Isolation and Characterization of a Deoxynivalenol-Degrading Bacterium Bacillus licheniformis YB9 with the Capability of Modulating Intestinal Microbial Flora of Mice

Abstract
:1. Introduction
2. Results
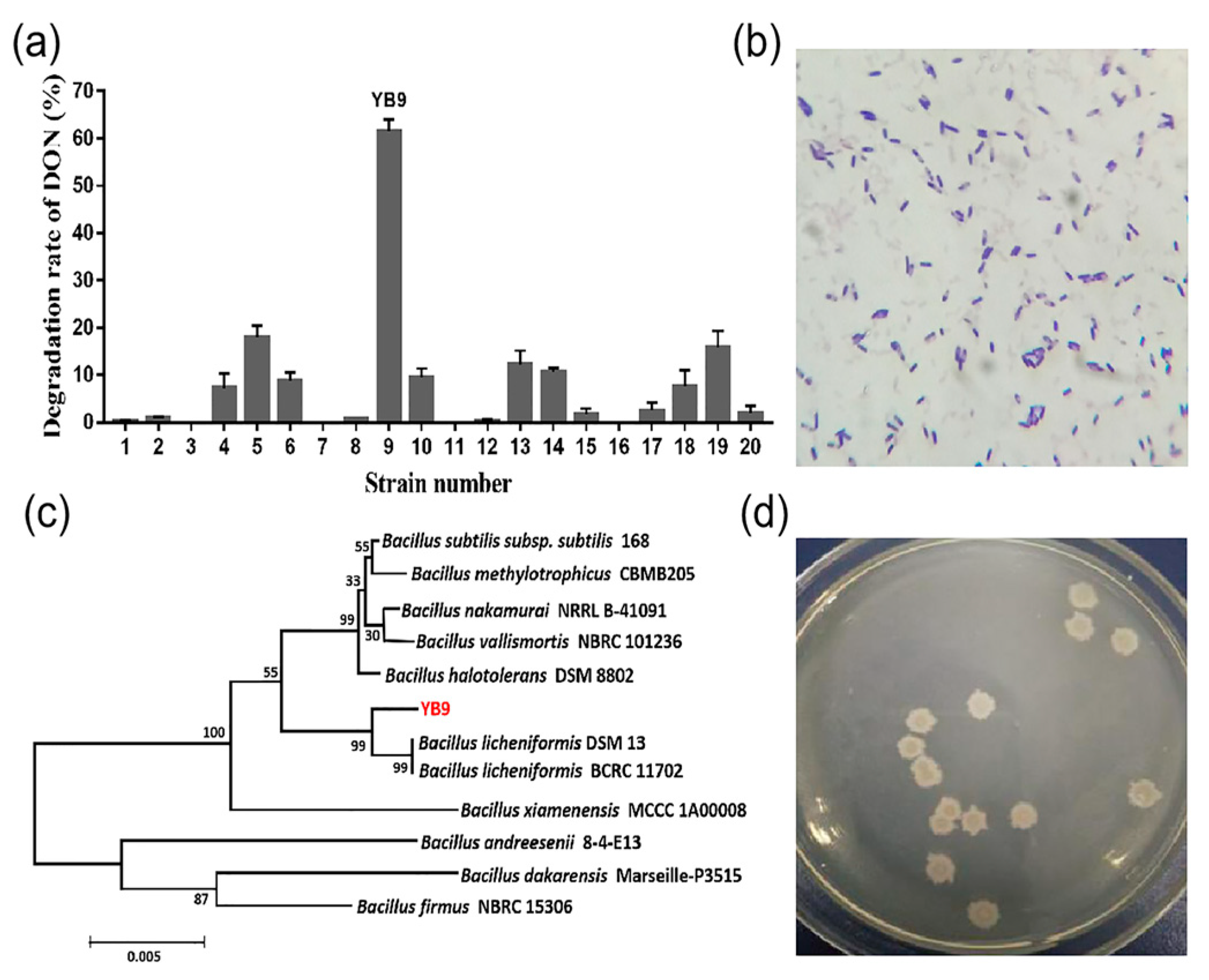
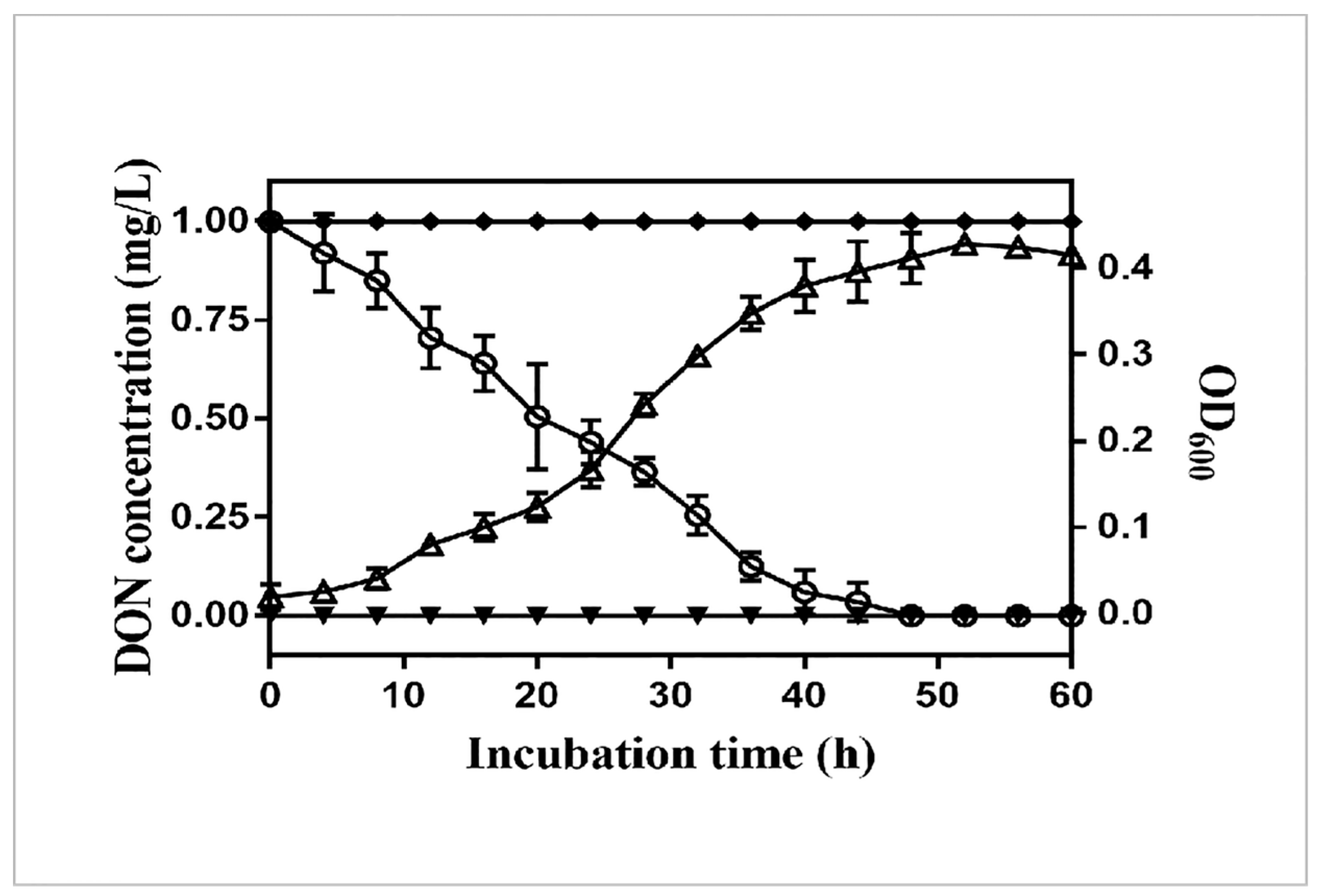
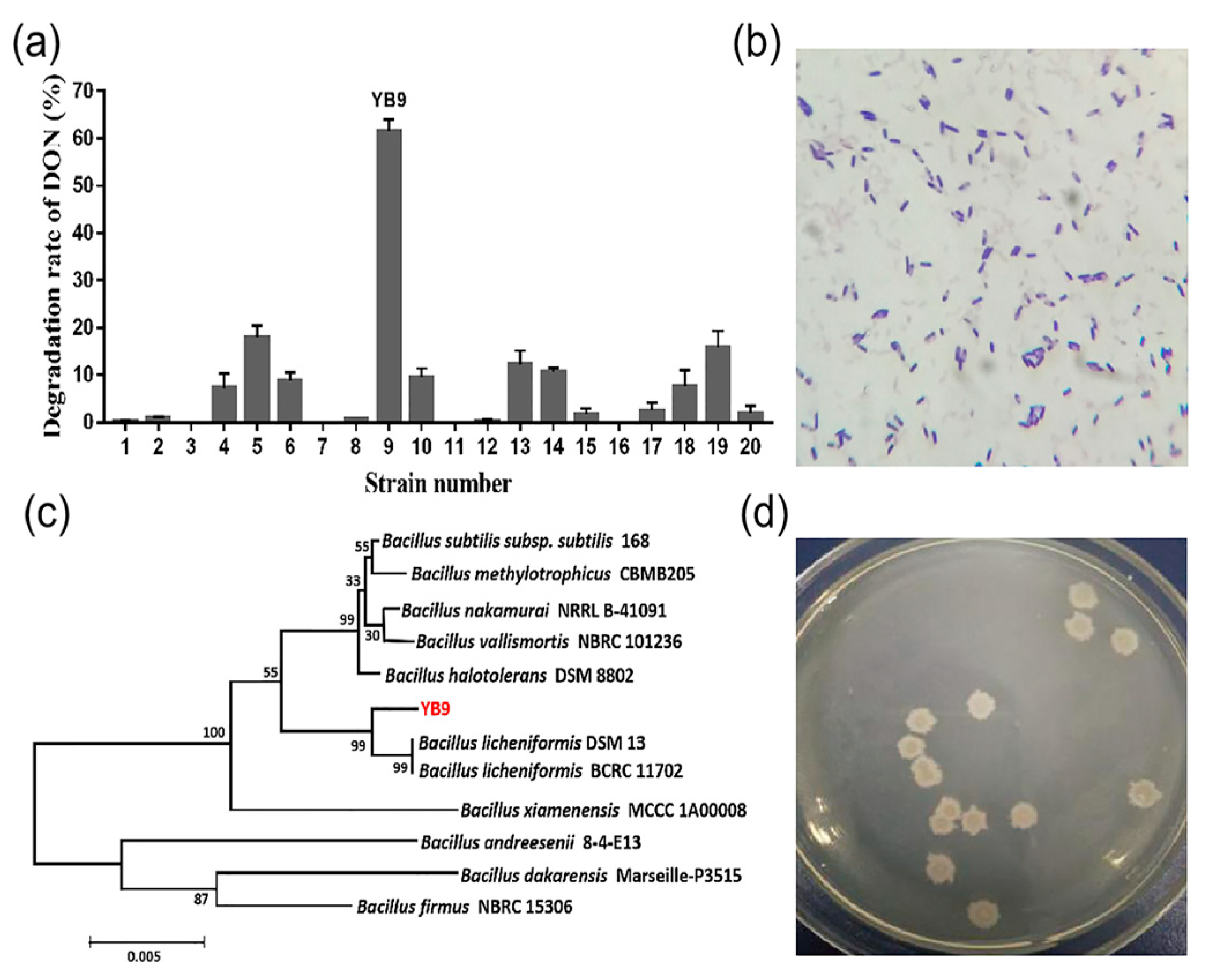
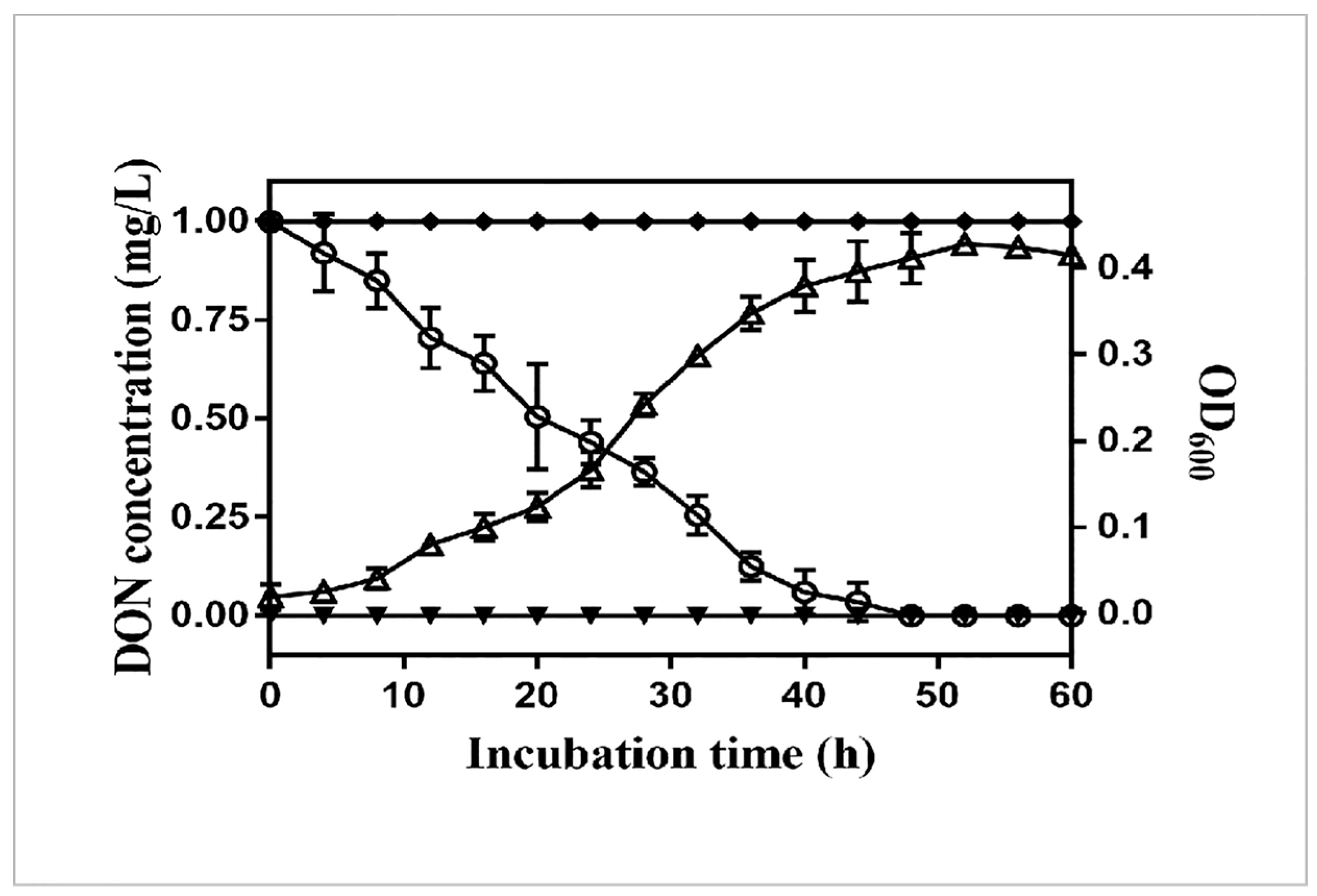
2.1. Isolation and Identification of the DON-Degrading Strain Bacillus Licheniformis YB9
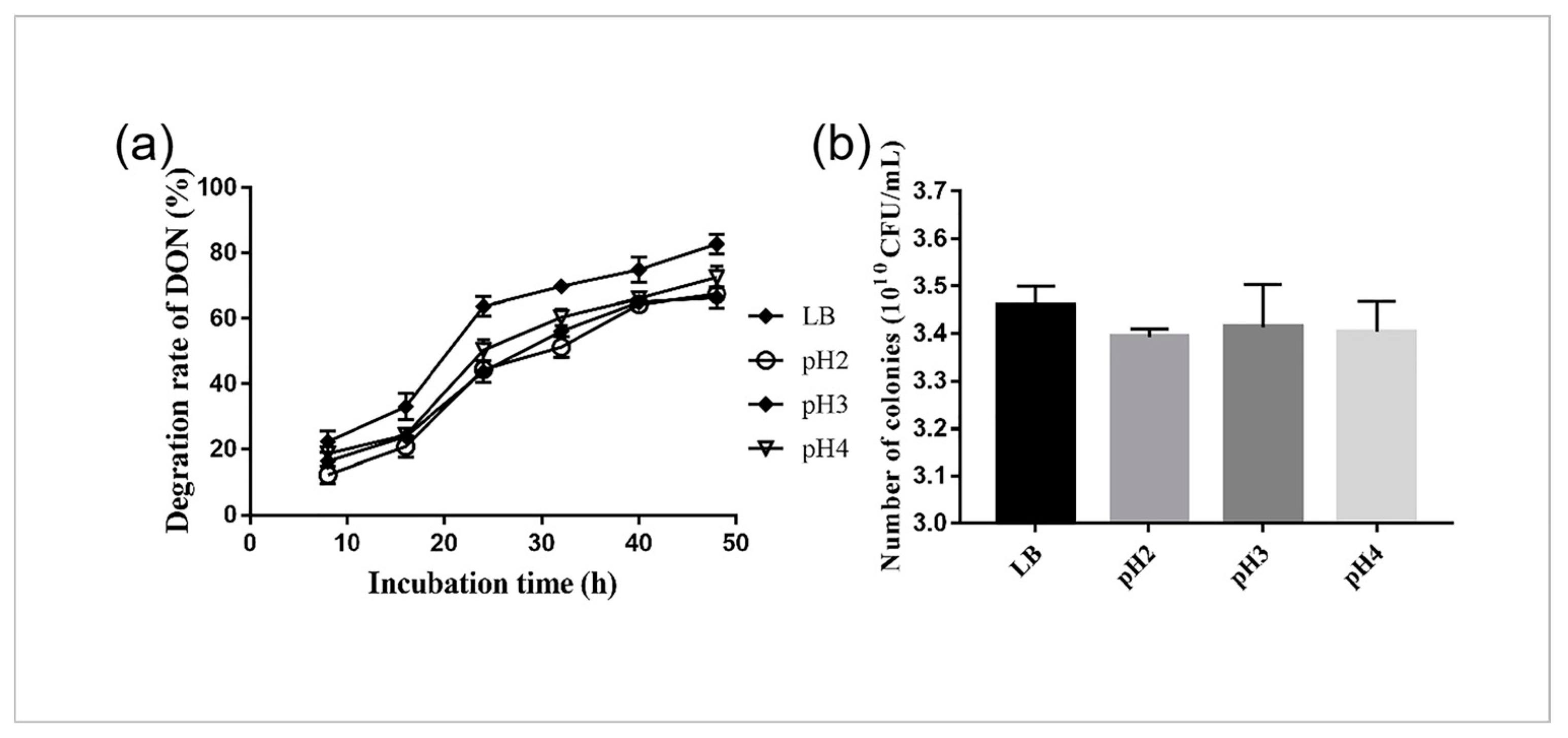
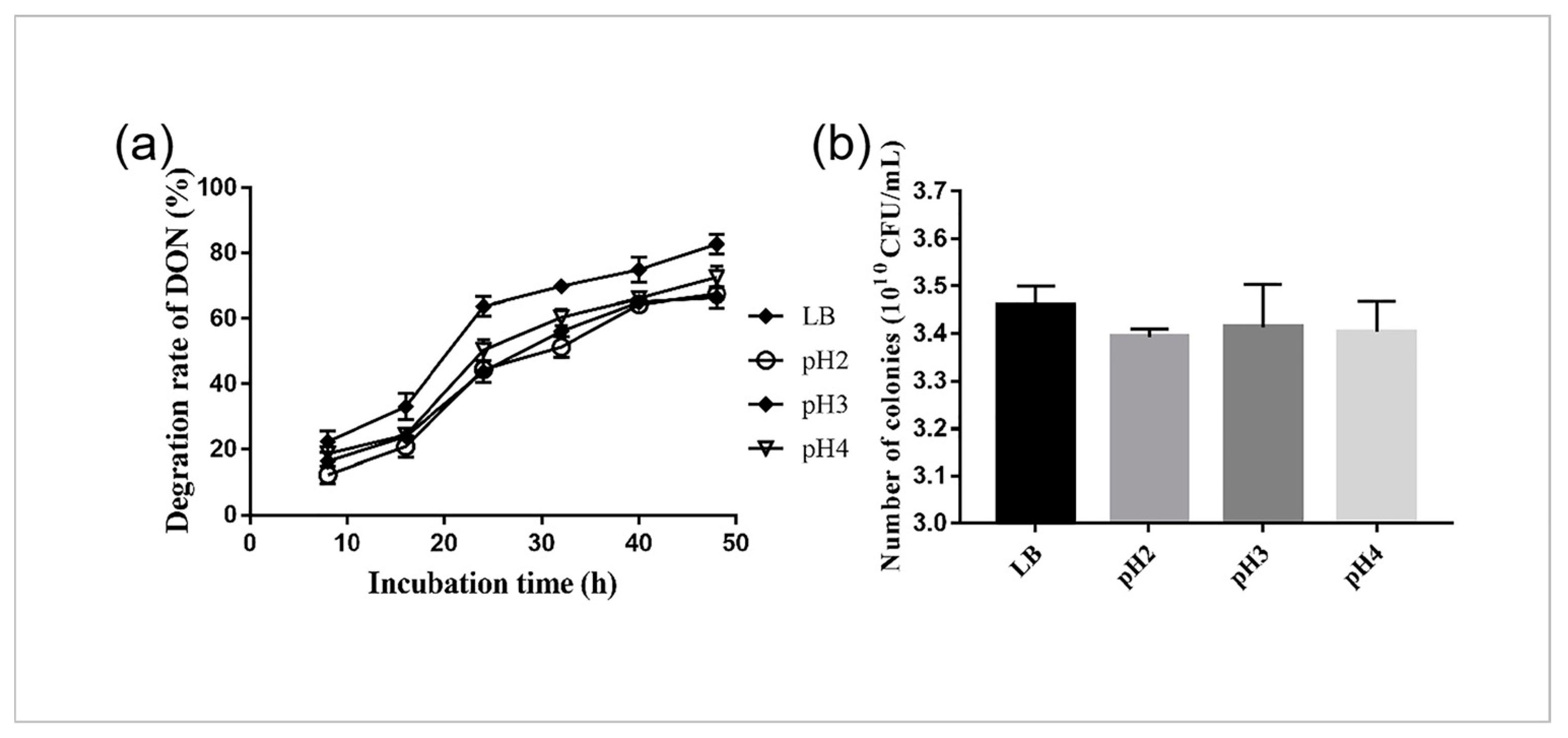
2.2. Survival and DON Degradation of YB9 in SGF
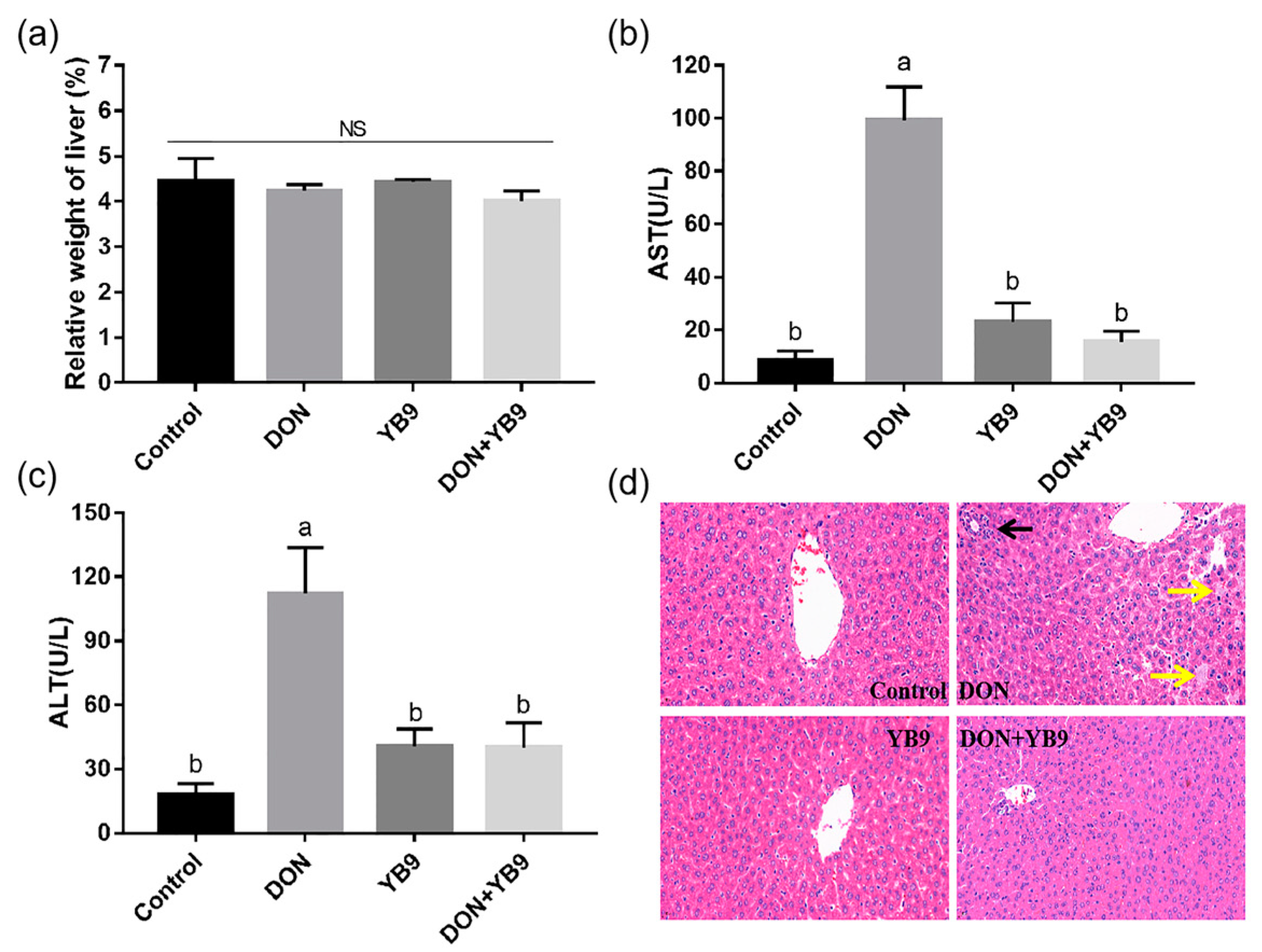
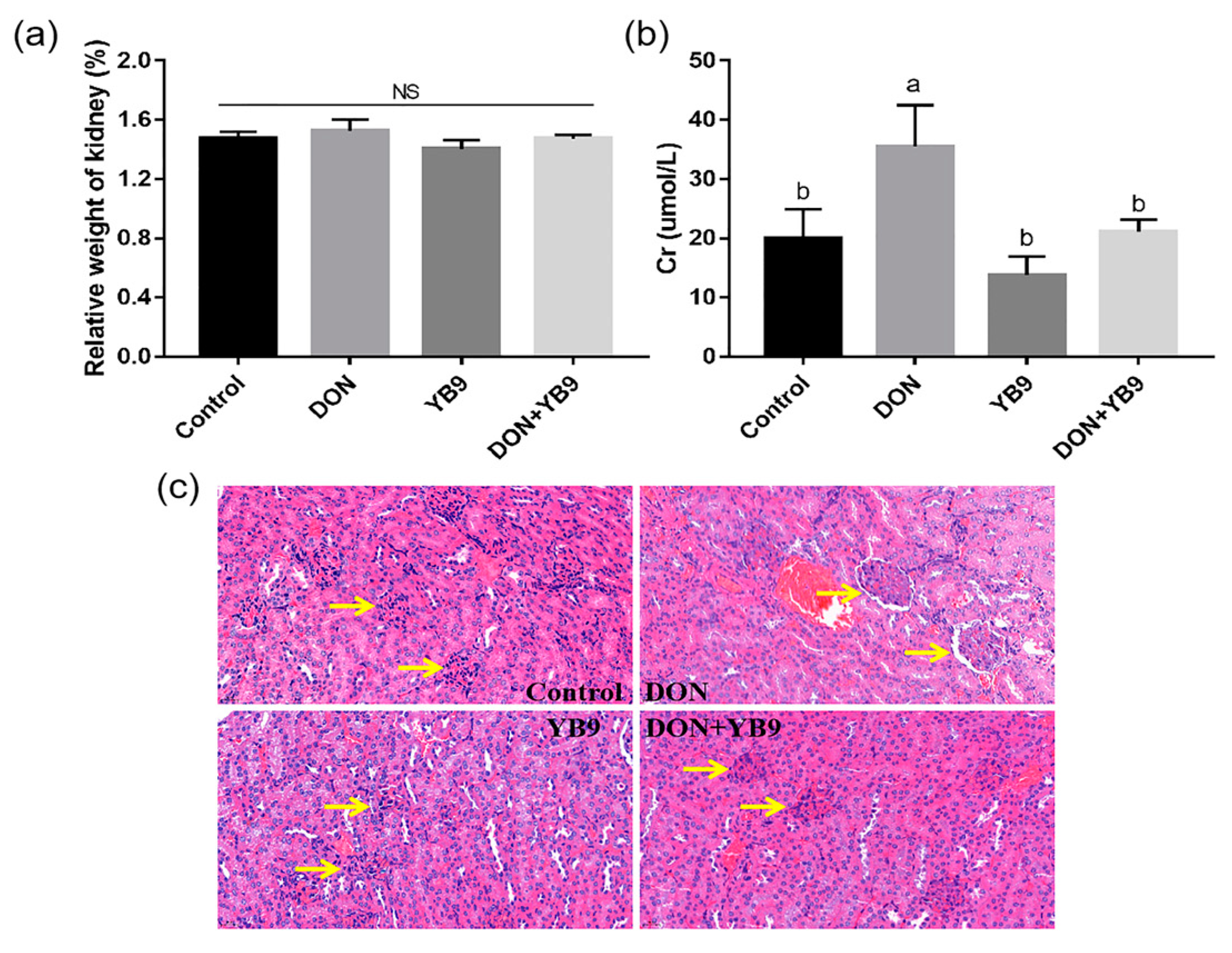
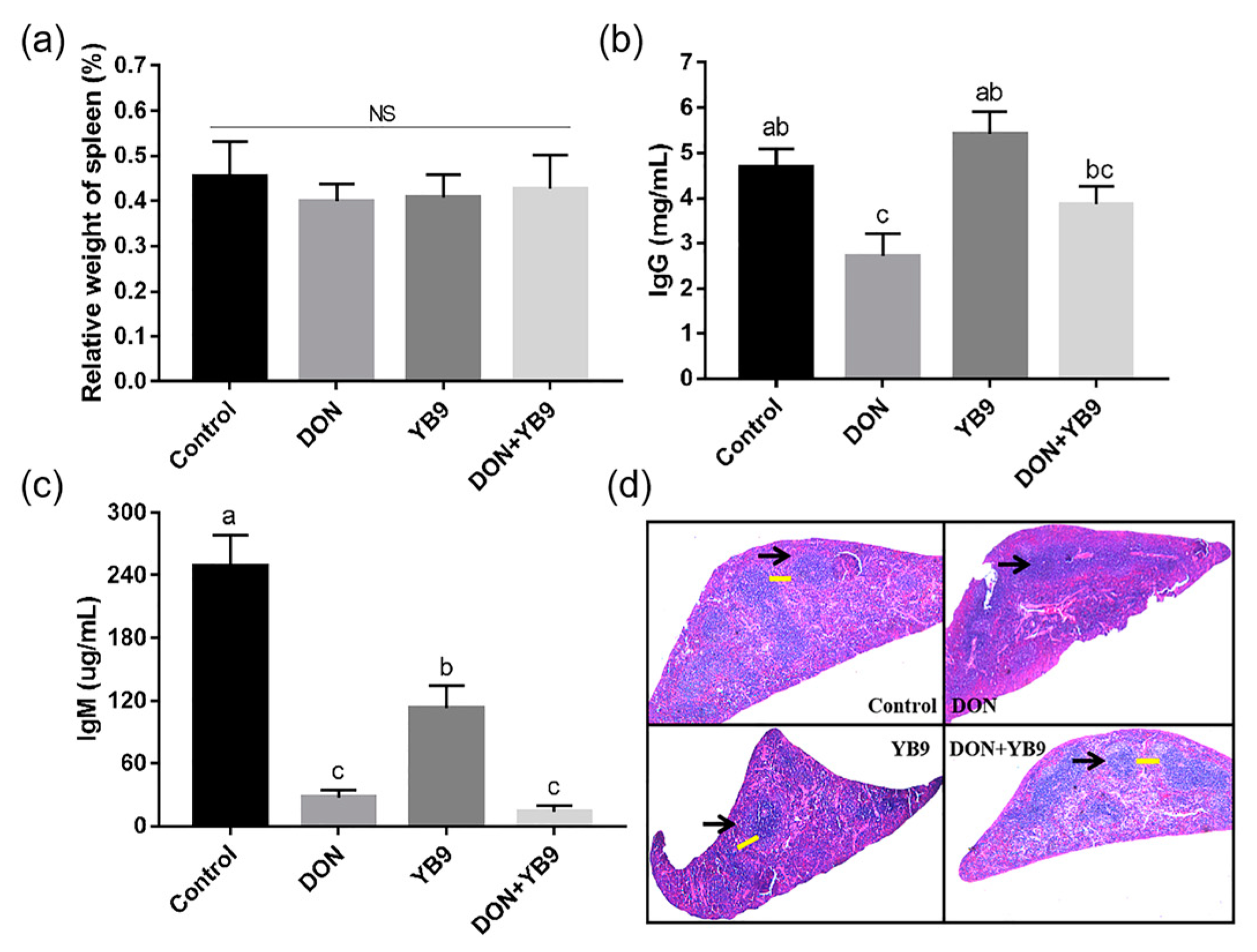
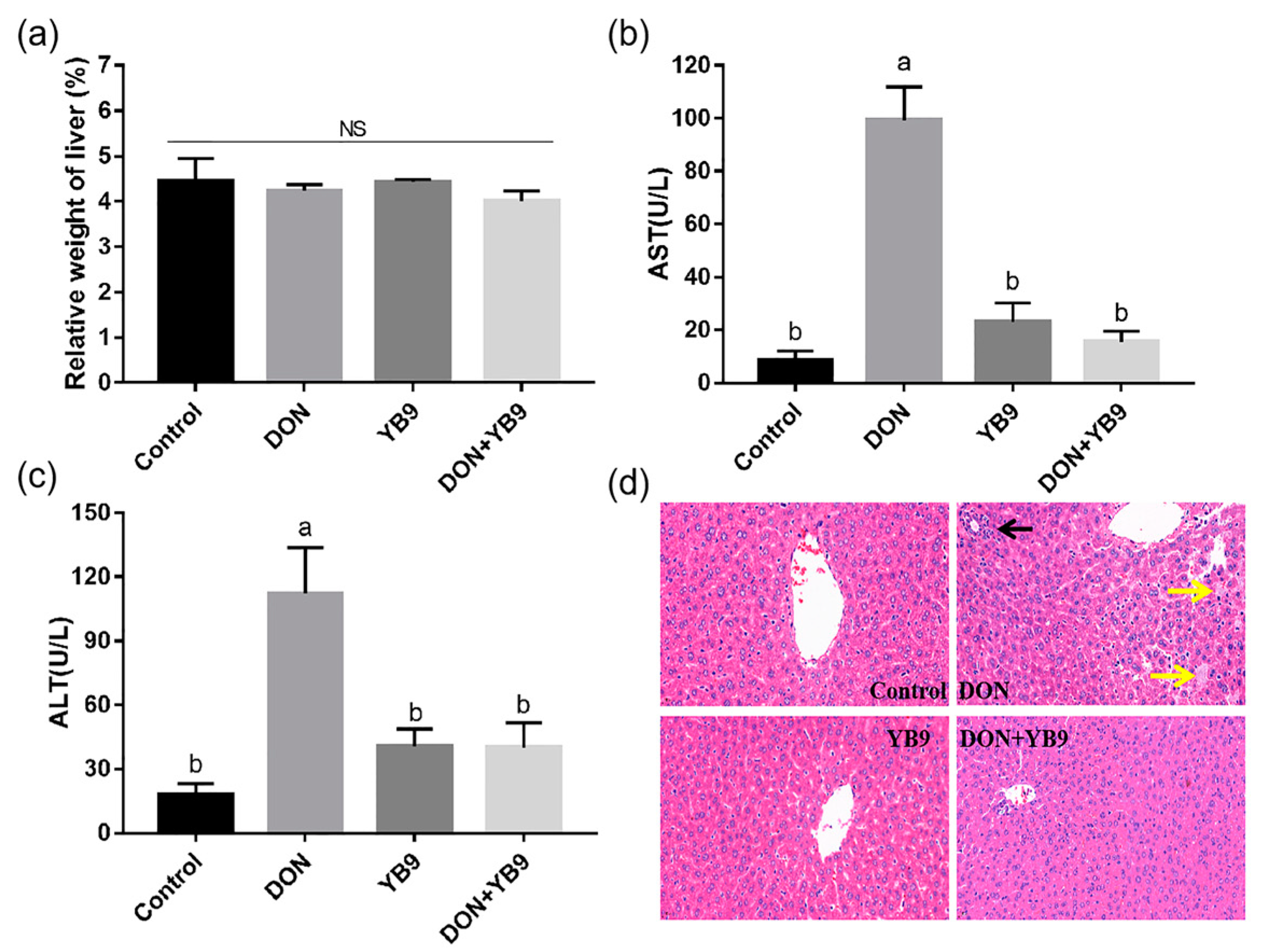
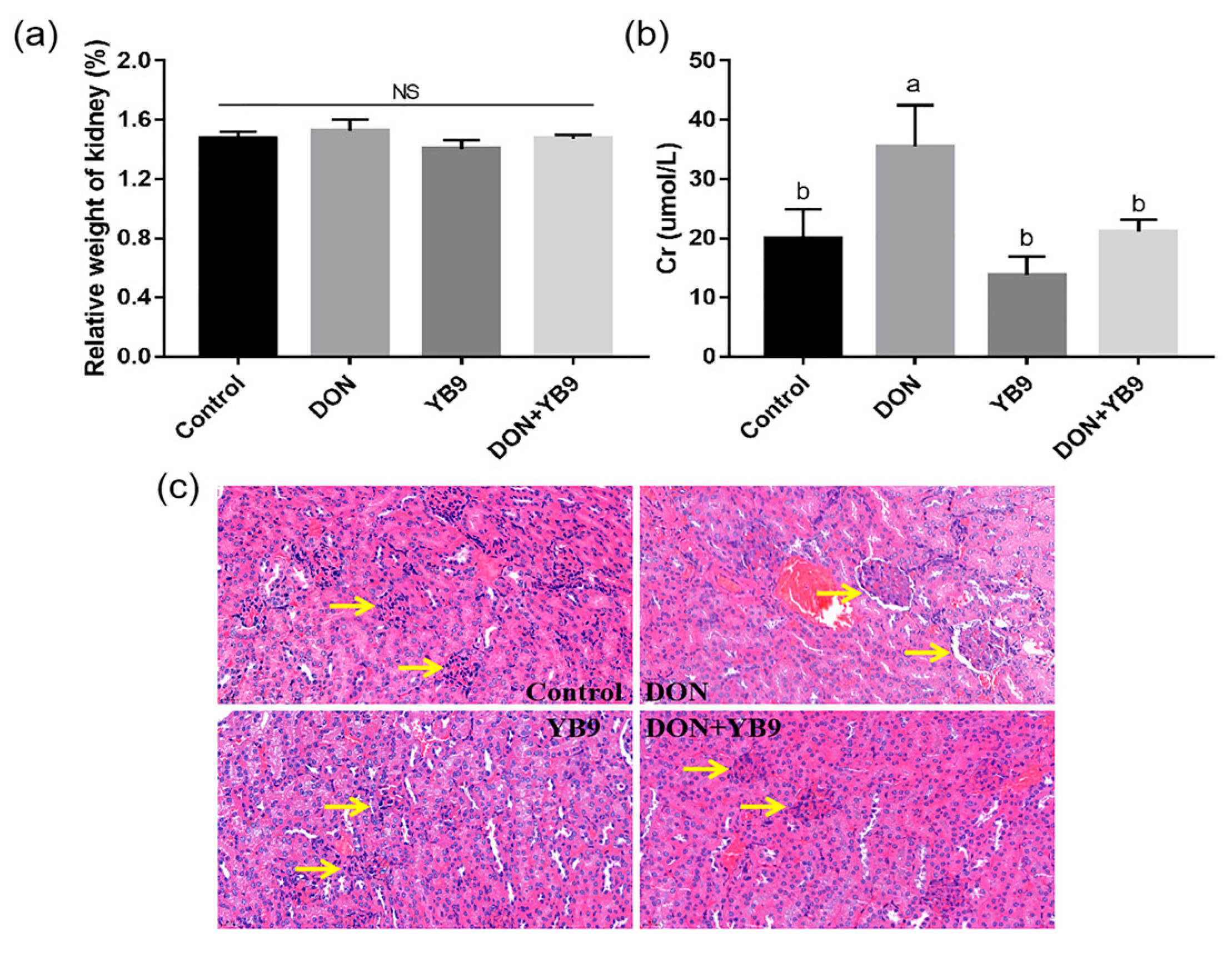
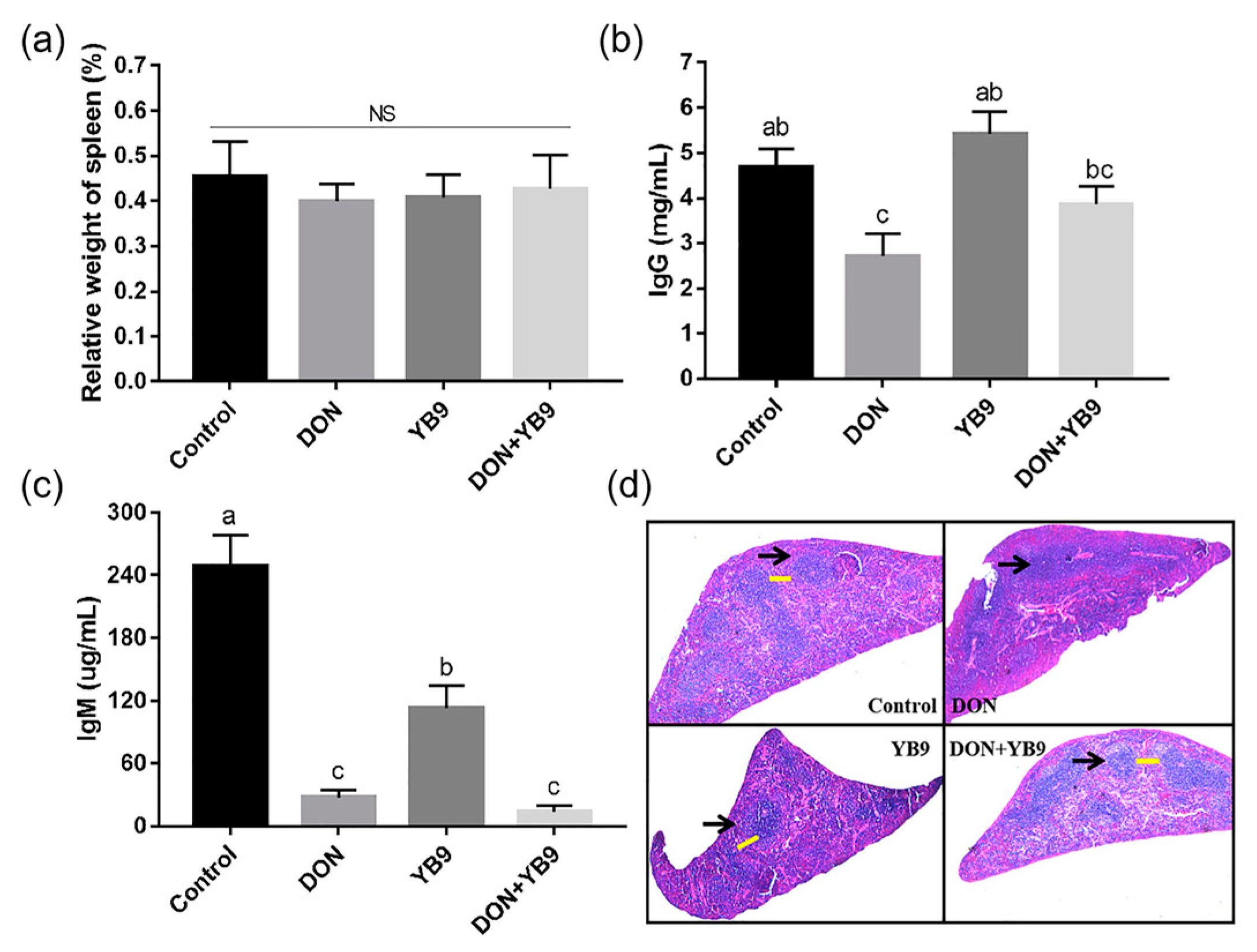
2.3. Protection of YB9 Against the Damage of Liver, Kidney, and Spleen Caused by DON
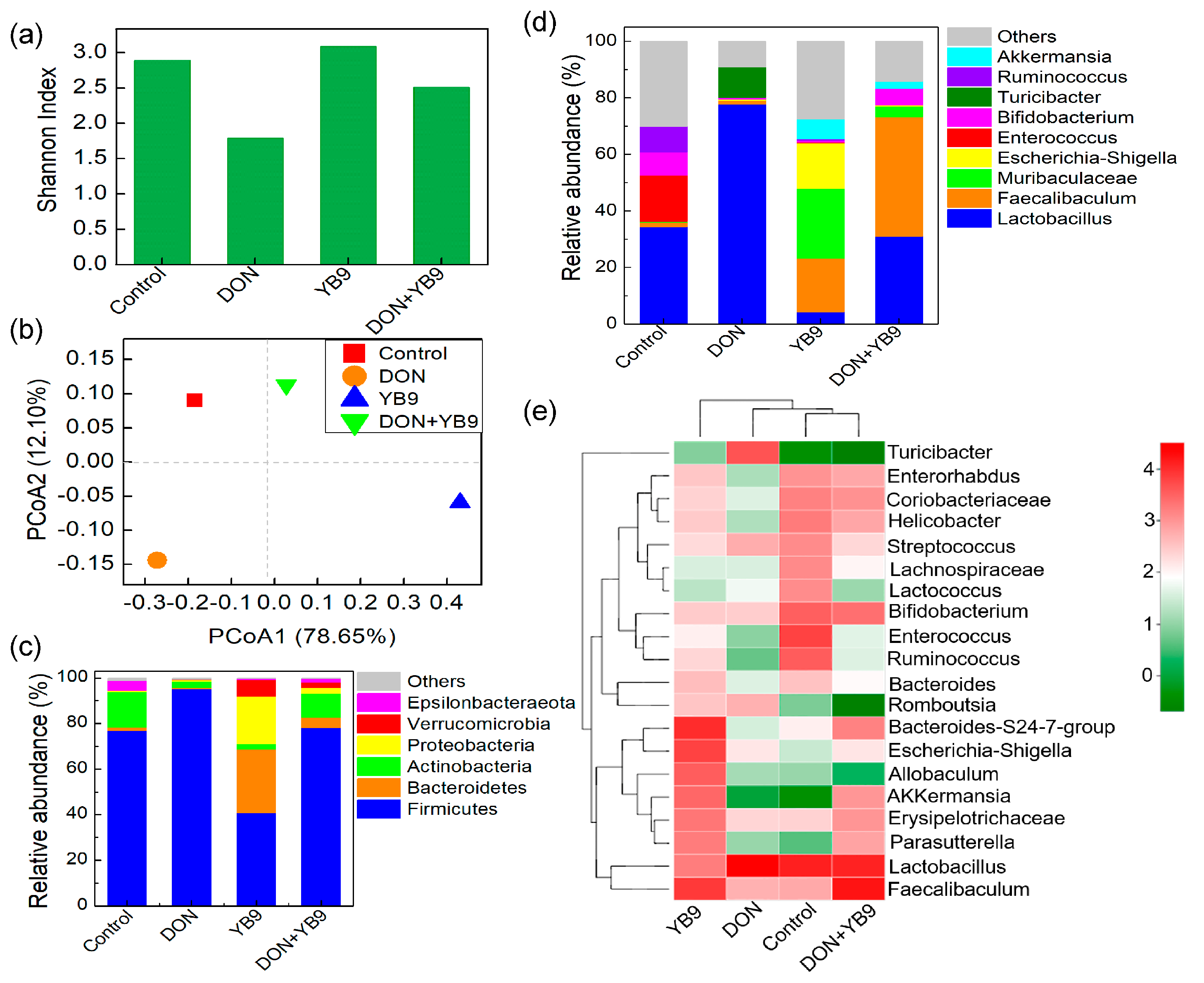
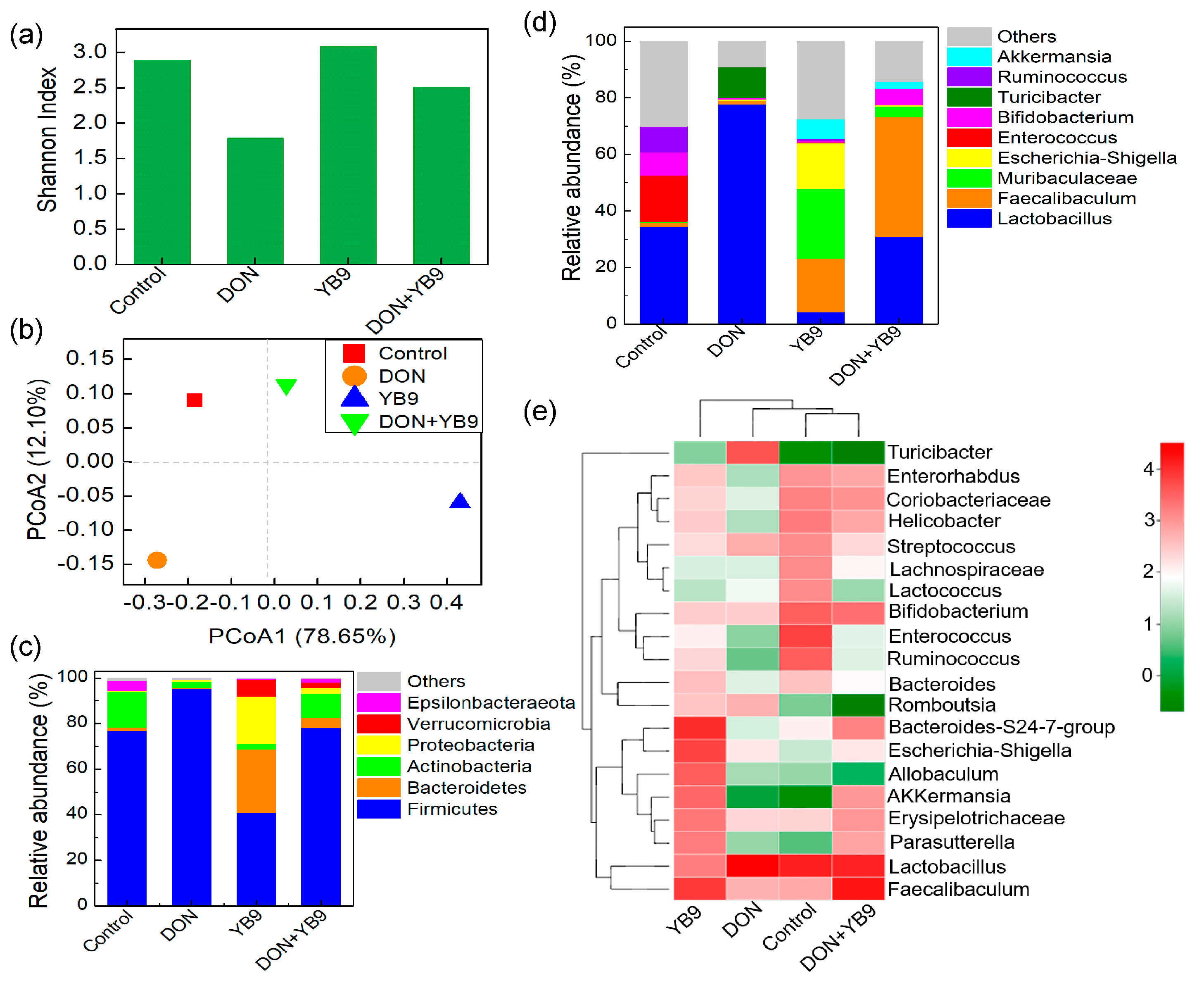
2.4. Effects of DON and YB9 on the Intestinal Flora of Mice
3. Discussion
4. Materials and Methods
4.1. Culture Media and Reagents
4.2. Isolation of Microorganisms with DON-Removal Ability
4.3. Characterization of the DON-Degrading Strain YB9
4.4. DON Degradation and Survival of YB9 in Simulated Gastric Fluid (SGF)
4.5. Design of Animal Experiments
4.5.1. Organ Weight
4.5.2. Biochemical Parameter Examination
4.5.3. Histopathological Examination
4.5.4. Determination of the Effects of YB9 and DON on Gut Microbiota
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Human and Animal Rights and Informed Consent
References
- Wang, L.L.; Kong, W.J.; Yang, M.H.; Han, J.P.; Chen, S.L. Safety issues and new rapid detection methods in traditional Chinese medicinal materials. Acta Pharm. Sin. B 2015, 5, 38–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delmulle, B.; Saeger, S.D.; Adams, A.; Kimpe, N.D.; Peteghem, C.V. Development of a liquid chromatography/tandem mass spectrometry method for the simultaneous determination of 16 mycotoxins on cellulose filters and in fungal cultures. Rapid Commun. Mass Spectrom. 2006, 20, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Wells, L.; Hardie, L.; Williams, C.; White, K.; Liu, Y.; De Santis, B.; Debegnach, F.; Moretti, G.; Greetham, S.; Brera, C.; et al. Determination of deoxynivalenol in the urine of pregnant women in the UK. Toxins 2016, 8, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.S.; Li, Y.H.; Lin, M.F. Chronic exposure to the Fusarium mycotoxin deoxynivalenol: Impact on performance, immune organ, and intestinal integrity of slow-growing chickens. Toxins 2017, 9, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Springler, A.; Hessenberger, S.; Reisinger, N.; Kern, C.; Nagl, V.; Schatzmayr, G.; Mayer, E. Deoxynivalenol and its metabolite deepoxy-deoxynivalenol: Multi-parameter analysis for the evaluation of cytotoxicity and cellular effects. Mycotoxin Res. 2017, 33, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Dabrowski, M.; Jakimiuk, E.; Baranowski, M.; Gajecka, M.; Zielonka, L.; Gajecki, M.T. The effect of deoxynivalenol on selected populations of immunocompetent cells in porcine blood-a preliminary study. Molecules 2017, 22, 691. [Google Scholar] [CrossRef] [Green Version]
- Gajecka, M.; Tarasiuk, M.; Zielonka, L.; Dabrowski, M.; Nicpon, J.; Baranowski, M.; Gajecki, M.T. Changes in the metabolic profile and body weight of pre-pubertal gilts during prolonged monotonic exposure to low doses of zearalenone and deoxynivalenol. Toxicon 2017, 125, 32–43. [Google Scholar] [CrossRef]
- Ji, J.; Zhu, P.; Cui, F.; Pi, F.; Zhang, Y.; Sun, X. The disorder metabolic profiling in kidney and spleen of mice induced by mycotoxins deoxynivalenol through gas chromatography mass spectrometry. Chemosphere 2017, 180, 267–274. [Google Scholar] [CrossRef]
- Liang, Z.; Ren, Z.H.; Gao, S.; Chen, Y.; Yang, Y.Y.; Yang, D.; Deng, J.L.; Zuo, Z.C.; Wang, Y.; Shen, L.H. Individual and combined effects of deoxynivalenol and zearalenone on mouse kidney. Environ. Toxicol. Pharmacol. 2015, 40, 686–691. [Google Scholar] [CrossRef]
- Peng, Z.; Chen, L.; Nussler, A.K.; Liu, L.; Yang, W. Current sights for mechanisms of deoxynivalenol-induced hepatotoxicity and prospective views for future scientific research: A mini review. J. Appl. Toxicol. 2017, 37, 518–529. [Google Scholar] [CrossRef]
- Peng, Z.; Liao, Y.; Chen, L.; Liu, S.; Shan, Z.; Nussler, A.K.; Yao, P.; Yan, H.; Liu, L.G.; Yang, W. Heme oxygenase-1 attenuates low-dose of deoxynivalenol-induced liver inflammation potentially associating with microbiota. Toxicol. Appl. Pharmacol. 2019, 374, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Tardivel, C.; Airault, C.; Djelloul, M.; Guillebaud, F.; Barbouche, R.; Troadec, J.D.; Gaigé, S.; Dallaporta, M. The food born mycotoxin deoxynivalenol induces low-grade inflammation in mice in the absence of observed-adverse effects. Toxicol. Lett. 2015, 232, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Liao, P.; He, L.Q.; Ren, W.K.; Yin, J.; Duan, J.L.; Li, T.J. Growth performance, serum biochemical profile, jejunal morphology, and the expression of nutrients transporter genes in deoxynivalenol (DON)-challenged growing pigs. BMC Vet. Res. 2015, 11, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.C.; Wang, J.Q.; Huang, L.B.; Chen, H.J.; Wang, C.Y. Effects of adding clostridium sp. WJ06 on intestinal morphology and microbial diversity of growing pigs fed with natural deoxynivalenol contaminated wheat. Toxins 2017, 9, 383. [Google Scholar] [CrossRef] [Green Version]
- Pestka, J.J.; Islam, Z.; Amuzie, C.J. Immunochemical assessment of deoxynivalenol tissue distribution following oral exposure in the mouse. Toxicol. Lett. 2008, 178, 83–87. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.J.; Jeong, S.H.; Cho, J.H.; Ku, H.O.; Pyo, H.M.; Kang, H.G.; Choi, K.H. Plasma haptoglobin and immunoglobulins as diagnostic indicators of deoxynivalenol intoxication. J. Vet. Sci. 2008, 9, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Reddy, K.E.; Song, J.; Lee, H.J.; Kim, M.; Kim, D.W.; Jung, H.J.; Kim, B.; Lee, Y.; Yu, D.; Kim, D.W.; et al. Effects of high levels of deoxynivalenol and zearalenone on growth performance, and hematological and immunological parameters in pigs. Toxins 2018, 10, 114. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.H.; Wang, Y.C.; Deng, H.D.; Deng, Y.T.; Deng, J.L.; Zuo, Z.C.; Wang, Y.; Peng, X.; Cui, H.M.; Shen, L.H. Deoxynivalenol induces apoptosis in chicken splenic lymphocytes via the reactive oxygen species-mediated mitochondrial pathway. Environ. Toxicol. Pharmacol. 2015, 39, 339–346. [Google Scholar] [CrossRef]
- Awad, W.; Ghareeb, K.; Bohm, J.; Zentek, J. The toxicological impacts of the Fusarium mycotoxin, deoxynivalenol, in poultry flocks with special reference to immunotoxicity. Toxins 2013, 5, 912–925. [Google Scholar] [CrossRef]
- Pestka, J.J.; Zhou, H.R.; Moon, Y.; Chung, Y.J. Cellular and molecular mechanisms for immune modulation by deoxynivalenol and other trichothecenes: Unraveling a paradox. Toxicol. Lett. 2004, 153, 61–73. [Google Scholar] [CrossRef]
- Moon, Y.; Kim, H.K.; Suh, H.; Chung, D.H. Toxic alterations in chick embryonic liver and spleen by acute exposure to Fusarium-producing mycotoxin deoxynivalenol. Biol. Pharm. Bull. 2007, 30, 1808–1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wache, Y.J.; Valat, C.; Postollec, G.; Bougeard, S.; Burel, C.; Oswald, I.P.; Fravalo, P. Impact of deoxynivalenol on the intestinal microflora of pigs. Int. J. Mol. Sci. 2009, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Payros, D.; Alassane-Kpembi, I.; Pierron, A.; Loiseau, N.; Pinton, P.; Oswald, I.P. Toxicology of deoxynivalenol and its acetylated and modified forms. Arch. Toxicol. 2016, 90, 2931–2957. [Google Scholar] [CrossRef] [PubMed]
- Sugita-Konishi, Y.; Park, B.J.; Kobayashi-Hattori, K.; Tanaka, T.; Chonan, T.; Yoshikawa, K.; Kumagai, S. Effect of cooking process on the deoxynivalenol content and its subsequent cytotoxicity in wheat products. Biosci. Biotechnol. Biochem. 2006, 70, 1764–1768. [Google Scholar] [CrossRef] [Green Version]
- Wan, P.; Peng, Y.J.; Chen, G.J.; Xie, M.H.; Dai, Z.Q.; Huang, K.Y.; Dong, W.; Zeng, X.X.; Sun, Y. Modulation of gut microbiota by ilex kudingcha improves dextran sulfate sodium-induced colitis. Food Res. Int. 2019, 126, 108595. [Google Scholar] [CrossRef]
- Islam, R.; Zhou, T.; Young, J.C.; Goodwin, P.H.; Pauls, K.P. Aerobic and anaerobic de-epoxydation of mycotoxin deoxynivalenol by bacteria originating from agricultural soil. World J. Microbiol. Biotechnol. 2012, 28, 7–13. [Google Scholar] [CrossRef]
- Ahad, R.; Zhou, T.; Leppa, D.; Pauls, K.P. Draft genome sequence of Citrobacter freundii strain A47, resistant to the mycotoxin deoxynivalenol. Genome Announc. 2017, 5, e17–e19. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.H.; Li, X.Y.; Ji, C.; Rong, X.P.; Liu, S.J.; Zhang, J.Y.; Ma, Q.G. Protective effect of Devosia sp. ANSB714 on growth performance, serum chemistry, immunity function and residues in kidneys of mice exposed to deoxynivalenol. Food Chem. Toxicol 2016, 92, 143–149. [Google Scholar] [CrossRef]
- Zhu, Y.; Hassan, Y.I.; Lepp, D.; Shao, S.Q.; Zhou, T. Strategies and nethodologies for developing microbial detoxification systems to mitigate mycotoxins. Toxins 2017, 9, 130. [Google Scholar] [CrossRef] [Green Version]
- Cao, G.T.; Dai, B.; Wang, K.L.; Yan, Y.; Xu, Y.L.; Wang, Y.X.; Yang, C.M. Bacillus licheniformis, a potential probiotic, inhibits obesity by modulating colonic microflora in C57BL/6J mice model. J. Appl. Microbiol. 2019, 127, 880–888. [Google Scholar] [CrossRef]
- Hsu, T.C.; Yi, P.J.; Lee, T.Y.; Liu, J.R. Probiotic characteristics and zearalenone-removal ability of a Bacillus licheniformis strain. PLoS ONE 2018, 13, e0194866. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Y.; Shao, H.L.; Luo, X.H.; Wang, R.; Li, Y.F.; Li, Y.N.; Luo, Y.P.; Zhang, D.J.; Chen, Z.X. In vivo toxicity assessment of deoxynivalenol-contaminated wheat after ozone degradation. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2017, 34, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Zhang, R.Q.; Zhai, Q.Y.; Liu, J.C.; Li, N.; Liu, W.X.; Li, L.; Shen, W. Metagenomic analysis of gut microbiota alteration in a mouse model exposed to mycotoxin deoxynivalenol. Toxicol. Appl. Pharmacol. 2019, 372, 47–56. [Google Scholar] [CrossRef] [PubMed]
- DelPiano, M.; Carmagnola, S.; Anderloni, A.; Andorno, S.; Ballare, M.; Balzarini, M.; Montino, F.; Orsello, M.; Pagliarulo, M.; Sartori, M.; et al. The use of probiotics in healthy volunteers with evacuation disorders and hard stools: A double-blind, randomized, placebo-controlled study. J. Clin. Gastroenterol. 2010, 44, S30–S34. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, J.; Wang, Y.L.; Wang, K.R.; Wei, H.; Shen, L. Isolation and characterization of the Bacillus cereus BC7 strain, which is capable of zearalenone removal and intestinal flora modulation in mice. Toxicon 2018, 155, 9–20. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, H.H.; Zhao, C.; Han, Y.T.; Liu, Y.C.; Zhang, X.L. Isolation and characterization of a novel deoxynivalenol-transforming strain paradevosia shaoguanensis DDB001 from wheat field soil. Lett. Appl. Microbiol. 2017, 65, 414–422. [Google Scholar] [CrossRef]
- Shieh, G. Power and sample size calculations for comparison of two regression lines with heterogeneous variances. PLoS ONE 2018, 13, e0207745. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate/Test | Result | Substrate/Test | Result |
|---|---|---|---|
| Sugar fermentation | + | Amylohydrolysis | + |
| 3%H2O2 | + | 7% NaCl | + |
| Semi-solid agar | + | Indole | − |
| Nitrate | + | Citrate utilization | + |
| Methyl red test | − | pH 5.5–8.7 | + |
| V-P | + | Casein hydrolysis | + |
| Yolk | − |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Hou, Q.; Guo, Q.; Zhang, J.; Sun, Y.; Wei, H.; Shen, L. Isolation and Characterization of a Deoxynivalenol-Degrading Bacterium Bacillus licheniformis YB9 with the Capability of Modulating Intestinal Microbial Flora of Mice. Toxins 2020, 12, 184. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12030184
Wang S, Hou Q, Guo Q, Zhang J, Sun Y, Wei H, Shen L. Isolation and Characterization of a Deoxynivalenol-Degrading Bacterium Bacillus licheniformis YB9 with the Capability of Modulating Intestinal Microbial Flora of Mice. Toxins. 2020; 12(3):184. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12030184
Chicago/Turabian StyleWang, Shiwei, Qiuqiu Hou, Qianqian Guo, Jian Zhang, Yanmei Sun, Hong Wei, and Lixin Shen. 2020. "Isolation and Characterization of a Deoxynivalenol-Degrading Bacterium Bacillus licheniformis YB9 with the Capability of Modulating Intestinal Microbial Flora of Mice" Toxins 12, no. 3: 184. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12030184