Persistent Exposure to Environmental Levels of Microcystin-LR Disturbs Cortisol Production via Hypothalamic-Pituitary-Interrenal (HPI) Axis and Subsequently Liver Glucose Metabolism in Adult Male Zebrafish (Danio rerio)

 , and
, and
Abstract
:1. Introduction
2. Results
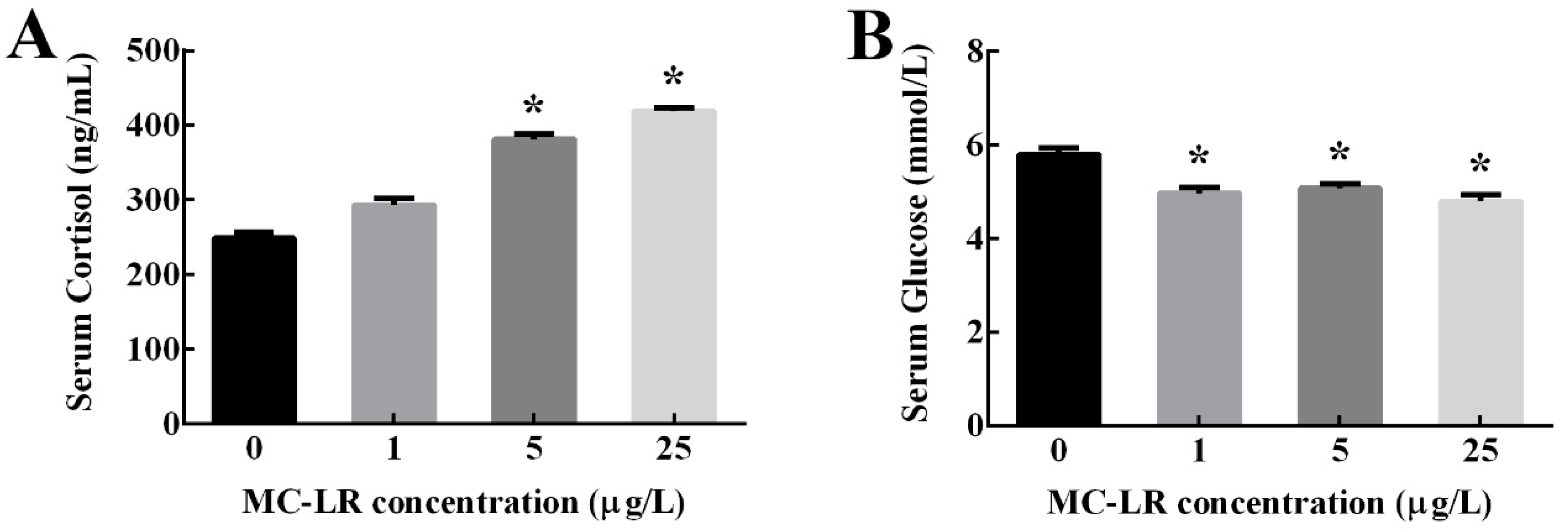
2.1. Serum Cortisol and Glucose Levels
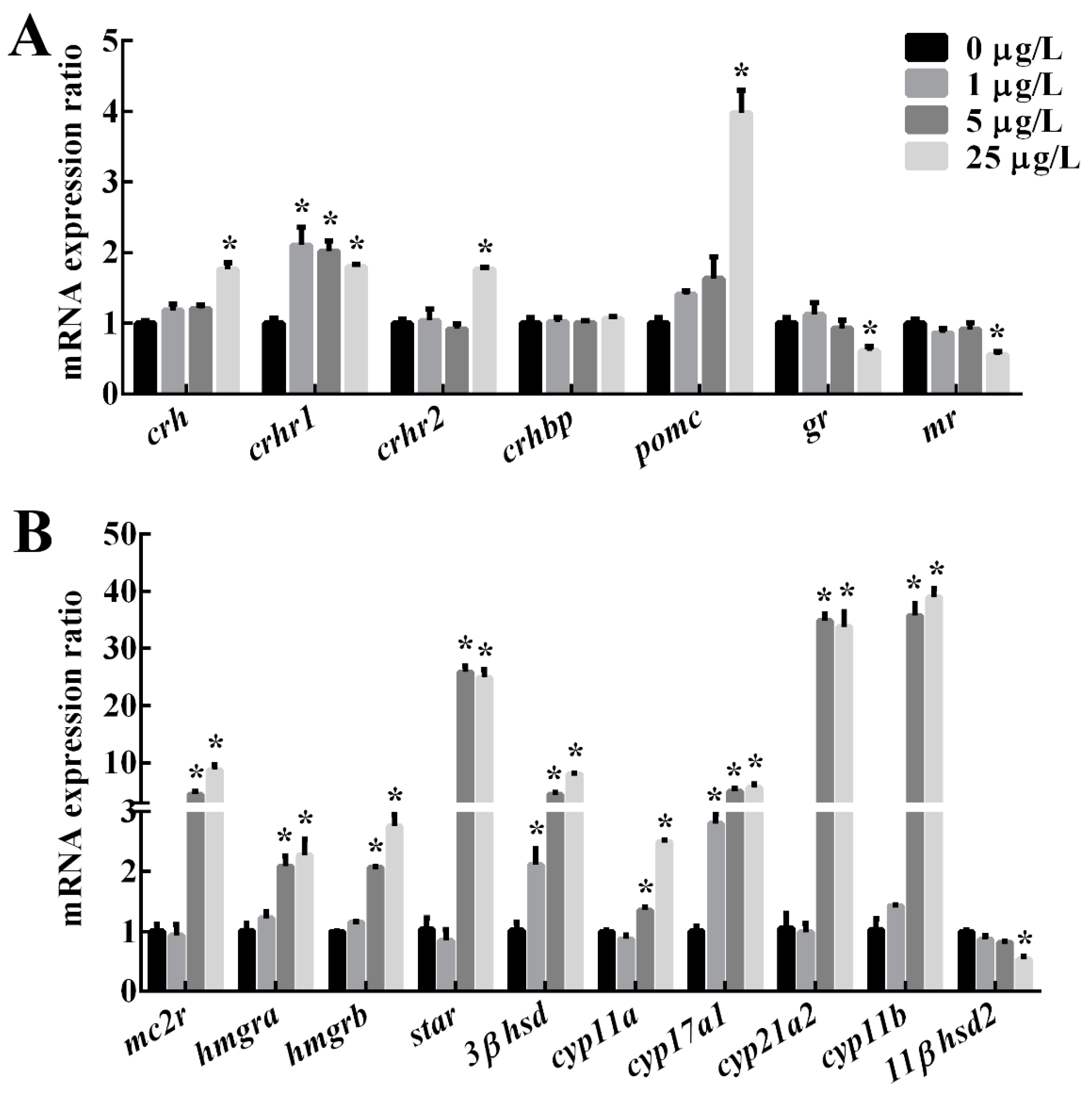
2.2. The Expression of Genes Related to HPI Axis
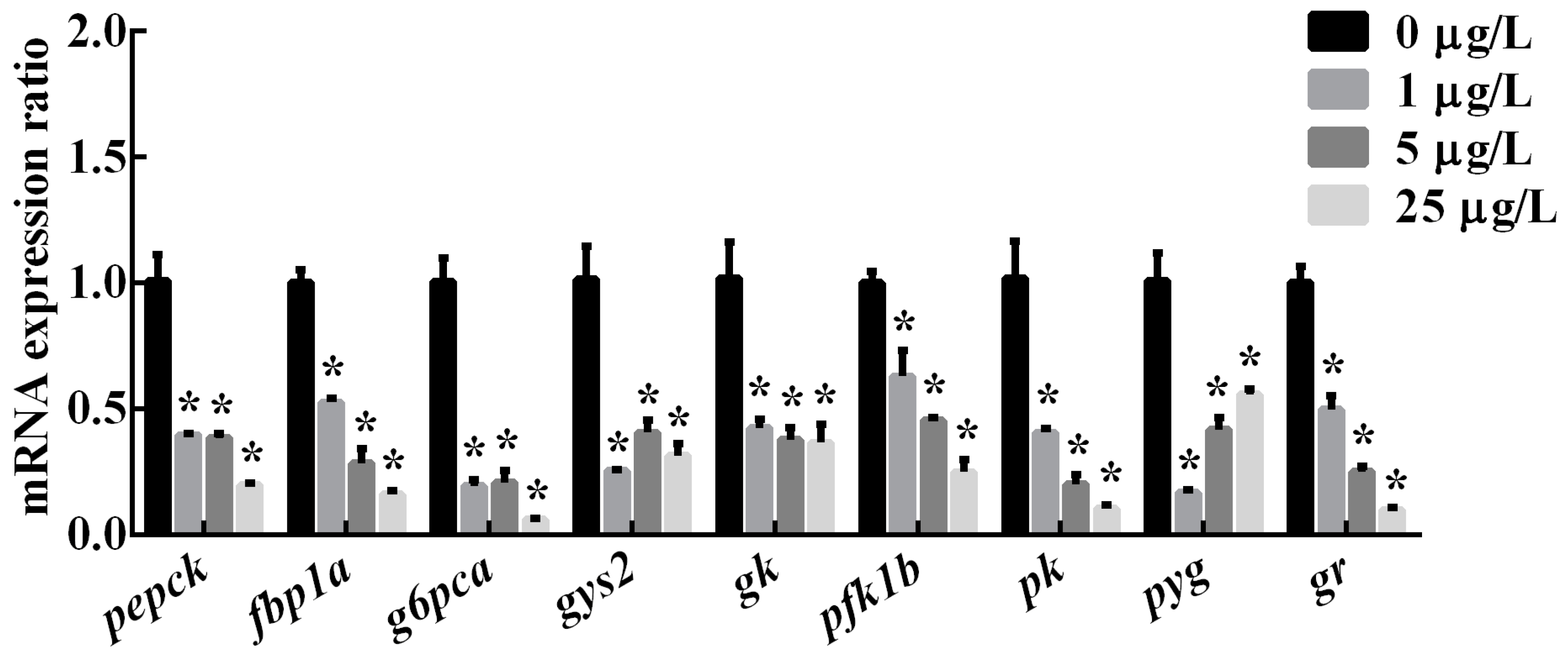
2.3. The Expression of Genes Related to Hepatic Glucose Metabolic Enzymes
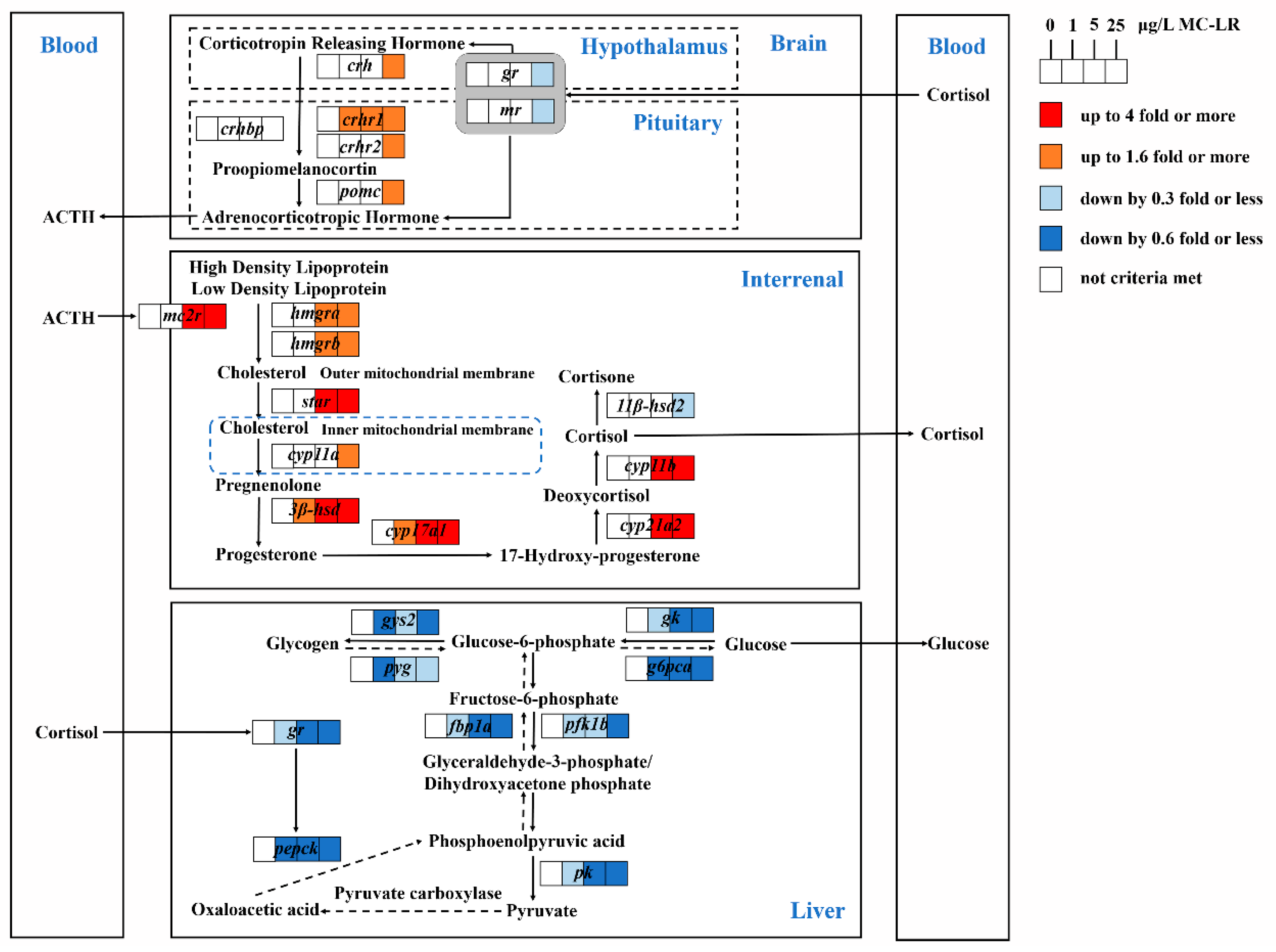
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Animal Maintenance and Experimental Design
5.3. Serum Cortisol and Glucose Analysis
5.4. Quantitative Real-Time PCR
5.5. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, L.; Xie, P.; Lei, H.; Zhang, X. Renal accumulation and effects of intraperitoneal injection of extracted microcystins in omnivorous crucian carp (Carassius auratus). Toxicon 2013, 70, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Su, Y.; Lin, W.; Guo, H.; Xie, P.; Chen, J.; Gu, Z.; Li, L. Microcystin-LR retards gonadal maturation through disrupting the growth hormone/insulin-like growth factors system in zebrafish. Ecotoxicol. Environ. Saf. 2017, 139, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Rezaitabar, S.; Sari, A.E.; Bahramifar, N.; Ramezanpour, Z. Transfer, tissue distribution and bioaccumulation of microcystin-LR in the phytoplanktivorous and carnivorous fish in Anzali wetland, with potential health risks to humans. Sci. Totol. Environ. 2017, 575, 1130–1138. [Google Scholar] [CrossRef]
- Bouaïcha, N.; Miles, C.O.; Beach, D.G.; Labidi, Z.; Djabri, A.; Benayache, N.Y.; Nguyen-Quang, T. Structural diversity, characterization and toxicology of microcystins. Toxins 2019, 11, 714. [Google Scholar] [CrossRef] [Green Version]
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef]
- World Health Organization. Health Organization. Health criteria and other supporting information: Addendum. In Guidelines for Drinking-Water Quality; No. WHO/EOS/98.1; World Health Organization: Geneva, Switzerland, 1998; Volume 2. [Google Scholar]
- Chen, J.; Xie, P.; Li, L.; Xu, J. First identification of the hepatotoxic microcystins in the serum of a chronically exposed human population together with indication of hepatocellular damage. Toxicol. Sci. 2009, 108, 81–89. [Google Scholar] [CrossRef]
- Wang, Q.; Niu, Y.; Xie, P.; Chen, J.; Ma, Z.; Tao, M.; Qi, M.; Wu, L.; Guo, L. Factors affecting temporal and spatial variations of microcystins in Gonghu Bay of Lake Taihu, with potential risk of microcystin contamination to human health. Sci. World J. 2010, 10, 1795–1809. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Xie, P.; Chen, J. In vivo studies on toxin accumulation in liver and ultrastructural changes of hepatocytes of the phytoplanktivorous bighead carp ip-injected with extracted microcystins. Toxicon 2005, 46, 533–545. [Google Scholar] [CrossRef]
- Li, D.; Xie, P.; Zhang, X.; Zhao, Y. Intraperitoneal injection of extracted microcystins results in hypovolemia and hypotension in crucian carp (Carassius auratus). Toxicon 2009, 53, 638–644. [Google Scholar] [CrossRef]
- Lin, W.; Hou, J.; Guo, H.; Qiu, Y.; Li, L.; Li, D.; Tang, R. Dualistic immunomodulation of sub-chronic microcystin-LR exposure on the innate-immune defense system in male zebrafish. Chemosphere 2017, 183, 315–322. [Google Scholar] [CrossRef]
- Li, G.; Cai, F.; Yan, W.; Li, C.; Wang, J. A proteomic analysis of MCLR-induced neurotoxicity: Implications for Alzheimer’s disease. Toxicol. Sci. 2012, 127, 485–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Xie, L.; Yan, Y. Microcystin-LR impairs zebrafish reproduction by affecting oogenesis and endocrine system. Chemosphere 2015, 120, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Li, L.; Wu, N.; Su, Y.; Lin, W.; Li, G.; Gu, Z. Reproduction impairment and endocrine disruption in female zebrafish after long-term exposure to MC-LR: A life cycle assessment. Environ. Pollut. 2016, 208, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Chen, C.; Chen, L.; Wang, L.; Li, J.; Chen, Y.; Jin, J.; Kawan, A.; Zhang, X. Sex-dependent effects of microcystin-LR on hypothalamic-pituitary-gonad axis and gametogenesis of adult zebrafish. Sci. Rep. 2016, 6, 22819. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Li, L.; Hou, J.; Wu, N.; Lin, W.; Li, G. Life-cycle exposure to microcystin-LR interferes with the reproductive endocrine system of male zebrafish. Aquat. Toxicol. 2016, 175, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Su, Y.; Lin, W.; Guo, H.; Li, L.; Anderson, D.M.; Li, D.; Tang, R.; Chi, W.; Zhang, X. Estrogenic potency of MC-LR is induced via stimulating steroidogenesis: In vitro and in vivo evidence. Environ. Pollut. 2018, 240, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, D.; Hu, Q.; Tang, R.; Li, L. Effects of exposure to microcystin-LR at environmentally relevant concentrations on the metabolism of thyroid hormones in adult zebrafish (Danio rerio). Toxicon 2016, 124, 15–25. [Google Scholar] [CrossRef]
- Cheng, H.; Yan, W.; Wu, Q.; Liu, C.; Gong, X.; Hung, T.C.; Li, G. Parental exposure to microcystin-LR induced thyroid endocrine disruption in zebrafish offspring, a transgenerational toxicity. Environ. Pollut. 2017, 230, 981–988. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, X.; Deng, J.; Hecker, M.; Al-Khedhairy, A.; Giesy, J.P.; Zhou, B. Effects of prochloraz or propylthiouracil on the cross-talk between the HPG, HPA, and HPT axes in zebrafish. Environ. Sci. Technol. 2011, 45, 769–775. [Google Scholar] [CrossRef]
- Bury, N.R.; Eddy, F.B.; Codd, G.A. Stress responses of brown trout, Salmo trutta L., to the cyanobacterium, Microcystis aeruginosa. Environ. Toxicol. Water Qual. 1996, 11, 187–193. [Google Scholar] [CrossRef]
- Li, D.; Xie, P.; Zhang, X. Changes in plasma thyroid hormones and cortisol levels in crucian carp (Carassius auratus) exposed to the extracted microcystins. Chemosphere 2008, 74, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Mommsen, T.P.; Vijayan, M.M.; Moon, T.W. Cortisol in teleosts: Dynamics, mechanisms of action, and metabolic regulation. Rev. Fish Biol. Fish. 1999, 9, 211–268. [Google Scholar] [CrossRef]
- Trenzado, C.E.; Carrick, T.R.; Pottinger, T.G. Divergence of endocrine and metabolic responses to stress in two rainbow trout lines selected for differing cortisol responsiveness to stress. Gen. Comp. Endocrinol. 2003, 133, 332–340. [Google Scholar] [CrossRef] [Green Version]
- López-Patiño, M.A.; Hernández-Pérez, J.; Gesto, M.; Librán-Pérez, M.; Míguez, J.M.; Soengas, J.L. Short-term time course of liver metabolic response to acute handling stress in rainbow trout, Oncorhynchus mykiss. Comp. Biochem. Phys. A 2014, 168, 40–49. [Google Scholar] [CrossRef]
- Ernst, B.; Hoeger, S.J.; O’Brien, E.; Dietrich, D.R. Oral toxicity of the microcystin-containing cyanobacterium Planktothrix rubescens in European whitefish (Coregonus lavaretus). Aqua. Toxicol. 2006, 79, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Woźny, M.; Lewczuk, B.; Ziółkowska, N.; Gomułka, P.; Dobosz, S.; Łakomiak, A.; Florczyk, M.; Brzuzan, P. Intraperitoneal exposure of whitefish to microcystin-LR induces rapid liver injury followed by regeneration and resilience to subsequent exposures. Toxicol. Appl. Pharm. 2016, 313, 68–87. [Google Scholar] [CrossRef]
- Paulino, M.G.; Rossi, P.A.; Venturini, F.P.; Tavares, D.; da Silva Souza, N.E.; Sakuragui, M.M.; Moraes, G.; Terezan, A.P.; Fernandes, J.B.; Giani, A.; et al. Hepatotoxicity and metabolic effects of cellular extract of cyanobacterium Radiocystis fernandoi containing microcystins RR and YR on neotropical fish (Hoplias malabaricus). Chemosphere 2017, 175, 431–439. [Google Scholar] [CrossRef]
- Mezhoud, K.; Praseuth, D.; Puiseux-Dao, S.; Francois, J.C. Global quantitative analysis of protein expression and phosphorylation status in the liver of the medaka fish (Oryzias latipes) exposed to microcystin-LR, Balneation study. Aquat. Toxicol. 2008, 86, 166–175. [Google Scholar] [CrossRef]
- Zhao, S.; Li, G.; Chen, J. A proteomic analysis of prenatal transfer of microcystin-LR induced neurotoxicity in rat offspring. J. Proteom. 2015, 114, 197–213. [Google Scholar] [CrossRef]
- Moon, T.W. Glucose intolerance in teleost fish: Fact or fiction? Comp. Biochem. Phys. B 2001, 129, 243–249. [Google Scholar] [CrossRef]
- Alsop, D.; Vijayan, M.M. Molecular programming of the corticosteroid stress axis during zebrafish development. Comp. Biochem. Phys. A 2009, 153, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Alsop, D.; Vijayan, M. The zebrafish stress axis: Molecular fallout from the teleost-specific genome duplication event. Gen. Comp. Endocrinol. 2009, 161, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yu, H.; Zhang, X. Zebrafish embryos/larvae for rapid determination of effects on hypothalamic-pituitary-thyroid (HPT) and hypothalamic-pituitary-interrenal (HPI) axis: mRNA expression. Chemosphere 2013, 93, 2327–2332. [Google Scholar] [CrossRef] [PubMed]
- Alderman, S.L.; Bernier, N.J. Ontogeny of the corticotropin-releasing factor system in zebrafish. Gen. Comp. Endocrinol. 2009, 164, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Yan, W.; Liu, C.S.; Li, L.; Yu, L.Q.; Zhao, S.J.; Li, G.Y. Microcystin-LR exposure induces developmental neurotoxicity in zebrafish embryo. Environ. Pollut. 2016, 213, 793–800. [Google Scholar] [CrossRef]
- Yan, W.; Zhou, Y.; Yang, J.; Li, S.; Hu, D.; Wang, J.; Chen, J.; Li, G. Waterborne exposure to microcystin-LR alters thyroid hormone levels and gene transcription in the hypothalamic–pituitary–thyroid axis in zebrafish larvae. Chemosphere 2012, 87, 1301–1307. [Google Scholar] [CrossRef]
- Liu, Z.; Tang, R.; Li, D.; Hu, Q.; Wang, Y. Subacute microcystin-LR exposure alters the metabolism of thyroid hormones in juvenile zebrafish (Danio Rerio). Toxins 2015, 7, 337–352. [Google Scholar] [CrossRef] [Green Version]
- Fuzzen, M.L.M.; Van Der Kraak, G.; Bernier, N.J. Stirring up new ideas about the regulation of the hypothalamic-pituitary-interrenal axis in zebrafish (Danio rerio). Zebrafish 2010, 7, 349–358. [Google Scholar] [CrossRef]
- Zhang, X.; Zhong, Y.; Tian, H.; Wang, W.; Ru, S. Impairment of the cortisol stress response mediated by the hypothalamus–pituitary–interrenal (HPI) axis in zebrafish (Danio rerio) exposed to monocrotophos pesticide. Comp. Biochem. Phys. C 2015, 176, 10–16. [Google Scholar] [CrossRef]
- Arukwe, A. Modulation of barin steroidogenesis by affecting transcriptional changes of steroidogenic acute regulatory (StAR) protein and cholesterol side chain cleavage (P450scc) in juvenile Atlantic salmon (Salmo salar) is a novel aspect of nonylphenol toxicity. Environ. Sci. Technol. 2005, 39, 9791–9798. [Google Scholar] [CrossRef]
- Nichols, J.W.; Breen, M.; Denver, R.J.; DiStefano, J.J.; Edwards, J.S.; Hoke, R.A.; Volz, D.C.; Zhang, X. Predicting chemical impacts on vertebrate endocrine systems. Environ. Toxicol. Chem. 2011, 30, 39–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bury, N.R.; Sturm, A. Evolution of the corticosteroid receptor signaling pathway in fish. Gen. Comp. Endocrinol. 2007, 153, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Schaaf, M.J.M.; Chatzopoulou, A.; Spaink, H.P. The zebrafish as a model system for glucocorticoid receptor research. Comp. Biochem. Phys. A 2009, 153, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, Y.; Giesy, J.P.; Chen, F.; Shi, T.; Chen, J.; Xie, P. Microcystin-LR affects the hypothalamic-pituitary-inter-renal (HPI) axis in early life stages (embryos and larvae) of zebrafish. Environ. Pollut. 2018, 241, 540–548. [Google Scholar] [CrossRef]
- Griffiths, B.; Schoonheim, P.J.; Ziv, L.; Voelker, L.; Baier, H.; Gahtan, E. A zebrafish model of glucocorticoid resistance shows serotonergic modulation of the stress response. Front. Behav. Neurosci. 2012, 6, 68. [Google Scholar] [CrossRef] [Green Version]
- Ziv, L.; Muto, A.; Schoonheim, P.J.; Meijsing, S.H.; Strasser, D.; Ingraham, H.A.; Schaaf, M.J.M.; Yamamoto, K.R.; Baier, H. An affective disorder in zebrafish with mutation of the glucocorticoid receptor. Mol. Psychiatry 2013, 18, 681–691. [Google Scholar] [CrossRef]
- Romero, L.M.; Dickens, M.J.; Cyr, N.E. The reactive scope model—A new model integrating homeostasis, allostasis, and stress. Horm. Behav. 2009, 55, 375–389. [Google Scholar] [CrossRef]
- Zhao, F.; Jiang, G.; Wei, P.; Wang, H.; Ru, S. Bisphenol S exposure impairs glucose homeostasis in male zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2018, 147, 794–802. [Google Scholar] [CrossRef]
- Chen, L.; Hu, Y.; He, J.; Chen, J.; Giesy, J.P.; Xie, P. Responses of the proteome and metabolome in livers of zebrafish exposed chronically to environmentally relevant concentrations of microcystin-LR. Environ. Sci. Technol. 2016, 51, 596–607. [Google Scholar] [CrossRef]
- Malbrouck, C.; Trausch, G.; Devos, P.; Kestemont, P. Effect of microcystin-LR on protein phosphatase activity and glycogen content in isolated hepatocytes of fed and fasted juvenile goldfish Carassius auratus L. Toxicon 2004, 44, 927–932. [Google Scholar] [CrossRef]
- Hou, J.; Li, L.; Xue, T.; Long, M.; Su, Y.; Wu, N. Hepatic positive and negative antioxidant responses in zebrafish after intraperitoneal administration of toxic microcystin-LR. Chemosphere 2015, 120, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Hou, J.; Guo, H.; Li, L.; Wang, L.; Zhang, D.; Li, D.; Tang, R. The synergistic effects of waterborne microcystin-LR and nitrite on hepatic pathological damage, lipid peroxidation and antioxidant responses of male zebrafish. Environ. Pollut. 2018, 235, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Rance, T.A.; Baker, B.I. The in vitro response of the trout interrenal to various fragments of ACTH. Gen. Comp. Endocrinol. 1981, 45, 497–503. [Google Scholar] [CrossRef]
- Trinder, P. Determination of glucose in blood using glucose oxidase with an alternative oxygen acceptor. Ann. Clin. Biochem. 1969, 6, 24–27. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Gene Name |
|---|---|
| crh | corticotropin releasing hormone |
| crhr1 | corticotropin releasing hormone receptor 1 |
| crhr2 | corticotropin releasing hormone receptor 2 |
| crhbp | corticotropin releasing hormone binding protein |
| pomc | proopiomelanocortin |
| gr | glucocorticoid receptor |
| mr | mineralocorticoid receptor |
| mc2r | melanocortin 2 receptor |
| hmgra | hydroxy-methylglutaryl-CoA reductase a |
| hmgrb | hydroxy-methylglutaryl-CoA reductase b |
| star | steroidogenic acute regulatory protein |
| cyp11a1 | cytochrome P450, family 11, subfamily A, polypeptide 1 |
| 3β-hsd | 3-beta-hydroxysteroid dehydrogenase |
| cyp17a1 | cytochrome P450, family 17, subfamily A, polypeptide 1 |
| cyp21a2 | cytochrome P450, family 21, subfamily A, polypeptide 2 |
| cyp11b | cytochrome P450, family 11, subfamily B |
| 11β-hsd2 | 11-beta-hydroxysteroid dehydrogenase-type 3 |
| pepck | phosphoenolpyruvate carboxykinase |
| fbp1a | fructose-1,6-bisphosphatase 1a |
| g6pca | glucose-6-phosphatase |
| gk | glucokinase |
| pfk1b | phosphofructokinase 1b |
| pk | pyruvate kinase |
| pyg | phosphorylase, glycogen |
| gys2 | glycogen synthase 2 |
| Gene | Sequence of the Primers (5′-3′) | Accession No. | Product Length (bp) | Amplification Efficiency (%) |
|---|---|---|---|---|
| crh | F: TTCGGGAAGTAACCACAAGC | NM_001007379.1 | 161 | 109.37% |
| R: CTGCACTCTATTCGCCTTCC | ||||
| crhr1 | F: TCGGTGTTGAGTCTCATGCC | XM_691254.6 | 186 | 93.89% |
| R: CGCGTGCGGGTAAAATGTAG | ||||
| crhr2 | F: TGAACTGTTGCTCCGAGTCC | NM_001113644.1 | 81 | 101.76% |
| R: GGAGAAACACGTCGCCGTTA | ||||
| crhbp | F: AGAGGTCCATTCTTGGCTGC | NM_001003459.1 | 170 | 98.52% |
| R: GCTCGACACCATTCTGACCA | ||||
| pomc | F: AGGTCGACTATCCGCAAGAA | AY158003 | 232 | 97.13% |
| R: CAACCTCTCCCCCTTAAAGC | ||||
| gr | F: TTCTACGTTGCTGACGATGC | EF567112 | 239 | 95.48% |
| R: CCGGTGTTCTCCTGTTTGAT | ||||
| mr | F: TGCCACTACGGGGTTGTTAC | EF567113 | 181 | 102.77% |
| R: GTGCCCCAAGATTCATCCCA | ||||
| mc2r | F: AAAGGGGTATTCGTGGCCTG | NM_180971 | 150 | 96.54% |
| R: TGCCGGATCAATAACAGCGT | ||||
| hmgra | F: GGAACATGCACTAAGCAGGC | NM_001079977.2 | 143 | 93.24% |
| R: AGAGAAGAAGGGATCGGTTGC | ||||
| hmgrb | F: CCGCTTCTATTGGACGGGAA | NM_001014292.2 | 200 | 90.61% |
| R: AACCAGCCCTTAACCTCTGC | ||||
| star | F: TTTCTGGCTGGGATGTCCAC | NM_131663 | 105 | 104.15% |
| R: ATCTGCACTTGGTCGCATGA | ||||
| cyp11a | F: TCCCGAAACCAGAGCAATAC | NM_152953 | 204 | 97.45% |
| R: GCTCAAACTTGCTCCTGACC | ||||
| 3βhsd | F: CGCACATCGTCTCAGGACAT | AY279108.1 | 122 | 106.82% |
| R: GTAGAGCGCTGCGTTGAAAG | ||||
| cyp17a1 | F: AGACCCACCACAGACCTTTAC | NM_212806.3 | 96 | 92.59% |
| R: TGAGCACAATCGGCCACTTA | ||||
| cyp21a2 | F: AATGGTCTGACTTTGCTGGGA | XM_021474355.1 | 222 | 95.38% |
| R: CAGCCTTTCCACTGTAGTCTCG | ||||
| cyp11b | F: TGTATCCTCTGGGTCGTTCC | DQ650710 | 228 | 99.68% |
| R: GCTCTTCTGTGGACGAAACC | ||||
| 11β-hsd2 | F: ATCAGAATCCATCCAGCCTTAG | NM_212720 | 113 | 92.76% |
| R: ATTAGCATCCACACCAATACATC | ||||
| pepck | F: TGCCTGGATGAAATTTGACA | NM_213192.1 | 106 | 98.67% |
| R: GGCATGAGGGTTGGTTTTTA | ||||
| fbp1a | F: CATCTGTATGGGATTGCTGG | NM_199942 | 173 | 93.12% |
| R: TTACCCCGTCTATCTGGCTC | ||||
| g6pca | F: GGCTGAACCTCGTCCTAAAGT | BC076446 | 203 | 93.11% |
| R: GATTGAAAGCAACGCTGTGAT | ||||
| gk | F: TGAGGATGAAGAGCGAGGC | BC122359 | 178 | 91.9% |
| R: AGAGAAGGTGAATCCCAGCG | ||||
| pfk1b | F: GCATGTGTGTCATTCCTGCC | NM_001328389.1 | 194 | 104.5% |
| R: GCCTGTAGTGGTTGCCAGAT | ||||
| pk | F: AGAAACAGCCAAAGGACA | BC152219 | 253 | 103.16% |
| R: ACGAGGACGATAACGAGA | ||||
| pyg | F: GAAAGGTTTCAACCGGCACC | NM_001008538.1 | 88 | 90.8% |
| R: GTGCGACAGCGCGAAATAAT | ||||
| gys2 | F: GTGCGATGCGACTATCCAGA | NM_001018679.1 | 188 | 106.6% |
| R: TTCACCCCATTCGTCCACAG | ||||
| gapdh | F: CTGGTGACCCGTGCTGCTT | NM_001115114 | 150 | 98.2% |
| R: TTTGCCGCCTTCTGCCTTA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Lin, W.; Zha, Q.; Guo, H.; Zhang, D.; Yang, L.; Li, L.; Li, D.; Tang, R. Persistent Exposure to Environmental Levels of Microcystin-LR Disturbs Cortisol Production via Hypothalamic-Pituitary-Interrenal (HPI) Axis and Subsequently Liver Glucose Metabolism in Adult Male Zebrafish (Danio rerio). Toxins 2020, 12, 282. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12050282
Wang L, Lin W, Zha Q, Guo H, Zhang D, Yang L, Li L, Li D, Tang R. Persistent Exposure to Environmental Levels of Microcystin-LR Disturbs Cortisol Production via Hypothalamic-Pituitary-Interrenal (HPI) Axis and Subsequently Liver Glucose Metabolism in Adult Male Zebrafish (Danio rerio). Toxins. 2020; 12(5):282. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12050282
Chicago/Turabian StyleWang, Lingkai, Wang Lin, Qingji Zha, Honghui Guo, Dandan Zhang, Liping Yang, Li Li, Dapeng Li, and Rong Tang. 2020. "Persistent Exposure to Environmental Levels of Microcystin-LR Disturbs Cortisol Production via Hypothalamic-Pituitary-Interrenal (HPI) Axis and Subsequently Liver Glucose Metabolism in Adult Male Zebrafish (Danio rerio)" Toxins 12, no. 5: 282. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12050282