Natural Thiols, but Not Thioethers, Attenuate Patulin-Induced Endoplasmic Reticulum Stress in HepG2 Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of Patulin on Metabolic Activity of HepG2 Cells
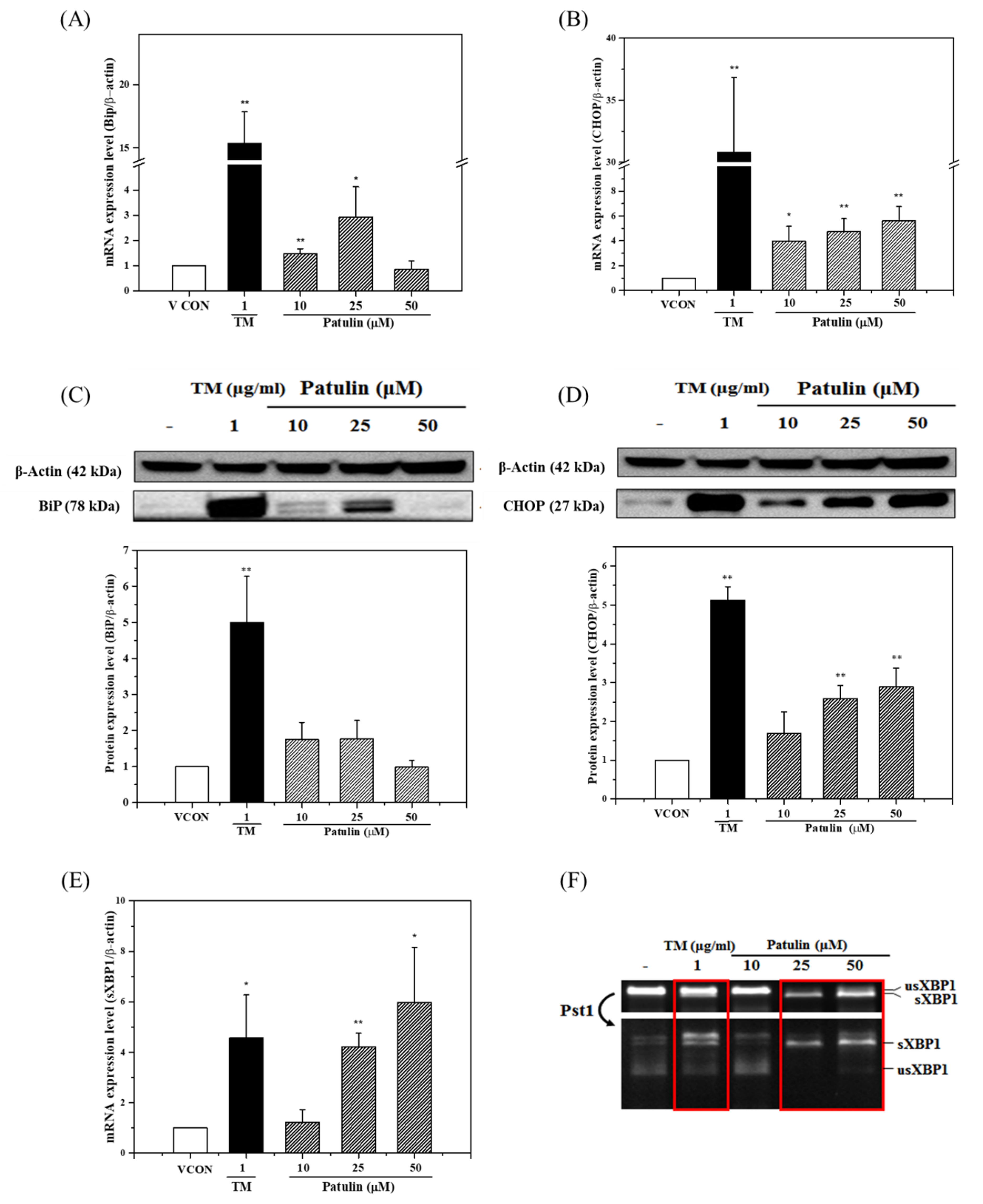
2.2. Effects of Patulin on the Expression Levels of ER Stress Marker Genes and Proteins
2.3. Attenuation of Patulin-Induced Cytotoxicity by Sulfur-Containing Compounds
2.4. Effects of Thiol-Containing Compounds on Patulin-Induced ER Stress
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture
4.3. Cell Metabolic Activity
4.4. Reverse Transcription-Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR) Analysis
4.5. XBP1 Splicing
4.6. Western Blot Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Baert, K.; Devlieghere, F.; Flyps, H.; Oosterlinck, M.; Ahmed, M.M.; Rajković, A.; Verlinden, B.; Nicolaï, B.; Debevere, J.; Meulenaer, B.D. Influence of storage conditions of apples on growth and patulin production by Penicillium expansum. Int. J. Food Microbiol. 2007, 119, 170–181. [Google Scholar] [CrossRef]
- Leggott, N.L.; Shephard, G.S. Patulin in South African commercial apple products. Food Control 2001, 12, 73–76. [Google Scholar] [CrossRef]
- Plunkett, L.M.; Turnbull, D.; Rudricks, J.V. Differences between adults and children affecting exposure assessment. In Similarities and Differences between Children and Adults; Guelian, P.S., Henry, C.J., Olin, S.S., Eds.; ILSI Press: Washington, DC, USA, 1992; pp. 79–94. [Google Scholar]
- Zaied, C.; Abid, S.; Hlel, W.; Bacha, H. Occurrence of patulin in apple-based-foods largely consumed in Tunisia. Food Control 2013, 31, 263–267. [Google Scholar] [CrossRef]
- Escoula, L.; Thomsen, M.; Bourdiol, D.; Pipy, B.; Peuriere, S.; Roubinet, F. Patulin immunotoxicology: Effect on phagocyte activation and the cellular and humoral immune system of mice and rabbits. Int. J. Immunopharmacol. 1988, 10, 983–989. [Google Scholar] [CrossRef]
- Walker, K.; Wiesner, B.P. Patulin and clavacin. Lancet 1944, 246, 294. [Google Scholar] [CrossRef]
- Hayes, A.W.; Phillips, T.D.; Williams, W.L.; Ciegler, A. Acute toxicity of patulin in mice and rats. Toxicology 1979, 13, 91–100. [Google Scholar] [CrossRef] [PubMed]
- McKinley, E.R.; Carlton, W.W.; Boon, G.D. Patulin mycotoxicosis in the rat: Toxicology, pathology, and clinical pathology. Food Chem. Toxicol. 1982, 20, 289–300. [Google Scholar] [CrossRef]
- Mahfoud, R.; Maresca, M.; Garmy, N.; Fantini, J. The mycotoxin patulin alters the barrier function of the intestinal epithelium: Mechanism of action of the toxin and protective effects of glutathione. Toxicol. Appl. Pharmacol. 2002, 181, 209–218. [Google Scholar] [CrossRef]
- Oswald, H.; Frank, H.K.; Komitowski, D.; Winter, H. Long-term testing of patulin administered orally to Sprague-Dawley rats and Swiss mice. Food Cosmet. Toxicol. 1978, 16, 243–247. [Google Scholar] [CrossRef]
- Pfeiffer, E.; Gross, K.; Metzler, M. Aneuploidogenic and clastogenic potential of the mycotoxins citrinin and patulin. Carcinogenesis 1998, 19, 1313–1318. [Google Scholar] [CrossRef] [Green Version]
- Wichmann, G.; Herbarth, O.; Lehmann, I. The mycotoxins citrinin, gliotoxin, and patulin affect interferon-rather than interleukin-4 production in human blood cells. Environ. Toxicol. 2002, 17, 211–218. [Google Scholar] [CrossRef]
- Lee, K.S.; Röschenthaler, R.J. DNA-damaging activity of patulin in Escherichia coli. Appl. Environ. Microbiol. 1986, 52, 1046–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roll, R.; Matthiaschk, G.; Korte, A. Embryotoxicity and mutagenicity of mycotoxins. J. Environ. Pathol. Toxicol. Oncol. 1990, 10, 1–7. [Google Scholar] [PubMed]
- Ciegler, A.; Beck, A.C.; Jackson, L.K. Teratogenicity of patulin and patulin adducts formed with cysteine. Appl. Environ. Microbiol. 1976, 31, 664–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickens, F.; Jones, H.E.H. Carcinogenic activity of a series of reactive lactones and related substances. Br. J. Cancer 1961, 15, 85–100. [Google Scholar] [CrossRef] [Green Version]
- International Agency for Research on Cancer (IARC). Patulin. In Monograph No. 40 on the Evaluation of Carcinogenic Risk of Chemicals to Man; IARC: Lyon, France, 1986; pp. 83–98. [Google Scholar]
- Liu, B.H.; Wu, T.S.; Yu, F.Y.; Su, C.C. Induction of oxidative stress response by the mycotoxin patulin in mammalian cells. Toxicol. Sci. 2006, 95, 340–347. [Google Scholar] [CrossRef] [Green Version]
- Morgavi, D.P.; Boudra, H.; Jouany, J.P.; Graviou, D. Prevention of patulin toxicity on rumen microbial fermentation by SH-containing reducing agents. J. Agric. Food Chem. 2003, 51, 6906–6910. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: Introductory remarks. In Oxidative Stress, 1st ed.; Sies, H., Ed.; Academic Press: London, UK, 1985; pp. 1–8. [Google Scholar]
- Henle, E.S.; Linn, S. Formation, prevention, and repair of DNA damage by iron/hydrogen peroxide. J. Biol. Chem. 1997, 272, 19095–19098. [Google Scholar] [CrossRef] [Green Version]
- Boussabbeh, M.; Salem, I.B.; Prola, A.; Guilbert, A.; Bacha, H.; Abid-Essefi, S.; Lemaire, C. Patulin induces apoptosis through ROS-mediated endoplasmic reticulum stress pathway. Toxicol. Sci. 2015, 144, 328–337. [Google Scholar] [CrossRef] [Green Version]
- Lindholm, D.; Wootz, H.; Korhonen, L. ER stress and neurodegenerative diseases. Cell Death Differ. 2006, 13, 385–392. [Google Scholar] [CrossRef]
- Wu, J.; Yuan, R.J. From acute ER stress to physiological roles of the unfolded protein response. Cell Death Differ. 2006, 13, 374–384. [Google Scholar] [CrossRef]
- Verkhratsky, A. Physiology and pathophysiology of the calcium store in the endoplasmic reticulum of neurons. Physiol. Rev. 2005, 85, 201–279. [Google Scholar] [CrossRef]
- Kim, I.; Xu, W.; Reed, J.C. Cell death and endoplasmic reticulum stress: Disease relevance and therapeutic opportunities. Nat. Rev. Drug Discov. 2008, 7, 1013–1030. [Google Scholar] [CrossRef]
- Hetz, C. The unfolded protein response: Controlling cell fate decisions under ER stress and beyond. Nat. Rev. Mol. Cell Biol. 2012, 13, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Tennant, B.C. Hepatic Function. In Clinical Biochemistry of Domestic Animals, 5th ed.; Kaneko, J.J., Harvey, J.W., Bruss, M.L., Eds.; Academic Press: San Diego, CA, USA, 1997; pp. 327–352. [Google Scholar]
- Ayed-Boussema, I.; Abassi, H.; Bouaziz, C.; Hlima, W.B.; Ayed, Y.; Bacha, H. Antioxidative and antigenotoxic effect of vitamin E against patulin cytotoxicity and genotoxicity in HepG2 cells. Environ. Toxicol. 2011, 28, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Diao, E.; Ren, D.; Liu, T.; Zhang, J.; Hu, W.; Hou, H. Ozone detoxification of patulin in aqueous solution and cytotoxic evaluation using human hepatic carcinoma cells. Toxicon 2018, 155, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Van Tonder, A.; Joubert, A.M.; Cromarty, A.D. Limitations of the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay when compared to three commonly used cell enumeration assays. BMC Res. Notes 2015, 8, 47. [Google Scholar] [CrossRef] [Green Version]
- Donato, M.T.; Tolosa, L.; Gómez-Lechón, M.J. Culture and functional characterization of human hepatoma HepG2 cells. Methods Mol. Biol. 2015, 1250, 77–93. [Google Scholar]
- Ellis, J. Proteins as molecular chaperones. Nature 1987, 328, 378–379. [Google Scholar] [CrossRef] [PubMed]
- Dorner, A.J.; Wasley, L.C.; Kaufman, R.J. Overexpression of GRP78 mitigates stress induction of glucose regulated proteins and blocks secretion of selective proteins in Chinese hamster ovary cells. EMBO J. 1992, 11, 1563–1571. [Google Scholar] [CrossRef]
- Jin, T.; Gu, Y.; Zanusso, G.; Sy, M.; Kumar, A.; Cohen, M.; Gambetti, P.; Singh, N. The chaperone protein BiP binds to a mutant prion protein and mediates its degradation by the proteasome. J. Biol. Chem. 2000, 275, 38699–38704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell. Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Gülow, K.; Bienert, D.; Haas, I.G. BiP is feed-back regulated by control of protein translation efficiency. J. Cell Sci. 2002, 115, 2443–2452. [Google Scholar] [CrossRef]
- Zha, L.; Fan, L.; Sun, G.; Wang, H.; Ma, T.; Zhong, F.; Wei, W. Melatonin sensitizes human hepatoma cells to endoplasmic reticulum stress-induced apoptosis. J. Pineal. Res. 2012, 52, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Guo, Y.; Tang, J.; Jiang, J.; Chen, Z. New insights into the roles of CHOP-induced apoptosis in ER stress. Acta Biochim. Biophys. Sin. 2014, 46, 629–640. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, H.; Matsui, T.; Yamamoto, A.; Okada, T.; Mori, K. XBP1 mRNA is induced by ATF6 and spliced by IRE1 in response to ER stress to produce a highly active transcription factor. Cell 2001, 107, 881–891. [Google Scholar] [CrossRef] [Green Version]
- Kwon, K.; Goo, T.W.; Kwon, O.Y. Development of rapid detection method for unfolded protein response in the mammalian cells. J. Exp. Biomed. Sci. 2005, 11, 249–252. [Google Scholar]
- Fliege, R.; Metzler, M. Electrophilic properties of patulin. N-acetylcysteine and glutathione adducts. Chem. Res. Toxicol. 2000, 13, 373–381. [Google Scholar] [CrossRef]
- Zhou, S.M.; Jiang, L.P.; Geng, C.Y.; Cao, J.; Zhong, L.F. Patulin-induced genotoxicity and modulation of glutathione in HepG2 cells. Toxicon 2009, 53, 584–586. [Google Scholar] [CrossRef]
- Atkuri, K.R.; Mantovani, J.J.; Herzenberg, L.A.; Herzenberg, L.A. N-acetylcysteine—A safe antidote for cysteine/glutathione deficiency. Curr. Opin. Pharm. 2007, 7, 355–359. [Google Scholar] [CrossRef] [Green Version]
- El-Ashmawy, N.E.; Khedr, N.F.; El-Bahrawy, H.A.; Hamada, O.B. Anti-inflammatory and antioxidant effects of captopril compared to methylprednisolone in l-arginine-induced acute pancreatitis. Dig. Dis. Sci. 2018, 63, 1497–1505. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.M.; Choi, H.Y.; Cho, G.H.; Im, J.H.; Hong, E.Y.; Chun, H.S. Natural Thiols, but Not Thioethers, Attenuate Patulin-Induced Endoplasmic Reticulum Stress in HepG2 Cells. Toxins 2021, 13, 727. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100727
Kim HM, Choi HY, Cho GH, Im JH, Hong EY, Chun HS. Natural Thiols, but Not Thioethers, Attenuate Patulin-Induced Endoplasmic Reticulum Stress in HepG2 Cells. Toxins. 2021; 13(10):727. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100727
Chicago/Turabian StyleKim, Hye Mi, Hwa Young Choi, Gun Hee Cho, Ju Hee Im, Eun Young Hong, and Hyang Sook Chun. 2021. "Natural Thiols, but Not Thioethers, Attenuate Patulin-Induced Endoplasmic Reticulum Stress in HepG2 Cells" Toxins 13, no. 10: 727. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100727