A Mediterranean Alexandrium taylorii (Dinophyceae) Strain Produces Goniodomin A and Lytic Compounds but Not Paralytic Shellfish Toxins


Abstract
:1. Introduction
2. Results
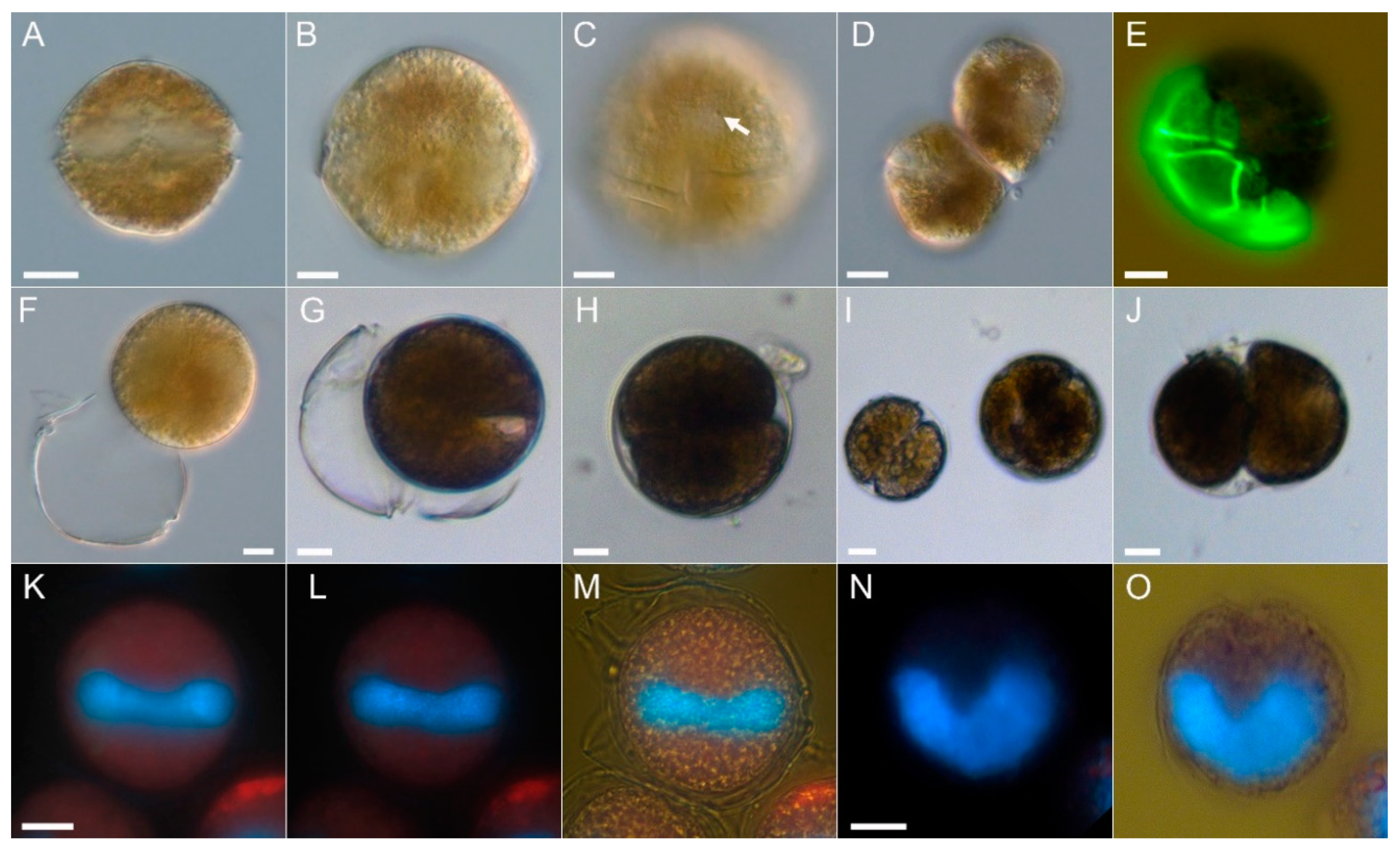
2.1. Species Identification
2.2. Toxin Analysis
2.2.1. PST
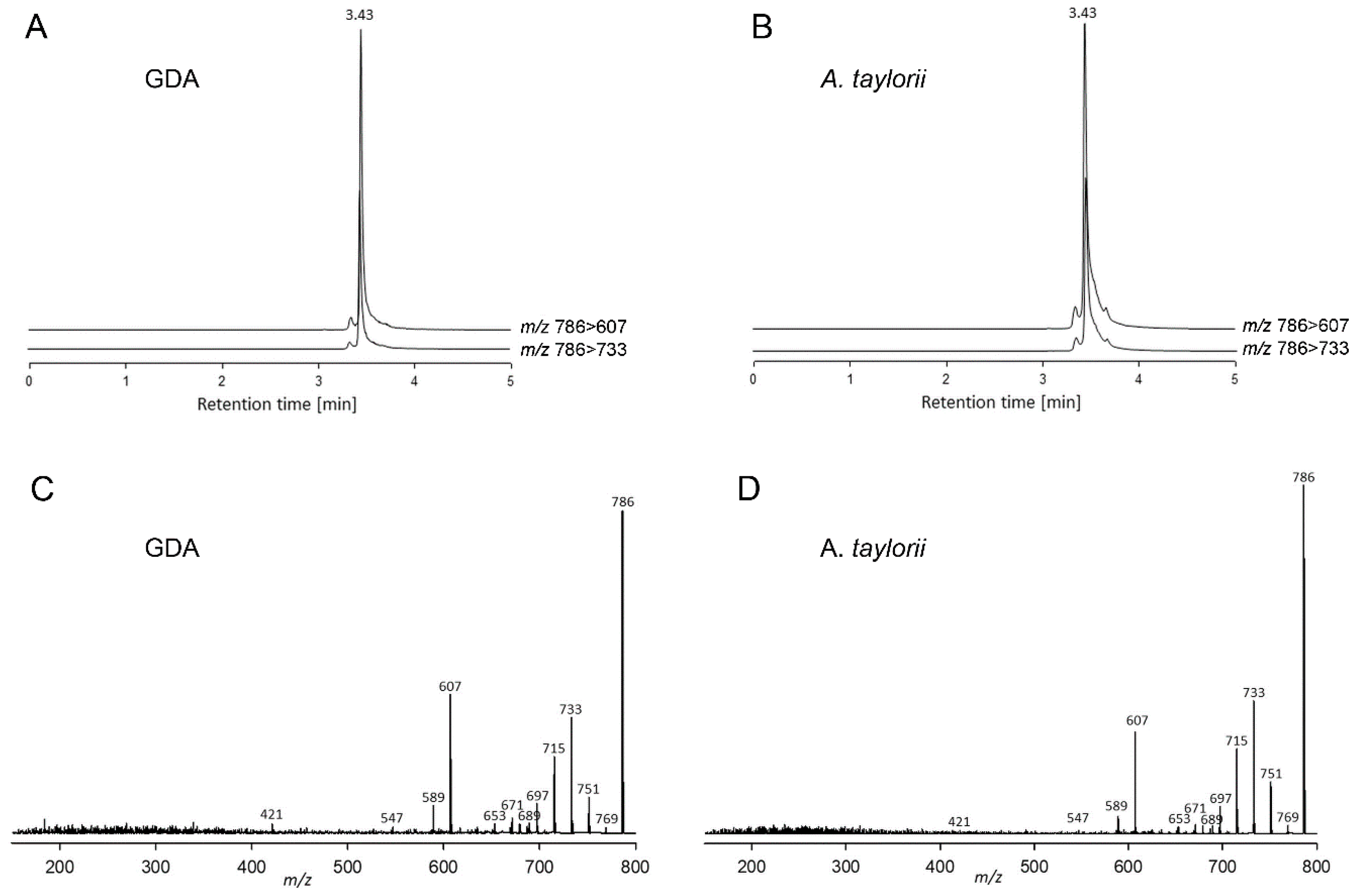
2.2.2. Lipophilic Compounds
2.2.3. Lytic Capacity
3. Discussion
4. Materials and Methods
4.1. Strain Isolation and Harvest
4.2. Microscopy
4.3. DNA Extraction and Sequencing
4.4. Toxin Analysis
4.4.1. Paralytic Shellfish Toxins
4.4.2. Lipophilic Compounds
4.4.3. Lytic Compounds
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Taylor, F.J.R.; Fukuyo, Y. The Neurotoxigenic Dinoflagellate Genus Alexandrium Halim: General Introduction. In Physiological Ecology of Harmful Algal Blooms, 1st ed.; Anderson, D.M., Cembella, A.D., Hallegraeff, G.M., Eds.; Springer: Berlin, Germany, 1998; Volume 41, pp. 3–12. [Google Scholar]
- Cembella, A. Alexandrium. In Harmful Algae Blooms, a Compendium Desk Reference; Shumway, S.E., Burkholder, J.A., Morton, S.L., Eds.; Wiley: Hoboken, NJ, USA, 2018; pp. 563–573. [Google Scholar]
- Anderson, D.M.; Alpermann, T.J.; Cembella, A.D.; Collos, Y.; Masseret, E.; Montresor, M. The globally distributed genus Alexandrium: Multifaceted roles in marine ecosystems and impacts on human health. Harmful Algae 2012, 14, 10–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez, G.; Díaz, P.A.; Godoy, M.; Araya, M.; Ganuza, I.; Pino, R.; Álvarez, F.; Rengel, J.; Hernández, C.; Uribe, E.; et al. Paralytic shellfish toxins in surf clams Mesodesma donacium during a large bloom of Alexandrium catenella dinoflagellates associated to an intense shellfish mass mortality. Toxins 2019, 11, 18. [Google Scholar] [CrossRef] [Green Version]
- Díaz, P.A.; Álvarez, G.; Varela, D.; Pérez-Santos, I.; Díaz, M.; Molinet, C.; Seguel, M.; Aguilera-Belmonte, A.; Guzmán, L.; Uribe, E.; et al. Impacts of harmful algal blooms on the aquaculture industry: Chile as a case study. Perspect. Phycol. 2019, 6, 39–50. [Google Scholar] [CrossRef]
- Balech, E. Three new species of the genus Alexandrium (Dinoflagellata). Trans. Am. Microsc. Soc. 1994, 113, 216–220. [Google Scholar] [CrossRef]
- Giacobbe, M.G.; Penna, A.; Gangemi, E.; Maso, M.; Garcés, E.; Fraga, S.; Bravo, I.; Azzaro, F.; Penna, A. Recurrent high-biomass blooms of Alexandrium taylorii (Dinophyceae), a HAB species expanding in the Mediterranean. Hydrobiologia 2007, 580, 125–133. [Google Scholar] [CrossRef]
- Matsuyama, Y.; Usuki, H.; Uchida, T.; Kotani, Y. Effects of harmful algae on the early planktonic larvae of the oyster, Crassostrea gigas. In IX International Conference on Harmful Algal Blooms; Hallegraeff, G.M., Blackburn, S.I., Bolch, J.S., Lewis, R.J., Eds.; Intergovernmental Oceanographic Commission: Hobart, Australia, 2000; pp. 173–176. [Google Scholar]
- Lim, P.T.; Usup, G.; Leaw, C.P.; Ogata, T. First report of Alexandrium taylori and Alexandrium peruvianum (Dinophyceae) in Malaysia waters. Harmful Algae 2005, 4, 391–400. [Google Scholar] [CrossRef]
- Emura, A.; Matsuyama, Y.; Oda, T. Evidence for the production of a novel proteinaceous hemolytic exotoxin by dinoflagellate Alexandrium taylori. Harmful Algae 2004, 3, 29–37. [Google Scholar] [CrossRef]
- Penna, A.; Giacobbe, M.G.; Penna, N.; Andreoni, F.; Magnani, M. Seasonal blooms of the HAB Dinoflagellate Alexandrium taylori Balech in a New Mediterranean Area (Vulcano, Aeolian Islands). PSZN Mar. Ecol. 2002, 23, 1–9. [Google Scholar] [CrossRef]
- Garcés, E.; Delgado, M.; Maso, M.; Camp, J. Life history and in situ growth rates of Alexandrium taylori. J. Phycol. 1998, 34, 880–887. [Google Scholar] [CrossRef] [Green Version]
- Garcés, E.; Masó, M.; Camp, J. A recurrent and localized dinoflagellate bloom in a Mediterranean beach. J. Plankton Res. 1999, 21, 2373–2391. [Google Scholar] [CrossRef] [Green Version]
- Delgado, M.; Garcés, E.; Vila, M.; Camp, J. Morphological variability in three populations of the dinoflagellate Alexandrium taylori. J. Plankton Res. 1997, 19, 749–757. [Google Scholar] [CrossRef] [Green Version]
- Satta, C.T.; Pulina, S.; Padedda, B.M.; Penna, A.; Sechi, N.; Luglié, A. Water discoloration events caused by the harmful dinoflagellate Alexandrium taylorii Balech in a new beach of the Western Mediterranean Sea (Platamona beach, North Sardinia). Adv. Oceanogr. Limnol. 2010, 1, 259–269. [Google Scholar] [CrossRef]
- Basterretxea, G.; Garcés, E.; Jordi, A.; Maso, M.; Tintore, J. Breeze conditions as a favoring mechanism of Alexandrium taylori blooms at a Mediterranean beach. Estuar. Coast. Shelf Sci. 2005, 62, 1–12. [Google Scholar] [CrossRef]
- Penna, A.; Garcés, E.; Vila, M.; Giacobbe, M.G.; Fraga, S.; Luglié, A.; Bravo, I.; Bertozzini, E.; Vernesi, C. Alexandrium catenella (Dinophyceae), a toxic ribotype expanding in the NW Mediterranean Sea. Mar. Biol. 2005, 148, 13–23. [Google Scholar] [CrossRef]
- Penna, A.; Fraga, S.; Maso, M.; Giacobbe, M.G.; Bravo, I.; Garcés, E.; Vila, M.; Bertozzini, E.; Andreoni, F.; Luglie, A.; et al. Phylogenetic relationships among the Mediterranean Alexandrium (Dinophyceae) species based on sequences of 5.8S gene and Internal Transcript Spacers of the rRNA operon. Eur. J. Phycol. 2008, 43, 163–178. [Google Scholar] [CrossRef] [Green Version]
- Giacobbe, M.G.; Yang, X. The life history of Alexandrium taylori (Dinophyceae). J. Phycol. 1999, 35, 331–338. [Google Scholar] [CrossRef]
- Tillmann, U.; John, U. Toxic effects of Alexandrium spp. on heterotrophic dinoflagellates: An allelochemical defence mechanism independent of PSP toxins. Mar. Ecol. Prog. Ser. 2002, 230, 47–58. [Google Scholar] [CrossRef]
- Balech, E. The genus Alexandrium Halim (Dinoflagellata); Sherkin Island Marine Station Publication, Sherkin Island, Co.: Cork, Ireland, 1995; p. 151. [Google Scholar]
- Murray, S.A.; Diwan, R.; Orr, R.J.S.; Kohli, G.S.; John, U. Gene duplication, loss and selection in the evolution of saxitoxin biosythesis in alveolates. Mol. Phylogenet. Evol. 2015, 92, 165–180. [Google Scholar] [CrossRef] [Green Version]
- Tillmann, U.; Bantle, A.; Krock, B.; Elbrächter, M.; Gottschling, M. Molecular phylogenetics of the Gonyaucales based on curated rRNA sequence data and recommendations for epitypication of dinophytes exemplified with Lingulodinium polyedra. Harmful Algae 2020. in revision. [Google Scholar]
- Harding, J.M.; Mann, R.; Moeller, P.D.R.; Hsia, M.H. Mortality of the veined rapa whelk, Rapana venosa, in relation to a bloom of Alexandrium monilatum in the York River, United States. J. Shellfish Res. 2009, 28, 363–367. [Google Scholar] [CrossRef]
- Sharma, G.M.; Michaels, L.; Burkholder, P.R. Goniodomin, a new antibiotic from a dinoflagellate. J. Antibiot. 1968, 21, 659–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, C.M.; Reece, K.; Stec, D.F.; Scott, G.P.; Jones, W.M.; Hobbs, P.L.M.; Harris, T.M. The toxin goniodomin, produced by Alexandrium spp., is identical to goniodomin A. Harmful Algae 2020, 92, 101707. [Google Scholar] [CrossRef]
- Murakami, M.; Makabe, K.; Yamaguchu, K.; Konosu, S.; Wälchli, M.R. Goniodomin A, a novel polyether macrilide from the dinoflagellate Goniodoma pseudogonyaulax. Tetrahedron Lett. 1988, 29, 1149–1152. [Google Scholar] [CrossRef]
- Hsia, M.H.; Morton, S.L.; Smith, L.L.; Beauchesne, K.R.; Muncik, K.M.; Moeller, P.D.R. Production of goniodomin A by the planktonic, chain-forming dinoflagellate Alexandrium monilatum (Howell) Balech isolated from the Gulf Coast of the United States. Harmful Algae 2006, 5, 290–299. [Google Scholar] [CrossRef]
- Zmerli Triki, H.; Daly-Yahia, O.K.; Malouche, D.; Komiha, Y.; Deidun, A.; Brahim, M.; Laabir, M. Distribution of resting cysts of the potentially toxic dinoflagellate Alexandrium pseudogonyaulax in recently-deposited sediment within Bizerte Lagoon (Mediterranean coast, Tunisia). Mar. Pollut. Bull. 2014, 84, 172–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lilly, E.L.; Halanych, K.M.; Anderson, D.M. Species boundaries and global biogeography of the Alexandrium tamarense species complex. J. Phycol. 2007, 43, 1329–1338. [Google Scholar] [CrossRef]
- Lilly, E.L.; Halanych, K.M.; Anderson, D.M. Phylogeny, biogeography, and species boundaries within the Alexandrium minutum group. Harmful Algae 2005, 4, 1004–1020. [Google Scholar] [CrossRef]
- John, U.; Litaker, R.W.; Montresor, M.; Murray, S.; Brosnahan, M.L.; Anderson, D.M. Formal revision of the Alexandrium tamarense species complex (Dinophyceae) taxonmomy: The introduction of five species with emphasis on molecular-based (rDNA) classification. Protist 2014, 165, 779–804. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhuang, Y.; Zhang, H.; Lin, X.; Lin, S. DNA barcoding species in Alexandrium tamarense complex using ITS and proposing designation of five species. Harmful Algae 2014, 31, 100–113. [Google Scholar] [CrossRef]
- Litaker, R.W.; Fraga, S.; Montresor, M.; Brosnahan, M.L.; Hoppenrath, M.; Murray, S.; Wolny, J.; John, U.; Sampedro, N.; Larsen, J.; et al. A practical guide to new nomenclature for species within the “Alexandrium tamarense species complex”. Harmful Algae News 2018, 61, 13–15. [Google Scholar]
- Hansen, G.; Daugbjerg, N.; Franco, J.M. Morphology, toxin composition and LSU rDNA phylogeny of Alexandrium minutum (Dinophyceae) from Denmark, with some morphological observations on other European strains. Harmful Algae 2003, 2, 317–335. [Google Scholar] [CrossRef]
- Bravo, I.; Garcés, E.; Diogéne, J.; Fraga, S.; Sampedro, N.; Figueroa, R. Resting cysts of the toxigenic dinoflagellate genus Alexandrium in recent sediments from the Western Mediterranean coast, including the first description of cysts of A. kutnerae and A. perivianum. Eur. J. Phycol. 2006, 41, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Boczar, B.A.; Beitler, M.K.; Liston, J.; Sullivan, J.J.; Cattolico, R.A. Paralytic shellfish toxins in Protogonyaulax tamarensis and Protogonyaulax catenella in axenic culture. Plant Physiol. 1988, 88, 1285–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, S.A.; Wiese, M.; Neilan, B.; Orr, R.J.S.; de Salas, M.; Brett, S.; Hallegraeff, G. A reinvestigation of saxitoxin production and sxtA in the "non-toxic" Alexandrium tamarense group V clade. Hamful Algae 2012, 18, 96–104. [Google Scholar] [CrossRef]
- Kremp, A.; Tahvanainen, P.; Litaker, W.; Krock, B.; Suikkanen, S.; Leaw, C.P.; Tomas, C. Phylogenetic relationships, morphological variation, and toxin pattern in the Alexandrium ostenfeldii (Dinopyhceae) complex: Implications for species boundaries and identities. J. Phycol. 2014, 50, 81–100. [Google Scholar] [CrossRef]
- Suikkanen, S.; Kremp, A.; Hautala, H.; Krock, B. Paralytic shellfish toxins or spirolides? The role of environmental and genetic factors in toxin production of the Alexandrium ostenfeldii complex. Harmful Algae 2013, 26, 52–59. [Google Scholar] [CrossRef]
- Touzet, N.; Franco, J.M.; Raine, R. Characterization of nontoxic and toxin-producing strains of Alexandrium minutum (Dinophyceae) in Irish coastal waters. Appl. Environ. Microbiol. 2007, 73, 3333–3342. [Google Scholar] [CrossRef] [Green Version]
- Nguyen-Ngoc, L. An autoecological study of the potentially toxic dinoflagellate Alexandrium affine isolated from Vietnamese waters. Harmful Algae 2004, 3, 117–129. [Google Scholar] [CrossRef]
- Orr, R.J.S.; Stuken, A.; Rundberget, T.; Eikrem, W.; Jakobsen, K.S. Improved phylogenetic resolution of toxic and non-toxic Alexandrium strains using a concatenated rDNA approach. Hamful Algae 2011, 10, 676–688. [Google Scholar] [CrossRef]
- Ciminiello, P.; Fatturoso, E.; Forino, M.; Montresor, M. Saxitoxin and neosaxitoxin as toxic principles of Alexandrium andersoni (Dinophyceae) from the Gulf of Naples, Italy. Toxicon 2000, 38, 1871–1877. [Google Scholar] [CrossRef]
- Usup, G.; Leaw, C.P.; Ahmad, A.; Lim, P.T. Alexandrium (Dinophyceae) species in Malaysian waters. Harmful Algae 2002, 1, 265–275. [Google Scholar] [CrossRef]
- Mertens, K.; Adachi, M.; Anderson, D.M.; Band-Schmidt, C.; Bravo, I.; Brosnahan, M.L.; Bolach, C.J.; Calado, A.; Carbonell-Moore, M.C.; Chomérat, N.; et al. Morphological and phylogenetic data do not support the split of Alexandrium into four genera. Harmful Algae 2020. in revision. [Google Scholar]
- Kita, T.; Fukuyo, Y. Description of the gonyaulacoid dinoflagellate Alexandrium hiranoi sp. nov. inhabiting tidepools on Japanese Pacific coast. Bull. Plankton Soc. Jpn. 1988, 35, 1–7. [Google Scholar]
- Lim, A.S.; Jeong, H.J.; Lee, S.Y. Description of the new phototrophic dinoflagellate Alexandrium pohangense sp. nov. from Korean coastal waters. Harmful Algae 2015, 46, 49–61. [Google Scholar] [CrossRef]
- Clemons, G.P.; Pinion, J.P.; Bass, E.; Pham, D.V.; Sharif, M.; Wutoh, J.G. A hemolytic principle associated with the red-tide dinoflagellate Gonyaulax monilata. Toxicon 1980, 18, 323–326. [Google Scholar] [CrossRef]
- Gates, J.A.; Wilson, W.B. The toxicity of Gonyaulax monilata Howell to Mugil Cephalus. Limnol. Oceanogr. 1960, 5, 171–174. [Google Scholar] [CrossRef]
- May, S.P.; Burkholder, J.M.; Shumway, S.E.; Hégaret, H.; Wikfors, G.H. Effects of the toxic dinoflagellate Alexandrium monilatum on survival, grazing and behavioral response of three ecologically important bivalve molluscs. Harmful Algae 2010, 9, 281–293. [Google Scholar] [CrossRef]
- Fritz, L.; Triemer, R.E. A rapid simple technique utilizing Calcofluor White M2R for the visualization of dinoflagellate thecal plates. J. Phycol. 2004, 21, 662–664. [Google Scholar] [CrossRef]
- Agatha, S.; Strüder-Kypke, M.C.; Beran, A. Morphological and genetic variability in the marine planktonic ciliate Laboea strobila Lohmann, 1908 (Ciliophora, Oligotrichia), with notes on its ontogenesis. J. Eukaryot. Microbiol. 2004, 51, 267–281. [Google Scholar] [CrossRef] [Green Version]
- Keller, M.D.; Selvin, R.C.; Claus, W.; Guillard, R.R.L. Media for the culture of oceanic ultraphytoplankton. J. Phycol. 1987, 23, 633–638. [Google Scholar] [CrossRef]
- Chomérat, N.; Gatti, C.M.I.; Nézan, E.; Chinain, M. Studies on the benthic genus Sinophysis (Dinophysales, Dinophyceae) II. S. canaliculata from Rapa Island (French Polynesia). Phycologia 2017, 56, 193–203. [Google Scholar]
- Tillmann, U.; Trefault, N.; Krock, B.; Parada-Pozo, G.; De la Iglesia, R.; Vásquez, M. Identification of Azadinium poporum (Dinophyceae) in the Southeast Pacific: Morphology, molecular phylogeny, and azaspiracid profile characterization. J. Plankton Res. 2017, 39, 350–367. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krock, B.; Tillmann, U.; Wen, Y.; Hansen, P.J.; Larsen, T.O.; Andersen, A.J.C. Development of a LC-MS/MS method for the quantification of goniodomins A and B and its application to Alexandrium pseudogonyaulax strains and plankton field samples of Danish coastal waters. Toxicon 2018, 155, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tillmann, U.; Alpermann, T.; da Purificação, R.C.; Krock, B.; Cembella, A. Intra-population clonal variability in allelochemical potency of the toxigenic dinoflagellate Alexandrium tamarense. Harmful Algae 2009, 8, 759–769. [Google Scholar] [CrossRef]
- Ma, H.; Krock, B.; Tillmann, U.; Cembella, A. Preliminary characterization of extracellular allelochemicals of the toxic marine dinoflagellate Alexandrium tamarense using a Rhodomonas salina bioassay. Mar. Drugs 2009, 7, 497–522. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Sequence (5′-3′) | Nucleotide Positions (bp) |
|---|---|---|
| AY7T (this study) | CATTAATTTGGACTTGGTGCAA | 547–568 |
| AY4T | ---------------------AG--------------------- | 555–576 |
| AY1T | ---------------------------------------------- | 516–537 |
| AY2T | ---------------------------------------------- | 551–572 |
| Strain | Sequence (5′-3′) | Nucleotide Positions (bp) |
|---|---|---|
| AY7T (this study) | GATCCAA……….AGGCATC | 354–360……….494–500 |
| CSIC-AV8 | ------T------………..-------------- | 314–320……….454–460 |
| VGO704, VGOE6 | --------------………..------A----- | 314–320……….454–460 |
| AY10T | --------------………..-------------- | 314–320……….454–460 |
| AY1T | --------------………..-------------- | 314–320……….454–460 |
| AY7T | --------------………..-------------- | 314–320……….454–460 |
| CBA-1 | --------------………..-------------- | 314–320……….454–460 |
| CNR-AT4 | --------------………..-------------- | 314–320……….454–460 |
| CNR-ATAYB2 | --------------………..-------------- | 314–320……….454–460 |
| Field sample | --------------………..-------------- | 314–320……….454–460 |
| Temporary-cyst | --------------………..-------------- | 314–320……….454–460 |
| VGO705 | --------------………..-------------- | 314–320……….454–460 |
| Toxin | LOD (FLD) [fg cell−1] | LOD (MS/MS) [fg cell−1] |
|---|---|---|
| C1 | 57 | 0.25 |
| C2 | 57 | 0.76 |
| C3 | nd | 0.49 |
| C4 | nd | 1.87 |
| B1 | 141 | 0.12 |
| B2 | nd | 0.49 |
| STX | 35 | 0.13 |
| NEO | 516 | 0.63 |
| GTX1 | 715 | 0.07 |
| GTX2 | 26 | 0.24 |
| GTX3 | 32 | 0.18 |
| GTX4 | 722 | 0.16 |
| dcSTX | 51 | 0.15 |
| dcNEO | nd | 0.34 |
| dcGTX1 | nd | 0.35 |
| dcGTX2 | 25 | 0.49 |
| dcGTX3 | 25 | 0.45 |
| dcGTX4 | nd | 0.84 |
| doSTX | nd | 0.09 |
| TTX | nd | 0.18 |
| Quantifier + | Qualifier + | Quantifier - | Qualifier - | |
|---|---|---|---|---|
| doSTX | 241 > 60 | 241 > 206 | ||
| dcSTX | 257 > 126 | 257 > 222 | ||
| dcNEO | 273 > 126 | 273 > 225 | ||
| STX | 300 > 126 | 300 > 204 | ||
| NEO | 316 > 126 | 316 > 220 | ||
| TTX | 320 > 302 | 320 > 162 | ||
| dcGTX2 | 351 > 164 | 351 > 333 | ||
| dcGTX3 | 353 > 255 | 351 > 333 | ||
| dcGTX1 | 367 > 274 | 367 > 349 | ||
| dcGTX4 | 369 > 271 | 367 > 349 | ||
| B1 | 380 > 300 | 378 > 122 | ||
| B2 | 396 > 316 | 394 > 122 | ||
| GTX2 | 394 > 351 | 394 > 333 | ||
| GTX3 | 396 > 298 | 394 > 333 | ||
| GTX1 | 410 > 367 | 410 > 349 | ||
| GTX4 | 412 > 314 | 410 > 367 | ||
| C1 | 474 > 122 | 474 > 351 | ||
| C2 | 396 > 298 | 474 > 122 | ||
| C3 | 412 > 332 | 490 > 410 | ||
| C4 | 412 > 314 | 490 > 392 | ||
| GC3 | 377 > 359 | 377 > 257 | ||
| GC3a | 393 > 375 | 393 > 257 | ||
| GC6 | 393 > 375 | 393 > 273 | ||
| GC6a | 409 > 391 | 409 > 273 | ||
| GC3b | 457 > 359 | 457 > 377 | ||
| GC1/2 | 473 > 375 | 473 > 455 | ||
| GC6b | 473 > 375 | 473 > 393 | ||
| GC1a/2a | 489 > 409 | 489 > 471 | ||
| GC4/5 | 489 > 489 | 489 > 471 | ||
| GC4a/5a | 505 > 425 | 505 > 487 | ||
| GC1b/GC2b | 553 > 393 | 553 > 473 | ||
| GC4b | 569 > 489 | 569 > 409 | ||
| GC5b | 569 > 409 | 569 > 489 |
| Toxin | Quantifier + | Qualifier + |
|---|---|---|
| GYM A | 508 > 490 | 508 > 162 |
| GYM D | 510 > 492 | |
| 12-me-GYM A | 522 > 504 | |
| GYM B/C | 524 > 506 | |
| GYM E | 526 > 508 | |
| GYM (uncharacterized) | 540 > 522 | |
| GYM (uncharacterized) | 542 > 524 | |
| SPX (uncharacterized) | 592 > 164 | |
| SPX (uncharacterized) | 618 > 164 | |
| SPX H | 650 > 164 | |
| SPX I | 652 > 164 | |
| SPX (uncharacterized) | 658 > 164 | |
| SPX (uncharacterized) | 666 > 164 | |
| SPX (uncharacterized) | 666 > 180 | |
| SPX (uncharacterized) | 678 > 150 | |
| 13,19-Didesmethyl-SPX C | 678 > 164 | |
| SPX (uncharacterized) | 686 > 164 | |
| SPX A | 692 > 150 | |
| 13-Desme-SPX C, SPX G | 692 > 164 | |
| SPX (uncharacterized) | 692 > 180 | |
| SPX B | 694 > 150 | |
| 13-Desme-SPX D, PnTx G, 20-Hydroxy-13,19-didesmethyl SPX C | 694 > 164 | |
| 27-Hydroxy-13-desmethyl SPX C | 694 > 180 | |
| 20-Hydroxy-13,19-didesmethyl-SPX D | 696 > 164 | |
| SPX (uncharacterized) | 698 > 164 | |
| SPX (uncharacterized) | 704 > 164 | |
| SPX (uncharacterized) | 706 > 150 | |
| SPX C, 20-Methyl-SPX G | 706 > 164 | |
| SPX D | 708 > 164 | |
| SPX (uncharacterized) | 708 > 180 | |
| SPX (uncharacterized) | 710 > 150 | |
| SPX (uncharacterized) | 710 > 164 | |
| SPX (uncharacterized) | 718 > 164 | |
| SPX (uncharacterized) | 720 > 150 | |
| SPX (uncharacterized) | 720 > 164 | |
| SPX (uncharacterized) | 722 > 164 | |
| SPX (uncharacterized) | 722 > 180 | |
| PnTx F | 766 > 164 | |
| PnTx E | 784 > 164 | |
| GDA | 786 > 607 | 786 > 733 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tillmann, U.; Krock, B.; Wietkamp, S.; Beran, A. A Mediterranean Alexandrium taylorii (Dinophyceae) Strain Produces Goniodomin A and Lytic Compounds but Not Paralytic Shellfish Toxins. Toxins 2020, 12, 564. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12090564
Tillmann U, Krock B, Wietkamp S, Beran A. A Mediterranean Alexandrium taylorii (Dinophyceae) Strain Produces Goniodomin A and Lytic Compounds but Not Paralytic Shellfish Toxins. Toxins. 2020; 12(9):564. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12090564
Chicago/Turabian StyleTillmann, Urban, Bernd Krock, Stephan Wietkamp, and Alfred Beran. 2020. "A Mediterranean Alexandrium taylorii (Dinophyceae) Strain Produces Goniodomin A and Lytic Compounds but Not Paralytic Shellfish Toxins" Toxins 12, no. 9: 564. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12090564