Recombinant Production and Characterization of a New Toxin from Cryptops iheringi Centipede Venom Revealed by Proteome and Transcriptome Analysis

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
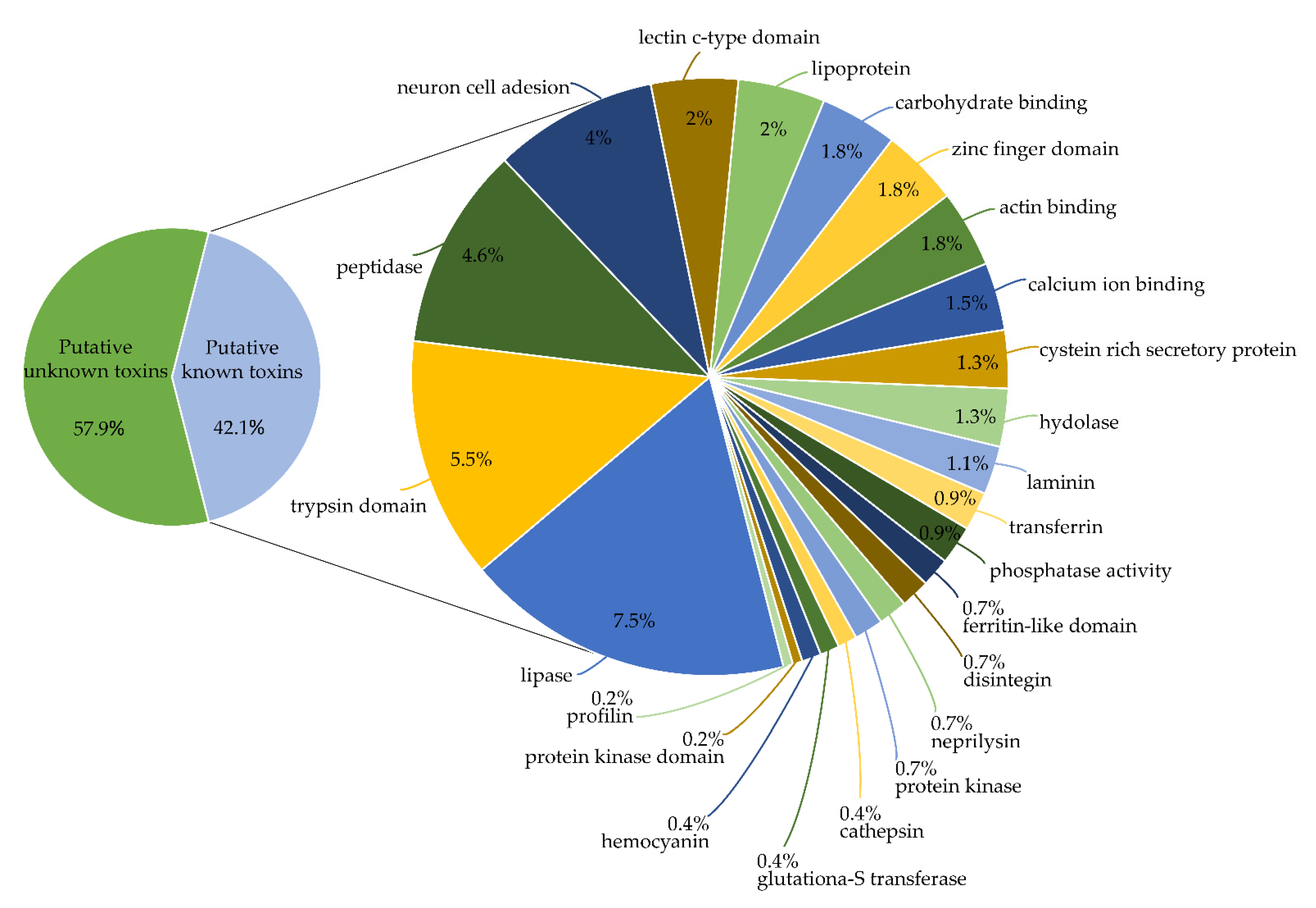
2.1. Identification of Toxins from Transcriptomic and Proteomic Analysis
2.2. Novel Toxins Identified by the Mass Spectrometry
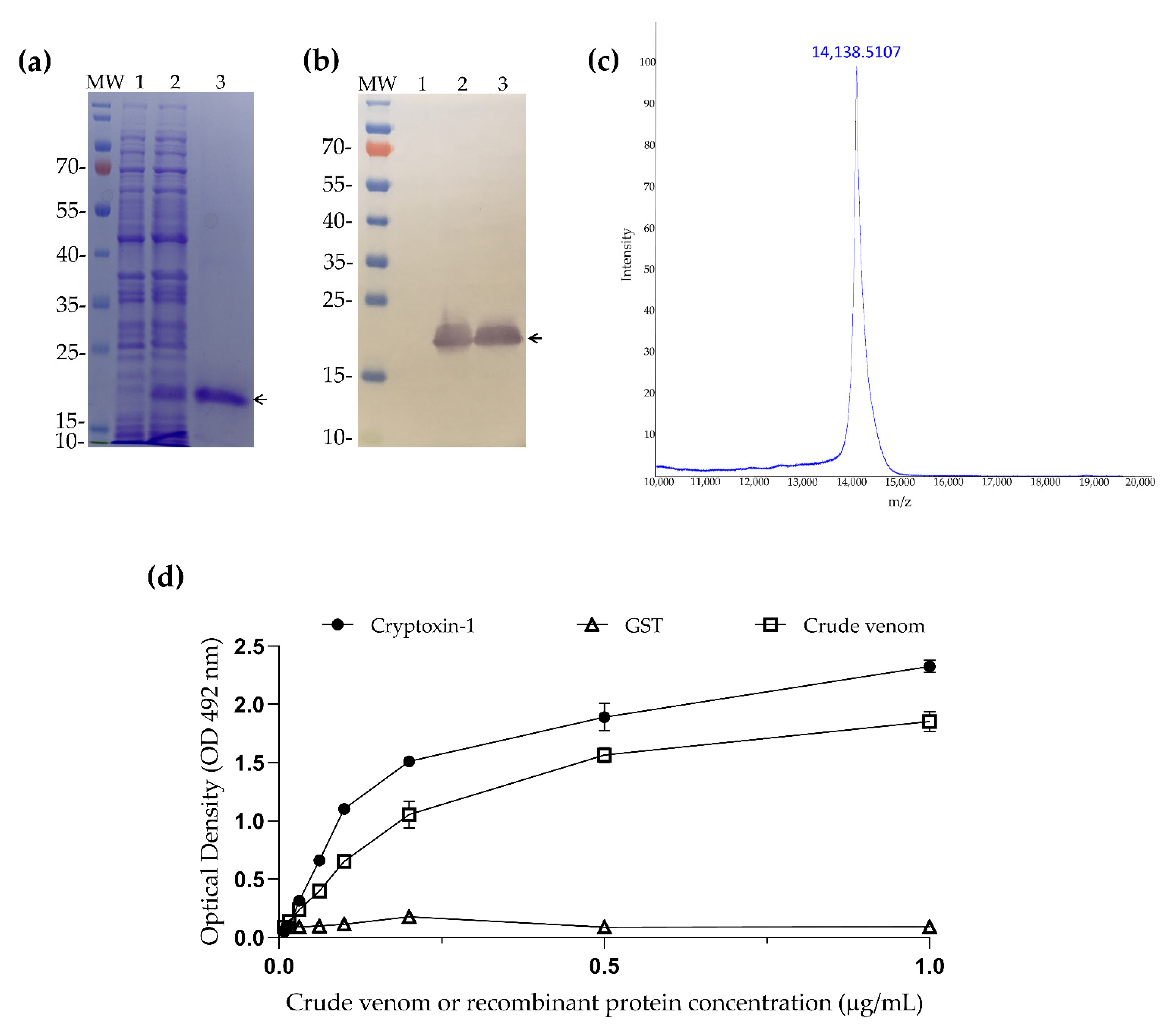
2.3. Cloning and Expression of Cryptoxin-1
2.4. Crypotoxin-1 Induces Edema in Mice Footpad
2.5. Cryptoxin-1 Induces Potent Neutrophil Migration in Mice Footpad
3. Discussion
4. Materials and Methods
4.1. Specimen Collection and Venom Extraction
4.2. RNA Isolation, Library Preparation, and Illumina Sequencing
4.3. RNA-Seq Raw Data Pre-Processing, De Novo Assembly, and Functional Annotation
4.4. SDS-PAGE and LC-MS/MS Analysis of C. iheringi
4.5. LC-MS/MS Data Analysis
4.6. Protein Expression and Purification
4.7. Mass Spectrometry Analysis
4.8. Rabbit Specific Antivenom Production
4.9. ELISA Immunoassay
4.10. Mice
4.11. Evaluation of Paw Edema
4.12. Histological Analysis
4.13. Analysis of the Neutrophil Infiltrate in Footpad Tissue by Flow Cytometry
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bücherl, W.B.; Buckley, E.E. Venomous Chilopods or Centipeds; Academic Press: New York, NY, USA, 1971; Volume 3. [Google Scholar]
- Knysak, I.; Martins, R.; Bertim, C.R. Epidemiological aspects of centipede (Scolopendromorphae: Chilopoda) bites registered in greater S. Paulo, SP, Brazil. Revista Saúde Pública 1998, 32, 514–518. [Google Scholar] [CrossRef] [Green Version]
- Attems, C.G. Myriapoda II Scolopendromorpha. Berl. Leipz. 1930, 54, 1–308. [Google Scholar]
- Bucherl, W. Ação do veneno dos escolopendromorfos do Brasil sobre alguns animais de laboratório. Mem. Inst. Butantan 1947, 19, 181–198. [Google Scholar]
- Antoniazzi, M.M.; Pedroso, C.M.; Knysak, I.; Martins, R.; Guizze, S.P.G.; Jared, C.; Barbaro, K.C. Comparative mor-phological study of the venom glands of the centipede Cryptops iheringi, Otostigmus pradoi and Scolopendra viridicomis. Toxicon 2009, 53, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Stankiewicz, M.; Hamon, A.; Benkhalifa, R.; Kadziela, W.; Hue, B.; Lucas, S.; Mebs, D.; Pelhate, M. Effects of a centipede venom fraction on insect nervous system, a native Xenopus oocyte receptor and on an expressed Drosophila muscarinic receptor. Toxicon 1999, 37, 1431–1445. [Google Scholar] [CrossRef]
- Undheim, E.A.B.; King, G.F. On the venom system of centipedes (Chilopoda), a neglected group of venomous animals. Toxicon 2011, 57, 512–524. [Google Scholar] [CrossRef]
- Liu, Z.-C.; Liang, J.-Y.; Lan, X.-Q.; Li, T.; Zhang, J.-R.; Zhao, F.; Li, G.; Chen, P.-Y.; Zhang, Y.; Lee, W.-H.; et al. Comparative analysis of diverse toxins from a new pharmaceutical centipede. Zool. Res. 2020, 41, 138. [Google Scholar]
- Yang, S.; Liu, Z.H.; Xiao, Y.; Li, Y.; Rong, M.Q.; Liang, S.P.; Zhang, Z.Y.; Yu, H.N.; King, G.F.; Lai, R. Chemical Punch Packed in Venoms Makes Centipedes Excellent Predators. Mol. Cell. Proteom. 2012, 11, 640–650. [Google Scholar] [CrossRef] [Green Version]
- Undheim, E.A.B.; Jones, A.; Clauser, K.R.; Holland, J.W.; Pineda, S.S.; King, G.F.; Fry, B.G. Clawing through Evolution: Toxin Diversification and Convergence in the Ancient Lineage Chilopoda (Centipedes). Mol. Biol. Evol. 2014, 31, 2124–2148. [Google Scholar] [CrossRef] [Green Version]
- Pemberton, R.W. Insects and other arthropods used as drugs in Korean traditional medicine. J. Ethnopharmacol. 1999, 65, 207–216. [Google Scholar] [CrossRef]
- Kong, Y.; Shao, Y.; Chen, H.; Ming, X.; Wang, J.B.; Li, Z.Y.; Wei, J.F. A Novel Factor Xa-Inhibiting Peptide from Cen-tipedes Venom. Int. J. Pept. Res. Ther. 2013, 19, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Medeiros, C.R.; Susaki, T.T.; Knysak, I.; Cardoso, J.L.; Málaque, C.M.S.; Fan, H.W.; Santoro, M.L.; França, F.O.S.; Barbaro, K.C. Epidemiologic and clinical survey of victims of centipede stings admitted to Hospital Vital Brazil (São Paulo, Brazil). Toxicon 2008, 52, 606–610. [Google Scholar] [CrossRef]
- Barroso, E.; Hidaka, A.; Santos, A.; França, J.; Sousa, A.; Valente, J.; Magalhães, A.; Pardal, P. Centipede accidents notified by “Centro de Informações Toxicológicas de Belém”, over a two-year period. Rev. Soc. Bras. Med. Trop. 2001, 34, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Mumcuoglu, K.Y.; Leibovici, V. Centipede (Scolopendra) bite: A case report. Isr. J. Med. Sci. 1989, 25, 47–49. [Google Scholar]
- Harada, K.; Asa, K.; Imachi, T.; Yamaguchi, Y.; Yoshida, K.-I. Centipede inflicted postmortem injury. J. Forensic Sci. 1999, 44, 849–850. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.-K.; Hsu, S.-P.; Chi, C.-C.; Lee, K.-F.; Lin, P.Y.; Chang, H.-W.; Chuang, F.-R. Rhabdomyolysis, acute renal failure, and multiple focal neuropathies after drinking alcohol soaked with centipede. Ren. Fail. 2004, 26, 93–97. [Google Scholar] [CrossRef]
- Ozsarac, M.; Karcioglu, O.U.; Ayrik, C.; Somuncu, F.; Gumrukcu, S. Acute coronary ischemia following centipede envenomation: Case report and review of the literature. Wilderness Environ. Med. 2004, 15, 109–112. [Google Scholar] [CrossRef]
- Yildiz, A.; Biceroglu, S.; Yakut, N.; Bilir, C.; Akdemir, R.; Akilli, A. Acute myocardial infarction in a young man caused by centipede sting. Emerg. Med. J. 2006, 23, 3. [Google Scholar] [CrossRef]
- Hasan, S.; Hassan, K. Proteinuria associated with centipede bite. Pediatr. Nephrol. 2004, 20, 550–551. [Google Scholar] [CrossRef]
- Mohamed, A.H.; Zaid, E.; El-Beih, N.M.; Abd El-Aal, A.A. Effects of an extract from the centipede Scolopendra moristans on intestine, uterus and heart contractions and on blood glucose and liver and muscle glycogen levels. Toxicon 1980, 18, 581–589. [Google Scholar] [CrossRef]
- Mohamed, A.H.; Abu-Sinna, G.; El-Shabaka, H.A.; Abd El-Aal, A. Proteins, lipids, lipoproteins and some enzyme char-acterizations of the venom extract from the centipede Scolopendra morsitans. Toxicon 1983, 21, 371–377. [Google Scholar] [CrossRef]
- Gomes, A.; Datta, A.; Sarangi, B.; Kar, P.K.; Lahiri, S.C. Isolation, purification & pharmacodynamics of a toxin from the venom of the centipede Scolopendra subspinipes dehaani Brandt. Indian J. Exp. Boil. 1983, 21, 203–207. [Google Scholar]
- Cooper, A.M.; Fox, G.A.; Nelsen, D.R.; Hayes, W.K. Variation in venom yield and protein concentration of the centipedes Scolopendra polymorpha and Scolopendra subspinipes. Toxicon 2014, 82, 30–51. [Google Scholar] [CrossRef] [PubMed]
- Malta, M.B.; Lira, M.S.; Soares, S.L.; Rocha, G.C.; Knysak, I.; Martins, R.; Guizze, S.P.G.; Santoro, M.L.; Barbaro, K.C. Toxic activities of Brazilian centipede venoms. Toxicon 2008, 52, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Fung, H.T.; Lam, S.K.; Wong, O.F. Centipede bite victims: A review of patients presenting to two emergency departments in Hong Kong. Hong Kong Med. J. 2011, 17, 381–385. [Google Scholar]
- Undheim, E.A.B.; Fry, B.G.; King, G.F. Centipede Venom: Recent Discoveries and Current State of Knowledge. Toxins 2015, 7, 679–704. [Google Scholar] [CrossRef] [Green Version]
- Undheim, E.A.; Jenner, R.A.; King, G.F. Centipede venoms as a source of drug leads. Expert Opin. Drug Discov. 2016, 11, 1139–1149. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.-C.; Zhang, R.; Zhao, F.; Chen, Z.-M.; Liu, H.-W.; Wang, Y.-J.; Jiang, P.; Zhang, Y.; Wu, Y.; Ding, J.-P.; et al. Venomic and Transcriptomic Analysis of Centipede Scolopendra subspinipes dehaani. J. Proteome Res. 2012, 11, 6197–6212. [Google Scholar] [CrossRef]
- González-Morales, L.; Pedraza-Escalona, M.; Diego-Garcia, E.; Restano-Cassulini, R.; Batista, C.V.F.; Gutiérrez, M.D.C.; Possani, L.D. Proteomic characterization of the venom and transcriptomic analysis of the venomous gland from the Mexican centipede Scolopendra viridis. J. Proteom. 2014, 111, 224–237. [Google Scholar]
- Rong, M.; Yang, S.; Wen, B.; Mo, G.; Kang, D.; Liu, J.; Lin, Z.; Jiang, W.; Li, B.; Du, C.; et al. Peptidomics combined with cDNA library unravel the diversity of centipede venom. J. Proteom. 2015, 114, 28–37. [Google Scholar] [CrossRef]
- Smith, J.J.; Undheim, E.A.B. True Lies: Using Proteomics to Assess the Accuracy of Transcriptome-Based Venomics in Centi-pedes Uncovers False Positives and Reveals Startling Intraspecific Variation in Scolopendra Subspinipes. Toxins 2018, 10, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, M.J.; Rokyta, D.R. Venom-gland transcriptomics and venom proteomics of the giant Florida blue centipede, Scolopendra viridis. Toxicon 2018, 152, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Yoo, W.G.; Lee, J.H.; Shin, Y.; Shim, J.-Y.; Jung, M.; Kang, B.-C.; Oh, J.; Seong, J.; Lee, H.K.; Kong, H.S.; et al. Antimicrobial peptides in the centipede Scolopendra subspinipes mutilans. Funct. Integr. Genom. 2014, 14, 275–283. [Google Scholar] [CrossRef]
- Zhao, F.; Lan, X.; Li, T.; Xiang, Y.; Zhao, F.; Zhang, Y.; Lee, W.-H. Proteotranscriptomic Analysis and Discovery of the Profile and Diversity of Toxin-like Proteins in Centipede. Mol. Cell. Proteom. 2018, 17, 709–720. [Google Scholar] [CrossRef] [Green Version]
- Undheim, E.A.; Sunagar, K.; Hamilton, B.R.; Jones, A.; Venter, D.J.; Fry, B.G.; King, G.F. Multifunctional warheads: Diversification of the toxin arsenal of centipedes via novel multidomain transcripts. J. Proteom. 2014, 102, 1–10. [Google Scholar] [CrossRef]
- Undheim, E.A.; Grimm, L.L.; Low, C.-F.; Morgenstern, D.; Herzig, V.; Zobel-Thropp, P.; Pineda, S.S.; Habib, R.; Dziemborowicz, S.; Fry, B.G.; et al. Weaponization of a Hormone: Convergent Recruitment of Hyperglycemic Hormone into the Venom of Arthropod Predators. Structure 2015, 23, 1283–1292. [Google Scholar] [CrossRef] [Green Version]
- Jenner, R.A.; von Reumont, B.M.; Campbell, L.I.; Undheim, E.A.B. Parallel Evolution of Complex Centipede Venoms Revealed by Comparative Proteotranscriptomic Analyses. Mol. Biol. Evol. 2019, 36, 2748–2763, Erratum in 2021, 38, 4057. [Google Scholar] [CrossRef] [Green Version]
- Undheim, E.A.; Sunagar, K.; Herzig, V.; Kely, L.; Low, D.H.W.; Jackson, T.N.W.; Jones, A.; Kurniawan, N.; King, G.F.; Ali, S.A.; et al. A Proteomics and Transcriptomics Investigation of the Venom from the Barychelid Spider Trittame loki (Brush-Foot Trapdoor). Toxins 2013, 5, 2488–2503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Li, C.; Xiu, C.; Zhang, J.; Li, J.; Huang, L.; Zhang, Y.; Liu, Z. Identification and Biochemical Properties of Two New Acetylcholinesterases in the Pond Wolf Spider (Pardosa pseudoannulata). PLoS ONE 2016, 11, e0158011. [Google Scholar] [CrossRef]
- Alex, A.B.; Deshpande, S.B. Indian red scorpion venom modulates spontaneous activity of rat right atria through the in-volvement of cholinergic and adrenergic systems. Indian J. Exp. Biol. 1999, 37, 455–460. [Google Scholar] [PubMed]
- Krayem, N.; Gargouri, Y. Scorpion venom phospholipases A2: A minireview. Toxicon 2020, 184, 48–54. [Google Scholar] [CrossRef]
- Gonzalez-Morales, L.; Diego-Garcia, E.; Segovia, L.; Gutierrez, M.D.; Possani, L.D. Venom from the centipede Scolo-pendra viridis Say: Purification, gene cloning and phylogenetic analysis of a phospholipase A2. Toxicon 2009, 54, 8–15. [Google Scholar] [CrossRef]
- Li, C.; Wang, F.; Aweya, J.J.; Yao, D.; Zheng, Z.; Huang, H.; Li, S.; Zhang, Y. Trypsin of Litopenaeus vannamei is required for the generation of hemocyanin-derived peptides. Dev. Comp. Immunol. 2018, 79, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Muhlia-Almazán, A.; Sánchez-Paz, A.; García-Carreño, F.L. Invertebrate trypsins: A review. J. Comp. Physiol. B 2008, 178, 655–672. [Google Scholar] [CrossRef]
- Bottrall, J.L.; Madaras, F.; Biven, C.D.; Venning, M.G.; Mirtschin, P.J. Proteolytic activity of Elapid and Viperid Snake venoms and its implication to digestion. J. Venom Res. 2010, 1, 18–28. [Google Scholar] [PubMed]
- Rokyta, D.R.; Ward, M.J. Venom-gland transcriptomics and venom proteomics of the black-back scorpion (Hadrurus spadix) reveal detectability challenges and an unexplored realm of animal toxin diversity. Toxicon 2017, 128, 23–37. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.P.; Ushkaryov, Y.A. The latrophilins, “split-personality” receptors. Adv. Exp. Med. Biol. 2010, 706, 59–75. [Google Scholar] [PubMed]
- Jackson, V.A.; del Toro, D.; Carrasquero, M.; Roversi, P.; Harlos, K.; Klein, R.; Seiradake, E. Structural basis of latro-philin-FLRT interaction. Structure 2015, 23, 774–781. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Guan, X.; Jiang, S.; Yang, J.; Sui, N.; Chen, A.; Kuang, P.; Zhang, X. Effect of batroxobin on expression of neural cell adhesion molecule in temporal infarction rats and spatial learning and memory disorder. J. Tradit. Chin. Med. = Chung i Tsa Chih Ying Wen Pan 2001, 21, 294–298. [Google Scholar]
- Eichberg, S.; Sanz, L.; Calvete, J.J.; Pla, D. Constructing comprehensive venom proteome reference maps for integrative venomics. Expert Rev. Proteom. 2015, 12, 557–573. [Google Scholar] [CrossRef] [PubMed]
- Tavora, B.; Kimura, L.F.; Antoniazzi, M.M.; Chiariello, T.M.; Faquim-Mauro, E.L.; Barbaro, K.C. Involvement of mast cells and histamine in edema induced in mice by Scolopendra viridicornis centipede venom. Toxicon 2016, 121, 51–60. [Google Scholar] [CrossRef]
- Kimura, L.F.; Prezotto-Neto, J.P.; Távora, B.D.C.L.F.; Antoniazzi, M.M.; Knysak, I.; Guizze, S.P.G.; Santoro, M.L.; Barbaro, K.C. Local inflammatory reaction induced by Scolopendra viridicornis centipede venom in mice. Toxicon 2013, 76, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Akdis, C.A.; Blaser, K. Histamine in the immune regulation of allergic inflammation. J. Allergy Clin. Immunol. 2003, 112, 15–22. [Google Scholar] [CrossRef]
- Jutel, M.; Akdis, M.; Akdis, C.A. Histamine, histamine receptors and their role in immune pathology. Clin. Exp. Allergy 2009, 39, 1786–1800. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform ex-traction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Aronesty, E. Command-Line Tools for Processing Biological Sequencing Data; Ea-Utils, Expression Analysis: Durham, NC, USA, 2011. [Google Scholar]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas Nordahl, P.; Søren, B.; Gunnar Von, H.; Henrik, N. SignalP 4.0: Discriminating signal peptides from transmem-brane regions. Nat. Methods 2011, 8, 785. [Google Scholar]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mistry, J.; Finn, R.D.; Eddy, S.R.; Bateman, A.; Punta, M. Challenges in homology search: HMMER3 and convergent evolution of coiled-coil regions. Nucleic Acids Res. 2013, 41, e121. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Number (%) |
|---|---|
| Total Raw Paird-end Reads | 15,904,398 |
| Total High-quality Paird-end Reads | 14,964,551 |
| Total transcripts | 88,774 |
| Percent GC Content | 42.76% |
| Transcript N50 | 1104 |
| Median transcript length | 416 |
| Average transcript length | 766.27 |
| Longest transcript length | 23,855 |
| Number of transcripts >1 kb | 16,266 (18.3%) |
| Shortest transcript length | 209 |
| Total assembled bases | 68,024,656 |
| Complete BUSCOs | 885 (92.7%) |
| Complete and single-copy BUSCOs | 713 (74.7%) |
| Complete and duplicated BUSCOs | 172 (18%) |
| Fragmented BUSCOs | 49 (5.1%) |
| Missing BUSCOs | 20 (2.2%) |
| Total BUSCO groups searched | 954 (100%) |
| Description | Total Transcripts | Number (%) |
|---|---|---|
| Total Cryptops iheringi Hits | - | 5328 (6%) |
| Cryptops anomalans | 33,662 | 4272 |
| Scolopocryptops rubiginosus | 28,965 | 575 |
| Scolopendra cingulata | 23,301 | 283 |
| Scolopocryptops sexspinosus | 1540 | 117 |
| Scolopendra subspinipes | 648 | 32 |
| Scolopendra viridis | 520 | 29 |
| Hemiscolopendra marginata | 764 | 17 |
| Scolopendra morsitans | 662 | 3 |
| Scolopendra alternans | 51 | 0 |
| Scolopendra dehaani | 16,084 | 0 |
| Band Group | Accession Number | Unique Peptides | Proteome Coverage | Molecular Weigth | Best Hit | Species | Identity and Acession Number |
|---|---|---|---|---|---|---|---|
| 1 | Ciheringi01366 | 32 | 53% | 152 kDa | uncharacterized protein | Centruroides sculpturatus | 25.07% XP_023226371.1 |
| 2 | Ciheringi05450 | 78 | 77% | 76 kDa | Hemocyanin | Scolopendra dehaani | 55.99% SMH67860.1 |
| 3 | Ciheringi11581 | 10 | 36% | 37 kDa | No hit | - | - |
| 3 | Ciheringi14246 | 8 | 29% | 27 kDa | Lipase | Centruroides sculpturatus | 29.74% XP_023229615.1 |
| 3 | Ciheringi16405 | 4 | 14% | 28 kDa | Venom allergen | Scolopendra subspinipes | 42.92% QEE04219.1 |
| 3 | Ciheringi21566 | 5 | 34% | 22 kDa | No hit | - | - |
| 4 | Ciheringi38643 | 12 | 56% | 11 kDa | No hit | - | - |
| 4 | Ciheringi10323 | 4 | 53% | 10 kDa | Lipase | Branchiostoma floridae | 37.39% XP_035699465.1 |
| 5 | Ciheringi24930 | 3 | 22% | 15 kDa | No hit | - | - |
| 5 | Cryptoxin-1 | 12 | 68% | 12 kDa | No hit | - | - |
| 6 | Ciheringi05125 | 7 | 74% | 14 kDa | Profilin | Orussus abietinus | 80.95% XP_012283556.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Lucca Caetano, L.H.; Nishiyama-Jr, M.Y.; de Carvalho Lins Fernandes Távora, B.; de Oliveira, U.C.; de Loiola Meirelles Junqueira-de-Azevedo, I.; Faquim-Mauro, E.L.; Magalhães, G.S. Recombinant Production and Characterization of a New Toxin from Cryptops iheringi Centipede Venom Revealed by Proteome and Transcriptome Analysis. Toxins 2021, 13, 858. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13120858
De Lucca Caetano LH, Nishiyama-Jr MY, de Carvalho Lins Fernandes Távora B, de Oliveira UC, de Loiola Meirelles Junqueira-de-Azevedo I, Faquim-Mauro EL, Magalhães GS. Recombinant Production and Characterization of a New Toxin from Cryptops iheringi Centipede Venom Revealed by Proteome and Transcriptome Analysis. Toxins. 2021; 13(12):858. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13120858
Chicago/Turabian StyleDe Lucca Caetano, Lhiri Hanna, Milton Yutaka Nishiyama-Jr, Bianca de Carvalho Lins Fernandes Távora, Ursula Castro de Oliveira, Inácio de Loiola Meirelles Junqueira-de-Azevedo, Eliana L. Faquim-Mauro, and Geraldo Santana Magalhães. 2021. "Recombinant Production and Characterization of a New Toxin from Cryptops iheringi Centipede Venom Revealed by Proteome and Transcriptome Analysis" Toxins 13, no. 12: 858. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13120858