Transcriptomic and Proteomic Analysis Reveals Mechanisms of Patulin-Induced Cell Toxicity in Human Embryonic Kidney Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
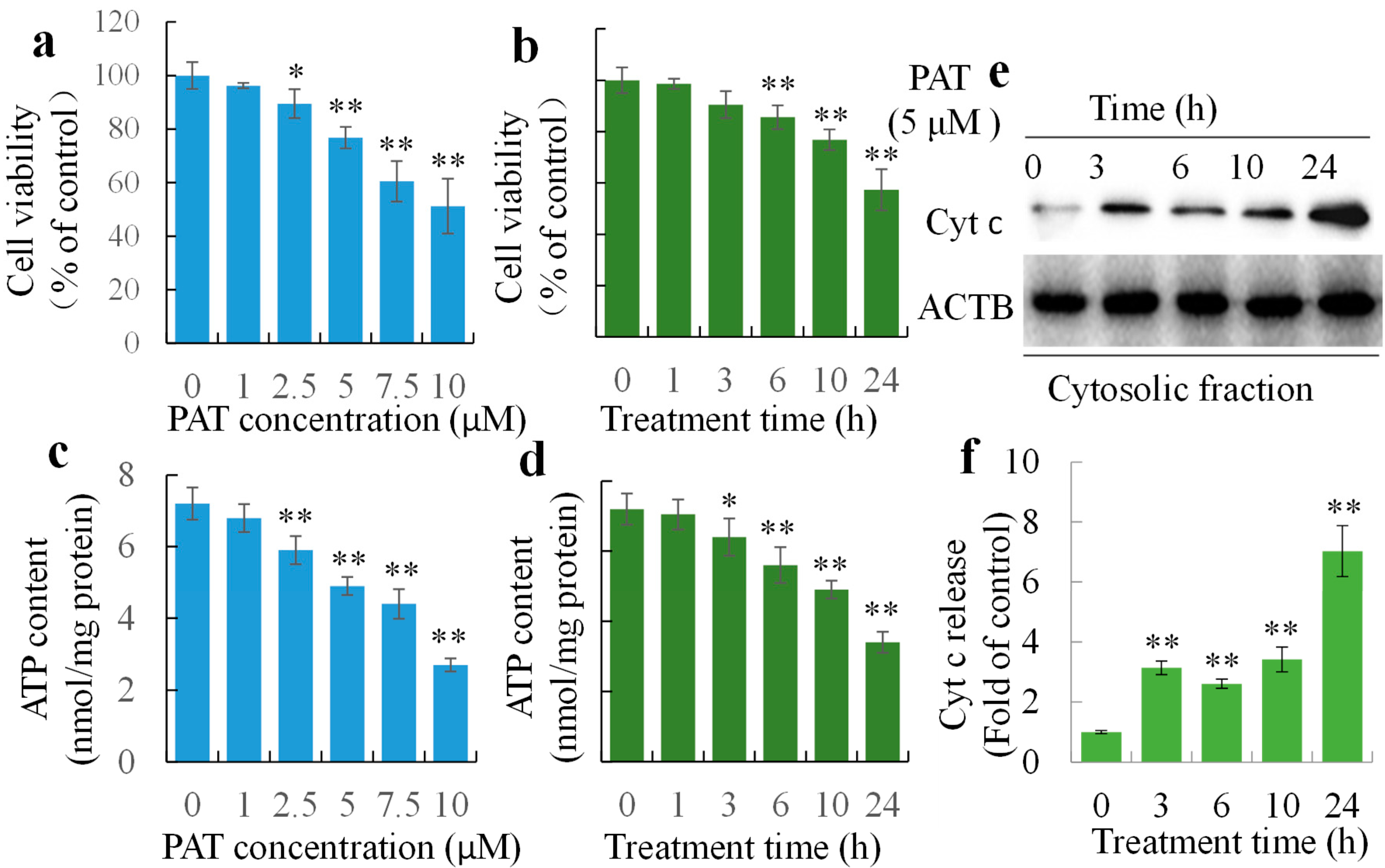
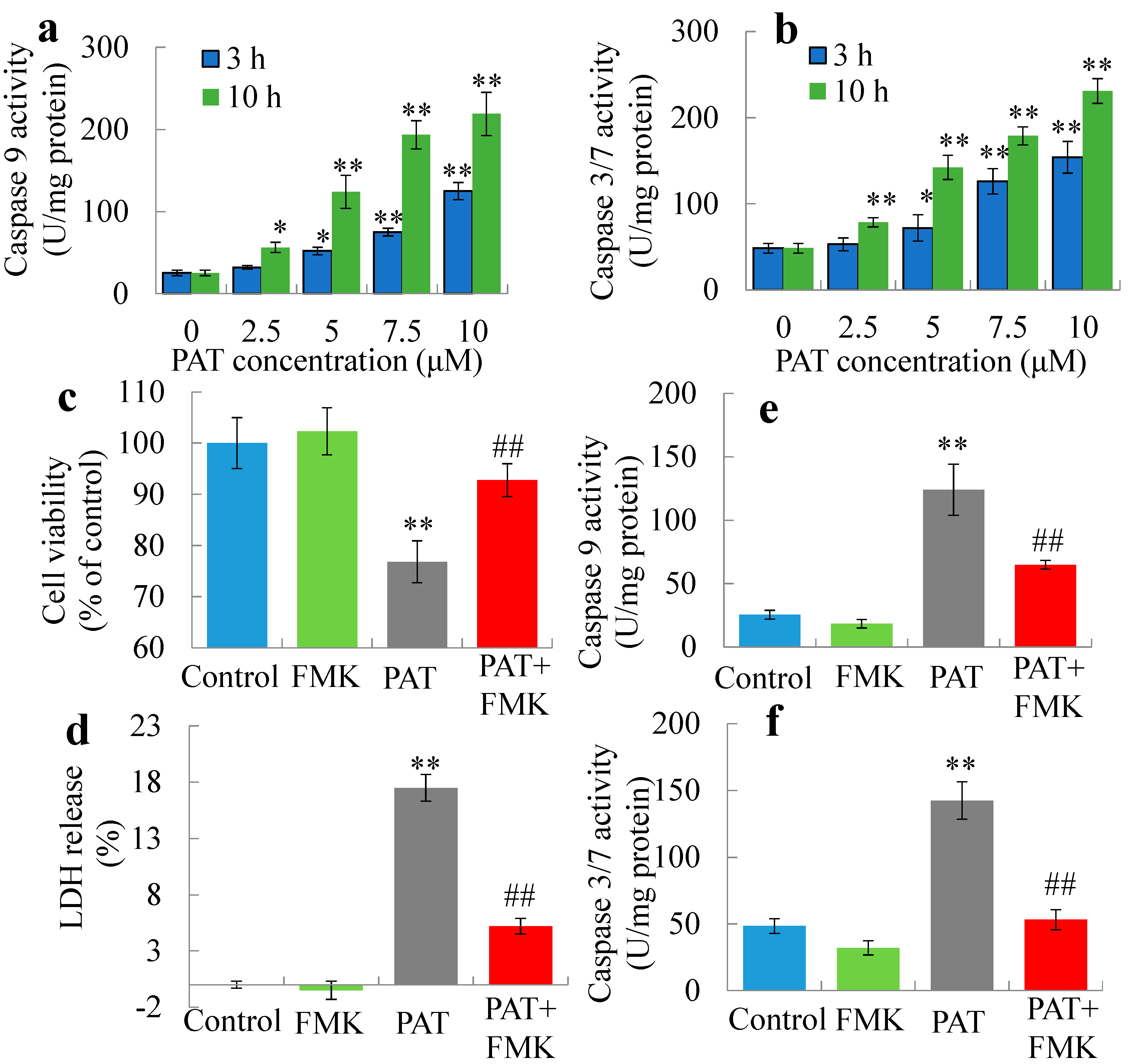
2.1. PAT Triggers Caspase-Dependent Cell Death via the Intrinsic Apoptotic Pathway
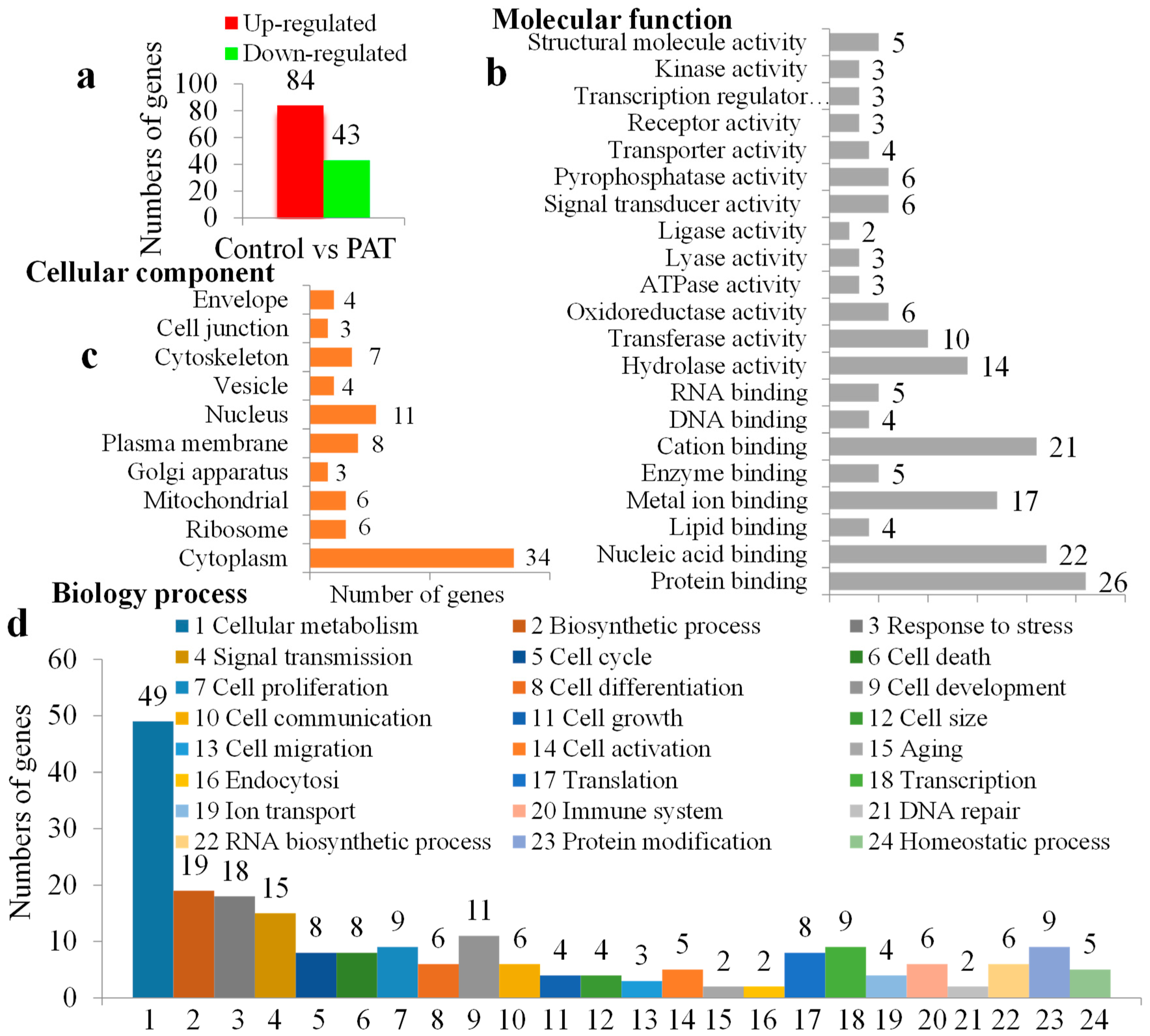
2.2. Transcriptome Analysis
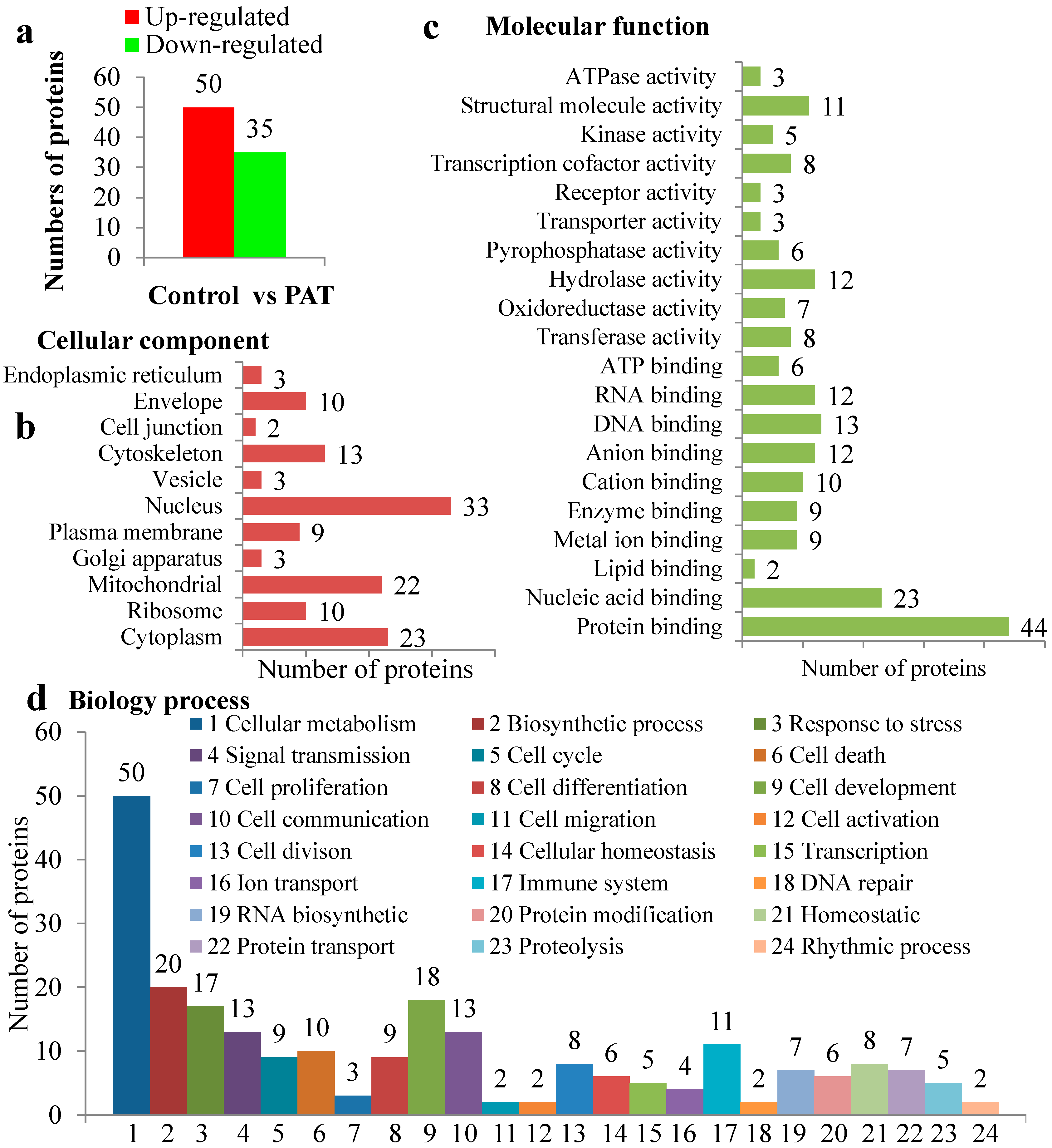
2.3. Proteomic Analysis
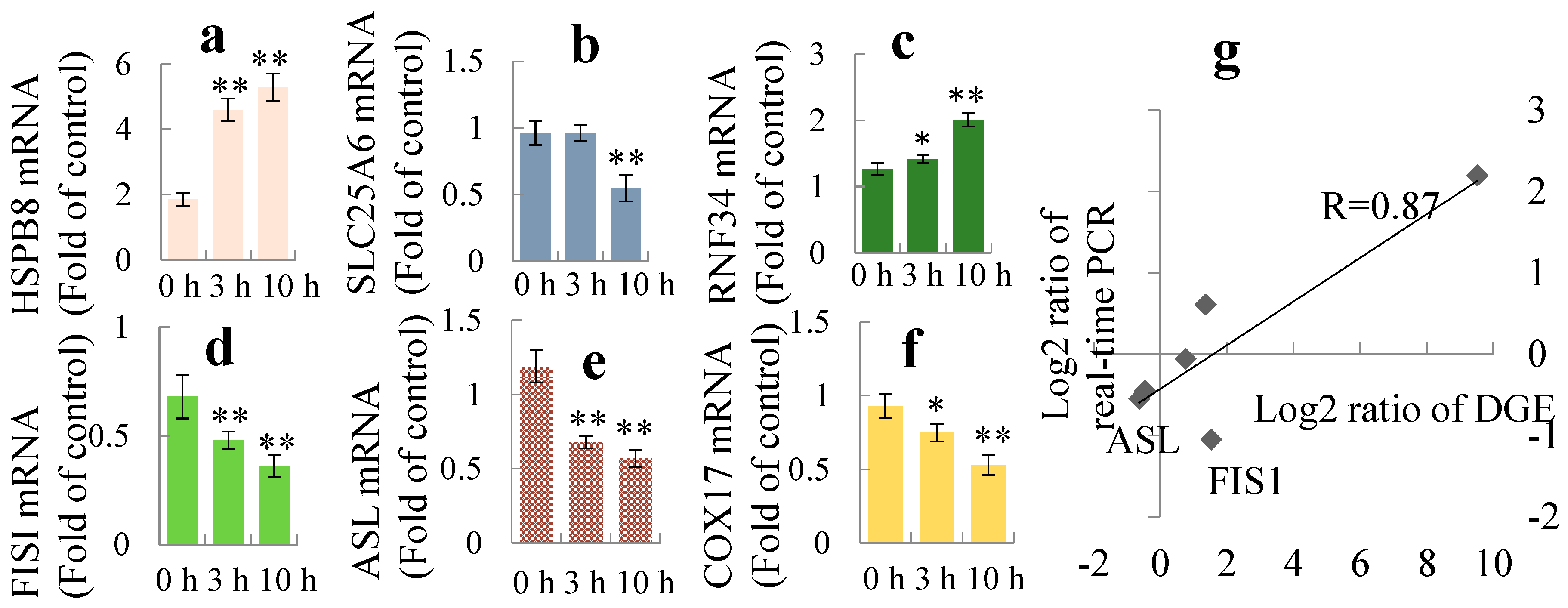
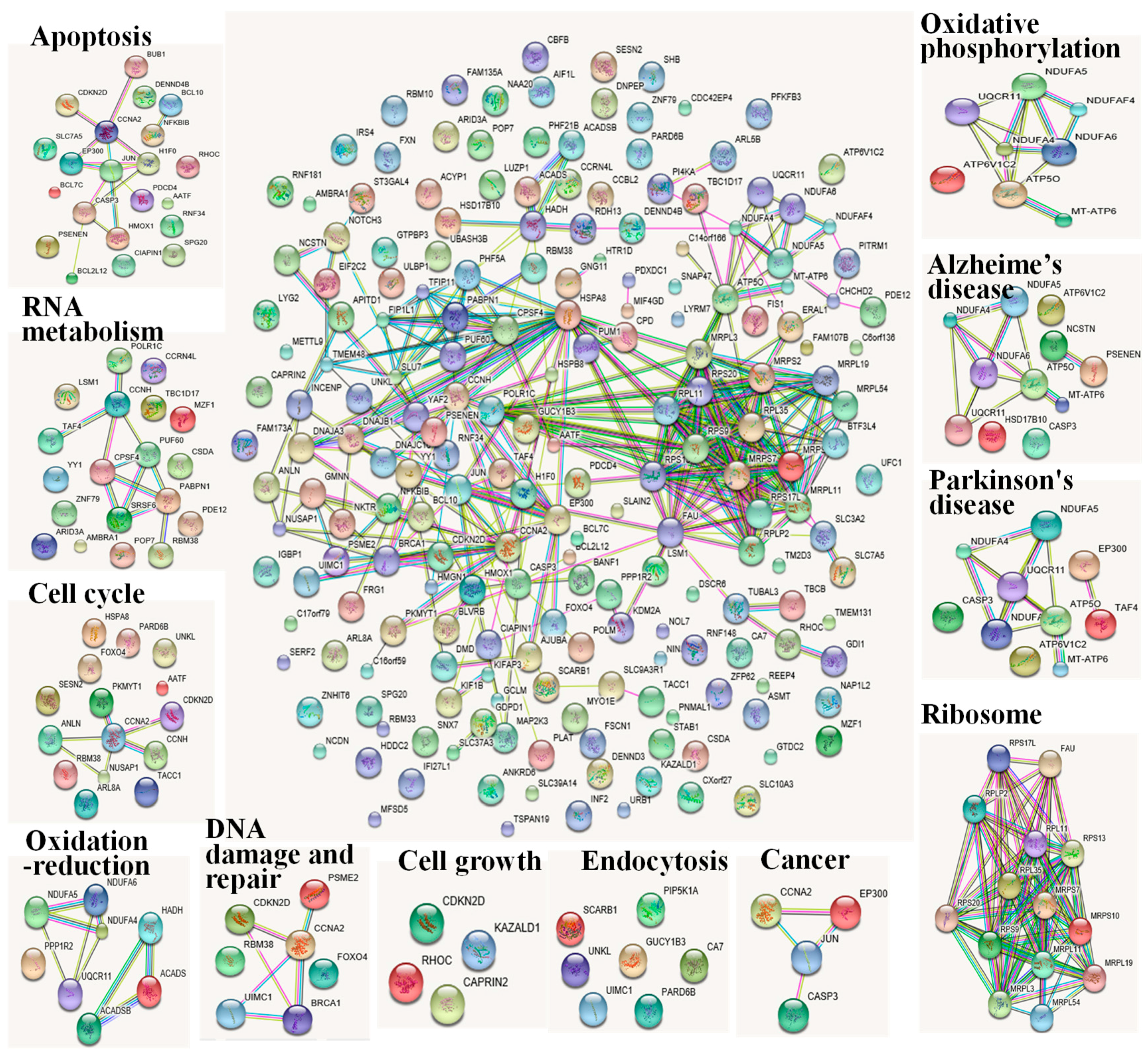
2.4. Integrated Transcriptome and Proteome Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cell Culture and PAT Treatment
5.2. Measuring Cell Viability, LDH Leakage and Caspase Activities
5.3. Digital Gene Expression (DGE)
5.4. iTRAQ Protein Profiling
5.5. Reverse Transcript and Real-Time Quantitative PCR Analysis of mRNA
5.6. Western Blot Analysis of Protein Expression
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Puel, O.; Galtier, P.; Oswald, I.P. Biosynthesis and toxicological effects of patulin. Toxins 2010, 2, 613–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.Q.; Chen, Y.; Zong, Y.Y.; Shang, Y.J.; Zhang, Z.Q.; Xu, X.D.; Wang, X.; Long, M.Y.; Tian, S.P. Dissection of patulin biosynthesis, spatial control and regulation mechanism in Penicillium expansum: Pathway control of patulin biosynthesis. Environ. Microbiol. 2019, 21, 1124–1139. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Iwahashi, Y. Gene expression profiles of Yeast Saccharomyces cerevisiae sod1 caused by patulin toxicity and evaluation of recovery potential of ascorbic acid. J. Agric. Food Chem. 2011, 59, 7145–7154. [Google Scholar] [CrossRef] [PubMed]
- Assunção, R.; Alcito, P.; Kleveland, C.R.; Lea, T.E. Characterization of in vitro effects of patulin on intestinal epithelial and immunes. Toxicol. Lett. 2016, 250, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Riley, R.T.; Showker, J.L. The mechanism of patulin’s cytotoxicity and its antioxidant activity of indoletetramic acids. Toxicol. Appl. Pharmacol. 1991, 109, 108–126. [Google Scholar] [CrossRef]
- Boussabbeh, M.; Ben, S.I.; Guilbert, A.; Bacha, H.; Abid-Essefi, S.; Lemaire, C. Patulin induces apoptosis through ROS-mediated endoplasmic reticulum stress pathway. Toxicol. Sci. 2015, 144, 328–337. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.H.; Yu, F.Y.; Wu, T.S.; Li, S.Y.; Su, M.C.; Wang, M.C.; Shih, S.M. Evaluation of genotoxic risk and oxidative DNA damage in mammalian cells exposed to mycotoxins, patulin and citrinin. Toxicol. Appl. Pharmacol. 2003, 191, 255–263. [Google Scholar] [CrossRef]
- Saxena, N.; Ansari, K.M.; Kumar, R.; Dhawan, A.; Dwivedi, P.D.; Das, M. Patulin causes DNA damage leading to cell cycle arrest and apoptosis through modulation of Bax, p53, p21/WAF1 protein in skin mice. Toxicol. Appl. Pharmacol. 2009, 234, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Peng, X.; Li, G.; Xu, Y.; Xia, X. Oxidative stress is involved in patulin induced apoptosis in HEK293 cells. Toxicon 2015, 94, 1–7. [Google Scholar] [CrossRef]
- Jin, H.; Yin, S.; Song, X.; Zhang, E.; Fan, L.; Hu, H. P53 activation contributes to patulin-induced nephrotoxicity via modulation of reactive oxygen species generation. Sci. Rep. 2015, 6, 24455. [Google Scholar] [CrossRef]
- Yang, G.; Bai, Y.; Wu, X.R.; Sun, X.C.; Sun, M.; Liu, X.; Yao, X.F.; Zhang, C.; Chu, Q.; Jiang, L.P.; et al. Patulin induced ROS-dependent autophagic cell death in human hepatoma G2 cells. Chem.-Biol. Interact. 2018, 288, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Kocharunchitt, C.; King, T.; Gobius, K.; Bowman, J.P. Integrated transcriptomic and proteomics analysis of the physiological response of Escherichia coli O157:H7 Sakai to steady-state conditions of cold and water activity stress. Mol. Cell. Proteom. 2012, 11, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.S.; Jiang, J.; Zhang, H.X.; Wang, J.P.; Cai, H.; Li, C.; Li, K.B.; Liu, J.; Guo, X.J.; Zou, G.X.; et al. Integrated transcriptional and proteomic analysis with vitro biochemical assay reveal the important role of CYP3A46 in T-2 toxin hydroxylation in porcine primary hepatocytes. Mol. Cell. Proteom. 2011, 10, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, S.; Kiyosawa, N.; Ando, Y.; Watanabe, Y.K.; Niino, N.; Ito, K.; Yamoto, T.; Manabe, S.; Sanbuissho, A. Hepatic transcriptome and proteome responses against diethyl maleate-induced glutathione depletion in the rat. Arch. Toxicol. 2011, 85, 1045–1056. [Google Scholar] [CrossRef]
- Mahfoud, R.; Maresca, M.; Garmy, N.; Fantini, J. The mycotoxin patulin alters the barrier function of the intestinal epithelium: Mechanism of action of the toxin and protective effects of glutathione. Toxicol. Appl. Pharmacol. 2002, 181, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.J.; Jin, C.N.; Gan, J.; Wang, X.R.; Shi, Z.Q.; Xia, X.D.; Peng, X.L. Apigenin attenuates patulin-induced apoptosis in HEK293 cells by modulating ROS-mediated mitochondrial dysfunction and caspase signal pathway. Toxicon 2017, 137, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.M.; Jiang, L.P.; Geng, C.Y.; Cao, J.; Zhong, L.F. Patulin-induced genotoxicity and modulation of glutathione in HepG2 cells. Toxicon 2009, 53, 584–586. [Google Scholar] [CrossRef]
- Nakagawa, T.; Zhu, H.; Morishima, N.; Li, E.; Xu, J.; Yankner, B.A.; Yuan, J.Y. Caspase-12 mediated endoplasmic-reticulum-specific apoptosis and cytotoxicity by amyloid-beta. Nature 2000, 403, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Asker, N.; Kristiansson, E.; Albertsson, E.; Larsson, J.; Förlin, L. Hepatic transcriptome profiling indicates differential mRNA expression of apoptosisand immune related genes in eelpout (Zoarcesviviparus) caught at Göteborgharbor, Sweden. Aquat. Toxicol. 2013, 130, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.S.; Liao, Y.C.; Yu, F.Y.; Chang, C.H.; Liu, B.H. Mechanism of patulin-induced apoptosis in human leukemia cells (HL-60). Toxicol. Lett. 2008, 183, 105–111. [Google Scholar] [CrossRef]
- Kwon, O.; Soung, N.K.; Thimmegowsa, N.R.; Jeong, S.J.; Jang, J.H.; Moon, D.O.; Chung, J.K.; Lee, K.S.; Kwon, Y.T.; Erikson, R.L.; et al. Patulin induces colorectal cancer cells apoptosis through EGR-1 dependent ATF3 up-regulation. Cell. Signal. 2012, 24, 943–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boussabbeh, M.; Salem, I.B.; Belguesmi, F.; Neffati, F.; Najjar, M.F.; Abid-Essefi, S.A.; Bacha, H. Crocin protects the liver and kidney from patulin-induced apoptosis in vivo. Environ. Sci. Pollut. Res. 2016, 23, 9799–9808. [Google Scholar] [CrossRef]
- Jayashree, G.V.; Krupashree, K.; Rachitha, P.; Khanum, F. Patulin Induced Oxidative Stress Mediated Apoptotic Damage in Mice, and its Modulation by Green Tea Leaves. J. Clin. Exp. Hepatol. 2017, 7, 127–134. [Google Scholar] [CrossRef]
- Wang, X.R.; Jin, C.N.; Zhong, Y.J.; Li, X.; Han, J.H.; Xue, W.; Wu, P.; Xia, X.D.; Peng, X.L. Glutathione reduction of patulin-evoked cytotoxicity in HEK293 cells by the prevention of oxidative damage and the mitochondrial apoptotic pathway. J. Agric. Food Chem. 2018, 66, 7775–7785. [Google Scholar] [CrossRef]
- Wang, Y.; Peng, X.L.; Yang, Z.J.; Zhao, W.W.; Xu, W.T.; Hao, J.R.; Wu, W.H.; Shen, X.L.; Luo, Y.B.; Huang, K.L. iTRAQ mitoproteome analysis reveals mechanisms of programmed cell death in Arabidopsis thaliana induced by ochratoxin A. Toxins 2017, 9, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handy, D.E.; Loscalzo, J. Redox regulation of mitochondrial function. Antioxid. Redox Signal. 2012, 16, 1323–1367. [Google Scholar] [CrossRef] [PubMed]
- Rohlena, J.; Dong, L.F.; Neuzil, J. Targeting the mitochondrial electron transport chain complexes for the induction of apoptosis and cancer treatment. Curr. Pharm. Biotechnol. 2013, 14, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Fang, J.; Leonard, S.S.; Krishna, R.K.M. Cadmium inhibits the electron transfer chain and induces reactive oxygen species. Free Radic. Biol. Med. 2004, 36, 1434–1443. [Google Scholar] [CrossRef] [PubMed]
- Hüttemann, M.; Lee, I.; Pecinova, A.; Pecina, P.; Przyklenk, K.; Doan, J.W. Regulation of oxidative phosphorylation, the mitochondrial membrane potential, and their role in human disease. J. Bioenerg. Biomembr. 2008, 40, 445–456. [Google Scholar] [CrossRef]
- Shen, X.L.; Zhang, Y.; Xu, W.; Liang, R.; Zheng, J.J.; Luo, Y.B.; Wang, Y.; Huang, K.L. An iTRAQ-based mitoproteomics approach for profiling the nephrotoxicity mechanisms of ochratoxin A in HEK293 cells. J. Proteom. 2013, 78, 398–415. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, N.; Luo, R.; Liu, J.; Guo, T.; Feng, J.; Peng, X. Transcriptomic and Proteomic Analysis Reveals Mechanisms of Patulin-Induced Cell Toxicity in Human Embryonic Kidney Cells. Toxins 2020, 12, 681. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110681
Han N, Luo R, Liu J, Guo T, Feng J, Peng X. Transcriptomic and Proteomic Analysis Reveals Mechanisms of Patulin-Induced Cell Toxicity in Human Embryonic Kidney Cells. Toxins. 2020; 12(11):681. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110681
Chicago/Turabian StyleHan, Nianfa, Ruilin Luo, Jiayu Liu, Tianmin Guo, Jiayu Feng, and Xiaoli Peng. 2020. "Transcriptomic and Proteomic Analysis Reveals Mechanisms of Patulin-Induced Cell Toxicity in Human Embryonic Kidney Cells" Toxins 12, no. 11: 681. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110681