Cylindrospermopsin- and Deoxycylindrospermopsin-Producing Raphidiopsis raciborskii and Microcystin-Producing Microcystis spp. in Meiktila Lake, Myanmar

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Physico-Chemical Parameters
2.2. Phytoplankton Community
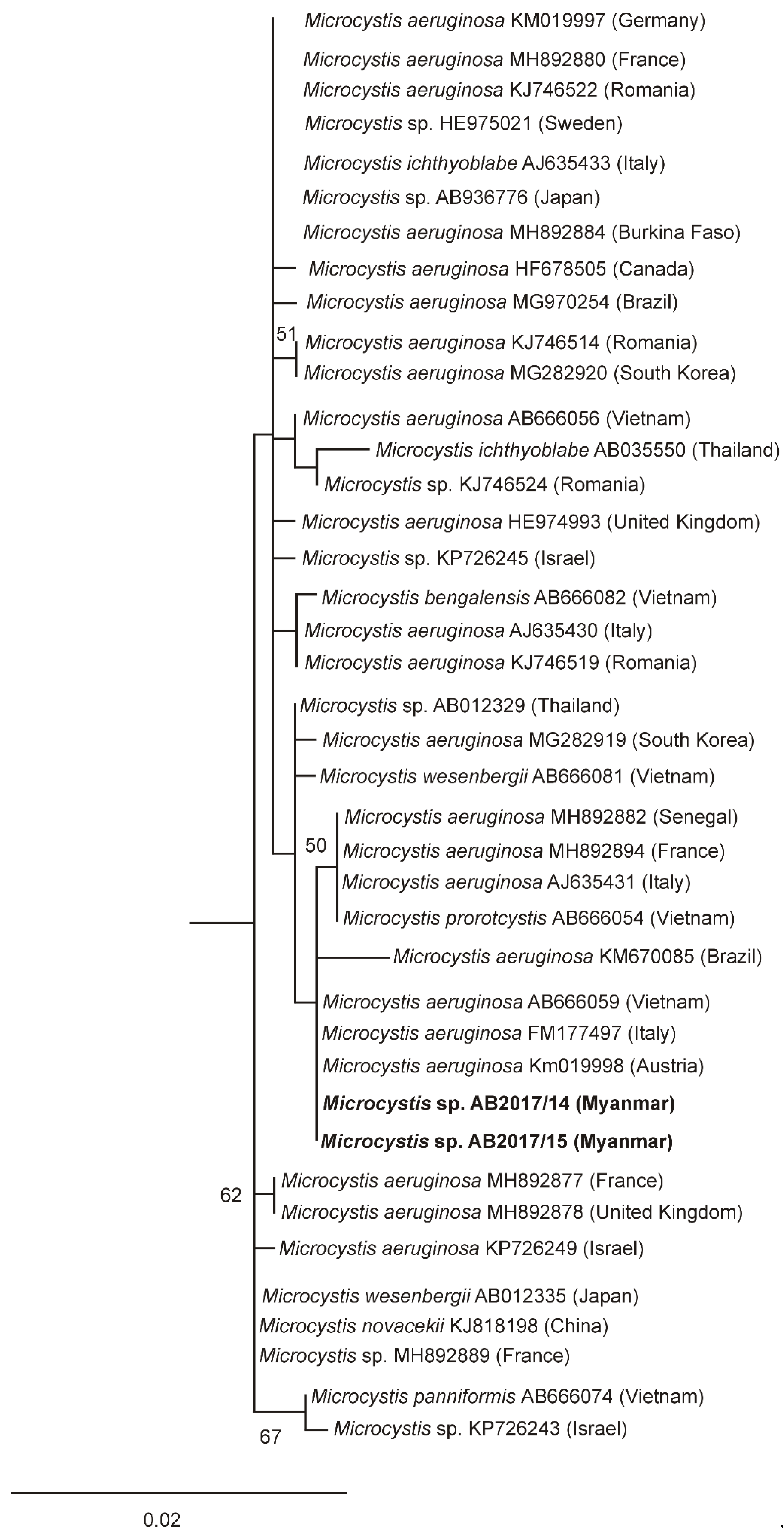
2.3. Morphological and Phylogenetic Characterization
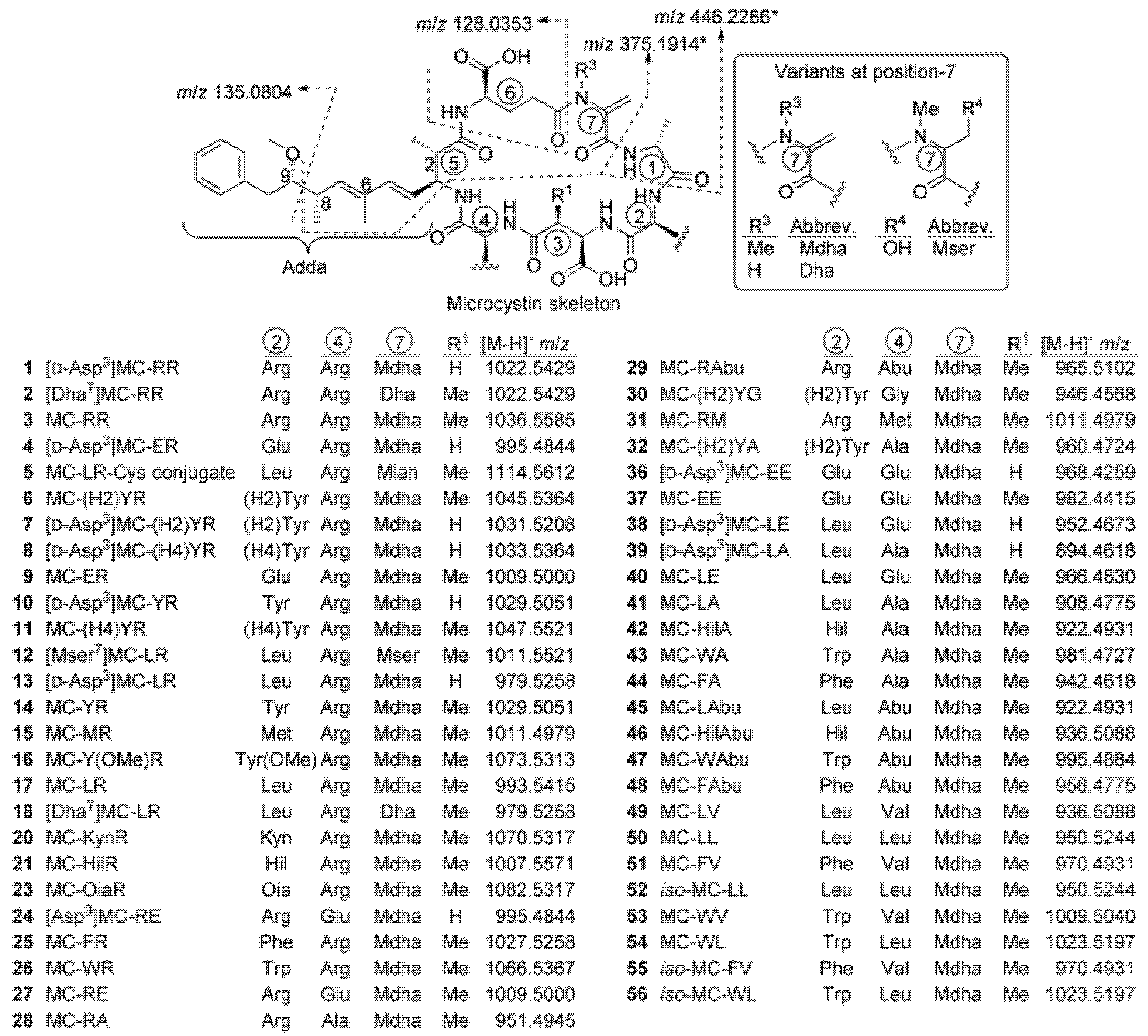
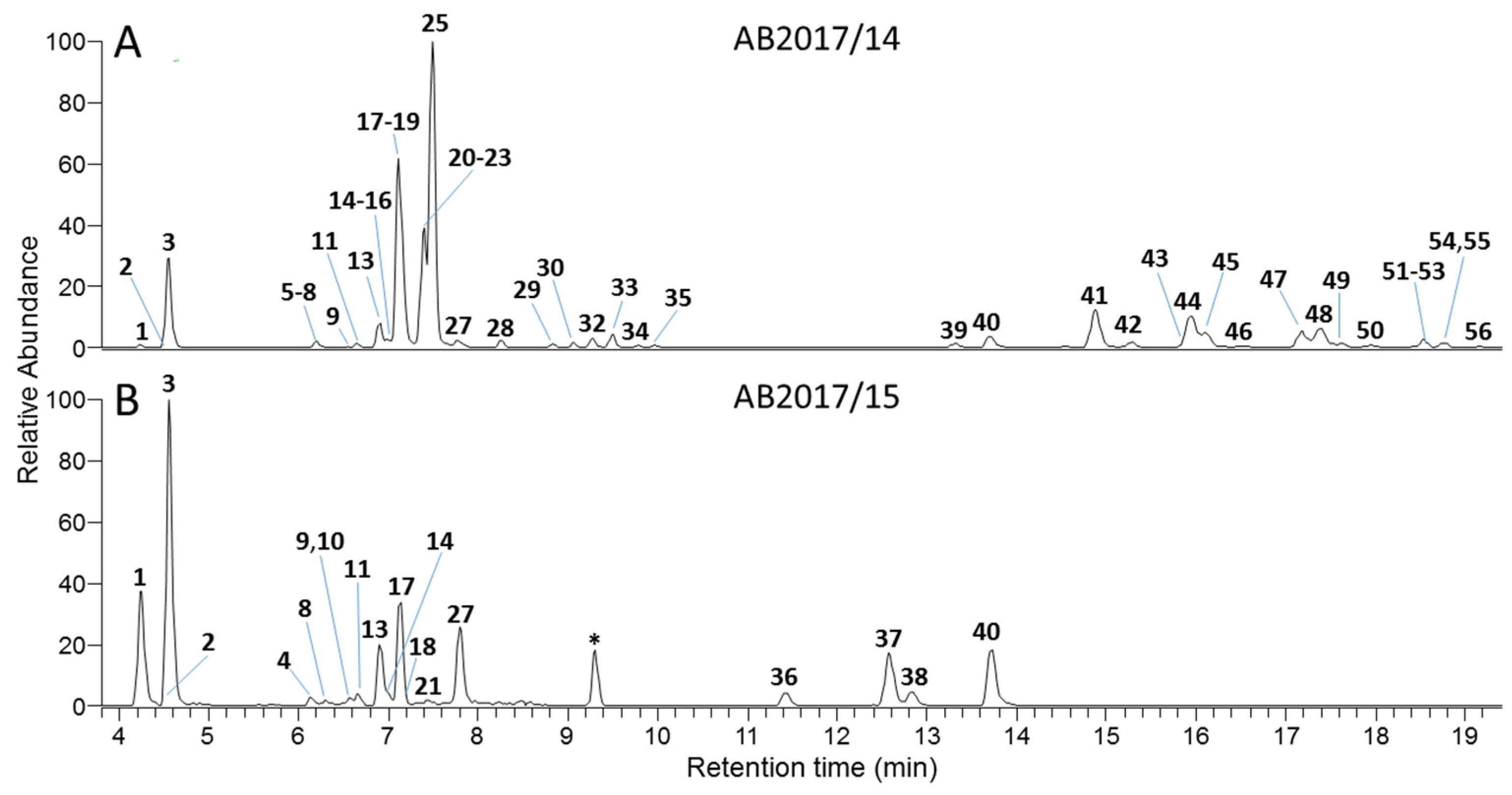
2.4. Identification of Cyanobacterial Toxins and Toxin-Producing Strains
3. Discussion
4. Conclusions
5. Materials and Methods
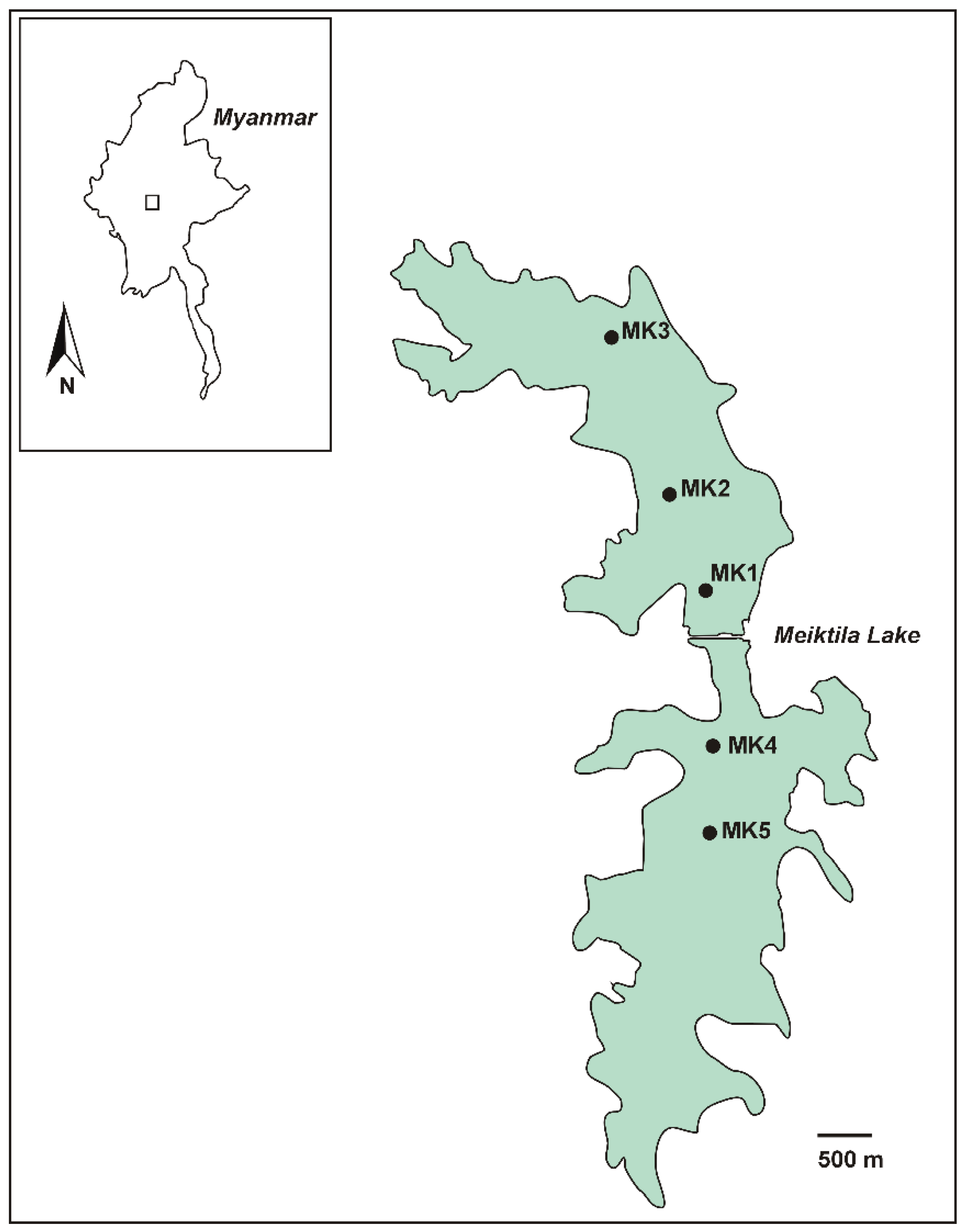
5.1. Study Area, Measurements and Sampling
5.2. Phytoplankton Analysis
5.3. Isolation of Strains and Morphological Characterization
5.4. Genomic DNA Extraction, PCR Amplification and Sequencing
5.5. Phylogenetic Analysis
5.6. Toxin Analysis
5.6.1. Material
5.6.2. ELISA for MCs, CYNs, ATXs and STXs
5.6.3. Microcystin Analysis by LC–HRMS
5.6.4. CYN, deoxyCYN, ATXs and STXs Analysis by LC-MS/MS
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Paerl, H.W.; Huisman, J. Climate change: a catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K. Cyanobacterial toxins. In Encyclopedia of Microbiology; Schaechter, M., Ed.; Elsevier: Oxford, UK, 2009; pp. 290–307. [Google Scholar]
- Harvey, G.E. History of Burma from the Earliest Times to 10 March 1824: The Beginning of the English Conquest, 1; Longmans, Green and Company: London, UK, 1925. [Google Scholar]
- Nyunt, K.K. Algal flora of Meiktila Lake and its surrounding area. Master´s Thesis, Mandalay University, Mandalay, Myanmar, 1995. [Google Scholar]
- Hlaing, N.K. Limnological study of Meiktila Lake, Mandalay Division. Ph.D. Thesis, University of Mandalay, Mandalay, Myanmar, 2014. [Google Scholar]
- Klink, T.; de Jongh, I.; Leushuis, M.; Bijlmakers, L.; Speets, R.; Brinkman, L. Scoping Study Meiktila Lake Area IWRM; Royal HaskoningDHV, Arcadis, Rebel Group, Dutch Water Authorities: Den Haag, The Netherlands, 2015; pp. 1–50. [Google Scholar]
- Aung, T.T.; Zin, T. Zooplankton community in Meiktila Lake, Meiktila, central Myanmar; Universities Research Journal; The Government of The Republic of the Union of Myanmar Ministry of Education: Yangon, Myanmar, 2015; Volume 7, pp. 1–11. [Google Scholar]
- Antunes, J.T.; Leao, P.N.; Vasconcelos, V.M. Cylindrospermopsis raciborskii: Review of the distribution, phylogeography, and ecophysiology of a global invasive species. Front. Microbiol. 2015, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burford, M.A.; Beardall, J.; Willis, A.; Orr, P.T.; Magalhaes, V.F.; Rangel, L.M.; Azevedo, S.; Neilan, B.A. Understanding the winning strategies used by the bloom-forming cyanobacterium Cylindrospermopsis raciborskii. Harmful Algae 2016, 54, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Mowe, M.A.D.; Mitrovic, S.M.; Lim, R.P.; Furey, A.; Yeo, D.C.J. Tropical cyanobacterial blooms: A review of prevalence, problem taxa, toxins and influencing environmental factors. J. Limnol. 2015, 74, 205–224. [Google Scholar] [CrossRef] [Green Version]
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A review of the global ecology, genomics, and biogeography of the toxic cyanobacterium, Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [Green Version]
- Mallia, V.; Uhlig, S.; Rafuse, C.; Meija, J.; Miles, C.O. Novel microcystins from Planktothrix prolifica NIVA-CYA 544 identified by LC-MS/MS, functional group derivatization and 15N-labeling. Mar. Drugs 2019, 17, 643. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, M.; Foss, A.J.; Miles, C.O.; Özen, M.; Demir, N.; Balci, M.; Beach, D.G. Comprehensive multi-technique approach reveals the high diversity of microcystins in field collections and an associated isolate of Microcystis aeruginosa from a Turkish lake. Toxicon 2019, 167, 87–100. [Google Scholar] [CrossRef]
- Miles, C.O.; Sandvik, M.; Haande, S.; Nonga, H.; Ballot, A. LC-MS analysis with thiol derivatization to differentiate [Dhb7]- from [Mdha7]-microcystins: analysis of cyanobacterial blooms, Planktothrix cultures and European crayfish from Lake Steinsfjorden, Norway. Environ. Sci. Technol. 2013, 47, 4080–4087. [Google Scholar] [CrossRef]
- Miles, C.O.; Melanson, J.E.; Ballot, A. Sulfide oxidations for LC-MS analysis of methionine-containing microcystins in Dolichospermum flos-aquae NIVA-CYA 656. Environ. Sci. Technol. 2014, 48, 13307–13315. [Google Scholar] [CrossRef]
- Lürling, M.; Van Oosterhout, F.; Faassen, E. Eutrophication and warming boost cyanobacterial biomass and microcystins. Toxins 2017, 9, 64. [Google Scholar] [CrossRef] [Green Version]
- Guedes, I.A.; Rachid, C.T.C.C.; Rangel, L.M.; Silva, L.H.S.; Bisch, P.M.; Azevedo, S.M.F.O.; Pacheco, A.B.F. Close link between harmful cyanobacterial dominance and associated bacterioplankton in a tropical eutrophic reservoir. Front. Microbiol. 2018, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Humpage, A.R.; Falconer, I.R. Oral toxicity of the cyanobacterial toxin cylindrospermopsin in male Swiss albino mice: determination of no observed adverse effect level for deriving a drinking water guideline value. Environ. Toxicol. 2003, 18, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Norris, R.L.; Eaglesham, G.K.; Pierens, G.; Shaw, G.R.; Smith, M.J.; Chiswell, R.K.; Seawright, A.A.; Moore, M.R. Deoxycylindrospermopsin, an analog of cylindrospermopsin from Cylindrospermopsis raciborskii. Environ. Toxicol. 1999, 14, 163–165. [Google Scholar] [CrossRef]
- Neumann, C.; Bain, P.; Shaw, G. Studies of the comparative in vitro toxicology of the cyanobacterial metabolite deoxycylindrospermopsin. J. Toxicol. Environ. Health, Part A 2007, 70, 1679–1686. [Google Scholar] [CrossRef]
- Machado, J.; Campos, A.; Vasconcelos, V.; Freitas, M. Effects of microcystin-LR and cylindrospermopsin on plant–soil systems: a review of their relevance for agricultural plant quality and public health. Environ. Res. 2017, 153, 191–204. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, D.J.; Saker, M.L. The Palm Island mystery disease 20 years on: A review of research on the cyanotoxin cylindrospermopsin. Environ. Toxicol. 2003, 18, 78–93. [Google Scholar] [CrossRef]
- Chiswell, R.K.; Shaw, G.R.; Eaglesham, G.; Smith, M.J.; Norris, R.L.; Seawright, A.A.; Moore, M.R. Stability of cylindrospermopsin, the toxin from the cyanobacterium, Cylindrospermopsis raciborskii: Effect of pH, temperature, and sunlight on decomposition. Environ. Toxicol. 1999, 14, 155–161. [Google Scholar] [CrossRef]
- Environmental Protection Agency. Cyanobacteria and Cyanotoxins: Information for Drinking Water Systems; United States Environmental Protection Agency: Washington, DC, USA, 2014; pp. 1–11. [Google Scholar]
- Li, R.H.; Carmichael, W.W.; Brittain, S.; Eaglesham, G.K.; Shaw, G.R.; Liu, Y.D.; Watanabe, M.M. First report of the cyanotoxins cylindrospermopsin and deoxycylindrospermopsin from Raphidiopsis curvata (cyanobacteria). J. Phycol. 2001, 37, 1121–1126. [Google Scholar] [CrossRef]
- Nguyen, T.T.L.; Hoang, T.H.; Nguyen, T.K.; Duong, T.T. The occurrence of toxic cyanobacterium Cylindrospermopsis raciborskii and its toxin cylindrospermopsin in the Huong River, Thua Thien Hue province, Vietnam. Environ. Monit. Assess. 2017, 189, 11. [Google Scholar] [CrossRef]
- Dao, T.S.; Cronberg, G.; Nimptsch, J.; Do-Hong, L.-C.; Wiegand, C. Toxic cyanobacteria from Tri An Reservoir, Vietnam. Nowa Hedwigia 2010, 90, 433–448. [Google Scholar] [CrossRef]
- Hawkins, P.R.; Chandrasena, N.R.; Jones, G.J.; Humpage, A.R.; Falconer, I.R. Isolation and toxicity of Cylindrospermopsis raciborskii from an ornamental lake. Toxicon 1997, 35, 341–346. [Google Scholar] [CrossRef]
- Saker, M.L.; Neilan, B.A. Varied diazotrophies, morphologies, and toxicities of genetically similar isolates of Cylindrospermopsis raciborskii (Nostocales, Cyanophyceae) from northern Australia. Appl. Environ. Microbiol. 2001, 67, 1839–1845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Carmichael, W.W.; Brittain, S.; Eaglesham, G.K.; Shaw, G.R.; Mahakhant, A.; Noparatnaraporn, N.; Yongmanitchai, W.; Kaya, K.; Watanabe, M.M. Isolation and identification of the cyanotoxin cylindrospermopsin and deoxy-cylindrospermopsin from a Thailand strain of Cylindrospermopsis raciborskii (Cyanobacteria). Toxicon 2001, 39, 973–980. [Google Scholar] [CrossRef]
- Chonudomkul, D.; Yongmanitchai, W.; Theeragool, G.; Kawachi, M.; Kasai, F.; Kaya, K.; Watanabe, M.M. Morphology, genetic diversity, temperature tolerance and toxicity of Cylindrospermopsis raciborskii (Nostocales, Cyanobacteria) strains from Thailand and Japan. FEMS Microbiol. Ecol. 2004, 48, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Zarenezhad, S.; Sano, T.; Watanabe, M.M.; Kawachi, M. Evidence of the existence of a toxic form of Cylindrospermopsis raciborskii (Nostocales, Cyanobacteria) in Japan. Phycol. Res. 2012, 60, 98–104. [Google Scholar] [CrossRef]
- Cirés, S.; Wörmer, L.; Ballot, A.; Agha, R.; Wiedner, C.; Velazquez, D.; Casero, M.C.; Quesada, A. Phylogeography of cylindrospermopsin and paralytic shellfish toxin-producing Nostocales cyanobacteria from Mediterranean Europe (Spain). Appl. Environ. Microbiol. 2014, 80, 1359–1370. [Google Scholar] [CrossRef] [Green Version]
- Panou, M.; Zervou, S.K.; Kaloudis, T.; Hiskia, A.; Gkelis, S. A Greek Cylindrospermopsis raciborskii strain: missing link in tropic invader’s phylogeography tale. Harmful Algae 2018, 80, 96–106. [Google Scholar] [CrossRef]
- Bouaïcha, N.; Miles, C.O.; Beach, D.G.; Labidi, Z.; Djabri, A.; Benayache, N.Y.; Nguyen-Quang, T. Structural diversity, characterization and toxicology of microcystins. Toxins 2019, 11, 714. [Google Scholar] [CrossRef] [Green Version]
- Foss, A.J.; Miles, C.O.; Samdal, I.A.; Lovberg, K.E.; Wilkins, A.L.; Rise, F.; Jaabaek, J.A.H.; McGowan, P.C.; Aubel, M.T. Analysis of free and metabolized microcystins in samples following a bird mortality event. Harmful Algae 2018, 80, 117–129. [Google Scholar] [CrossRef]
- Okello, W.; Portmann, C.; Erhard, M.; Gademann, K.; Kurmayer, R. Occurrence of microcystin-producing cyanobacteria in Ugandan freshwater habitats. Environ. Toxicol. 2010, 25, 367–380. [Google Scholar] [CrossRef] [Green Version]
- LeBlanc, P.; Merkley, N.; Thomas, K.; Lewis, N.I.; Békri, K.; Renaud, S.L.; Pick, F.R.; McCarron, P.; Miles, C.O.; Quilliam, M.A. Isolation and characterization of [D-Leu1]microcystin-LY from Microcystis aeruginosa CPCC-464. Toxins 2020, 12, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namikoshi, M.; Yuan, M.; Sivonen, K.; Carmichael, W.W.; Rinehart, K.L.; Rouhiainen, L.; Sun, F.; Brittain, S.; Otsuki, A. Seven new microcystins possessing two L-glutamic acid units, isolated from Anabaena sp. strain 186. Chem. Res. Toxicol. 1998, 11, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Foss, A.J.; Aubel, M.T. Using the MMPB technique to confirm microcystin concentrations in water measured by ELISA and HPLC (UV, MS, MS/MS). Toxicon 2015, 104, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Rinehart, K.L.; Namikoshi, M.; Choi, B.W. Structure and biosynthesis of toxins from blue–green algae (cyanobacteria). J. Appl. Phycol. 1994, 6, 159–176. [Google Scholar] [CrossRef]
- Ballot, A.; Sandvik, M.; Rundberget, T.; Botha, C.J.; Miles, C.O. Diversity of cyanobacteria and cyanotoxins in Hartbeespoort Dam, South Africa. Mar. Freshwat. Res. 2014, 65, 175–189. [Google Scholar] [CrossRef] [Green Version]
- Fewer, D.P.; Rouhiainen, L.; Jokela, J.; Wahlsten, M.; Laakso, K.; Wang, H.; Sivonen, K. Recurrent adenylation domain replacement in the microcystin synthetase gene cluster. BMC Evol. Biol. 2007, 7, 183. [Google Scholar] [CrossRef] [Green Version]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef]
- van Donk, E.; van de Bund, W.J. Impact of submerged macrophytes including charophytes on phyto- and zooplankton communities: allelopathy versus other mechanisms. Aquat. Bot. 2002, 72, 261–274. [Google Scholar] [CrossRef]
- Gao, Y.N.; Dong, J.; Fu, Q.Q.; Wang, Y.P.; Chen, C.; Li, J.H.; Li, R.; Zhou, C.J. Allelopathic effects of submerged macrophytes on phytoplankton. Allelopathy J. 2017, 40, 1–22. [Google Scholar]
- Mjelde, M.; Faafeng, B.A. Ceratophyllum demersum hampers phytoplankton development in some small Norwegian lakes over a wide range of phosphorus concentrations and geographical latitude. Freshwat. Biol. 1997, 37, 355–365. [Google Scholar] [CrossRef]
- Blindow, I.; Hargeby, A.; Andersson, G. Seasonal changes of mechanisms maintaining clear water in a shallow lake with abundant Chara vegetation. Aquat. Bot. 2002, 72, 315–334. [Google Scholar] [CrossRef]
- Lombardo, P. Applicability of littoral food-web biomanipulation for lake management purposes: snails, macrophytes, and water transparency in northeast Ohio shallow lakes. Lake Reserv. Manage. 2005, 21, 186–202. [Google Scholar] [CrossRef]
- Utermöhl, H. Zur Vervollkommnung der quantitativen Phytoplankton-Methodik. Mitt. Int. Ver. Theor. Angew. Limnol. 1958, 9, 1–38. [Google Scholar] [CrossRef]
- Kotai, J. Instructions for Preparation of Modified Nutrient Solution Z8 for Algae.—Publication B-11/69; Norwegian Institute for Water Research: Oslo, Norway, 1972. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 1. Chroococcales.—19/1. In Süsswasserflora von Mitteleuropa; Ettl, H., Gärtner, G., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer: Jena-Stuttgart-Lübeck-Ulm, Germany, 1999; p. 548. [Google Scholar]
- Komárek, J. Cyanoprokaryota. 3. Heterocytous genera. In Süsswasserflora von Mitteleuropa/Freshwater flora of Central Europe; Büdel, B., Krienitz, L., Gärtner, G., Schagerl, M., Eds.; Springer Spektrum: Berlin-Heidelberg, Germany, 2013; p. 1130. [Google Scholar]
- Ballot, A.; Cerasino, L.; Hostyeva, V.; Cirés, S. Variability in the sxt gene clusters of PSP toxin producing Aphanizomenon gracile strains from Norway, Spain, Germany and North America. PLoS One 2016, 11, e0167552. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, W.J.; Garthwaite, I.; Miles, C.O.; Ross, K.M.; Aggen, J.B.; Chamberlin, A.R.; Towers, N.R.; Dietrich, D.R. Congener-independent immunoassay for microcystins and nodularins. Environ. Sci. Technol. 2001, 35, 4849–4856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miles, C.O.; Sandvik, M.; Nonga, H.E.; Rundberget, T.; Wilkins, A.L.; Rise, F.; Ballot, A. Thiol derivatization for LC-MS identification of microcystins in complex matrices. Environ. Sci. Technol. 2012, 46, 8937–8944. [Google Scholar] [CrossRef] [Green Version]
- Miles, C.O.; Stirling, D. Toxin mass list, version 15b. Available online: https://www.researchgate.net/publication/324039408_Toxinmasslist_COM_v15b (accessed on 11 October 2018).
- Cerasino, L.; Meriluoto, J.; Bláha, L.; Carmeli, S.; Kaloudis, T.; Mazur-Marzec, A. SOP 6. Extraction of cyanotoxins from cyanobacterial biomass. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; Wiley: Chichester, UK, 2017; pp. 350–353. [Google Scholar]
- Cerasino, L.; Capelli, C.; Salmaso, N. A comparative study of the metabolic profiles of common nuisance cyanobacteria in southern perialpine lakes. Adv. Oceanogr. Limnol. 2017, 8, 22–32. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Point | MK1 | MK 1 | MK 2 | MK 2 | MK 3 | MK 3 | MK 4 | MK 4 | MK 5 | MK 5 |
|---|---|---|---|---|---|---|---|---|---|---|
| Sampling Date | Mar | Nov | Mar | Nov | Mar | Nov | Mar | Nov | Mar | Nov |
| Phytoplankton Group | ||||||||||
| Bacillariophyceae | 0.525 | 0.149 | 1.316 | 0.101 | 0.035 | 0.301 | 0.083 | 0.064 | 0.088 | 0.059 |
| Chlorophyceae | 0.143 | 0.055 | 0.182 | 0.146 | 0.102 | 0.045 | 0.101 | 0.102 | 0.107 | 0.165 |
| Chrysophyceae | 0.011 | 0 | 0.002 | 0 | 0.045 | 0.021 | 0.006 | 0.007 | 0 | 0.004 |
| Conjugatophyceae | 0.099 | 0.006 | 0.141 | 0 | 0.013 | 0 | 0.008 | 0.017 | 0.045 | 0 |
| Cryptophyceae | 0.023 | 0.132 | 0.007 | 0.170 | 0.155 | 0.299 | 0.127 | 0.275 | 0.106 | 0.138 |
| Cyanobacteria | 1.063 | 1.965 | 1.640 | 1.050 | 0.606 | 0.078 | 1.064 | 1.990 | 0.751 | 2.397 |
| Dinophyceae | 0.186 | 0.005 | 0.222 | 0 | 0.285 | 0 | 0.084 | 0.006 | 0.078 | 0 |
| Euglenophyceae | 0.255 | 0.029 | 0.159 | 0.005 | 0.014 | 0.050 | 0.009 | 0.002 | 0.053 | 0.021 |
| Eustigmatophyceae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 |
| Klebsormidiophyceae | 0 | 0 | 0 | 0 | 0 | 0.023 | 0 | 0 | 0.002 | 0 |
| Prymnesiophyceae | 0 | 0.002 | 0.000 | 0.002 | 0.003 | 0.004 | 0.002 | 0.003 | 0.008 | 0.007 |
| Trebouxiophyceae | 0 | 0 | 0.007 | 0 | 0 | 0.005 | 0.005 | 0 | 0 | 0.001 |
| Xanthophyceae | 0 | 0 | 0.008 | 0 | 0.005 | 0 | 0.003 | 0.001 | 0 | 0 |
| Total | 2.306 | 2.344 | 3.684 | 1.474 | 1.264 | 0.827 | 1.493 | 2.467 | 1.238 | 2.790 |
| Species | Strain | Accession nr. 16S rRNA Gene |
|---|---|---|
| Raphidiopsis | ||
| R. raciborskii | AB2017/05 | LR590626 |
| R. raciborskii | AB2017/09 | LR590627 |
| R. raciborskii | AB2017/12 | LR590628 |
| R. raciborskii | AB2017/13 | LR590629 |
| R. raciborskii | AB2017/16 | LR746263 |
| Microcystis | ||
| Microcystis | AB2017/14 | LR590630 |
| Microcystis | AB2017/15 | LR590631 |
| Strain | ELISA | LC-MS/MS | |||
|---|---|---|---|---|---|
| CYNs | CYN | deoxyCYN | CYN (%) | deoxyCYN (%) | |
| AB2017/09 | 2.18 | - | - | 38 | 62 |
| AB2017/05 | n.d. | n.d. | n.d. | n.d. | n.d. |
| AB2017/16 | 1.84 | 1.65 | 1.25 | 57 | 43 |
| AB2017/12 | n.d. | n.d. | n.d. | n.d. | n.d. |
| AB2017/13 | 4.31 | 2.46 | 7.29 | 25 | 75 |
| m/z | Compound Name | Confidence | tR (min) | Concentrationb | ||||
|---|---|---|---|---|---|---|---|---|
| AB2017/14 | AB2017/15 | |||||||
| μg g−1 | % | μg g−1 | % | |||||
| 1 | 1022.5443 | [D-Asp3]MC-RRc | Confirmed | 4.18 | 2.4 | 0.21 | 1002.4 | 7.21 |
| 2 | 1022.5454 | [Dha7]MC-RRc | Probable | 4.53 | 0.1 | 0.01 | 31.8 | 0.23 |
| 3 | 1036.5597 | MC-RRc | Confirmed | 4.55 | 19.4 | 1.74 | 1458.0 | 10.49 |
| 4 | 995.4858 | [D-Asp3]MC-ERc | Probable | 6.14 | ND | - | 119.6 | 0.86 |
| 5 | 1114.5657 | MC-LR–Cysd | Probable | 6.16 | 0.1 | 0.01 | ND | - |
| 6 | 1045.5378 | MC-(H2)YRc | Probable | 6.18 | 32.4 | 2.90 | ND | - |
| 7 | 1031.5224 | [D-Asp3]MC-(H2)YRc | Probable | 6.20 | 32.9 | 2.95 | ND | - |
| 8 | 1033.5381 | [D-Asp3]MC-(H4)YRc | Probable | 6.27 | 0.6 | 0.05 | 148.1 | 1.07 |
| 9 | 1009.5015 | MC-ERc | Probable | 6.55 | 0.6 | 0.05 | 37.8 | 0.27 |
| 10 | 1029.5072 | [D-Asp3]MC-YRc | Probable | 6.59 | ND | - | 144.1 | 1.04 |
| 11 | 1047.5540 | MC-(H4)YRc | Probable | 6.64 | 6.4 | 0.57 | 310.8 | 2.24 |
| 12 | 1011.5530 | [Mser7]MC-LR | Probable | 6.80 | 0.4 | 0.04 | 13.3 | 0.10 |
| 13 | 979.5273 | [D-Asp3]MC-LRc | Confirmed | 6.89 | 40.8 | 3.65 | 1870.3 | 13.45 |
| 14 | 1043.5224 | MC-YRc | Confirmed | 6.99 | 9.1 | 0.81 | 317.4 | 2.28 |
| 15 | 1011.4999 | MC-MRc,d,e | Probable | 6.99 | 3.6 | 0.32 | ND | - |
| 16 | 1073.5329 | MC-Y(OMe)Rc | Probable | 7.07 | 7.2 | 0.64 | ND | - |
| 17 | 993.5435 | MC-LRc | Confirmed | 7.12 | 183.0 | 16.38 | 3332.5 | 23.97 |
| 18 | 979.5289 | [Dha7]MC-LRc | Confirmed | 7.13 | 0.8 | 0.07 | 37.4 | 0.27 |
| 19 | 1080.5170 | Unidentified MCc | Unidentified | 7.15 | 59.0 | 5.28 | ND | - |
| 20 | 1070.5332 | MC-KynRc | Probable | 7.35 | 4.5 | 0.40 | ND | - |
| 21 | 1007.5582 | MC-HilRc | Confirmed | 7.38 | 41.8 | 3.74 | 17.9 | 0.13 |
| 22 | 1080.5169 | Unidentifiedc | Unidentified | 7.38 | 59.6 | 5.34 | ND | - |
| 23 | 1082.5311 | MC-OiaRc | Tentative | 7.40 | 102.4 | 9.17 | ND | - |
| 24 | 995.4857 | [D-Asp3]MC-REc | Probable | 7.43 | ND | - | 110.1 | 0.79 |
| 25 | 1027.5269 | MC-FRc | Confirmed | 7.48 | 255.2 | 22.85 | ND | - |
| 26 | 1066.5380 | MC-WRc | Confirmed | 7.64 | 0.7 | 0.06 | ND | - |
| 27 | 1009.5015 | MC-REc | Probable | 7.79 | 3.8 | 0.34 | 1281.0 | 9.21 |
| 28 | 951.4955 | MC-RAc | Probable | 8.24 | 13 | 1.16 | ND | - |
| 29 | 965.5116 | MC-RAbuc | Probable | 8.83 | 7.1 | 0.64 | ND | - |
| 30 | 946.4577 | MC-(H2)YGc | Probable | 9.06 | 5.4 | 0.48 | ND | - |
| 31 | 1011.4966 | MC-RMc,d,e | Probable | 9.23 | 0.2 | 0.02 | ND | - |
| 32 | 960.4738 | MC-(H2)YAc | Probable | 9.34 | 10.1 | 0.90 | ND | - |
| 33 | 953.4783 | Unidentified MCc | Unidentified | 9.50 | 12 | 1.07 | ND | - |
| 34 | 967.4942 | Unidentified MCc | Unidentified | 9.78 | 2.3 | 0.21 | ND | - |
| 35 | 984.4735 | Unidentified MCc | Unidentified | 9.96 | 3.7 | 0.33 | ND | - |
| 36 | 968.4272 | [D-Asp3]MC-EEc | Probable | 11.43 | ND | - | 403.3 | 2.90 |
| 37 | 982.4432 | MC-EEc | Probable | 12.58 | 0.9 | 0.08 | 1533.5 | 11.03 |
| 38 | 952.4690 | [D-Asp3]MC-LEc | Probable | 12.82 | 1.2 | 0.11 | 358.8 | 2.58 |
| 39 | 894.4631 | [D-Asp3]MC-LAc | Probable | 13.32 | 6.5 | 0.58 | ND | - |
| 40 | 966.4849 | MC-LEc | Probable | 13.69 | 12.9 | 1.15 | 1376.1 | 9.90 |
| 41 | 908.4788 | MC-LAc | Confirmed | 14.87 | 44.8 | 4.01 | ND | - |
| 42 | 922.4942 | MC-HilAc | Probable | 15.29 | 5.9 | 0.53 | ND | - |
| 43 | 981.4737 | MC-WAc | Probable | 15.88 | 4.4 | 0.39 | ND | - |
| 44 | 942.4629 | MC-FAc | Probable | 15.95 | 42.3 | 3.79 | ND | - |
| 45 | 922.4943 | MC-LAbuc | Probable | 16.09 | 16.6 | 1.49 | ND | - |
| 46 | 936.5110 | MC-HilAbuc | Probable | 16.55 | 2.2 | 0.20 | ND | - |
| 47 | 995.4898 | MC-WAbuc | Probable | 17.20 | 13.4 | 1.20 | ND | - |
| 48 | 956.4784 | MC-FAbuc | Probable | 17.39 | 26.4 | 2.36 | ND | - |
| 49 | 936.5106 | MC-LVc | Probable | 17.62 | 3.6 | 0.32 | ND | - |
| 50 | 950.5254 | MC-LLc | Probable | 17.93 | 3.9 | 0.35 | ND | - |
| 51 | 970.4944 | MC-FVc | Probable | 18.40 | 0.7 | 0.06 | ND | - |
| 52 | 950.5262 | iso-MC-LLc | Tentative | 18.49 | 1.2 | 0.11 | ND | - |
| 53 | 1009.5041 | MC-WVc | Probable | 18.54 | 2.9 | 0.26 | ND | - |
| 54 | 1023.5210 | MC-WLc | Probable | 18.74 | 2.8 | 0.25 | ND | - |
| 55 | 970.4942 | iso-MC-FVc | Tentative | 18.79 | 2.1 | 0.19 | ND | - |
| 56 | 1023.5216 | iso-MC-WLc | Tentative | 19.16 | 1.3 | 0.12 | ND | - |
| Sampling Point | Water Depth (m) | Depth of Integrated Sample (m) | Geographical Position |
|---|---|---|---|
| MK1 | 3.3 | 0–1 | N 20° 52’ 59.196, E 95° 51’ 12.204 |
| MK2 | 2 | 0–1 | N 20° 53’ 21.48, E 95° 51’ 2.124 |
| MK3 | 2.5 | 0–1 | N 20° 54’ 16.38, E 95° 50’ 40.092 |
| MK4 | 4.4 | 0–2 | N 20° 52’ 21.468, E 95° 51’ 12.528 |
| MK5 | 7.1 | 0–3 | N 20° 51’ 58.752, E 95° 51’ 18.936 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballot, A.; Swe, T.; Mjelde, M.; Cerasino, L.; Hostyeva, V.; Miles, C.O. Cylindrospermopsin- and Deoxycylindrospermopsin-Producing Raphidiopsis raciborskii and Microcystin-Producing Microcystis spp. in Meiktila Lake, Myanmar. Toxins 2020, 12, 232. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12040232
Ballot A, Swe T, Mjelde M, Cerasino L, Hostyeva V, Miles CO. Cylindrospermopsin- and Deoxycylindrospermopsin-Producing Raphidiopsis raciborskii and Microcystin-Producing Microcystis spp. in Meiktila Lake, Myanmar. Toxins. 2020; 12(4):232. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12040232
Chicago/Turabian StyleBallot, Andreas, Thida Swe, Marit Mjelde, Leonardo Cerasino, Vladyslava Hostyeva, and Christopher O. Miles. 2020. "Cylindrospermopsin- and Deoxycylindrospermopsin-Producing Raphidiopsis raciborskii and Microcystin-Producing Microcystis spp. in Meiktila Lake, Myanmar" Toxins 12, no. 4: 232. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12040232