotoxin-like basic polypeptide ah | P0C547 | 2 | Naja atra | |
| 54 | cardiotoxin 1e | AAA90960 | 4 | Naja atra | |
| 55 | Venom complement C3-like | Venom factor | AAX86641 | 5 | Austrelaps superbus |
| 56 | Cobra venom factor | Q91132 | 31 | Naja kaouthia | |
| 57 | Cobra venom factor gamma chain | Q91132 | 2 | Naja kaouthia | |
| 58 | Cobra venom factor alpha chain | Q91132 | 2 | Naja kaouthia | |
| 59 | cobra venom factor precursor | AAA68989 | 1 | Naja kaouthia | |
| 60 | venom factor-like, partial | XP_025025833 | 2 | Python bivittatus | |
| 61 | cobra venom factor 1, partial | AXL96620 | 13 | Ahaetulla prasina | |
| 62 | cobra venom factor, partial | AXL95279 | 1 | Spilotes sulphureus | |
| 63 | cobra venom factor, partial | AWX67646 | 1 | Boiga irregularis | |
| 64 | Ophiophagus venom factor | I2C090 | 3 | Ophiophagus hannah | |
| 66 | Venom Kunitz-type family | Kunitz-type serine protease inhibitor | P19859 | 1 | Naja naja |
| 67 | Kunitz-type serine protease inhibitor | P20229 | 6 | Naja naja | |
| 68 | Kunitz-type serine protease inhibitor isoform 7 | ACY68703 | 1 | Parasuta nigriceps | |
| 69 | Kunitz inhibitor b, partial | AAL30069 | 1 | Bungarus candidus | |
| 70 | protease inhibitor | AFA90080 | 1 | Daboia siamensis | |
| 71 | Venom basic protease inhibitor 2 | P00986 | 1 | Naja nivea | |
| 72 | Kunitz-type protease inhibitor, partial | AWX67660 | 1 | Boiga irregularis | |
| 73 | papilin-like, partial | XP_025032351 | 1 | Python bivittatus | |
| 74 | Kunitz inhibitor I | ABX82867 | 1 | Walterinnesia aegyptia | |
| 75 | natriuretic peptide family | Natriuretic peptide Na-NP | D9IX97 | 2 | Naja atra |
| 76 | natriuretic peptide | ADK12001 | 1 | Naja atra | |
| 77 | natriuretic peptide | ADK12001 | 1 | Naja atra | |
| 78 | cystatin | Cystatin | E3P6P4 | 4 | Naja kaouthia |
| 79 | NGF-beta family | Venom nerve growth factor 2 | Q5YF89 | 2 | Naja sputatrix |
| 80 | Venom nerve growth factor 3 | Q3HXY1 | 7 | Pseudechis australis | |
| 81 | nerve growth factor, partial | AAR24530 | 1 | Bitis gabonica | |
| 82 | nerve growth factor | BAN82142 | 4 | Ovophis okinavensis | |
| 83 | nerve growth factor beta chain precursor | A59218 | 1 | Naja kaouthia | |
| 84 | Ohanin/vespryn family. | Ohanin | P83234 | 4 | Ophiophagus hannah |
| 85 | Thaicobrin | P82885 | 2 | Naja kaouthia | |
| 86 | Venom PRY-SPRY domain-containing protein, partial | AHZ08803 | 4 | Micropechis ikaheca | |
| 87 | Vespryn | AEJ32004 | 1 | Crotalus adamanteus | |
| 88 | Insulin family | Insulin-like growth factor-binding protein 3, partial | XP_025032248 | 1 | Python bivittatus |
| 89 | Insulin enhancer protein ISL-1, partial | ETE72105 | 1 | Ophiophagus hannah | |
| 90 | Snake venom VEGF subfamily | Snake venom vascular endothelial growth factor toxin barietin | C0K3N1 | 1 | Bitis arietans |
| 91 | CRISP | Cysteine-rich venom protein 25 | P84806 | 6 | Naja haje haje |
| 92 | cysteine-rich seceretory protein Ts-CRPM | ACE73574 | 2 | Trimeresurus stejnegeri | |
| 93 | Cysteine-rich venom protein mossambin | P0DL16 | 2 | Naja mossambica | |
| 94 | Cysteine-rich venom protein natrin-1 | Q7T1K6 | 16 | Naja atra | |
| 95 | Cysteine-rich venom protein ophanin | Q7ZT98 | 3 | Ophiophagus hannah | |
| 96 | cysteine-rich venom protein, partial | BAP39957 | 1 | Protobothrops flavoviridis | |
| 97 | Cysteine-rich venom protein natrin-2 | Q7ZZN8 | 3 | Naja atra | |
| 98 | Cysteine-rich seceretory protein Ts-CRPM | N-ACE73574 | 1 | Trimeresurus stejnegeri | |
| 99 | Cysteine-rich venom protein 25-A | P84807 | 1 | Naja haje haje | |
| 100 | Helicopsin | P0DJG8 | 2 | Helicops angulatus | |
| 101 | Cysteine-rich venom protein bucarin | P81993 | 1 | Bungarus candidus | |
| 102 | Cysteine-rich venom protein latisemin | Q8JI38 | 1 | Laticauda semifasciata | |
| 103 | Cysteine-rich venom protein ophanin | AAO62996 | 1 | Ophiophagus hannah | |
| 104 | cysteine-rich secretory protein 4, partial | AXL96584 | 2 | Borikenophis portoricensis | |
| 105 | Cysteine-rich venom protein kaouthin-1 | P84805 | 1 | Naja kaouthia | |
| 106 | Cysteine-rich venom protein annuliferin-b | P0DL15 | 1 | Naja annulifera | |
| 107 | Cysteine-rich venom protein | AAP20603 | 2 | Naja atra | |
| 108 | Cysteine-rich secretory protein | AJB84505 | 1 | Philodryas chamissonis | |
| 109 | Opharin precursor | AAP81292 | 1 | Ophiophagus hannah | |
| 110 | Cysteine rich secretory protein 2, partial | AXL96629 | 4 | Ahaetulla prasina | |
| 111 | Cathelicidin family | Cathelicidin-related protein precursor | ACF21000 | 1 | Naja atra |
| 112 | TGF-beta family | Transforming growth factor beta-3, partial | ETE71774 | 1 | Ophiophagus hannah |
| 113 | Glial cell line-derived neurotrophic factor, partial | ETE67324 | 1 | Ophiophagus hannah | |
| 114 | Phospholipase A2 | Acidic phospholipase A2 3 | P60045 | 4 | Naja sagittifera |
| 115 | 85 kDa calcium-independent phospholipase A2, partial | ETE71158 | 2 | Ophiophagus hannah | |
| 116 | Acidic phospholipase A2 1 | P00596 | 4 | Naja kaouthia | |
| 117 | Acidic phospholipase A2 1 | Q9W7J4 | 6 | Pseudonaja textilis | |
| 118 | Basic phospholipase A2 T1-2 A chain | P84472 | 2 | Bungarus candidus | |
| 119 | Acidic phospholipase A2 C | Q92086 | 5 | Naja sputatrix | |
| 120 | Acidic phospholipase A2 1 | P00598 | 3 | Naja naja | |
| 121 | Acidic phospholipase A2 2 | P60044 | 1 | Naja sagittifera | |
| 122 | Acidic phospholipase A2 1 | P00596 | 4 | Naja kaouthia | |
| 123 | Phospholipase A2 | BAA36403 | 1 | Naja kaouthia | |
| 124 | Acidic phospholipase A2 beta-bungarotoxin A4 chain | P17934 | 2 | Bungarus multicinctus | |
| 125 | Phospholipase A2-III | ABD24038 | 1 | Daboia russelii russelii | |
| 126 | Basic phospholipase A2 homolog 1 | P10117 | 1 | Laticauda colubrina | |
| 127 | Phospholipase A2 | AAL55555 | 1 | Hydrophis hardwickii | |
| 128 | Phospholipase A2 | P15445 (2WQ5) | 1 | Naja naja | |
| 129 | Phospholipase A2 3 | P21792 | 3 | Micrurus nigrocinctus | |
| 130 | Phospholipase A2I precursor | BAC77655 | 1 | Bungarus flaviceps | |
| 131 | Phospholipase a2 | CAA45372 | 1 | Naja naja | |
| 132 | Phospholipase A2 | AAA66029 | 1 | Naja naja | |
| 133 | Phosphatidylcholine 2-acylhydrolase T1-2 A | P84472 | 2 | Bungarus candidus | |
| 134 | Phospholipase B-like family | Phospholipase B-like 1, partial | ETE59578 | 1 | Ophiophagus hannah |
| 135 | CNF-like-inhibitor family | Phospholipase A2 inhibitor subunit gamma A | Q9PWI4 | 1 | Elaphe quadrivirgata |
| 136 | Phospholipase A2 inhibitor beta subunit isoform OMI-2B | AAF21049 | 1 | Oxyuranus microlepidotus | |
| 137 | Phospholipase A2 inhibitor 31 kDa subunit | Q7LZI1 | 1 | Naja kaouthia | |
| 138 | SVMP (PIII) | Acutolysin e precursor | AAD27891 | 1 | Deinagkistrodon acutus |
| 139 | Snake venom metalloproteinase | D5LMJ3 | 12 | Naja atra | |
| 140 | Snake venom metalloproteinase | D3TTC1 | 20 | Naja atra | |
| 141 | Snake venom metalloproteinase | D3TTC2 | 8 | Naja atra | |
| 142 | Snake venom metalloproteinase-disintegrin-like mocarhagin | Q10749 | 7 | Naja mossambica | |
| 143 | Snake venom metalloproteinase | Q9PVK7 | 5 | Naja kaouthia | |
| 144 | Snake venom metalloproteinase | A8QL49 | 2 | Bungarus multicinctus | |
| 145 | Snake venom metalloproteinase | P82942 | 8 | Naja kaouthia | |
| 146 | Snake venom metalloprotease(ADAM) | ACS74986 | 1 | Philodryas olfersii | |
| 147 | Snake venom metalloproteinase 27, partial | AXL96577 | 1 | Borikenophis portoricensis | |
| 148 | Disintegrin and metalloproteinase domain-containing protein 21, partial | ETE71596 | 2 | Ophiophagus hannah | |
| 149 | Microlepidotease-1 | ABQ01137 | 1 | Oxyuranus microlepidotus | |
| 150 | Metalloproteinase atrase B, partial | ADD14036 | 1 | Naja atra | |
| 151 | Metalloproteinase 7, partial | AXL96626 | 1 | Ahaetulla prasina | |
| 152 | Snake venom metalloproteinase | P0DM46 | 1 | Micrurus corallinus | |
| 153 | K-like metalloprotease precursor, partial | ACN50005 | 1 | Naja atra | |
| 154 | Snake venom serine proteinase (peptidase S1 family) | Tissue-type plasminogen activator, partial | ETE66683 | 3 | Ophiophagus hannah |
| 155 | Tissue-type plasminogen activator-like, partial | XP_025033187 | 3 | Python bivittatus | |
| 156 | Complement factor B precursor | AAR21601 | 1 | Naja kaouthia | |
| 157 | Thrombin-like enzyme TLP | P86545 | 2 | Naja naja | |
| 158 | Serine endopeptidase | AUS82567 | 1 | Crotalus tigris | |
| 159 | Snake venom serine protease NaSP | A8QL53 | 1 | Naja atra | |
| 160 | Snake venom serine protease catroxase-1 | Q8QHK3 | 1 | Crotalus atrox | |
| 161 | Anionic trypsin-1-like | XP_007434941 | 1 | Python bivittatus | |
| 162 | Coagulation factor X isoform 1, partial | ETE73401 | 1 | Ophiophagus hannah | |
| 163 | Serine endopeptidase | AUS82552 | 1 | Crotalus scutulatus | |
| 164 | 5’-nucleotidase family | 5-nucleotidase | BAP39972 | 5 | Protobothrops flavoviridis |
| 165 | Venom 5’-nucleotidase | A0A2I4HXH5 | 3 | Naja atra | |
| 166 | 5’-nucleotidase, partial | ETE67245 | 1 | Ophiophagus hannah | |
| 167 | Snake venom 5’-nucleotidase | B6EWW8 | 1 | Gloydius brevicaudus | |
| 168 | Aminopeptidase | Aminopeptidase N, partial | ETE61021 | 1 | Ophiophagus hannah |
| 169 | Aminopeptidase N | BAG82599 | 6 | Gloydius brevicaudus | |
| 170 | Type-B carboxylesterase/lipase | N-acetylcholinesterase | AAC59905 | 1 | Bungarus fasciatus |
| 171 | Phosphodiesterase | Snake venom Phosphodiesterase | A0A2D0TC04 | 3 | Naja atra |
| 172 | Phosphodiesterase | AHJ80885 | 1 | Macrovipera lebetina | |
| 173 | Phosphodiesterase, partial | AXL96599 | 2 | Borikenophis portoricensis | |
| 174 | Phosphodiesterase | BAN89425 | 2 | Ovophis okinavensis | |
| 175 | Flavin monoamine oxidase family | L-amino acid oxidase, partial | AAZ08620 | 1 | Daboia siamensis |
| 176 | L-amino acid oxidase, partial | AVX27607 | 4 | Naja atra | |
| 177 | L-amino-acid oxidase | Q4JHE1 | 5 | Pseudechis australis | |
| 178 | L-amino-acid oxidase | P0C2D5 | 1 | Protobothrops flavoviridis | |
| 179 | L-amino-acid oxidase | A8QL51 | 1 | Bungarus multicinctus | |
| 180 | L-amino-acid oxidase | Q4JHE3 | 3 | Oxyuranus scutellatus scutellatus | |
| 181 | L-amino acid oxidase, partial | AVX27607 | 4 | Naja atra | |
| 182 | L-amino-acid oxidase | A8QL58 | 6 | Naja atra | |
| 183 | L-amino-acid oxidase | Q4JHE3 | 3 | Oxyuranus scutellatus scutellatus | |
| 184 | L-amino acid oxidase precursor | AAY89682 | 2 | Pseudechis australis | |
| 185 | L-amino-acid oxidase | CAQ72894 | 1 | Echis ocellatus | |
| 186 | True venom lectin family | C-type lectin galactose-binding isoform | D2YVK1 | 2 | Hoplocephalus stephensii |
| 187 | BJcuL precursor | AAQ92957 | 1 | Bothrops jararacussu | |
| 188 | Ankyrin SOCS box (ASB) family | Ankyrin repeat and SOCS box protein 7, partial | ETE63895 | 1 | Ophiophagus hannah |
| 189 | Ankyrin repeat domain-containing protein 50, partial | ETE61041 | 1 | Ophiophagus hannah | |
| 190 | Transferrin | Transferrin | CAK18221 | 2 | Natrix natrix |
| 191 | Cobra serum albumin | Cobra serum albumin | S59517 | 1 | Naja kaouthia |
| 192 | Serum albumin precursor | S59517 | 3 | Naja naja | |
| 193 | Cobra serum albumin | CAA55333 | 3 | Naja naja | |
| 194 | Serum albumin/Alpha-fetoprotein/Afamin | Alpha-fetoprotein, partial | ETE59846 | 3 | Ophiophagus hannah |
| 195 | Leucine repeat | Leucine-rich repeat neuronal protein 4 | XP_007424790 | 1 | Python bivittatus |
| 196 | Small leucine-rich proteoglycan (SLRP) family | Decorin, partial | ETE60606 | 1 | Ophiophagus hannah |
| 197 | Leucine-rich repeat and WD repeat-containing protein, partial | ETE61323 | 1 | Ophiophagus hannah | |
| 198 | XPG/RAD2 endonuclease family | Endonuclease domain-containing 1 protein, partial | ETE59939 | 2 | Ophiophagus hannah |
| 199 | Deoxyribonuclease-2-alpha, partial | ETE73206 | 1 | Ophiophagus hannah | |
| 200 | NHS Family | NHS-like protein 1, partial | ETE71282 | 1 | Ophiophagus hannah |
| 201 | G-protein coupled receptor | G-protein coupled receptor 161 | XP_007428215 | 1 | Python bivittatus |
| 202 | Putative G-protein coupled receptor | ETE61591 | 2 | Ophiophagus hannah | |
| 203 | Melanocyte-stimulating hormone receptor, partial | ETE69163 | 1 | Ophiophagus hannah | |
| 204 | Latrophilin-2, partial | ETE73569 | 1 | Ophiophagus hannah | |
| 205 | Cadherin EGF LAG seven-pass G-type receptor 2, partial | ETE72621 | 1 | Ophiophagus hannah | |
| 206 | Putative G-protein coupled receptor, partial | ETE70400 | 1 | Ophiophagus hannah | |
| 207 | Zinc finger protein | Thioredoxin domain-containing protein 11, partial | ETE72118 | 1 | Ophiophagus hannah |
| 208 | Zinc finger protein 91-like isoform X2 | XP_007443313 | 1 | Python bivittatus | |
| 209 | Zinc finger protein 687 isoform X1 | XP_025027118 | 1 | Python bivittatus | |
| 210 | Zinc finger FYVE domain-containing protein 16, partial | ETE66135 | 1 | Ophiophagus hannah | |
| 211 | Zinc finger and BTB domain-containing protein 14, partial | XP_026555390 | 1 | Pseudonaja textilis | |
| 212 | Zinc finger protein 609 isoform X1 | XP_007426825 | 1 | Python bivittatus | |
| 213 | Ras-like protein | Ras GTPase-activating protein 3, partial | ETE71570 | 1 | Ophiophagus hannah |
| 214 | Rac GTPase-activating protein 1, partial | ETE61861 | 1 | Ophiophagus hannah | |
| 215 | Ras-related protein Rap-2a, partial | ETE66602 | 1 | Ophiophagus hannah | |
| 216 | RalA-binding protein 1, partial | ETE67818 | 1 | Ophiophagus hannah | |
| 217 | Guanylate-binding protein 1-like | XP_007444632 | 1 | Python bivittatus | |
| 218 | Glutathione peroxidase family | Glutathione peroxidase 3, partial | ETE68810 | 9 | Ophiophagus hannah |
| 219 | Protein family not assigned | Octapeptide-repeat protein T2, partial | ETE65834 | 1 | Ophiophagus hannah |
| 220 | Atrial natriuretic peptide receptor 2, partial | ETE58463 | 1 | Ophiophagus hannah | |
| 221 | Octapeptide-repeat protein T2, partial | ETE61441 | 1 | Ophiophagus hannah | |
| 222 | GAS2-like protein 2, partial | ETE67730 | 1 | Ophiophagus hannah | |
| 223 | Exocyst complex component 3, partial | ETE60130 | 1 | Ophiophagus hannah | |
| 224 | Vacuolar protein sorting-associated protein 54 | ETE70627 | 1 | Ophiophagus hannah | |
| 225 | Cohesin subunit SA-2, partial | ETE63002 | Ophiophagus hannah | ||
| 226 | Zona pellucida sperm-binding protein 3 receptor, partial | ETE59512 | 1 | Ophiophagus hannah | |
| 227 | Ubiquitin carboxyl-terminal hydrolase 32, partial | ETE63263 | 1 | Ophiophagus hannah | |
| 228 | Putative E3 ubiquitin-protein ligase UBR7 | ETE67503 | 1 | Ophiophagus hannah | |
| 229 | Mdm2-binding protein, partial | ETE64533 | 1 | Ophiophagus hannah | |
| 230 | E3 ubiquitin-protein ligase TTC3, partial | ETE73451 | 1 | Ophiophagus hannah | |
| 231 | Protocadherin-23 | XP_007425673 | 1 | Python bivittatus | |
| 232 | Nucleolar complex protein 4-like protein, partial | ETE59886 | 1 | Ophiophagus hannah | |
| 233 | Low molecular weight phosphotyrosine protein phosphatase, partial | ETE66708 | 1 | Ophiophagus hannah | |
| 234 | Major histocompatibility complex class I-related protein, partial | ETE56816 | 1 | Ophiophagus hannah | |
| 235 | Beta-2-microglobulin, partial | ETE58426 | 1 | Ophiophagus hannah | |
| 236 | GRAM domain-containing protein 1B, partial | ETE59875 | 1 | Ophiophagus hannah | |
| 237 | von Willebrand factor A domain-containing protein 3B, partial | ETE71898 | 1 | Ophiophagus hannah | |
| 238 | Homeobox protein PKNOX1, partial | XP_007435014 | 1 | Python bivittatus | |
| 239 | Homeobox protein prophet of Pit-1, partial | ETE69018 | 1 | Ophiophagus hannah | |
| 240 | Homeobox protein cut-like 2, partial | ETE71612 | 1 | ||
| 241 | Inosine-uridine preferring nucleoside hydrolase, partial | ETE68936 | 1 | Ophiophagus hannah | |
| 242 | Signal recognition particle receptor subunit beta | ETE61181 | 1 | Ophiophagus hannah | |
| 243 | Sodium channel protein type 1 subunit alpha | XP_025024892 | 1 | Python bivittatus | |
| 244 | Small serum protein-4 | BAJ14709 | 1 | Gloydius blomhoffii blomhoffii | |
| 245 | Clathrin heavy chain 1, partial | ETE68739 | 1 | Ophiophagus hannah | |
| 246 | Neutral amino acid transporter A, partial | ETE71889 | 1 | Ophiophagus hannah | |
| 247 | Bystin | ETE67512 | 1 | Ophiophagus hannah | |
| 248 | Peroxisome biogenesis factor 1-like isoform X1 | XP_025032182 | 1 | Python bivittatus | |
| 249 | Dapper-like 1, partial | ETE59781 | 1 | Ophiophagus hannah | |
| 250 | Protein patched-like 2, partial | ETE72035 | 1 | Ophiophagus hannah | |
| 251 | Keratin, type II cytoskeletal 1, partial | ETE67131 | 1 | Ophiophagus hannah | |
| 252 | Keratin, type II cytoskeletal 6A-like | XP_007441333 | 1 | Python bivittatus | |
| 253 | Cytosolic carboxypeptidase 2, partial | ETE72716 | 1 | Ophiophagus hannah | |
| 254 | NADH dehydrogenase subunit 4 | YP_003540795 | 1 | Hypsiglena ochrorhyncha klauberi | |
| 255 | Olfactory receptor 2D2-like | XP_007442854 | 1 | Python bivittatus | |
| 256 | Histone-lysine N-methyltransferase SETD1B, partial | ETE63606 | 1 | Ophiophagus hannah | |
| 257 | Helicase SRCAP, partial | ETE66458 | 1 | Ophiophagus hannah | |
| 258 | Tyrosine-protein phosphatase non-receptor type 11-like | XP_015743235 | 1 | Python bivittatus | |
| 259 | Glycerol-3-phosphate acyltransferase 4 | ETE64295 | 1 | Ophiophagus hannah | |
| 260 | NEDD4-binding protein 1, partial | ETE71789 | 1 | Ophiophagus hannah | |
| 261 | Nuclear pore complex protein, partial | ETE72717 | 1 | Ophiophagus hannah | |
| 262 | G1/S-specific cyclin-E1, partial | ETE69419 | 1 | Ophiophagus hannah | |
| 263 | Copine-3 | ETE62235 | 1 | Ophiophagus hannah | |
| 264 | Disks large-like 1, partial | ETE60775 | 1 | Ophiophagus hannah | |
| 265 | Tumor necrosis factor receptor superfamily member 11B | ETE67452 | 1 | Ophiophagus hannah | |
| 266 | Extracellular matrix protein 1, partial | ETE63009 | 3 | Ophiophagus hannah | |
| 267 | Protein PRRC2C isoform X7 | XP_025025988 | 1 | Python bivittatus | |
| 268 | Protein dispatched-like 2, partial | ETE65280 | 1 | Ophiophagus hannah | |
| 269 | Cytoplasmic FMR1-interacting protein 1 | ETE70074 | 1 | Ophiophagus hannah | |
| 270 | Sushi domain-containing protein 2 isoform X1 | XP_007439094 | 1 | Python bivittatus | |
| 271 | POU domain, class 2, transcription factor 1, partial | ETE68887 | 1 | Ophiophagus hannah | |
| 272 | Vomeronasal type-2 receptor 26-like | XP_015746172 | 1 | Python bivittatus | |
| 273 | snRNA-activating protein complex subunit 4, partial | ETE66257 | 1 | Ophiophagus hannah | |
| 274 | Small subunit processome component 20-like protein, partia | ETE62675 | 1 | Ophiophagus hannah | |
| 275 | Retrotransposon-derived protein PEG10, partial | ETE60414 | 1 | Ophiophagus hannah | |
| 276 | Heterogeneous nuclear ribonucleoprotein R | ETE70095 | 1 | Ophiophagus hannah | |
| 277 | Sacsin, partial | ETE73074 | 1 | Ophiophagus hannah | |
| 278 | Trafficking protein particle complex subunit 3 | XP_007439119 | 1 | Python bivittatus | |
| 279 | Putative protein C4orf34 | ETE61848 | 1 | Ophiophagus hannah | |
| 280 | Sulfate transporter, partial | ETE72250 | 1 | Ophiophagus hannah | |
| 281 | Solute carrier family 2, facilitated glucose transporter member 11, partial | ETE65979 | 1 | Ophiophagus hannah | |
| 282 | Solute carrier family 25 member 47, partial | ETE64737 | 1 | Ophiophagus hannah | |
| 283 | Citrate synthase, mitochondrial | ETE71902 | 1 | Ophiophagus hannah | |
| 284 | Separin, partial | ETE71706 | 1 | Ophiophagus hannah | |
| 285 | 5,6-dihydroxyindole-2-carboxylic acid oxidase, partial | ETE63759 | 1 | Ophiophagus hannah | |
| 286 | Protocadherin-15, partial | ETE73122 | 1 | Ophiophagus hannah | |
| 287 | Tumor necrosis factor receptor superfamily member 11B isoform X2 | XP_025019261 | 1 | Python bivittatus | |
| 288 | Microtubule-actin cross-linking factor 1, isoforms 1/2/3/5, partial | ETE72267 | 1 | Ophiophagus hannah | |
| 289 | Ubiquitin carboxyl-terminal hydrolase CYLD | XP_015680147 | 1 | Protobothrops mucrosquamatus | |
| 290 | Peroxidasin, partial | ETE57820 | 1 | Ophiophagus hannah | |
| 291 | Serine palmitoyltransferase small subunit B | XP_025028624 | 1 | Python bivittatus | |
| 292 | C-terminal-binding protein 1, partial | ETE64323 | 1 | Ophiophagus hannah | |
| 293 | StAR-related lipid transfer protein 13 | ETE69978 | 1 | Ophiophagus hannah | |
| 294 | Ty3b-i, partial | ETE59080 | 1 | Ophiophagus hannah | |
| 295 | E3 ubiquitin-protein ligase RNF19B, partial | ETE68153 | 1 | Ophiophagus hannah | |
| 296 | PDZ domain-containing protein 6, partial | ETE69093 | 1 | Ophiophagus hannah | |
| 297 | Nebulin, partial | ETE70906 | 2 | Ophiophagus hannah | |
| 298 | Myoferlin, partial | ETE66870 | 1 | Ophiophagus hannah | |
| 299 | Protein mago nashi-like 2 | ETE70612 | 1 | Ophiophagus hannah | |
| 300 | H(+)/Cl(-) exchange transporter 7, partial | ETE72134 | 1 | Ophiophagus hannah | |
| 301 | Membrane cofactor protein-like | XP_025021316 | 2 | Python bivittatus | |
| 302 | Holliday junction recognition protein isoform X1 | XP_025025001 | 1 | Python bivittatus | |
| 303 | Adenylate cyclase type 2, partial | ETE62750 | 1 | Ophiophagus hannah | |
| 304 | Transmembrane protein, partial | ETE59610 | 1 | Ophiophagus hannah | |
| 305 | Transmembrane protein, partial | ETE58244 | 1 | Ophiophagus hannah | |
| 306 | Type I inositol 3,4-bisphosphate 4-phosphatase | XP_015686159 | 1 | Protobothrops mucrosquamatus | |
| 307 | Complement decay-accelerating factor transmembrane isoform, partial | ETE63384 | 8 | Ophiophagus hannah | |
| 308 | NACHT, LRR and PYD domains-containing protein 6(Belongs to NLRP family) | XP_015679160 | 1 | Protobothrops mucrosquamatus | |
| 309 | Ubiquitin carboxyl-terminal hydrolase 24 | ETE67725 | 1 | Ophiophagus hannah | |
| 310 | Epiplakin, partial | ETE58258 | 1 | Ophiophagus hannah | |
| 311 | 5’ nucleotidase, partial | AXL95273 | 1 | Spilotes sulphureus | |
| 312 | GTP-binding protein 2, partial | ETE70473 | 1 | Ophiophagus hannah | |
| 313 | Transmembrane protein 41A | XP_007420693 | 1 | Python bivittatus | |
| 314 | Serine/threonine-protein kinase TAO2, partial | ETE67077 | 1 | Ophiophagus hannah | |
| 315 | Serine/threonine-protein kinase WNK1, partial | ETE61641 | 1 | Ophiophagus hannah | |
| 316 | cilia- and flagella-associated protein 57-like, partial | XP_007436852 | 1 | Python bivittatus | |
| 317 | Lymphocyte antigen 6 complex locus protein G6d | ETE61452 | 1 | Ophiophagus hannah | |
| 318 | Histamine H3 receptor, partial | ETE72972 | 1 | Ophiophagus hannah | |
| 319 | Glycerol-3-phosphate acyltransferase 1, mitochondrial, partial | ETE59719 | 1 | Ophiophagus hannah | |
| 320 | Cleft lip and palate transmembrane protein 1-like protein, partial | ETE61569 | 1 | Ophiophagus hannah | |
| 321 | Complement factor B precursor | AAR21601 | 1 | Naja kaouthia | |
| 322 | Selenocysteine lyase | XP_015669194 | 1 | Protobothrops mucrosquamatus | |
| 323 | Serine/threonine-protein kinase Nek1, partial | ETE68306 | 1 | Ophiophagus hannah | |
| 324 | Collagen alpha-1(IV) chain, partial | ETE60834 | 1 | Ophiophagus hannah | |
| 325 | DmX-like protein 2, partial | ETE63888 | 1 | Ophiophagus hannah | |
| 326 | Aldehyde dehydrogenase family 3 member B1, partial | ETE72723 | 1 | Ophiophagus hannah | |
| 327 | Putative ATP-dependent RNA helicase DHX40, partial | ETE68740 | 1 | Ophiophagus hannah | |
| 328 | Immunoglobulin Y2 heavy chain, partial | AFR33766 | 1 | Python bivittatus | |
| 329 | Myomesin-1, partial | ETE65385 | 1 | Ophiophagus hannah | |
| 330 | Cyclic AMP-dependent transcription factor ATF-1, partial | ETE65149 | 1 | Ophiophagus hannah | |
| 331 | Toll-like receptor 4, partial | ETE72495 | 1 | Ophiophagus hannah | |
| 332 | Serine palmitoyltransferase small subunit B | XP_025028624 | 1 | Python bivittatus | |
| 333 | Histone-lysine N-methyltransferase, H3 lysine-79 specific, partial | ETE65559 | 1 | Ophiophagus hannah | |
| 334 | Creatine kinase B-type, partial | ETE69249 | 1 | Ophiophagus hannah | |
| 335 | Fibroblast growth factor 3, partial | ETE69378 | 1 | Ophiophagus hannah | |
| 336 | RB1-inducible coiled-coil protein 1, partial | ETE67067 | 1 | Ophiophagus hannah | |
| 337 | Phosphoinositide 3-kinase regulatory subunit 5, partial | ETE74144 | 1 | Ophiophagus hannah | |
| 338 | Cadherin EGF LAG seven-pass G-type receptor 2, partial | ETE72621 | 1 | Ophiophagus hannah | |
| 339 | Trafficking kinesin-binding protein 1, partial | ETE68220 | 1 | Ophiophagus hannah | |
| 340 | YTH domain family protein 2 | ETE65464 | 1 | Ophiophagus hannah | |
| 341 | Vigilin, partial | ETE61946 | 1 | Ophiophagus hannah | |
| 342 | 39S ribosomal protein L44, mitochondrial, partial | ETE68399 | 1 | Ophiophagus hannah | |
| 343 | Pseudouridine-5’-monophosphatase, partial | ETE71697 | 1 | Ophiophagus hannah | |
| 344 | Kelch-like protein 13, partial | ETE71947 | 1 | Ophiophagus hannah | |
| 345 | Maleylacetoacetate isomerase | ETE68752 | 1 | Ophiophagus hannah | |
| 346 | Neurexophilin-2, partial | ETE71784 | 1 | Ophiophagus hannah | |
| 347 | Myocyte-specific enhancer factor 2A isoform X1 | XP_007425135 | 1 | Python bivittatus | |
| 348 | Membrane cofactor protein-like isoform X1 | XP_015743425 | 1 | Python bivittatus | |
| 349 | Ninein-like protein, partial | ETE70166 | 1 | Ophiophagus hannah | |
| 350 | Keratin, type I cytoskeletal 19, partial | ETE70217 | 1 | Ophiophagus hannah | |
| 351 | Intraflagellar transport protein 88-like protein | ETE73657 | 1 | Ophiophagus hannah | |
| 352 | Complement receptor type 2, partial | ETE63383 | 1 | Ophiophagus hannah | |
| 353 | Complement decay-accelerating factor, partial | ETE59511 | 1 | Ophiophagus hannah | |
| 354 | Keratin, type II cytoskeletal 5-like | XP_025030548 | 1 | Python bivittatus | |
| 355 | 7-dehydrocholesterol reductase, partial | ETE67784 | 1 | Ophiophagus hannah | |
| 356 | La-related protein 4B | ETE62671 | 1 | Ophiophagus hannah | |
| 357 | Intelectin-1a, partial | ETE57886 | 1 | Ophiophagus hannah | |
| 358 | Cation-independent mannose-6-phosphate receptor | ETE64374 | 2 | Ophiophagus hannah | |
| 359 | Cerebellin-4 | ETE65277 | 1 | Ophiophagus hannah | |
| 360 | C3 and PZP-like alpha-2-macroglobulin domain-containing protein 8, partial | ASU45032 | 1 | Ophiophagus hannah | |
| 361 | Neuronal PAS domain-containing protein 2, partial | ETE63668 | 1 | Ophiophagus hannah | |
| 362 | Interferon-induced transmembrane protein 10, partial | ETE66904 | 1 | Ophiophagus hannah | |
| 363 | Myotubularin-related protein 11, partial | ETE72068 | 1 | Ophiophagus hannah | |
| 364 | Tyrosyl-DNA phosphodiesterase 2 | XP_026525751 | 1 | Notechis scutatus | |
| 365 | Phosphoinositide 3-kinase regulatory subunit 5, partial | ETE74144 | 1 | Ophiophagus hannah |
| S. No. | Protein Family | Protein | Accession Code | Number of Matched Peptides | Homology with Protein from the Venom of Snake Species | |
|---|---|---|---|---|---|---|
| 1 | 3FTX (Neurotoxin | Neurotoxin homolog NL1 | Q9DEQ3 | 1 | Naja atra | |
| 2 | Short neurotoxin SNTX-1 | A6MFK6 | 1 | Demansia vestigiata | ||
| 3 | Neurotoxin II | P01427 | 1 | Naja oxiana | ||
| 4 | Cobrotoxin-b | P80958 | 1 | Naja atra | ||
| 5 | Alpha-cobratoxin | P01391 | 3 | Naja kaouthia | ||
| 6 | Weak toxin 2 | Q8AY50 | 2 | Bungarus candidus | ||
| 7 | Weak neurotoxin 6 | O42256 | 1 | Naja sputatrix | ||
| 8 | Weak neurotoxin 7 | P29181 | 2 | Naja naja | ||
| 9 | Weak toxin S4C11 | P01400 | 1 | Naja melanoleuca | ||
| 10 | Muscarinic toxin-like protein 3 | P82464 | 4 | Naja kaouthia | ||
| 11 | Muscarinic toxin-like protein 2 | P82463 | 4 | Naja kaouthia | ||
| 12 | Muscarinic toxin-like protein | Q9W727 | 1 | Bungarus multicinctus | ||
| 13 | Three-finger toxin precursor, partial | ADN67582 | 1 | Naja atra | ||
| 14 | Three-finger toxin precursor, partial | ADN67582 | 1 | Naja atra | ||
| 15 | 3FTXs (cytotoxins) | Cytotoxin Vc-5 | Q9PS34 | 2 | Naja oxiana | |
| 16 | Cytotoxin homolog | P14541 | 1 | Naja kaouthia | ||
| 17 | Cytotoxin homolog 5V | Q9W716 | 1 | Naja atra | ||
| 18 | Cytotoxin SP15c | P60308 | 1 | Naja atra | ||
| 19 | Cytotoxin 8 | P86540 | 2 | Naja naja | ||
| 20 | Cytotoxin 1 | P01447 | 2 | Naja naja | ||
| 21 | Cardiotoxin 7a | Q91126 | 6 | Naja atra | ||
| 22 | Cardiotoxin 1e | AAA90960 | 2 | Naja atra | ||
| 23 | Venom Complement C3-like | Venom factor | AAX86641 | 1 | Austrelaps superbus | |
| 24 | Cobra venom factor | Q91132 | 10 | Naja kaouthia | ||
| 25 | A.superbus venom factor 1 | Q0ZZJ6 | 1 | Austrelaps superbus | ||
| 26 | Cobra venom factor alpha chain | Q91132 | 1 | Naja kaouthia | ||
| 27 | Cobra venom factor 1, partial | AXL96620 | 6 | Ahaetulla prasina | ||
| 28 | Cobra venom factor, partial | AWX67646 | 2 | Boiga irregularis | ||
| 29 | Ophiophagus venom factor | I2C090 | 1 | Ophiophagus hannah | ||
| 30 | Venom Kunitz-type family | Kunitz-type serine protease inhibitor | P20229 | 2 | Naja naja | |
| 31 | BPTI/Kunitz domain-containing protein-like | XP_026546510 | 1 | Notechis scutatus | ||
| 32 | Kunitz/BPTI-like toxin | XP_026579467 | 1 | Pseudonaja textilis | ||
| 33 | natriuretic peptide family | Natriuretic peptide PaNP-c precursor, partial | AAZ82822 | 1 | Pseudechis australis | |
| 34 | NGF-beta family | Venom nerve growth factor 2 | Q5YF89 | 5 | Naja sputatrix | |
| 35 | Nerve growth factor, partial | AAR24530 | 1 | Bitis gabonica | ||
| 36 | Nerve growth factor | BAN82142 | 4 | Ovophis okinavensis | ||
| 37 | Venom nerve growth factor 2 | Q3HXX9 | 1 | Hoplocephalus stephensii | ||
| 38 | ohanin/vespryn family. | Thaicobrin | P82885 | 1 | Naja kaouthia | |
| 39 | venom PRY-SPRY domain-containing protein, partial | AHZ08803 | 1 | Micropechis ikaheca | ||
| 40 | CRISP | Cysteine-rich venom protein natrin-1 | Q7T1K6 | 3 | Naja atra | |
| 41 | Cysteine-rich secretory protein 1, partial | AXL96607 | 1 | Ahaetulla prasina | ||
| 42 | Cysteine-rich venom protein ophanin | Q7ZT98 | 1 | Ophiophagus hannah | ||
| 43 | Cysteine-rich venom protein, partial | BAP39957 | 1 | Protobothrops flavoviridis | ||
| 44 | Cysteine-rich venom protein 2 | Q7ZZN8 | 1 | Naja atra | ||
| 45 | Phosoholipase A2 | Acidic phospholipase A2 3 | P60045 | 1 | Naja sagittifera | |
| 46 | Acidic phospholipase A2 2 | P00597 | 1 | Naja kaouthia | ||
| 47 | Phospholipase a2 | CAA45372 | 3 | Naja naja | ||
| 48 | Neutral phospholipase A2 paradoxin-like beta chain | Q45Z46 | 2 | Oxyuranus microlepidotus | ||
| 49 | Phospholipase A2 | AHZ08810 | 1 | Micropechis ikaheca | ||
| 50 | Phospholipase A2 | AAA66029.1 | 1 | Naja naja | ||
| 51 | Acidic phospholipase A2 2 | P15445 | 1 | Naja naja | ||
| 52 | Acidic phospholipase A2 1 | P00596 | 6 | Naja kaouthia | ||
| 53 | Acidic phospholipase A2 1 | Q9W7J4 | 1 | Pseudonaja textilis | ||
| 54 | Basic phospholipase A2 T1-2 A chain | P84472 | 1 | Bungarus candidus | ||
| 55 | Acidic phospholipase A2 C | Q92086 | 11 | Naja sputatrix | ||
| 56 | Acidic phospholipase A2 1 | P00598 | 1 | Naja naja | ||
| 57 | Acidic phospholipase A2 beta-bungarotoxin A4 chain | P17934 | 1 | Bungarus multicinctus | ||
| 58 | Phospholipase A2 3 | P21792 | 1 | Micrurus nigrocinctus | ||
| 59 | Phospholipase B | Phospholipase B, partial | AXL95274 | 1 | Spilotes sulphureus | |
| 60 | Phospholipase B1, partial | AXL96606 | 2 | Ahaetulla prasina | ||
| 61 | Phospholipase B1, membrane-associated | XP_02653746 | 1 | Notechis scutatus | ||
| 62 | SVMP | Snake venom metalloproteinase | D3TTC2 | 4 | Naja atra | |
| 63 | Snake venom metalloproteinase | F8RKW1 | 1 | Drysdalia coronoides | ||
| 64 | Snake venom metalloproteinase | Q9PVK7 | 1 | Naja kaouthia | ||
| 65 | Disintegrin and metalloproteinase domain-containing protein 20, partial | ETE72945 | 1 | Ophiophagus hannah | ||
| 66 | Disintegrin and metalloproteinase domain-containing protein 21, partial | ETE71596 | 1 | Ophiophagus hannah | ||
| 67 | disintegrin and metalloproteinase domain-containing protein 10-like, partial | XP_026580760 | 1 | Pseudonaja textilis | ||
| 68 | P-III snake venom metalloprotease, partial | AHZ08819 | 1 | Micropechis ikaheca | ||
| 69 | Zinc metalloproteinase-disintegrin-like kaouthiagin-like | D3TTC1 | 7 | Naja atra | ||
| 70 | Zinc metalloproteinase-disintegrin-like atrase-A | D5LMJ3 | 14 | Naja atra | ||
| 71 | Hemorrhagic metalloproteinase-disintegrin-like kaouthiagin | P82942 | 2 | Naja kaouthia | ||
| 72 | metalloproteinase 7, partial | AXL96626 | 1 | Ahaetulla prasina | ||
| 73 | metalloproteinase, partial | AWX67576 | 1 | Boiga irregularis | ||
| 74 | Snake venom metalloproteinase-disintegrin-like mocarhagin | Q10749 | 3 | Naja mossambica | ||
| 75 | Snake venom metalloproteinase | Q9W6M5 | 1 | Deinagkistrodon acutus | ||
| 76 | Snake venom serine proteinase (peptidase S1 family) | Tissue-type plasminogen activator, partial | ETE66683 | 3 | Ophiophagus hannah | |
| 77 | tissue-type plasminogen activator, partial | XP_026544671 | 2 | Notechis scutatus | ||
| 78 | Snake venom serine protease 3 | O13058 | 1 | Protobothrops flavoviridis | ||
| 79 | Serine protease 27, partial | ETE64653 | 1 | Ophiophagus hannah | ||
| 80 | Thrombin-like enzyme TLP | P86545 | 1 | Naja naja | ||
| 81 | Snake venom serine protease 3 | AAG10790 | 1 | Protobothrops jerdonii | ||
| 82 | Snake venom serine protease Dav-PA | Q9I8X1 | 1 | Deinagkistrodon acutus | ||
| 83 | serine protease 53 | XP_026576912 | 1 | Pseudonaja textilis | ||
| 84 | 5’-nucleotidase family | 5’ nucleotidase, partial | AXL95273 | 1 | Spilotes sulphureus | |
| 85 | Aminopeptidase | aminopeptidase N isoform X2 | XP_026565037 | 4 | Pseudonaja textilis | |
| 86 | type-B carboxylesterase/lipase | acetylcholinesterase | XP_026549820 | 1 | Notechis scutatus | |
| 87 | Phosphodiesterase | Phosphodiesterase | BAN89425 | 2 | Ovophis okinavensis | |
| 88 | Phosphodiesterase partial | ALA20853 | 1 | Macropisthodon rudis | ||
| 89 | Phosphodiesterase partial | AXL96599 | 1 | Borikenophis portoricensis | ||
| 90 | Ectonucleotide pyrophosphatase/phosphodiesterase family member 3 isoform X2 | XP_026561286 | 2 | Pseudonaja textilis | ||
| 91 | Snake venom Phosphodiesterase | A0A2D0TC04 | 2 | Naja atra | ||
| 92 | Flavin monoamine oxidase family | L-amino acid oxidase, partial | AVX27607 | 7 | Naja atra | |
| 93 | L-amino-acid oxidase | XP_026538830 | 4 | Notechis scutatus | ||
| 94 | L-amino-acid oxidase | Q4JHE3 | 1 | Oxyuranus scutellatus scutellatus | ||
| 95 | L-amino-acid oxidase | Q4JHE1 | 1 | Pseudechis australis | ||
| 96 | L-amino-acid oxidase | A8QL58 | 1 | Naja atra | ||
| 97 | True venom lectin family | C-type lectin Cal | P21963 | 1 | Crotalus atrox | |
| 98 | Glutathione peroxidase | Glutathione peroxidase 3, partial | ETE68810 | 1 | Ophiophagus hannah | |
| 99 | Glutathione peroxidase 3 isoform X1 | XP_026541908 | 1 | Notechis scutatus | ||
| 100 | Glutathione peroxidase 3 isoform X1 | XP_026552406 | 1 | Pseudonaja textilis | ||
| 101 | Leucine repeat | Leucine-rich repeat and death domain-containing protein 1 | XP_026543987 | 1 | Notechis scutatus | |
| 102 | TNF receptor superfamily | Tumor necrosis factor receptor superfamily member 11B | XP_026545353 | 1 | Notechis scutatus | |
| 103 | Tumor necrosis factor receptor superfamily member 11B | XP_026559377 | 1 | Pseudonaja textilis | ||
| 104 | Tumor necrosis factor receptor superfamily member 11B | ETE67452 | 1 | Ophiophagus hannah | ||
| 105 | Intermediate filament family | Keratin, type II cytoskeletal 1, partia | ETE67131 | 1 | Ophiophagus hannah | |
| 106 | Keratin, type II cytoskeletal 4-like | XP_026539658 | 1 | Notechis scutatus | ||
| 107 | Keratin, type II cytoskeletal 5, partial | ETE59039 | 1 | Ophiophagus hannah | ||
| 108 | Keratin, type II cytoskeletal 5, partial | ETE59038 | 2 | Ophiophagus hannah | ||
| 109 | Keratin, type II cytoskeletal 1-like | XP_026573193 | 1 | Pseudonaja textilis | ||
| 110 | Keratin, type I cytoskeletal 19, partial | ETE70217 | 2 | Ophiophagus hannah | ||
| 111 | Keratin, type I cytoskeletal 18-like isoform X1 | XP_026521302 | 1 | Notechis scutatus | ||
| 112 | Serpin Family | Serpin B5, partial | ETE65002 | 1 | Ophiophagus hannah | |
| 113 | Ankyrin repeat domain | M-phase phosphoprotein 8, partial | ETE73652 | 1 | Ophiophagus hannah | |
| 114 | Zinc finger containing proteins | Zinc finger protein, partial | ETE62318 | 1 | Ophiophagus hannah | |
| 115 | Zinc finger protein, partial | ETE62303 | 1 | Ophiophagus hannah | ||
| 116 | Zinc finger protein 804A | XP_026552505 | 1 | Pseudonaja textilis | ||
| 117 | Zinc finger SWIM domain-containing protein 6 | XP_026572863 | 1 | Pseudonaja textilis | ||
| 118 | Zinc finger MYM-type protein 2 isoform X1 | XP_026564670 | 1 | Pseudonaja textilis | ||
| 119 | Zinc finger BED domain-containing protein 1 | XP_026522663 | 1 | Notechis scutatus | ||
| 120 | NHS Family | NHS-like protein 1 isoform X1 | XP_026561348 | 1 | Pseudonaja textilis | |
| 121 | Protein family not assigned | Holliday junction recognition protein | XP_026519764 | 1 | Notechis scutatus | |
| 122 | N-acetylgalactosaminyltransferase 7 isoform X1 | XP_026555474 | 1 | Pseudonaja textilis | ||
| 123 | PHD finger protein 3 | XP_026520899 | 1 | Notechis scutatus | ||
| 124 | Sulfhydryl oxidase 1(contains FAD binding domain) | ETE70041 | 1 | Ophiophagus hannah | ||
| 125 | C-C chemokine receptor type 10, partial | ETE65216 | 1 | Ophiophagus hannah | ||
| 126 | Cytosolic carboxypeptidase 2 | XP_026521145 | 1 | Notechis scutatus | ||
| 127 | SUMO-specific isopeptidase USPL1 isoform X1 | XP_026564646 | 1 | Pseudonaja textilis | ||
| 128 | Protein VPRBP | ETE70381 | 1 | Ophiophagus hannah | ||
| 129 | Cilia- and flagella-associated protein 97 | XP_026553667 | 1 | Pseudonaja textilis | ||
| 130 | lpxK, partial | ETE68446 | 1 | Ophiophagus hannah | ||
| 131 | Zinc phosphodiesterase ELAC protein 2, partial | ETE70777 | 1 | Ophiophagus hannah | ||
| 132 | NHS-like protein 1 isoform X1 | XP_026561348 | 1 | Pseudonaja textilis | ||
| 133 | Pro-cathepsin H | XP_026565144 | 1 | Pseudonaja textilis | ||
| 134 | C4b-binding protein alpha chain-like isoform X1 | XP_026571379 | 2 | Pseudonaja textilis | ||
| 135 | Janus kinase and microtubule-interacting protein 3 isoform X1 | XP_026566312 | 1 | Pseudonaja textilis | ||
| 136 | WD and tetratricopeptide repeats protein 1 | XP_026558310 | 1 | Pseudonaja textilis | ||
| 137 | Pro-cathepsin H | XP_026565144 | 1 | Pseudonaja textilis | ||
| 138 | C4b-binding protein alpha chain-like isoform X1 | XP_026571379 | 2 | Pseudonaja textilis | ||
| 139 | Janus kinase and microtubule-interacting protein 3 isoform X1 | XP_026566312 | 1 | Pseudonaja textilis | ||
| 140 | WD and tetratricopeptide repeats protein 1 | XP_026558310 | 1 | Pseudonaja textilis | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manuwar, A.; Dreyer, B.; Böhmert, A.; Ullah, A.; Mughal, Z.; Akrem, A.; Ali, S.A.; Schlüter, H.; Betzel, C. Proteomic Investigations of Two Pakistani Naja Snake Venoms Species Unravel the Venom Complexity, Posttranslational Modifications, and Presence of Extracellular Vesicles. Toxins 2020, 12, 669. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110669
Manuwar A, Dreyer B, Böhmert A, Ullah A, Mughal Z, Akrem A, Ali SA, Schlüter H, Betzel C. Proteomic Investigations of Two Pakistani Naja Snake Venoms Species Unravel the Venom Complexity, Posttranslational Modifications, and Presence of Extracellular Vesicles. Toxins. 2020; 12(11):669. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110669
Chicago/Turabian StyleManuwar, Aisha, Benjamin Dreyer, Andreas Böhmert, Anwar Ullah, Zia Mughal, Ahmed Akrem, Syed Abid Ali, Hartmut Schlüter, and Christian Betzel. 2020. "Proteomic Investigations of Two Pakistani Naja Snake Venoms Species Unravel the Venom Complexity, Posttranslational Modifications, and Presence of Extracellular Vesicles" Toxins 12, no. 11: 669. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110669