Natural Occurrence of Mycotoxin-Producing Fusaria in Market-Bought Peruvian Cereals: A Food Safety Threat for Andean Populations

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
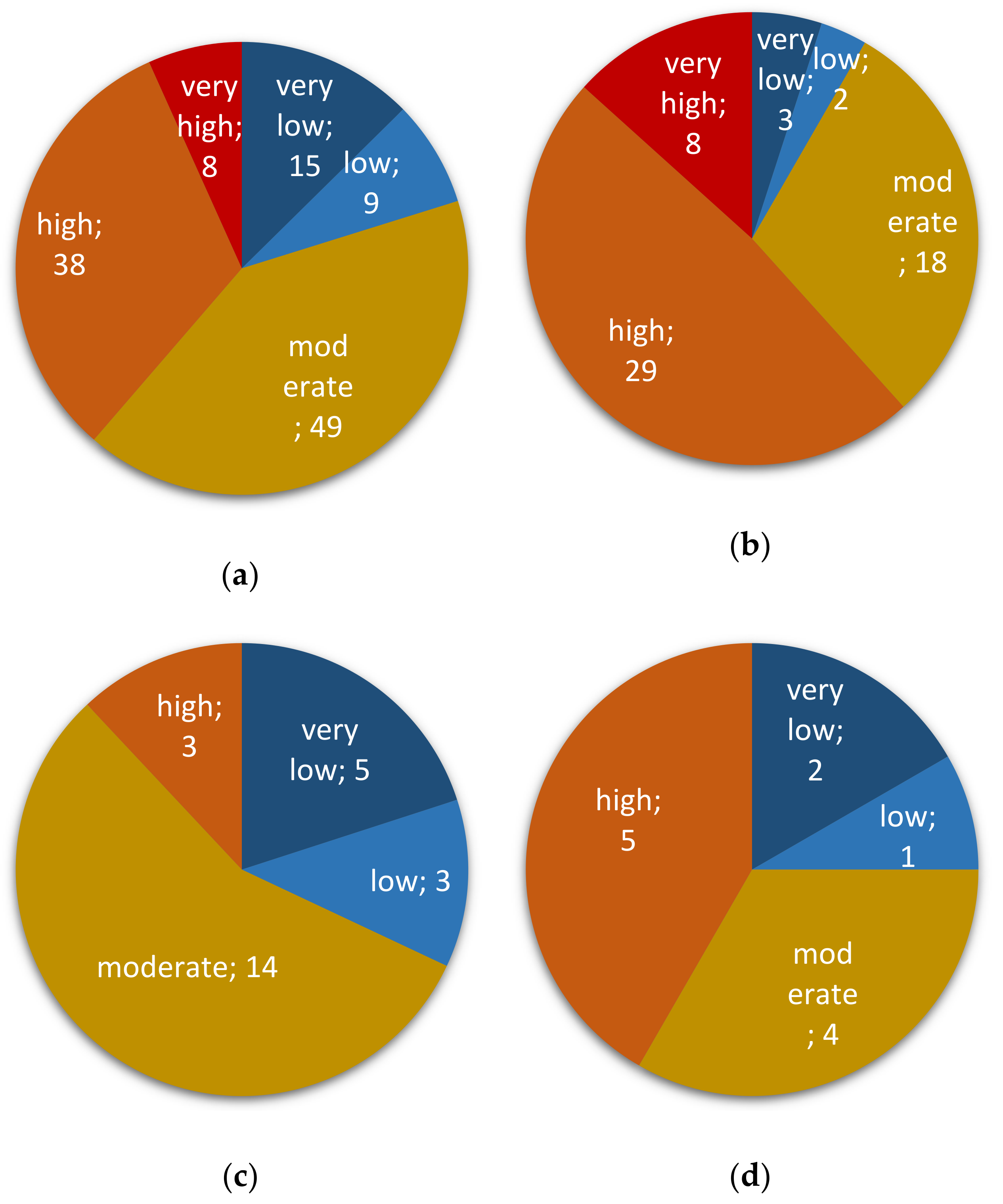
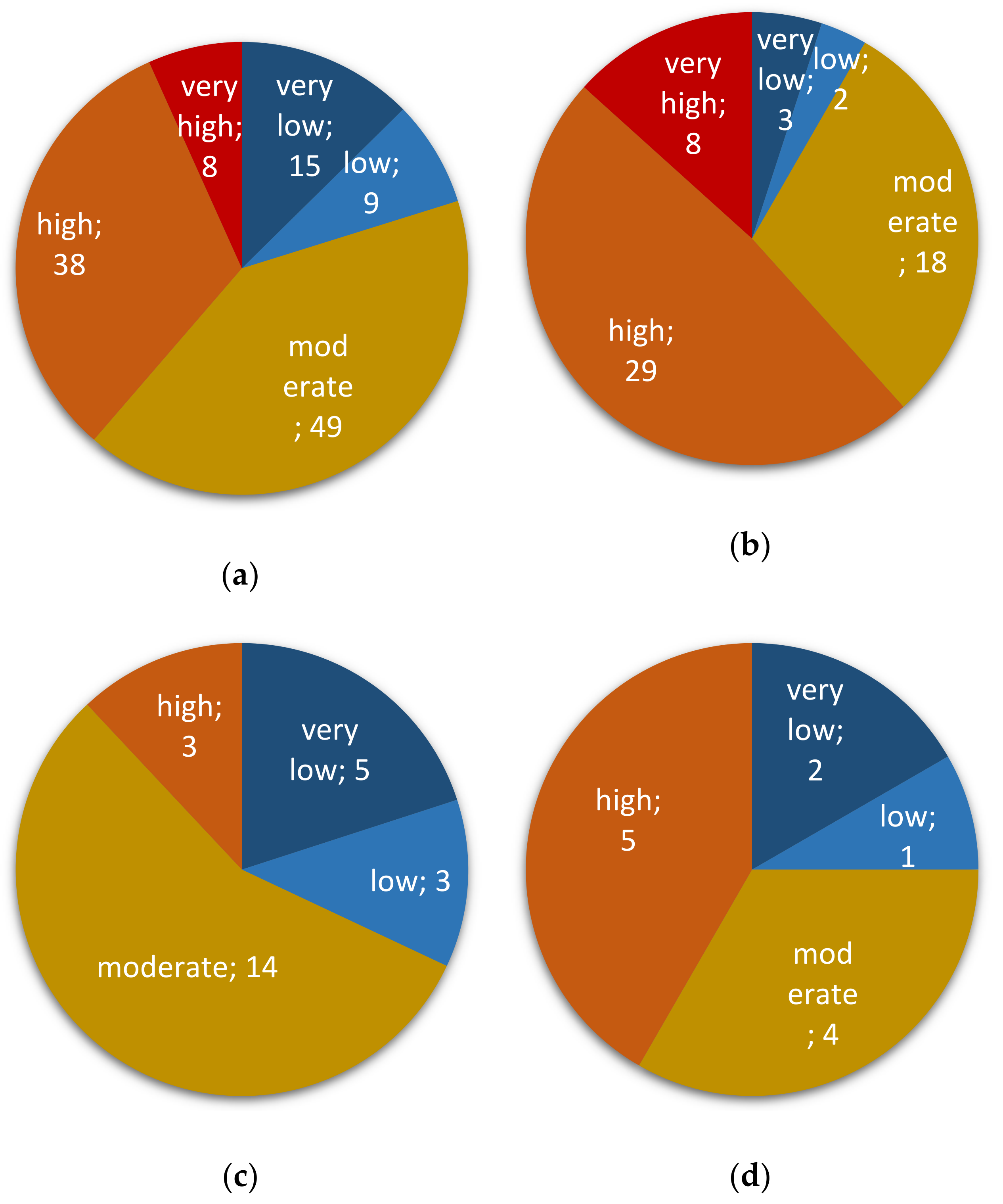
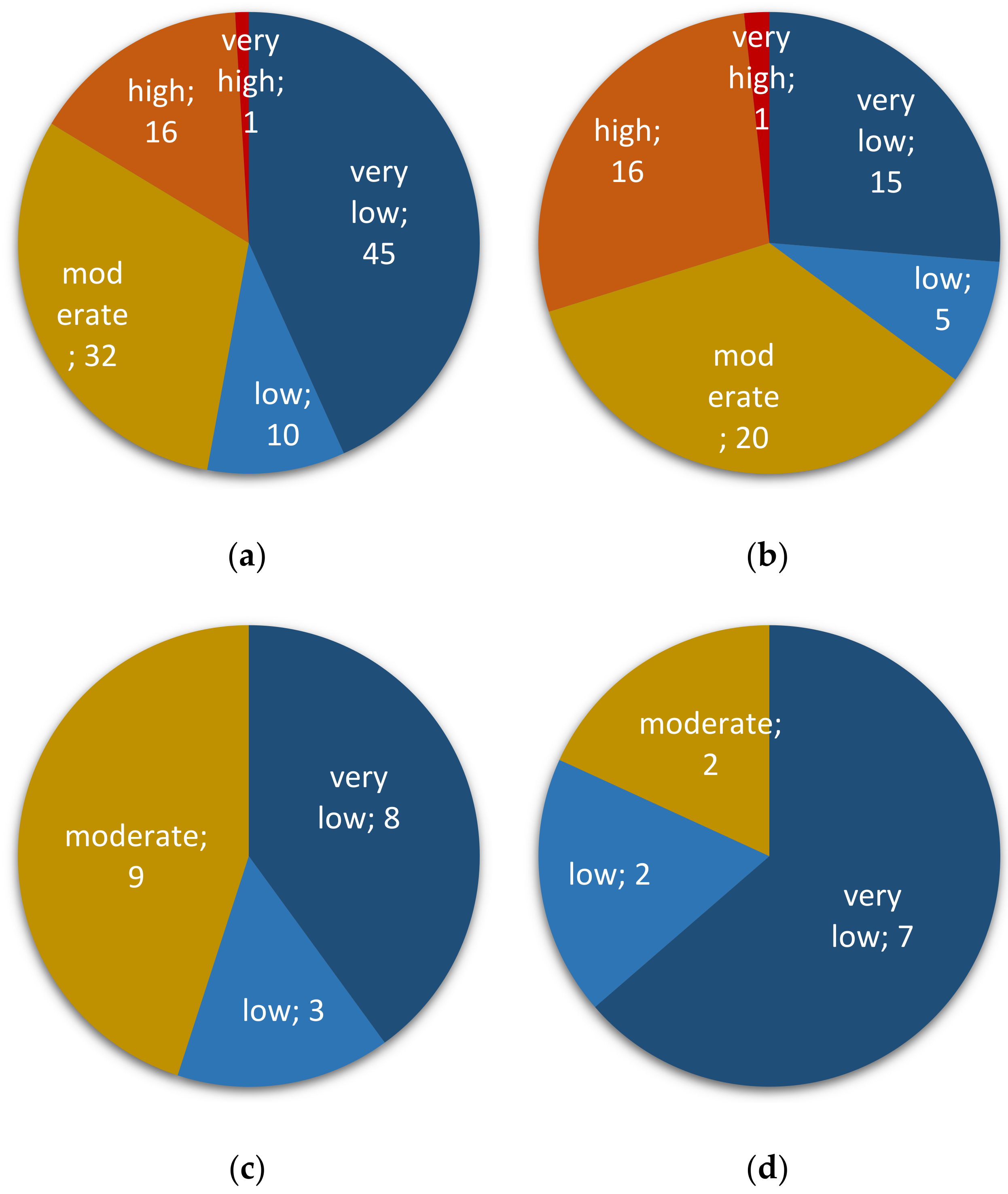
2.1. Occurrence of Fusarium spp. in Peruvian Cereals Destined for Direct Consumption
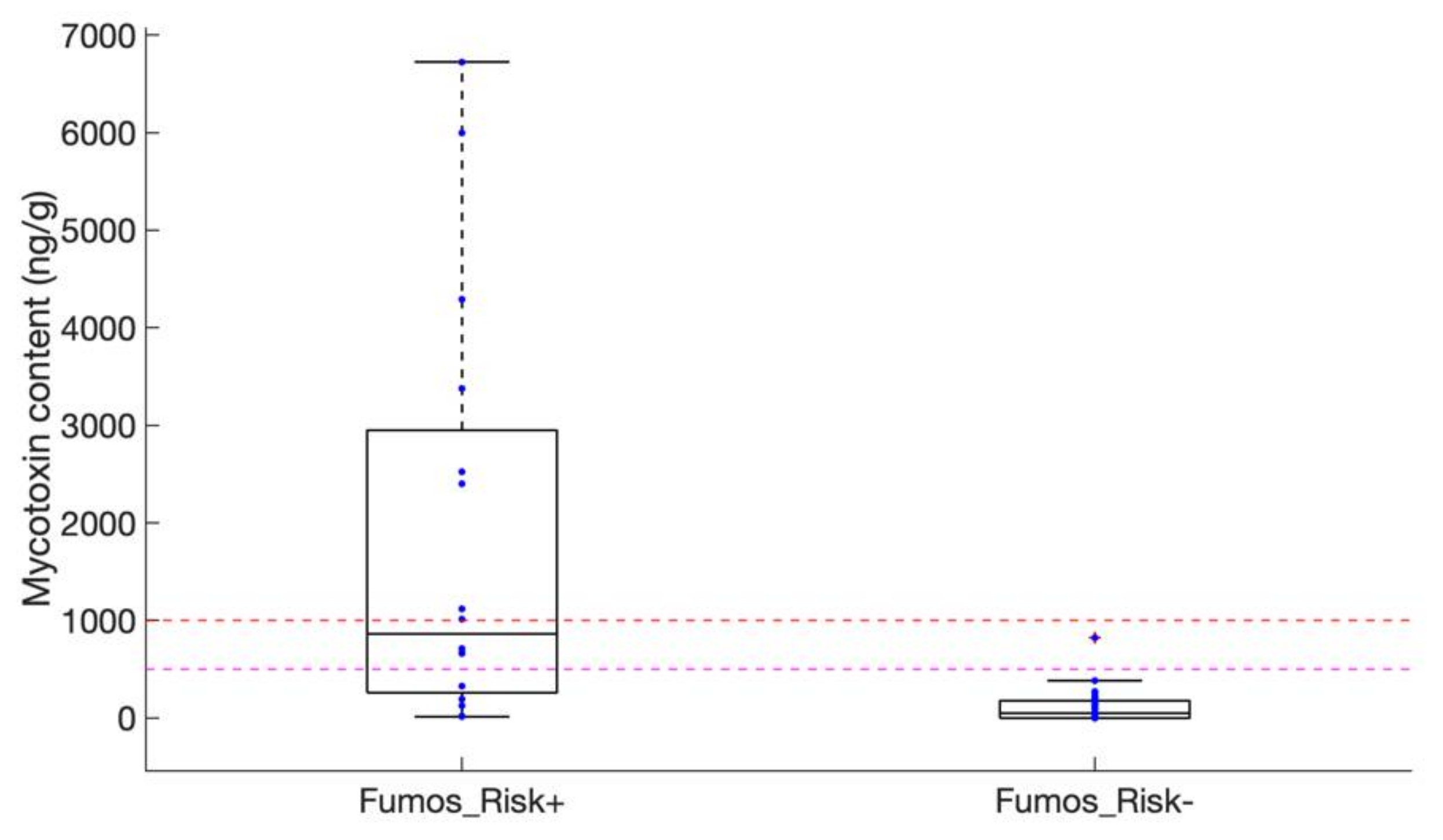
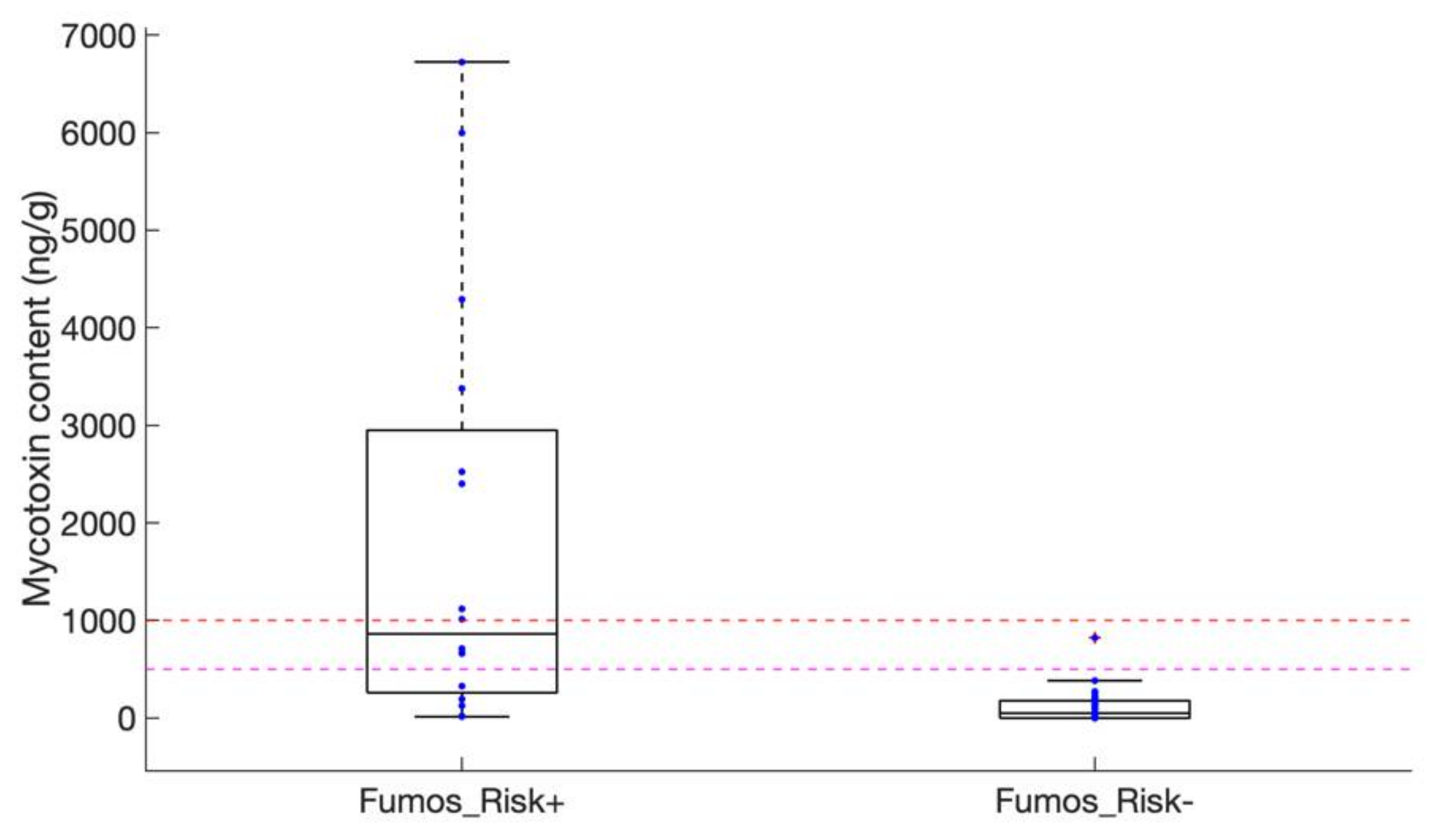
2.2. Assessing Contaminations with Fumonisins and Fumonisin-Producing Fusarium Species in Crop Samples
2.3. Assessing Contaminations with Trichothecenes and Zearalenone as well as Their Corresponding Producing Fusarium species in Crop Samples
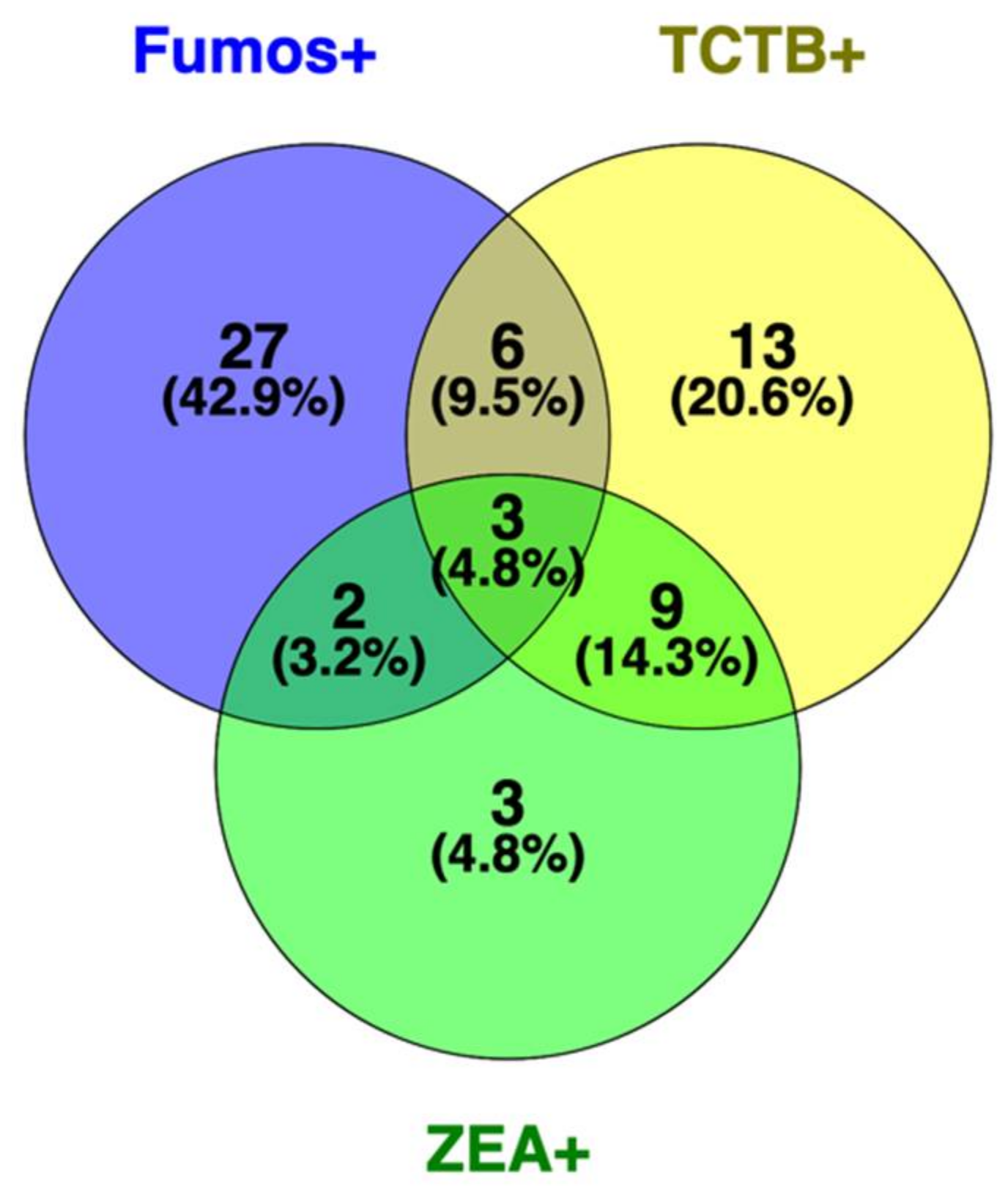
2.4. Frequencies of Co-Contaminations with Fusarium Species and The Mycotoxins Fumonisins, Trichothecenes, and Zearalenone
3. Discussion
4. Materials and Methods
4.1. Access to Genetic Resources and Benefit-Sharing
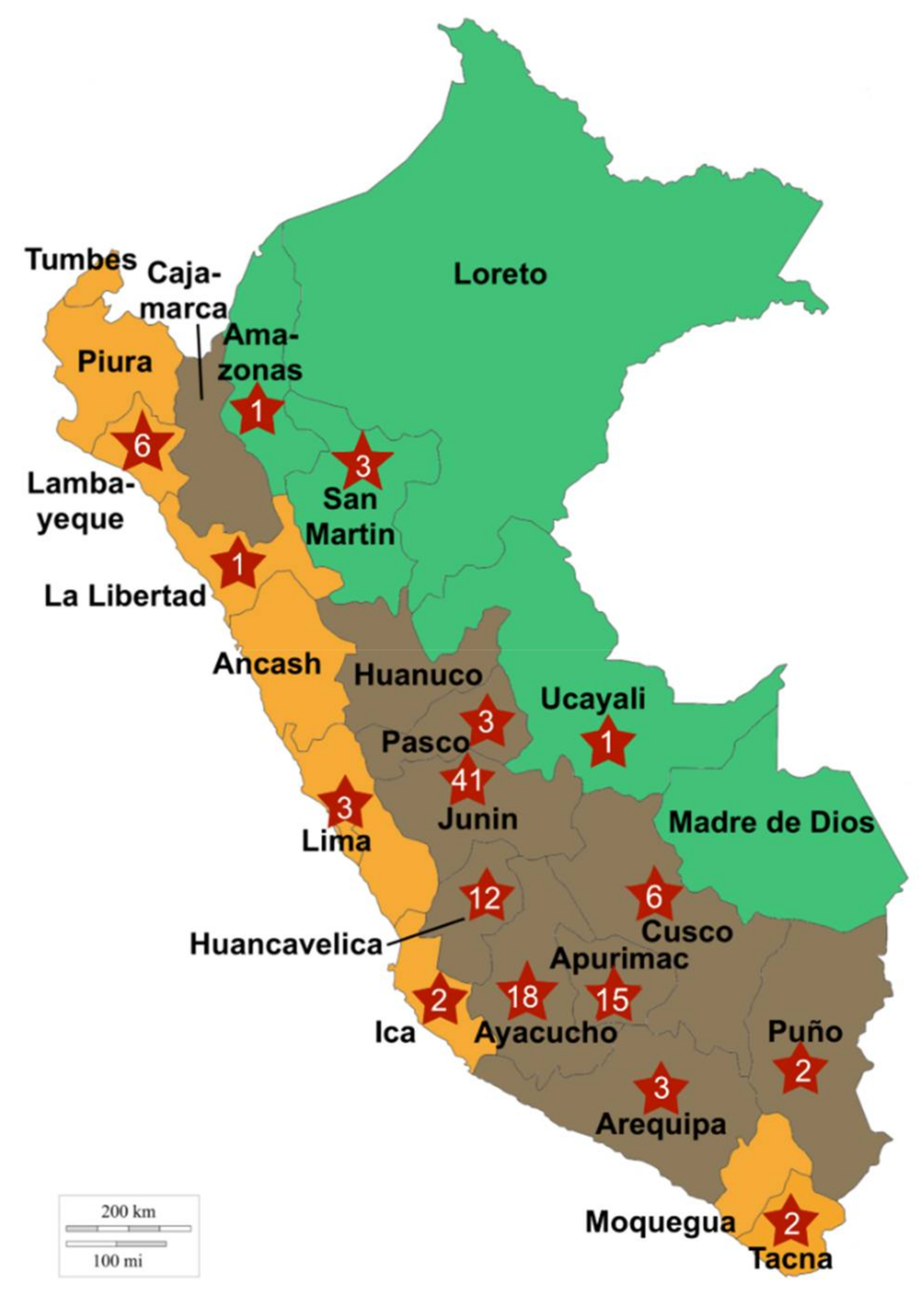
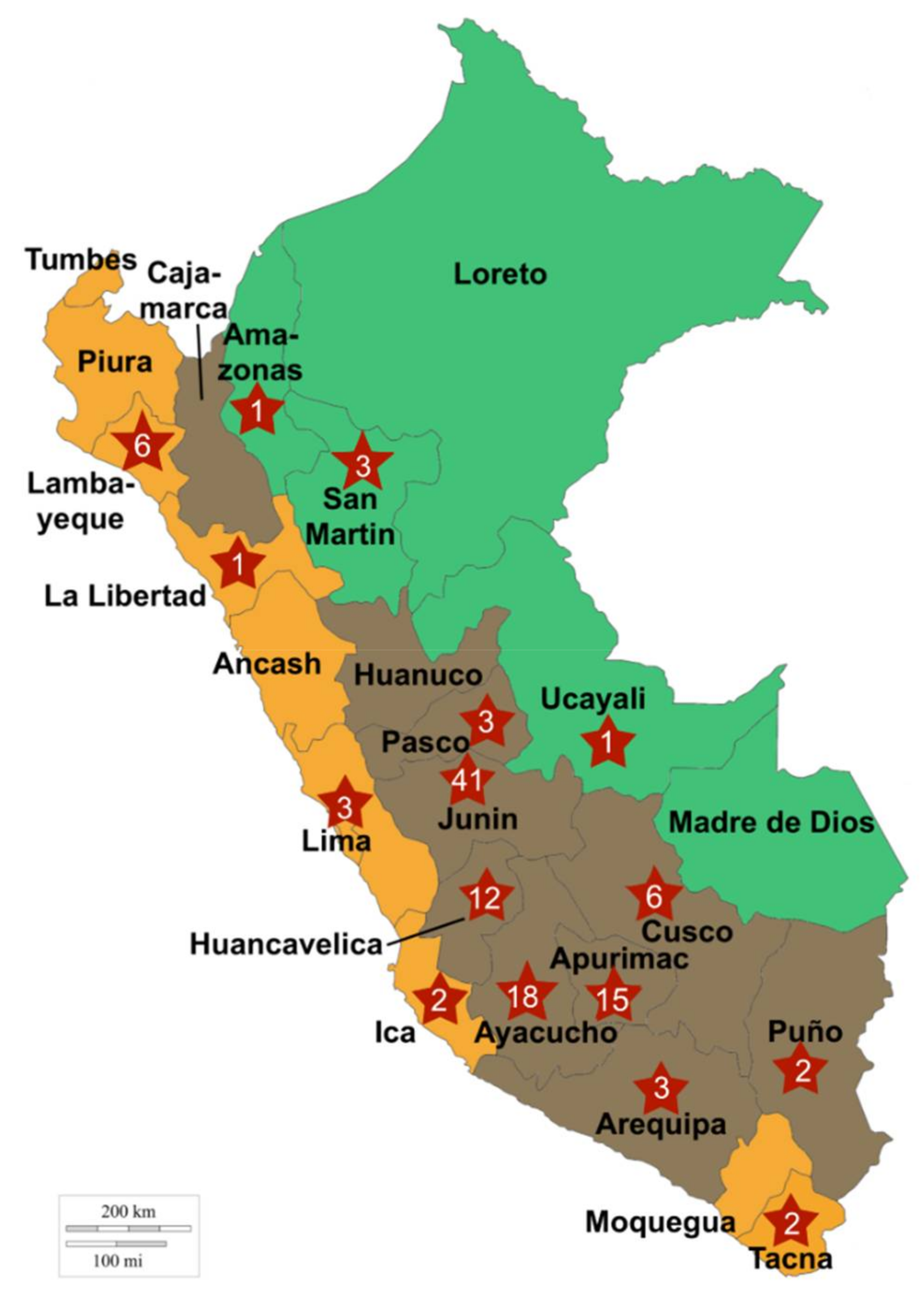
4.2. Experimental Design and Sampling
4.3. DNA Extraction
4.4. Fusarium Contaminations and Mycotoxin Risk Assessment by qPCR Assays
4.5. Mycotoxin Content Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R. Worldwide Contamination of Food-Crops with Mycotoxins: Validity of the Widely Cited ‘FAO Estimate’ of 25%. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef]
- BIOMIN. World Mycotoxin Survey the Global Threat January September 2020. Available online: https://ahead.biomin.net/hubfs/Downloads/REP_MTXsurvey_Q3_2020_EN.pdf (accessed on 4 January 2021).
- Braun, M.S.; Wink, M. Exposure, Occurrence, and Chemistry of Fumonisins and Their Cryptic Derivatives. Compr. Rev. Food Sci. Food Saf. 2018, 17, 769–791. [Google Scholar] [CrossRef] [Green Version]
- Osborne, L.E.; Stein, J.M. Epidemiology of Fusarium Head Blight on Small-Grain Cereals. Int. J. Food Microbiol. 2007, 119, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Wegulo, S.N. Factors Influencing Deoxynivalenol Accumulation in Small Grain Cereals. Toxins 2012, 4, 1157–1180. [Google Scholar] [CrossRef]
- Ropejko, K.; Twarużek, M. Zearalenone and Its Metabolites—General Overview, Occurrence, and Toxicity. Toxins 2021, 13, 35. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wei, Z.; Wang, Y.; Long, M.; Wu, W.; Kuca, K. Fumonisin B1: Mechanisms of Toxicity and Biological Detoxification Progress in Animals. Food Chem. Toxicol. 2021, 111977. [Google Scholar] [CrossRef] [PubMed]
- Foroud, N.A.; Baines, D.; Gagkaeva, T.Y.; Thakor, N.; Badea, A.; Steiner, B.; Bürstmayr, M.; Bürstmayr, H. Trichothecenes in Cereal Grains An Update. Toxins 2019, 11, 634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.; Srivastava, S.; Dewangan, J.; Divakar, A.; Rath, S.K. Global Occurrence of Deoxynivalenol in Food Commodities and Exposure Risk Assessment in Humans in the Last Decade: A Survey. Crit. Rev. Food Sci. Nutr. 2020, 60, 1346–1374. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Cui, L.; Sajid, A.; Zainab, F.; Wu, Q.; Wang, X.; Yuan, Z. The Epigenetic Mechanisms in Fusarium Mycotoxins Induced Toxicities. Food Chem. Toxicol. 2019, 123, 595–601. [Google Scholar] [CrossRef]
- Janik, E.; Niemcewicz, M.; Ceremuga, M.; Stela, M.; Saluk-Bijak, J.; Siadkowski, A.; Bijak, M. Molecular Aspects of Mycotoxins—A Serious Problem for Human Health. Int. J. Mol. Sci. 2020, 21, 8187. [Google Scholar] [CrossRef]
- Raj, J.; Farkas, H.; Jakovcevic, Z.; Todorovic, N.; Bosnjak-Neumuller, J.; Vasiljevic, M. Multiple Mycotoxins Detected in Corn Harvested in 2019; PATENT CO. DOO.: Misicevo, Serbia, 2020. [Google Scholar]
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global Mycotoxin Occurrence in Feed: A Ten-Year Survey. Toxins 2019, 11, 375. [Google Scholar] [CrossRef] [Green Version]
- Ellis, E.C.; Ramankutty, N. Putting People in the Map: Anthropogenic Biomes of the World. Front. Ecol. Environ. 2008, 6, 439–447. [Google Scholar] [CrossRef] [Green Version]
- Peru Agriculture Opportunities ASA. Gaining Momentum in Peruvian Agriculture: Opportunities to Increase Productivity and Enhance Competitiveness; World Bank: Washington, DC, USA, 2017; p. 150. [Google Scholar]
- Ramirez-Hernandez, A.; Galagarza, O.A.; Álvarez Rodriguez, M.V.; Pachari Vera, E.; Valdez Ortiz, M.D.C.; Deering, A.J.; Oliver, H.F. Food Safety in Peru: A Review of Fresh Produce Production and Challenges in the Public Health System. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3323–3342. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E.; Plattner, R.D.; Proctor, R.H. Genetic and Biochemical Aspects of Fumonisin Production. Adv. Exp. Med. Biol. 1996, 392, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Logrieco, A.; Moretti, A.; Altomare, C.; Bottalico, A.; Carbonell Torres, E. Occurrence and Toxicity of Fusarium Subglutinans from Peruvian Maize. Mycopathologia 1993, 122, 185–190. [Google Scholar] [CrossRef]
- Merhej, J.; Richard-Forget, F.; Barreau, C. Regulation of Trichothecene Biosynthesis in Fusarium: Recent Advances and New Insights. Appl. Microbiol. Biotechnol. 2011, 91, 519–528. [Google Scholar] [CrossRef]
- Laraba, I.; McCormick, S.P.; Vaughan, M.M.; Geiser, D.M.; O’Donnell, K. Phylogenetic Diversity, Trichothecene Potential, and Pathogenicity within Fusarium Sambucinum Species Complex. PLoS ONE 2021, 16, e0245037. [Google Scholar] [CrossRef]
- Oliveros, J.C. Venny. An Interactive Tool for Comparing Lists with Venn’s Diagrams. (2007–2015). Available online: https://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 6 November 2020).
- Food and Agriculture Organization/World Health Organization Expert Committee on Food Additives. Evaluation of Certain Food Additives and Contaminants. In World Health Organization Technical Report Series; World Health Organization: Geneva, Switzerland, 2001; Volume 901, pp. 1–107. [Google Scholar]
- World Health Organization. Evaluation of Certain Contaminants in Food. In World Health Organization Technical Report Series; World Health Organization: Geneva, Switzerland, 2011; pp. 1–105. [Google Scholar]
- World Health Organization. Evaluation of Certain Contaminants in Food. In World Health Organization Technical Report Series; World Health Organization: Geneva, Switzerland, 2017; pp. 1–166. [Google Scholar]
- Burger, H.-M.; Lombard, M.J.; Shephard, G.S.; Rheeder, J.R.; van der Westhuizen, L.; Gelderblom, W.C.A. Dietary Fumonisin Exposure in a Rural Population of South Africa. Food Chem. Toxicol. 2010, 48, 2103–2108. [Google Scholar] [CrossRef]
- Torres, O.; Matute, J.; Gelineau-van Waes, J.; Maddox, J.R.; Gregory, S.G.; Ashley-Koch, A.E.; Showker, J.L.; Zitomer, N.C.; Voss, K.A.; Riley, R.T. Urinary Fumonisin B1 and Estimated Fumonisin Intake in Women from High- and Low-Exposure Communities in Guatemala. Mol. Nutr. Food Res. 2014, 58, 973–983. [Google Scholar] [CrossRef]
- Wild, C.P.; Gong, Y.Y. Mycotoxins and Human Disease: A Largely Ignored Global Health Issue. Carcinogenesis 2010, 31, 71–82. [Google Scholar] [CrossRef]
- Coloma, Z.N.; Oliveira, M.S.; Dilkin, P.; Mallmann, A.O.; Almeida, C.A.A.; Mallmann, C.A. Mycotoxin Occurrence in Peruvian Purple Maize. World Mycotoxin J. 2019, 12, 307–315. [Google Scholar] [CrossRef]
- Rychlik, M.; Humpf, H.-U.; Marko, D.; Dänicke, S.; Mally, A.; Berthiller, F.; Klaffke, H.; Lorenz, N. Proposal of a Comprehensive Definition of Modified and Other Forms of Mycotoxins Including “Masked” Mycotoxins. Mycotoxin Res. 2014, 30, 197–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez Tuppia, C.; Atanasova-Penichon, V.; Chéreau, S.; Ferrer, N.; Marchegay, G.; Savoie, J.-M.; Richard-Forget, F. Yeast and Bacteria from Ensiled High Moisture Maize Grains as Potential Mitigation Agents of Fumonisin B1. J. Sci. Food Agric. 2017, 97, 2443–2452. [Google Scholar] [CrossRef]
- Bertani, S.; Pineau, P.; Loli, S.; Moura, J.; Zimic, M.; Deharo, E.; Ruiz, E. An Atypical Age-Specific Pattern of Hepatocellular Carcinoma in Peru: A Threat for Andean Populations. PLoS ONE 2013, 8, e67756. [Google Scholar] [CrossRef]
- Debes, J.D.; Chan, A.J.; Balderramo, D.; Kikuchi, L.; Gonzalez Ballerga, E.; Prieto, J.E.; Tapias, M.; Idrovo, V.; Davalos, M.B.; Cairo, F.; et al. Hepatocellular Carcinoma in South America: Evaluation of Risk Factors, Demographics and Therapy. Liver Int. 2018, 38, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Cano, L.; Cerapio, J.P.; Ruiz, E.; Marchio, A.; Turlin, B.; Casavilca, S.; Taxa, L.; Marti, G.; Deharo, E.; Pineau, P.; et al. Liver Clear Cell Foci and Viral Infection Are Associated with Non-Cirrhotic, Non-Fibrolamellar Hepatocellular Carcinoma in Young Patients from South America. Sci. Rep. 2018, 8, 9945. [Google Scholar] [CrossRef]
- Marchio, A.; Bertani, S.; Rojas Rojas, T.; Doimi, F.; Terris, B.; Deharo, E.; Dejean, A.; Ruiz, E.; Pineau, P. A Peculiar Mutation Spectrum Emerging from Young Peruvian Patients with Hepatocellular Carcinoma. PLoS ONE 2014, 9, e114912. [Google Scholar] [CrossRef] [Green Version]
- Thoolen, B.; Ten Kate, F.J.W.; van Diest, P.J.; Malarkey, D.E.; Elmore, S.A.; Maronpot, R.R. Comparative Histomorphological Review of Rat and Human Hepatocellular Proliferative Lesions. J. Toxicol. Pathol. 2012, 25, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Hailey, J.R.; Walker, N.J.; Sells, D.M.; Brix, A.E.; Jokinen, M.P.; Nyska, A. Classification of Proliferative Hepatocellular Lesions in Harlan Sprague-Dawley Rats Chronically Exposed to Dioxin-like Compounds. Toxicol. Pathol. 2005, 33, 165–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debes, J.D. Early Hepatocellular Carcinoma in South America: What Is to Blame? Liver Int. 2018, 38, 957–958. [Google Scholar] [CrossRef]
- Pineau, P.; Ruiz, E.; Deharo, E.; Bertani, S. On Hepatocellular Carcinoma in South America and Early-Age Onset of the Disease. Clin. Res. Hepatol. Gastroenterol. 2019, 43, 522–526. [Google Scholar] [CrossRef]
- Lemmer, E.R.; de la Motte Hall, P.; Omori, N.; Omori, M.; Shephard, E.G.; Gelderblom, W.C.; Cruse, J.P.; Barnard, R.A.; Marasas, W.F.; Kirsch, R.E.; et al. Histopathology and Gene Expression Changes in Rat Liver during Feeding of Fumonisin B1, a Carcinogenic Mycotoxin Produced by Fusarium Moniliforme. Carcinogenesis 1999, 20, 817–824. [Google Scholar] [CrossRef] [Green Version]
- Scauflaire, J.; Gourgue, M.; Munaut, F. Fusarium Temperatum Sp. Nov. from Maize, an Emergent Species Closely Related to Fusarium Subglutinans. Mycologia 2011, 103, 586–597. [Google Scholar] [CrossRef] [Green Version]
- Scauflaire, J.; Gourgue, M.; Callebaut, A.; Munaut, F. Fusarium Temperatum, a Mycotoxin-Producing Pathogen of Maize. Eur. J. Plant Pathol. 2012, 133, 911–922. [Google Scholar] [CrossRef]
- Tagele, S.B.; Kim, S.W.; Lee, H.G.; Lee, Y.S. Aggressiveness and Fumonisins Production of Fusarium Subglutinans and Fusarium Temperatum on Korean Maize Cultivars. Agronomy 2019, 9, 88. [Google Scholar] [CrossRef] [Green Version]
- Vaclavikova, M.; Malachova, A.; Veprikova, Z.; Dzuman, Z.; Zachariasova, M.; Hajslova, J. ‘Emerging’ Mycotoxins in Cereals Processing Chains: Changes of Enniatins during Beer and Bread Making. Food Chem. 2013, 136, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Fumero, M.V.; Reynoso, M.M.; Chulze, S. Fusarium Temperatum and Fusarium Subglutinans Isolated from Maize in Argentina. Int. J. Food Microbiol. 2015, 199, 86–92. [Google Scholar] [CrossRef]
- Fumero, M.V.; Villani, A.; Susca, A.; Haidukowski, M.; Cimmarusti, M.T.; Toomajian, C.; Leslie, J.F.; Chulze, S.N.; Moretti, A. Fumonisin and Beauvericin Chemotypes and Genotypes of the Sister Species Fusarium Subglutinans and Fusarium Temperatum. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef]
- Heino, M.; Puma, M.J.; Ward, P.J.; Gerten, D.; Heck, V.; Siebert, S.; Kummu, M. Two-Thirds of Global Cropland Area Impacted by Climate Oscillations. Nat. Commun. 2018, 9, 1257. [Google Scholar] [CrossRef] [PubMed]
- Bruns, H.A. Controlling Aflatoxin and Fumonisin in Maize by Crop Management. J. Toxicol. Toxin Rev. 2003, 22, 153–173. [Google Scholar] [CrossRef]
- Rojas Rojas, T.; Bourdy, G.; Ruiz, E.; Cerapio, J.-P.; Pineau, P.; Gardon, J.; Doimi, F.; Deparis, X.; Deharo, E.; Bertani, S. Herbal Medicine Practices of Patients with Liver Cancer in Peru: A Comprehensive Study Toward Integrative Cancer Management. Integr. Cancer 2018, 17, 52–64. [Google Scholar] [CrossRef] [Green Version]
- Glass, N.L.; Donaldson, G.C. Development of Primer Sets Designed for Use with the PCR to Amplify Conserved Genes from Filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Pinson-Gadais, L.; Barreau, C.; Chaurand, M.; Gregoire, S.; Monmarson, M.; Richard-Forget, F. Distribution of Toxigenic Fusarium Spp. and Mycotoxin Production in Milling Fractions of Durum Wheat. Food Addit. Contam. 2007, 24, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Doohan, F.M.; Weston, G.; Rezanoor, H.N.; Parry, D.W.; Nicholson, P. Development and Use of a Reverse Transcription-PCR Assay to Study Expression of Tri5 by Fusarium Species in Vitro and in Planta. Appl. Environ. Microbiol. 1999, 65, 3850–3854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picot, A.; Hourcade-Marcolla, D.; Barreau, C.; Pinson-Gadais, L.; Caron, D.; Richard-Forget, F.; Lannou, C. Interactions between Fusarium Verticillioides and Fusarium Graminearum in Maize Ears and Consequences for Fungal Development and Mycotoxin Accumulation. Plant Pathol. 2012, 61, 140–151. [Google Scholar] [CrossRef]
- Aguado, A.; Savoie, J.-M.; Chéreau, S.; Ducos, C.; Aguilar, M.; Ferrer, N.; Aguilar, M.; Pinson-Gadais, L.; Richard-Forget, F. Priming to Protect Maize from Fusarium Verticillioides and Its Fumonisin Accumulation. J. Sci. Food Agric. 2019, 99, 64–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, P.; Simpson, D.R.; Weston, G.; Rezanoor, H.N.; Lees, A.K.; Parry, D.W.; Joyce, D. Detection and Quantification of Fusarium Culmorum and Fusarium Graminearumin Cereals Using PCR Assays. Physiol. Mol. Plant Pathol. 1998, 53, 17–37. [Google Scholar] [CrossRef]
- Aoki, T.; O’Donnell, K. Morphological and Molecular Characterization of Fusarium Pseudograminearum Sp. Nov., Formerly Recognized as the Group 1 Population of F. Graminearum. Mycologia 1999, 91, 597–609. [Google Scholar] [CrossRef]
- Picot, A.; Barreau, C.; Pinson-Gadais, L.; Piraux, F.; Caron, D.; Lannou, C.; Richard-Forget, F. The Dent Stage of Maize Kernels Is the Most Conducive for Fumonisin Biosynthesis under Field Conditions. Appl. Environ. Microbiol. 2011, 77, 8382–8390. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, J.L.; Nielsen, K.F.; Rasmussen, P.H.; Thrane, U. Development of a LC-MS/MS Method for the Analysis of Enniatins and Beauvericin in Whole Fresh and Ensiled Maize. J. Agric. Food Chem. 2008, 56, 10439–10443. [Google Scholar] [CrossRef] [PubMed]
- Sulyok, M.; Berthiller, F.; Krska, R.; Schuhmacher, R. Development and Validation of a Liquid Chromatography/Tandem Mass Spectrometric Method for the Determination of 39 Mycotoxins in Wheat and Maize. Rapid Commun. Mass Spectrom. 2006, 20, 2649–2659. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Grain | Number of Samples | Landscape of Origin | ||
|---|---|---|---|---|
| Costa | Sierra | Selva | ||
| corn | 60 | 7 | 51 | 2 |
| wheat | 25 | 1 | 24 | 0 |
| amaranth | 12 | 0 | 12 | 0 |
| rice | 12 | 4 | 5 | 3 |
| barley | 6 | 0 | 6 | 0 |
| oat | 4 | 2 | 2 | 0 |
| Total | 119 | 14 | 100 | 5 |
| Primer Names | Primer Sequences | Ta 1 (°C) | Fusarium Species/Toxin Producers Targeted | References |
|---|---|---|---|---|
| ßtub2a ßt-2 | GGTAACCAAATCGGTGCTGCTTTC GATTGACCGAAAACGAAGTTG | 58 | Broad range 2 | [49,50,51] |
| Fum1-F Fum1-R | GGATTGGCTGGATCTTCACAG GAAGATGGCATTGATTGCCTC | 57 | Fumonisins 3 | [52] |
| Tri5-TCTA-F Tri5-TCTA-R | CTATTCCTTGAGATTACAT CCTTGTAGAATGACATAAGA | 58 | TCTA 4 | This paper |
| Tri5-TCTB-F Tri5-TCTB-R | GATGGACACGATTGAGTG GCTCAAAGAACTTGCAGA | 58 | TCTB 5 | This paper |
| Fum1-656F Fum1-1158R | CGGTTGTTCATCATCTCTGA GCTCCCGATGTAGAGCTTCTT | 60 | F. verticillioides | [53] |
| Fum1-656F Fum1-872R | TGCTCGTCATCCCTGATAG GAAGATGGCATTGATTGCCTC | 60 | F. proliferatum | [52] |
| Fg16N-F Fg16N-R | ACAGATGACAAGATTCAGGCACA TTCTTTGACATCTGTTCAACCCA | 60 | F. graminearum | [54] |
| FC01-F FC01-R | ATGGTGAACTCGTCGTGGC CCCTTCTTACGCCAATCTCG | 60 | F. culmorum | [54] |
| FcroA-F FcroA-R | CTCAGTGTCCACCGCGTTGCGTAG CTCAGTGTCCCAATCAAATAGTCC | 62 | F. cerealis | [54] |
| Fp1-F Fp1-R | CGGGGTAGTTTCACATTTC(C/T)G GAGAATGTGATGA(C/G)GACAATA | 57 | F. pseudograminearum | [55] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ducos, C.; Pinson-Gadais, L.; Chereau, S.; Richard-Forget, F.; Vásquez-Ocmín, P.; Cerapio, J.P.; Casavilca-Zambrano, S.; Ruiz, E.; Pineau, P.; Bertani, S.; et al. Natural Occurrence of Mycotoxin-Producing Fusaria in Market-Bought Peruvian Cereals: A Food Safety Threat for Andean Populations. Toxins 2021, 13, 172. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020172
Ducos C, Pinson-Gadais L, Chereau S, Richard-Forget F, Vásquez-Ocmín P, Cerapio JP, Casavilca-Zambrano S, Ruiz E, Pineau P, Bertani S, et al. Natural Occurrence of Mycotoxin-Producing Fusaria in Market-Bought Peruvian Cereals: A Food Safety Threat for Andean Populations. Toxins. 2021; 13(2):172. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020172
Chicago/Turabian StyleDucos, Christine, Laetitia Pinson-Gadais, Sylvain Chereau, Florence Richard-Forget, Pedro Vásquez-Ocmín, Juan Pablo Cerapio, Sandro Casavilca-Zambrano, Eloy Ruiz, Pascal Pineau, Stéphane Bertani, and et al. 2021. "Natural Occurrence of Mycotoxin-Producing Fusaria in Market-Bought Peruvian Cereals: A Food Safety Threat for Andean Populations" Toxins 13, no. 2: 172. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020172