Carbon Catabolite Repression Gene AoCreA Regulates Morphological Development and Ochratoxin A Biosynthesis Responding to Carbon Sources in Aspergillus ochraceus

 and
and 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Identification of AoCreA in A. ochraceus
2.2. Identification of AoCreA Gene Disruption in A. ochraceus
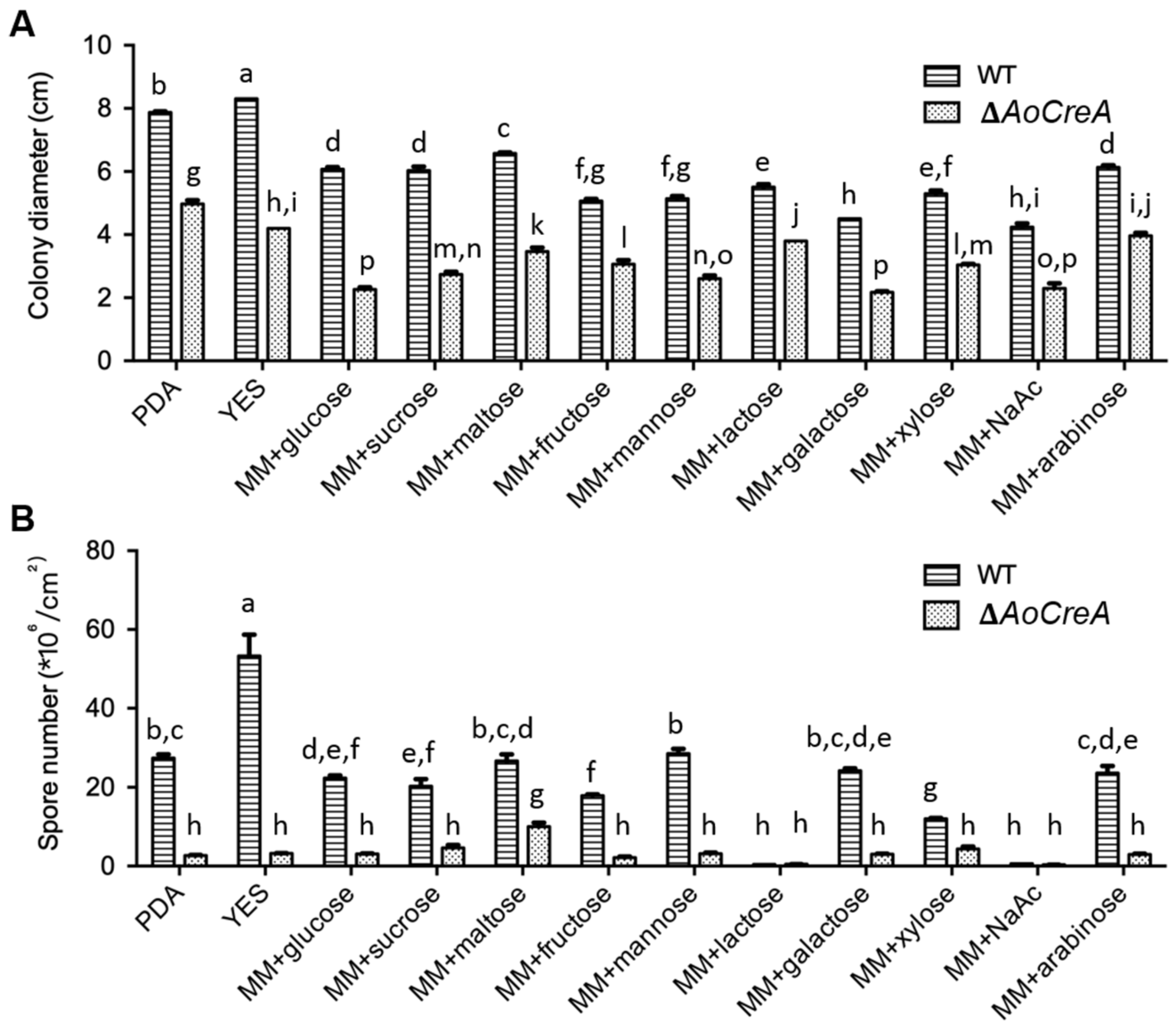
2.3. Growth and Conidiospore Production Were Affected by Carbon Sources and Modulated by AoCreA
2.4. AoCreA Was Involved in OTA Biosynthesis
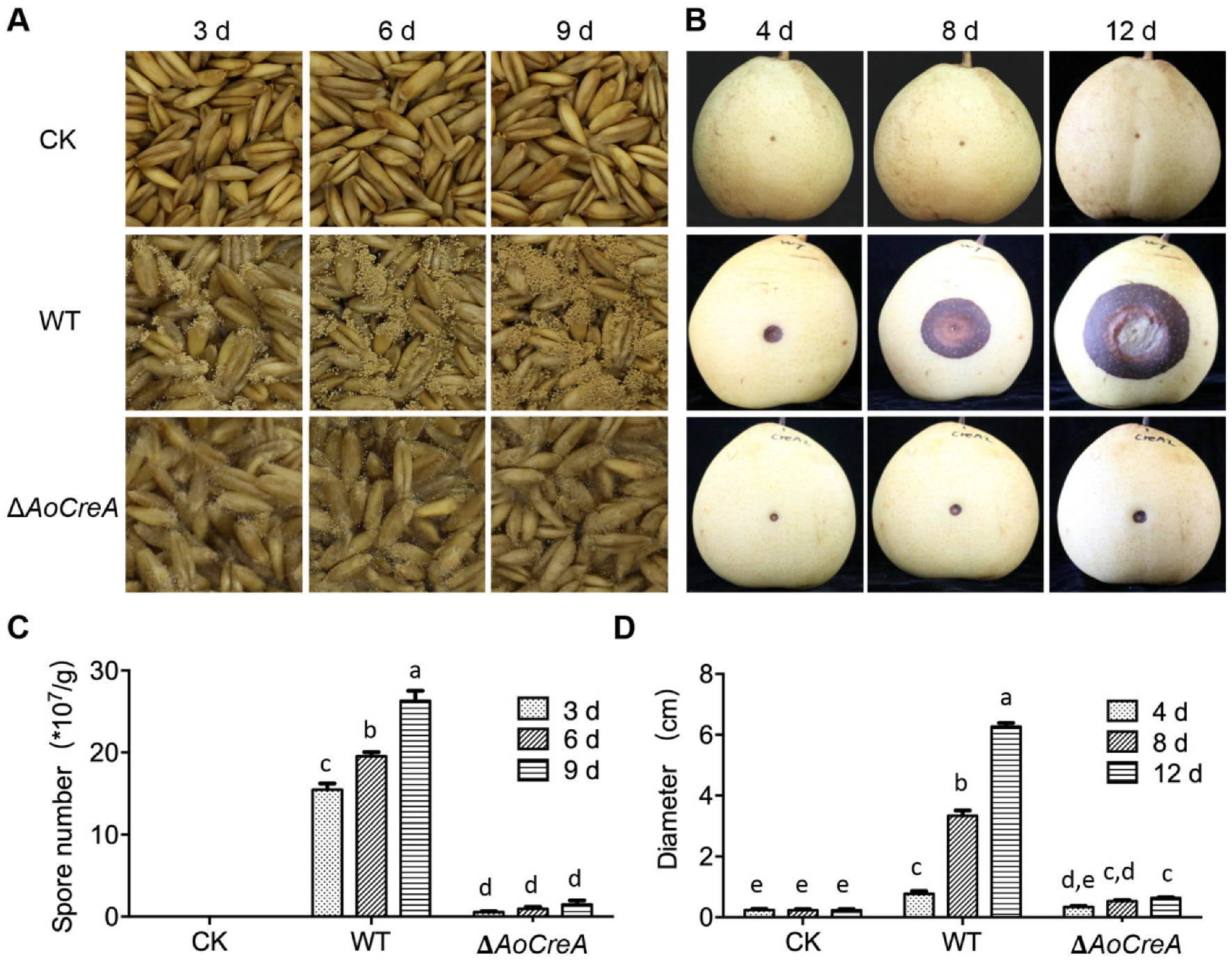
2.5. AoCreA in A. ochraceus Is Required for Fungal Infection
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Strains and Culture Conditions
5.2. AoCreA Identification and Phylogenetic Analysis
5.3. Construction of Gene Deletion Mutants
5.4. Phenotypic Studies
5.5. DNA and RNA Extraction
5.6. OTA Extraction and Production Analysis
5.7. Gene Expression Analysis
5.8. Pathogenicity Assay
5.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ruprich, J.; Ostry, V. Health risk assessment of the mycotoxin ochratoxin A to humans: Czech Republic--Brno--1991/92. Cent. Eur. J. Public Health 1993, 1, 86–93. [Google Scholar]
- Bayman, P.; Baker, J.L.; Doster, M.A.; Michailides, T.J.; Mahoney, N.E. Ochratoxin production by the Aspergillus ochraceus group and Aspergillus alliaceus. Appl. Environ. Microbiol. 2002, 68, 2326–2329. [Google Scholar] [CrossRef] [Green Version]
- Varga, J.; Kevei, E.; Rinyu, E.; Teren, J.; Kozakiewicz, Z. Ochratoxin production by Aspergillus species. Appl. Environ. Microbiol. 1996, 62, 4461–4464. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.P.; Mantle, P.G. Biosynthesis of ochratoxins by Aspergillus ochraceus. Phytochemistry 2001, 58, 709–716. [Google Scholar] [CrossRef]
- Van der Merwe, K.J.; Steyn, P.S.; Fourie, L. Mycotoxins. II. The constitution of ochratoxins A, B, and C, metabolites of Aspergillus ochraceus Wilh. J. Chem. Soc. 1965, 1, 7083–7088. [Google Scholar] [CrossRef]
- Wu, Q.; Dohnal, V.; Huang, L.; Kuca, K.; Wang, X.; Chen, G.; Yuan, Z. Metabolic pathways of ochratoxin A. Curr. Drug Metab. 2011, 12, 1–10. [Google Scholar] [CrossRef]
- IARC. Some naturally occurring substances: Food items and constituents, heterocyclic aromatic amines and mycotoxins. IARC Monogr. Eval. Carcinog. Risks Hum. 1993, 56, 245–395. [Google Scholar]
- Wang, G.; Zhang, H.; Wang, Y.; Liu, F.; Li, E.; Ma, J.; Yang, B.; Zhang, C.; Li, L.; Liu, Y. Requirement of LaeA, VeA, and VelB on asexual development, ochratoxin A biosynthesis, and fungal virulence in Aspergillus ochraceus. Front. Microbiol. 2019, 10, 2759. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, L.; Wu, F.; Liu, F.; Wang, Q.; Zhang, X.; Selvaraj, J.N.; Zhao, Y.; Xing, F.; Yin, W.B.; et al. A consensus ochratoxin A biosynthetic pathway: Insights from the genome sequence of Aspergillus ochraceus and a comparative genomic analysis. Appl. Environ. Microbiol. 2018, 84, e01009-18. [Google Scholar] [CrossRef] [Green Version]
- Crespo-Sempere, A.; Marin, S.; Sanchis, V.; Ramos, A.J. VeA and LaeA transcriptional factors regulate ochratoxin A biosynthesis in Aspergillus carbonarius. Int. J. Food Microbiol. 2013, 166, 479–486. [Google Scholar] [CrossRef]
- Gerin, D.; De Miccolis Angelini, R.M.; Pollastro, S.; Faretra, F. RNA-Seq reveals OTA-related gene transcriptional changes in Aspergillus carbonarius. PLoS ONE 2016, 11, e0147089. [Google Scholar]
- Castellá, G.; Bragulat, M.R.; Cigliano, R.A.; Cabañes, F.J. Transcriptome analysis of non-ochratoxigenic Aspergillus carbonarius strains and interactions between some black aspergilli species. Int. J. Food Microbiol. 2020, 317, 108498. [Google Scholar] [CrossRef]
- Gallo, A.; Ferrara, M.; Perrone, G. Recent advances on the molecular aspects of ochratoxin A biosynthesis. Curr. Opin. Food Sci. 2017, 17, 49–56. [Google Scholar]
- Luchese, R.H.; Harrigan, W.F. Biosynthesis of aflatoxin-the role of nutritional factors. J. Appl. Bacteriol. 1993, 74, 5–14. [Google Scholar]
- Szilágyi, M.; Miskei, M.; Karanyi, Z.; Lenkey, B.; Pocsi, I.; Emri, T. Transcriptome changes initiated by carbon starvation in Aspergillus nidulans. Microbiology 2013, 159, 176–190. [Google Scholar]
- Bailey, C.; Arst, H.N., Jr. Carbon catabolite repression in Aspergillos nidulans. Eur. J. Biochem. 1975, 51, 573–577. [Google Scholar]
- Dowzer, C.E.; Kelly, J.M. Analysis of the creA gene, a regulator of carbon catabolite repression in Aspergillus nidulans. Mol. Cell Biol. 1991, 11, 5701–5709. [Google Scholar]
- Hynes, M.J.; Kelly, J.M. Pleiotropic mutants of Aspergillus nidulans altered in carbon metabolism. Mol. Gen. Genet. 1977, 150, 193–204. [Google Scholar]
- Ruijter, G.J.; Visser, J. Carbon repression in Aspergilli. FEMS Microbiol. Lett. 1997, 151, 103–114. [Google Scholar]
- Kelly, J.M. The regulation of carbon metabolism in filamentous fungi. In Biochemistry and Molecular Biology, the Mycota; Brambl, R., Marzluf, G., Eds.; Springer: Berlin, Germany, 2016; Volume 3, pp. 385–401. [Google Scholar]
- Fasoyin, O.E.; Wang, B.; Qiu, M.; Han, X.; Chung, K.R.; Wang, S. Carbon catabolite repression gene creA regulates morphology, aflatoxin biosynthesis and virulence in Aspergillus flavus. Fungal Genet. Biol. 2018, 115, 41–51. [Google Scholar]
- Ichinose, S.; Tanaka, M.; Shintani, T.; Gomi, K. Improved α-amylase production by Aspergillus oryzae after a double deletion of genes involved in carbon catabolite repression. Appl. Microbiol. Biot. 2014, 98, 335–343. [Google Scholar] [CrossRef]
- Jin, C.; Barrientos, A.; Epstein, C.B.; Butow, R.A.; Tzagoloff, A. SIT4 regulation of Mig1p-mediated catabolite repression in Saccharomyces cerevisiae. FEBS Lett. 2007, 581, 5658–5663. [Google Scholar] [CrossRef] [Green Version]
- Zaragoza, O.; Rodriguez, C.; Gancedo, C. Isolation of the MIG1 gene from Candida albicans and effects of its disruption on catabolite repression. J. Bacteriol. 2000, 182, 320–326. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Li, T.; Gong, L.; Wang, Y.; Jiang, Y. Effects of different carbon sources on fumonisin production and FUM gene expression by Fusarium proliferatum. Toxins 2019, 11, 289. [Google Scholar] [CrossRef] [Green Version]
- Lillehoj, E.B.; Hesseltine, C.W. Aflatoxin control during plant growth and harvest of corn. In Mycotoxins in Human and Animal Health; Rodricks, J.V., Hesseltine, C.W., Eds.; Pathotox: Park Forest South, IL, USA, 1977; pp. 107–119. [Google Scholar]
- Brown, N.A.; Ries, L.N.A.; Goldman, G.H. How nutritional status signalling coordinates metabolism and lignocellulolytic enzyme secretion. Fungal Genet. Biol. 2014, 72, 48–63. [Google Scholar] [CrossRef]
- Cziferszky, A.; Mach, R.L.; Kubicek, C.P. Phosphorylation positively regulates DNA binding of the carbon catabolite repressor Cre1 of Hypocrea jecorina (Trichoderma reesei). J. Biol. Chem. 2002, 277, 14688–14694. [Google Scholar] [CrossRef] [Green Version]
- Kubicek, C.P.; Mikus, M.; Schuster, A.; Schmoll, M.; Seiboth, B. Metabolic engineering strategies for the improvement of cellulase production by Hypocrea jecorina. Biotechnol. Biofuels 2009, 2, 19. [Google Scholar] [CrossRef] [Green Version]
- Nahas, E.; Waldemarin, M.M. Control of amylase production and growth characteristics of Aspergillus ochraceus. Rev. Latinoam. Microbiol. 2002, 44, 5–10. [Google Scholar]
- Shimizu, K.; Keller, N.P. Genetic involvement of a cAMP-dependent protein kinase in a G protein signaling pathway regulating morphological and chemical transitions in Aspergillus nidulans. Genetics 2001, 157, 591–600. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA 5: Molecular evolutionary genetics analysis using maximumlikelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Liu, Z.; Lin, R.; Li, E.; Mao, Z.; Ling, J.; Yang, Y.; Yin, W.; Xie, B. Biosynthesis of antibiotic leucinostatins and their inhibition on Phytophthora on bio-control Purpureocillium lilacinum. PLoS Pathog. 2016, 12, e1005685. [Google Scholar] [CrossRef] [Green Version]
- Solomon, P.S.; Ipcho, S.V.S.; Hane, J.K.; Tan, K.C.; Oliver, R.P. A quantitative PCR approach to determine gene copy number. Fungal Genet. Rep. 2008, 55, 5–8. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.; Wang, Y.; Yang, B.; Zhang, C.; Zhang, H.; Xing, F.; Liu, Y. Carbon Catabolite Repression Gene AoCreA Regulates Morphological Development and Ochratoxin A Biosynthesis Responding to Carbon Sources in Aspergillus ochraceus. Toxins 2020, 12, 697. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110697
Wang G, Wang Y, Yang B, Zhang C, Zhang H, Xing F, Liu Y. Carbon Catabolite Repression Gene AoCreA Regulates Morphological Development and Ochratoxin A Biosynthesis Responding to Carbon Sources in Aspergillus ochraceus. Toxins. 2020; 12(11):697. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110697
Chicago/Turabian StyleWang, Gang, Yulong Wang, Bolei Yang, Chenxi Zhang, Haiyong Zhang, Fuguo Xing, and Yang Liu. 2020. "Carbon Catabolite Repression Gene AoCreA Regulates Morphological Development and Ochratoxin A Biosynthesis Responding to Carbon Sources in Aspergillus ochraceus" Toxins 12, no. 11: 697. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110697