1. Introduction
Fescue toxicosis, resulting from consumption of ergot alkaloids commonly found in endophyte (
Epichloë coenophiala)-infected tall fescue (
Lolium arundinaceum Schreb. Darbysh), significantly impacts livestock health and production globally. Direct exposure to ergot alkaloids from tall fescue and other grass species occurs in production systems that heavily rely on grazing, such as in cow calf and stocker programs in the United States, New Zealand, and Australia; whereas importation of feedstuff has led to ergot alkaloid-induced effects in several Asian countries, including Japan and Korea [
1,
2,
3]. Due to the agronomic benefits resulting from symbiosis with the endophyte, including stand persistence and drought tolerance, it is expected that endophyte-infected tall fescue will become the primary forage across more growing zones as the global climate warms. McCulley et al. [
4] recently reported that warming air temperatures (+3 °C, day and night, year-round) significantly increased concentrations of ergovaline and total ergot alkaloids found in endophyte-infected tall fescue, which may exacerbate fescue toxicosis in future years. Complex plant–fungus–environment interactions exist due to variation in endophyte infection rate, alkaloid production (125 to 5000 µg/Kg), and the animal’s voluntary intake [
4], all of which can lead to high variation in individual animal responses. Individual variation in response to fescue toxicosis might be, in part, due to host genetics. Galliou et al. [
5] tested the association between genotypes from a commercial genetic test for fescue toxicosis and performance in pregnant Angus cows. These authors found significant associations with growth, hair shedding, and calf weaning weight. Altogether, these pose a major challenge to understanding the underlying mechanisms of action responsible for the decreases in livestock productivity. While advancements have been made to better understand the mechanisms associated with fescue toxicosis symptoms, such as focusing on improving growth and reproductive performance of beef cattle grazing endophyte-infected tall fescue, a greater understanding is needed to identify markers for tolerance to fescue toxicosis, which is critical for the sustainability of the global beef industry.
Tall fescue, and more specifically the cultivar Kentucky 31, is the most abundant and economically important cool-season perennial grass in U.S. agriculture. Approximately 8.5 million head of beef cattle reside in southeast or central U.S., where fescue is the predominant forage available, most of which contains the endophyte that causes toxicosis in grazing livestock. While precise data on the rate and composition of ergot alkaloid intake in grazing cattle is limited, the symptoms of this multifaceted disease range from acute outbreaks of gangrenous ergotism to more subtle and chronic decreases in livestock productivity characterized by decreased feed intake and growth performance, compromised reproduction (pregnancy and calving rates), elevated body temperature, and reduced blood flow to the extremities, as reviewed by Strickland et al. [
1]. While the gangrenous ergotism is rare and most likely due to acute exposure to very high concentrations of ergot alkaloids, combined with cold temperatures, the decreased livestock health, welfare, and productivity is expected as a result of chronic exposure to ergot alkaloids.
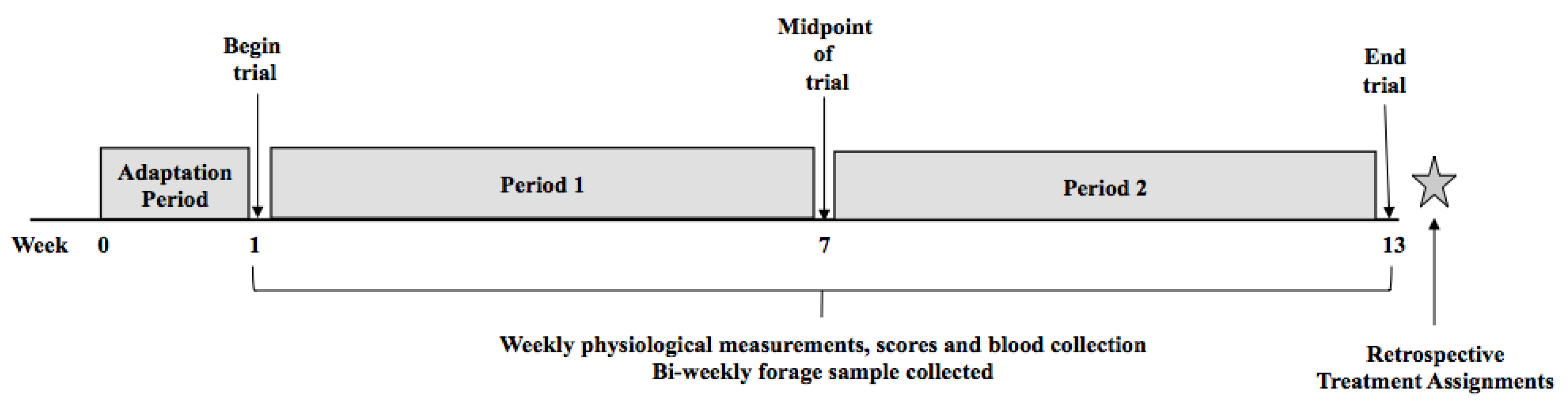
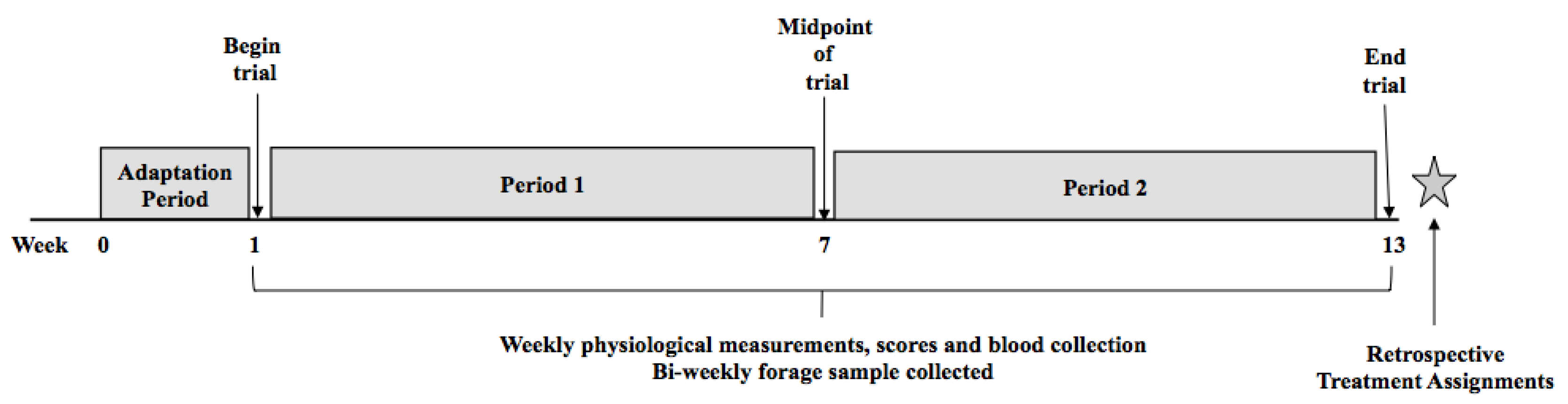
While there have been extensive efforts to develop an innovative and strategic pasture and grazing management system to convert toxic tall fescue to the non-toxic “Novel Endophyte Tall Fescue”, many farmers have exercised some caution and hesitation to adopt the technology. This appears to be due to lack of understanding of how to renovate, expense, time, challenging terrain, and lack of feed resources to utilize while the new forage is being established. Essentially, the goal of converting all of the toxic fescue to novel endophyte tall fescue is likely impractical and highly unlikely. Therefore, there has been concurrent interest in identifying ways that producers can select animals based on their tolerance to the adverse effects of fescue toxicosis. As previously stated, animals suffering from fescue toxicosis exhibit decreased growth capability, poor reproductive performance, and have a lack of adequate heat tolerance at least in part due to their retained winter hair coat. Focusing on growth performance selection parameters and the animal’s cytokine response to the toxin could be utilized to identify animals that are tolerant to the effects of fescue toxicosis. Therefore, the goals of this study were to develop selection criteria to identify tolerant and susceptible animals based on phenotypic animal performance traits, to evaluate how our criteria matches with the information from a commercial test for fescue toxicosis, and then examine responsive cytokine profiles to identify beef cattle displaying tolerance to fescue toxicosis.
3. Discussion
Due to the extensive use of endophyte-infected tall fescue across the globe, the goals of this study were to develop selection criteria to identify tolerant and susceptible animals based on phenotypic animal performance traits, and then examine responsive cytokine profiles to identify beef cattle displaying tolerance to fescue toxicosis. Several studies described breed differences in response to ergot alkaloids, in which breeds such as Senepol and Brahman, which are better able to handle the heat and humidity, outperformed British breeds such as Angus and Hereford [
8,
10,
11,
12] when exposed to toxic tall fescue. These breeds have been used in crossbreeding programs to address fescue toxicosis, but it is unclear if their apparent tolerance is to fescue or adaptation to elevated heat and humidity. However, genetic progress is obtained within the breed, and the identification of genetic variation in animals from the same breed should be exploited to allow for selection for improved response (e.g., tolerance) to fescue toxicosis. Gray et al. [
13] examined Angus cattle performance in North Carolina and Mississippi on endophyte-infected fescue pasture. These researchers observed genetic variation for hair coat shedding, indicating that Angus cattle that shed their winter hair coat earlier in the year may be more heat tolerant.
At the genomic level, genes regulating prolactin production have been targeted to identify genetic markers for tolerance to fescue toxicosis. Campbell et al. [
14] recently identified a single nucleotide polymorphism (SNP) within the dopamine receptor D2 gene that was associated with variation in calving rates when grazing endophyte-infected tall fescue. In addition, using part of the same data in this study, Galliou et al. [
5] showed that a commercial genetic test for fescue toxicosis, T-Snip (AgBotanica, LCC, Columbia, MO, USA), is associated with growth, hair shedding, and calf weaning weight in pregnant Angus cows. With the available T-Snip data, we tested whether the distribution of T-Snip genotypes changed between EI-TOL and EI-SUS animals in our study. According to the company’s instructions, T-Snip genotypes range from 1 to 5, with levels of tolerance to fescue toxicosis increasing with the value of the genotype. Although both groups of animals showed T-Snip genotypes 1 to 4, there was a greater proportion of genotype 3 in EI-TOL than in EI-SUS. Similarly, there was a greater proportion of genotype 2 in EI-SUS than in EI-TOL. Therefore, the expected genetic values based on this commercial genetic test was greater in EI-TOL than in EI-SUS. Hence, although the selection of animals in this study was fully based on their phenotypic performance, results from T-Snip genotypes further support that this selection method was able to identify animals with contrasting tolerance to fescue toxicosis. It is important to note that this commercial genetic test does not have a perfect accuracy to predict with phenotypic performance in animals during fescue toxicosis [
5]. Therefore, the presence of animals with lower and greater genotype values in EI-TOL and EI-SUS groups, respectively, was expected. Considering these factors, developing a selection method based on the phenotypic performance parameter in cattle from a single breed source in controlled experiments would aid in identifying animals that are potentially tolerant or susceptible to ergot alkaloids.
A major challenge with studying this multifaceted syndrome is that endophyte production of ergot alkaloids varies with season and plant maturity, and contributes to variation in ergot alkaloid intake by grazing cattle. Total ergot alkaloids or ergovaline, the most abundant alkaloid, has been measured in fescue to describe its potential toxicity. Concentrations of ergovaline increase from 250 to 500 µg/Kg in leaf blades and from 500 to 1300 µg/Kg in leaf sheaths from April to May. Seed heads contain the greatest concentration of toxins, and reach concentrations as high as 5000 µg/Kg in June. Ergovaline concentrations decline in August and increase again during fall regrowth [
9]. Total ergot alkaloid concentration displays the same seasonal changes as ergovaline [
15]. The concentration of total ergot alkaloids and ergovaline declines by 81% to 85%, respectively, from December to March in stockpiled fescue (accumulated during the growing season for grazing during dormancy; [
16,
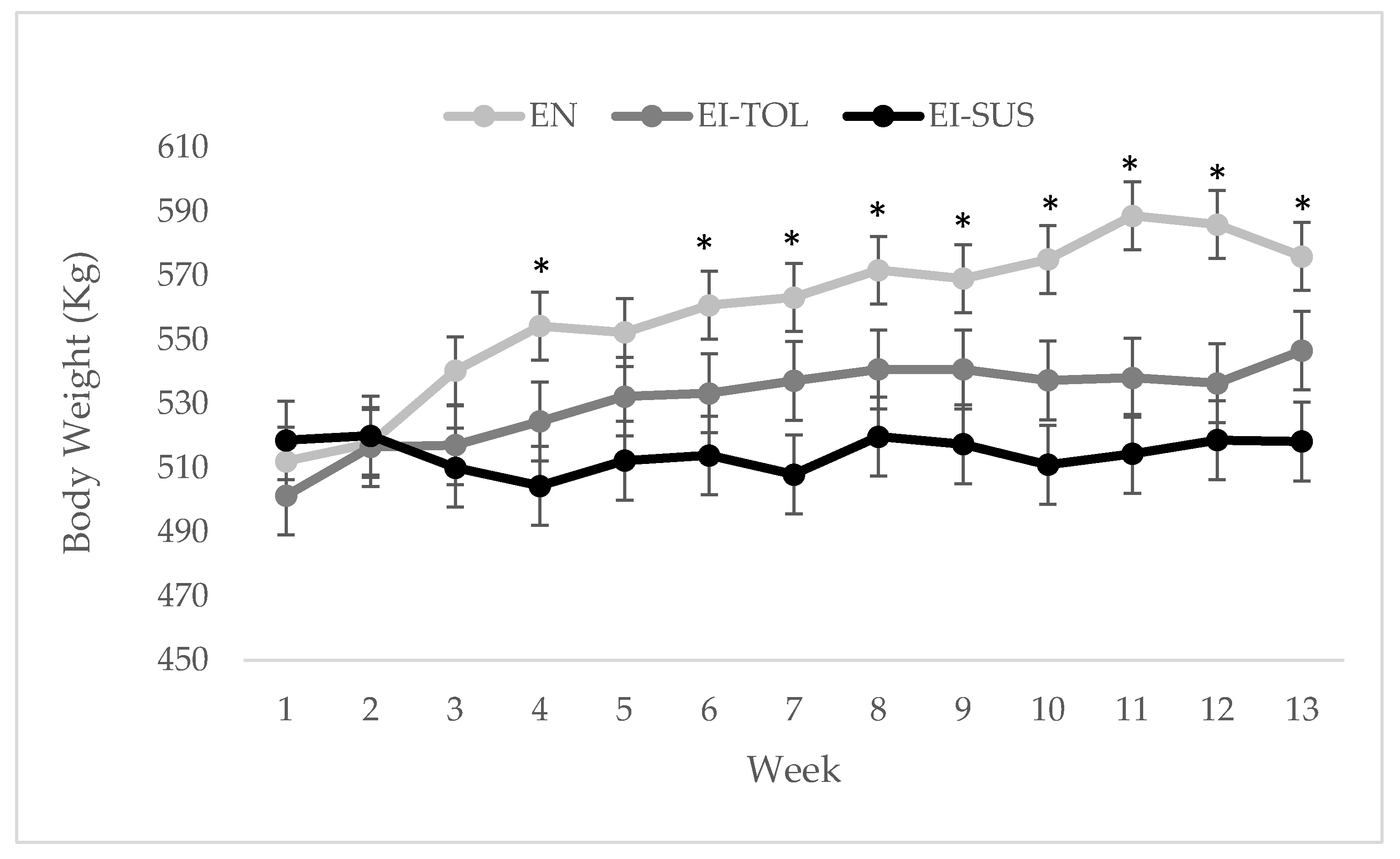
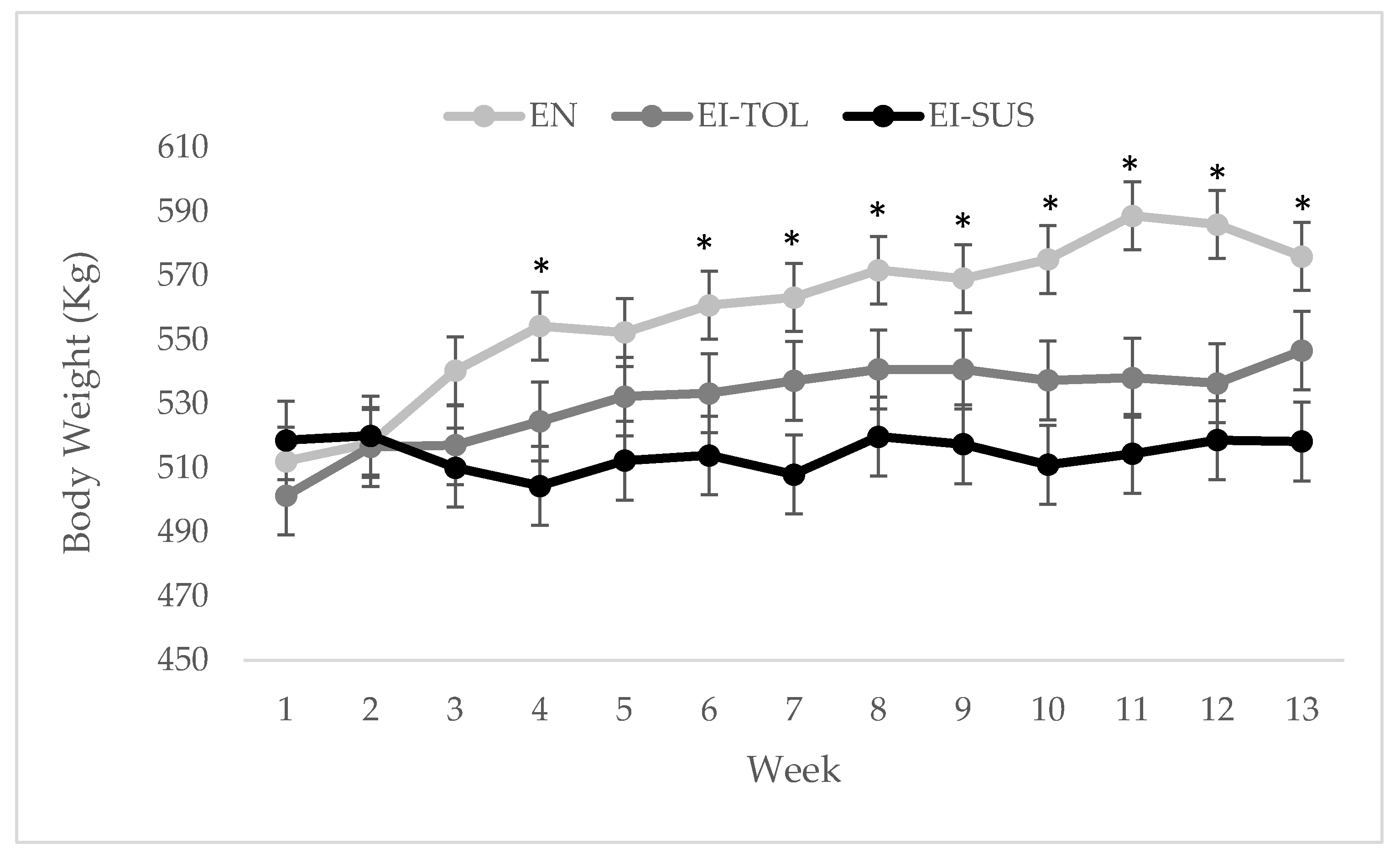
17]). There were differences in the percentage of fescue and endophyte infection rate between locations used in this study, which then led to differences in ergovaline concentrations between locations, as well as variation in ergovaline concentration throughout the trial. The increase in ergot alkaloid consumption during period 1 resulted in greater differences in the phenotypic performance parameter, including average daily gain (ADG), body condition scores (BCS), body weight (BW), and rectal temperatures that were evaluated in this study.
The difference in ADG and BCS in response to increased consumption of ergot alkaloids is similar to data reported Thompson et al. [
18], which identified a direct negative correlation to an animal’s ADG when the pasture infection rate increased. Moreover, comparison of phenotypic performance parameters of animals exposed to ergot alkaloids (EI) to animals not exposed to ergot alkaloids (EN) demonstrated the severity of exposure throughout the trial. Additionally, sufficient body condition has been shown to be one of the most important traits to cattle reproductive efficiency in a cow–calf herd, and decreases as a result of endophyte-infected tall fescue consumption [
19].
Aside from animal growth performance, issues with thermotolerance have also frequently been seen in animals suffering from severe cases of fescue toxicosis [
20,
21]. Many of the symptoms of fescue toxicosis (ergot alkaloid exposure) are amplified during a period of heat stress, and the greatest loss in animal performance and consequently increased production losses occur during the summer months when grazing endophyte infected tall fescue [
22]. It has been speculated that heat tolerant (i.e., Bos indicus) breeds of cattle have improved resistance to fescue toxicosis [
10,
23]. Several studies, including Poole et al. [
8], have demonstrated that cattle with adaptions to manage heat stress perform better when exposed to ergot alkaloids and increased environmental temperatures. However, there is conflicting data that report no difference in growth rate [
24], hormone concentration [
23], or milk production [
11] between Bos indicus and Bos taurus cattle grazing endophyte-infected tall fescue. Therefore, it remains unknown if the traits for heat tolerance and tolerance to ergot alkaloids are synonymous.
Correlations have been previously made in which cattle that shed their winter hair coats earlier in the fescue season suffer less from the negative effects of fescue toxicosis [
13]. In this study, weekly hair shedding scores (HSS) and hair coat scores (HCS) were taken to evaluate differences based on tolerance designation week to week, as well as initial rate of shedding. Both measures were significantly different between tolerant and susceptible animals grazing the endophyte-infected tall fescue. Both HCS and HSS were similar at the start of the study, and deviation between EI-TOL and EI-SUS animal occurred at week three of the study and continued through week seven (P-1). Interestingly, HSS and HCS re-converged to similar numeric values when ergot alkaloid concentrations decreased, yet ambient temperatures and humidity remained elevated consequentially when prolactin concentrations increased. Decreases in serum prolactin have been associated with fescue toxicity, and are often used as an indicator of ergot alkaloid exposure in cattle [
25]. However, increased prolactin concentrations are associated with increasing day lengths and ambient temperatures, and initiate hair shedding [
26,
27]. Prolactin secretion has been linked with changes in environmental temperature, such that prolactin concentrations are elevated during warmer verses cooler months [
28,
29], and has been shown to regulate hair shedding [
30,
31]. It has been speculated that fescue toxicosis-induced hypoprolactinemia prevents shedding of the winter hair coat, therefore, cattle have an elevated body temperature and increased vulnerability to heat stress [
32]. In the current study, EI-TOL animals had a tendency for lower serum prolactin concentrations when compared to EI-SUS animals as determined by the FTSM, thus, it can be assumed that those animals deemed tolerant had sufficient concentrations of serum prolactin to initiate hair shedding. Based on the other physiological measurements, the cattle in this study displayed many other symptoms of fescue toxicosis, and the lack of hypoprolactinemia was unexpected. The interaction of increased THI and exposure to ergot alkaloids impacts the severity of fescue toxicosis (reviewed by one), and the relatively low inclusion rate of the ergot alkaloids in the infected pastures may have altered serum prolactin concentrations. With EI-TOL animals having lower average serum prolactin concentrations than EI-SUS animals, it is hypothesized that these cattle were genetically predisposed to eliciting a shedding response at lower thresholds of prolactin. Using the same animals from this study, Koester et al. [
33] showed that EI-TOL and EI-SUS animals have distinct fecal bacterial and fungal communities, further supporting that the proposed FTSM also results in differences in the microbiome of animals under FT stress. This classification using FTSM is further supported by the differences in T-Snip genotypes between EI-TOL and EI-SUS presented in the current study. These hypotheses build from the speculation of Aiken et al. [
32] that genetic predispositions exist for necessary prolactin levels to initiate shedding.
More recent studies in various livestock species have reported that immune parameters have moderate to high heritability, as well as high genetic correlation with reproductive performance (lowly heritable traits), indicating that immune parameters can be used as a genetic trait for selecting animals that are resistant to specific diseases [
34]. Using the protein array approach, numerous growth factors, chemokines, cytokine receptor antagonists, and anti-inflammatory and pro-inflammatory cytokines were examined during ergot alkaloid exposure. One of the major outcomes of this analysis is that many of the factors examined were significantly different or showed a statistical tendency to be higher at UPRS compared to BBCFL (
Table 5 and
Table 6). While cytokine differences between locations was not the primary focus of this study, these data demonstrate the immune system’s responsiveness to varying ergot alkaloid exposure, and warrants further investigation with known concentrations of ergovaline to better understand this interaction. Based on these data, it is hypothesized that cattle at the UPRS location experienced a greater immune response in response to higher ergot alkaloid exposure. A similar report described a hyperactive innate immune response, which may lead to an immuno-compromised animal in stocker steers when chronically exposed to ergovaline [
35].
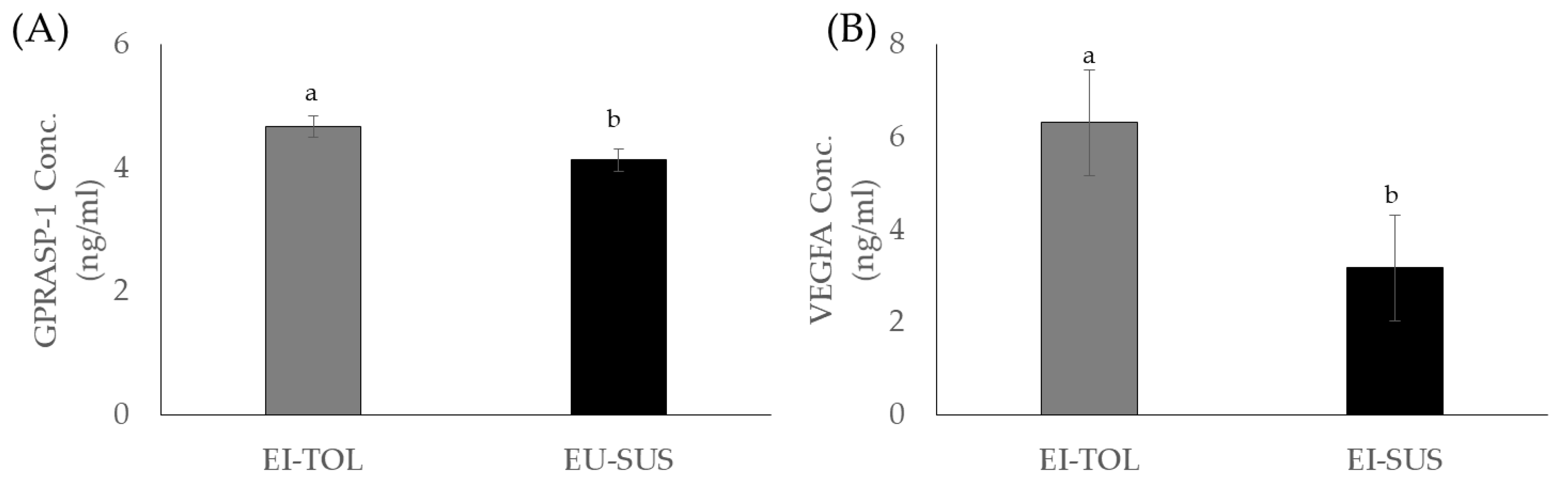
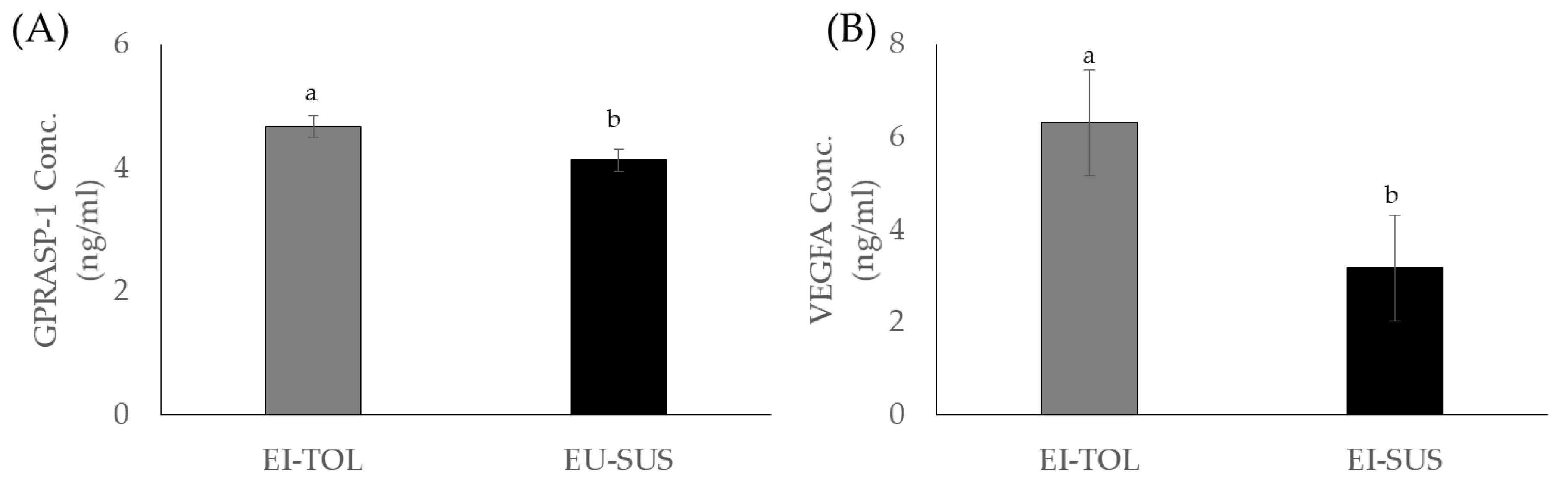
In the current study, vascular endothelial growth factor A (VEGFA) and G Protein-Coupled Receptor Associated Sorting Protein 1 (GPRASP-1) concentrations were greater in EI-TOL animals compared to EI-SUS animals. G Protein-Coupled Receptor Associated Sorting Protein 1 has been associated with downregulation of a variety of G Protein-Coupled receptors, including the D2-dopamine receptor, through lysosomal degradation. The D2-dopamine receptor has been shown to play a direct effect in cattle prolactin secretion, and is also involved in ergot alkaloids exposure’s decrease in prolactin secretion [
36,
37]. Prolactin is decreased when dopamine binds to the D2-dopamine receptor, and the cyclic ring structure of ergovaline closely mimics the ring structure of dopamine, allowing ergovaline to bind the D2-dopamine receptor and thus inhibit prolactin secretion [
38,
39]. In the current study, EI-TOL had greater GPRASP-1 concentrations, potentially causing the D2-dopamine receptor to be downregulated at a greater rate. This decrease in the prevalence of the D2-dopamine receptor decreased the opportunity for ergovaline to bind and decrease prolactin secretion, giving EI-TOL animals an advantage when grazing endophyte-infected tall fescue. Furthermore, this could contribute to EI-TOL animals having greater sensitivity to prolactin through downregulation of the entire prolactin secretion pathway, resulting in lower biological thresholds needed to initiate the biological roles of the hormone, which was previously discussed.
Vascular endothelial growth factor A is a known vasodilator, and has been shown to increase micro-vascular permeability. In addition, it is known to play a particularly important role in vascular endothelial cells, where higher concentrations increase vascular permeability [
40] and could provide protective effects against ergot alkaloid exposure. Vasoconstriction caused by exposure to ergot alkaloids further compounds the issues associated with fescue toxicosis, such as its ability to decrease the animal’s capacity for evaporative cooling [
41]. Increased VEGFA concentrations and lower rectal temperature observed in the EI-TOL animals provide evidence that the EI-TOL animals selected using the FTSM have a greater ability to avoid heat stress and the vasoconstrictions effects of fescue toxicosis compared to EI-SUS animals. Several studies have examined changes in VEGFA to heat stress, and have focused on tissue and/or cellular responsiveness as opposed to whole animal changes. Vascular endothelial growth factor concentrations in the blood of heat tolerant (Bos Indicus) cattle compared to cattle that are more sensitive to heat stress (Bos Taurus) has not been directly investigated; however, Jyotiranjan et al. [
42] and Iqbal et al. [
43] reported increases in VEGF gene expression in response to thermal stress in goats and Bos indicus cattle breeds, respectively. Interestingly, of the eight Bos indicus breeds examined, only two breeds (Bhagnari and Lohani) displayed increased VEGF expression, but the authors indicate that this increase in VEGF is associated with adaptation to high altitude as opposed to heat tolerances [
43]. Additionally, Jones et al. [
44] and Aiken et al. [
45] have speculated that decreases in serum progesterone concentrations could be caused by vasoconstriction of blood flow to the ovary in cattle consuming ergot alkaloids. Poole and colleagues [
7] confirmed that ergovaline exposure reduced the diameter of the ovarian artery, leading to the functional corpus luteum, thus decreasing circulating progesterone concentrations. In the current study, EI-TOL animals tended to have higher progesterone concentrations compared to EI-SUS animals, which may be linked to greater ovarian artery diameter due to greater concentrations of circulating VEGFA in these animals. Thus, the EI-TOL animal’s ability to synthesize higher concentrations of VEGFA may provide the capacity to increase vessel diameter, which would result in substantial mitigation of the vasoconstriction seen with fescue toxicosis. Ultimately, further investigation is needed to evaluate these cytokines and their role in fescue toxicosis, as well as the tolerant animal’s ability to mount a stronger immune response to mitigate some of the negative effects of fescue toxicosis.
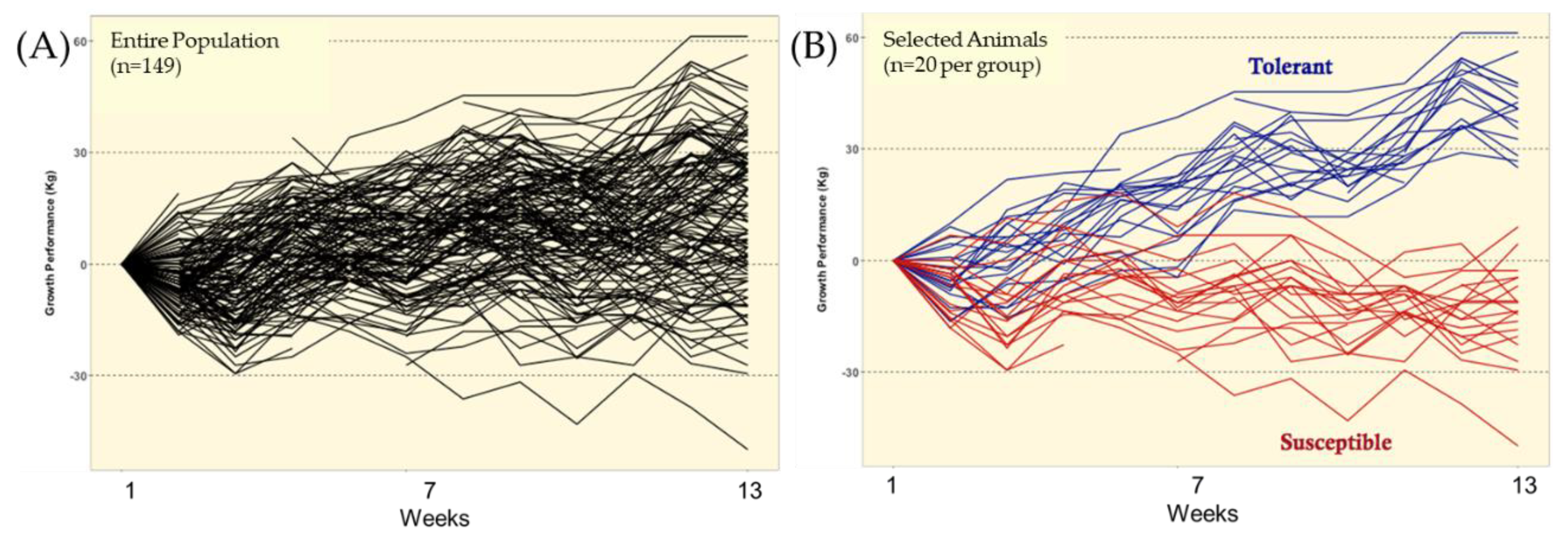
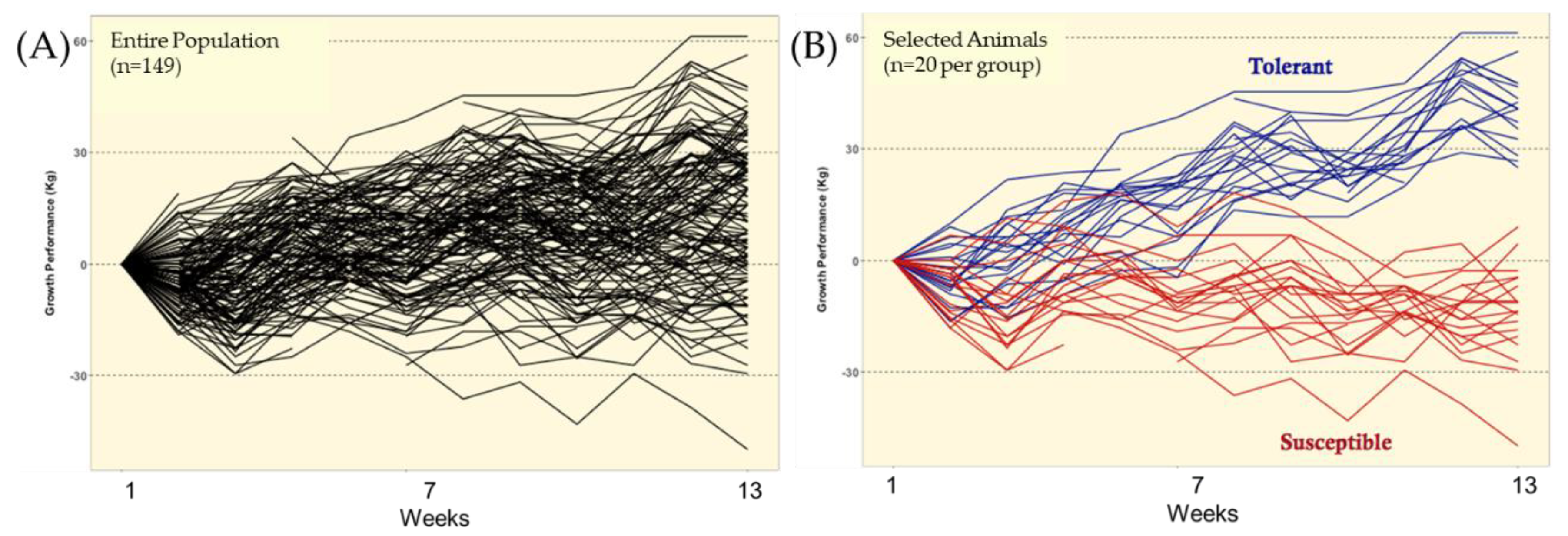
Taken together, the selection strategy used in this study (i.e., FTSM) was showed to be effective in identifying groups of individuals expressing contrasting responses to ergot alkaloid exposure, i.e., fescue toxicosis [
33]. Nonetheless, three items must be further discussed: first, the selection of these animals was based solely on their (adjusted) phenotypic performance. Although the T-Snip data presented in this study support that TOL animals have greater genetic tolerance to ergot alkaloid exposure than SUS animals, the accuracy of identifying individual differences from ergot alkaloid exposure using this test was not 100% [
5]. Hence, there is still genetic variation in the animal’s response to ergot alkaloid exposure that could be explored. One way of getting ahold of individual genetic variation is through using, for example, the expected progeny differences (EPDs) for the growth rate of these animals as part of the selection criteria. However, there are some limitations on using the current EPDs for this purpose. These EPDs are based on a nationwide genetic evaluation (American Angus Association;
http://www.angus.org/), which does not take into consideration the presence of GxE, which seems to be the case for ergot alkaloid exposure and for other stress-related traits [
46]. Thus, the use of EPDs for this purpose may bias the selection of animals with greater tolerance to ergot alkaloid exposure. Although no source of genetic information was used in our selection process, it was expected that our approach captured part of the genetic potential of these animals. Assuming a) unbiased estimates for the fixed effects and b) independence between fixed and random effects included in the statistical model used for selection of TOL and SUS animals (see details in Koester at al. [
33]), the estimated residuals that were used for classification of TOL and SUS animals should include both genotypic effects (additive and non-additive) and the true residual effects. Thus, assuming at least a moderate narrow-sense heritability for growth rate under ergot alkaloid exposure, it is expected that the estimated residuals include a substantial contribution of additive genetic values. Hence, although the selection process used in this study did not include prior genetic information from these animals, the data presented in this study demonstrate that TOL and SUS animals differed, at some level, in their genetic potential when exposed to ergot alkaloids.
Additionally, another consideration is the potential confounding between the growth rate in the presence or absence of ergot alkaloids. In this study, it was assumed that animals expressed their growth as a function of response to ergot alkaloid exposure. In fact, the difference in results obtained between the two locations with endophyte-infected (toxic) fescue suggests that some minimum concentrations of ergot alkaloids are needed for animals to express a tolerant-related phenotype. In order to obtain accurate information on the relationship between the growth rate in the presence or absence of ergot alkaloids at the genetic level, a much larger sample size, with animals from a substantial number of sires represented across locations with varying concentrations of ergot alkaloids, would be needed to estimate the genetic correlation between the growth rate in the presence or absence of ergot alkaloids. Albeit needed, such a scenario would be difficult to achieve in a timely manner. At the phenotypic level, the use of biomarkers for ergot alkaloid exposure, such as prolactin [
25] or those identified in our study (e.g., VEGFA), could provide additional information on the level of ergot alkaloid exposure on animals. This information could be used to evaluate whether a significant physiological response to ergot alkaloid exposure was observed in each animal, potentially allowing for better separation between animals showing a growth rate in the presence or absence of ergot alkaloids.
Lastly, our method was unidimensional, using only growth rate data for the selection of TOL and SUS animals. Hence, the evaluation of a fescue toxicosis selection index using multiple parameters and including other sources of information could be beneficial for more accurate selection of animals for response to ergot alkaloid exposure. Additional sources of information include the use of T-Snip, the biomarkers identified in this population (this study and Koester et al. [
33]), genotypes at the dopamine receptor D2 gene locus [
14], and more. Opportunities exist for the identification of additional biomarkers based on response-related traits other than growth rate, as used in this study. There is also a need for large-scale genomic studies for the identification of SNPs associated with variation in ergot alkaloid exposure related traits, which could be used to better identify animals with different genetic potential for fescue toxicosis.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}