Influence of H2O2-Induced Oxidative Stress on In Vitro Growth and Moniliformin and Fumonisins Accumulation by Fusarium proliferatum and Fusarium subglutinans

 , and
, and
Abstract
:1. Introduction
2. Results
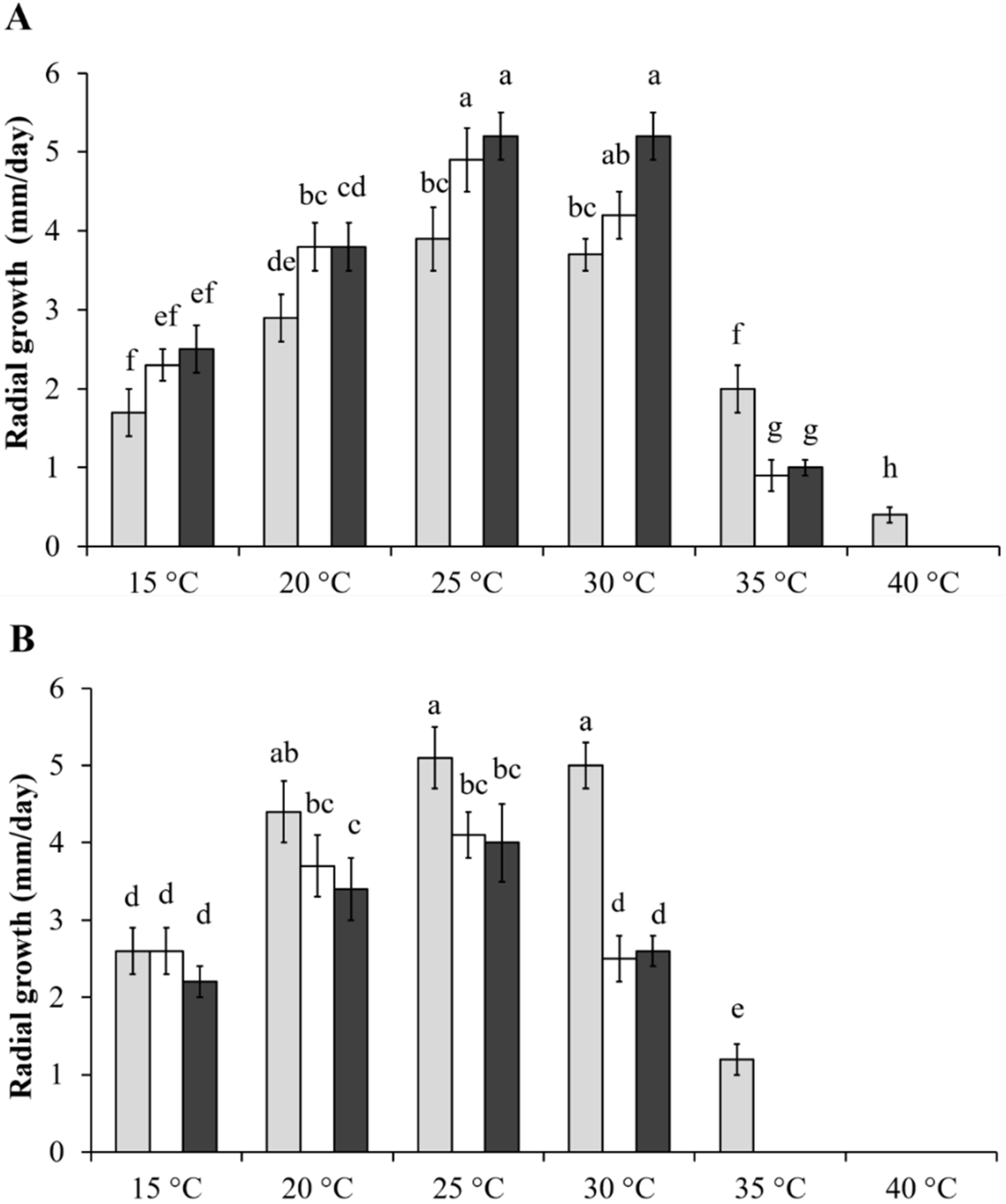
2.1. Effect of Temperature on Fusarium Growth
2.2. Effect of Hydrogen Peroxide Treatments on Fungal Biomass and Mycotoxin Content
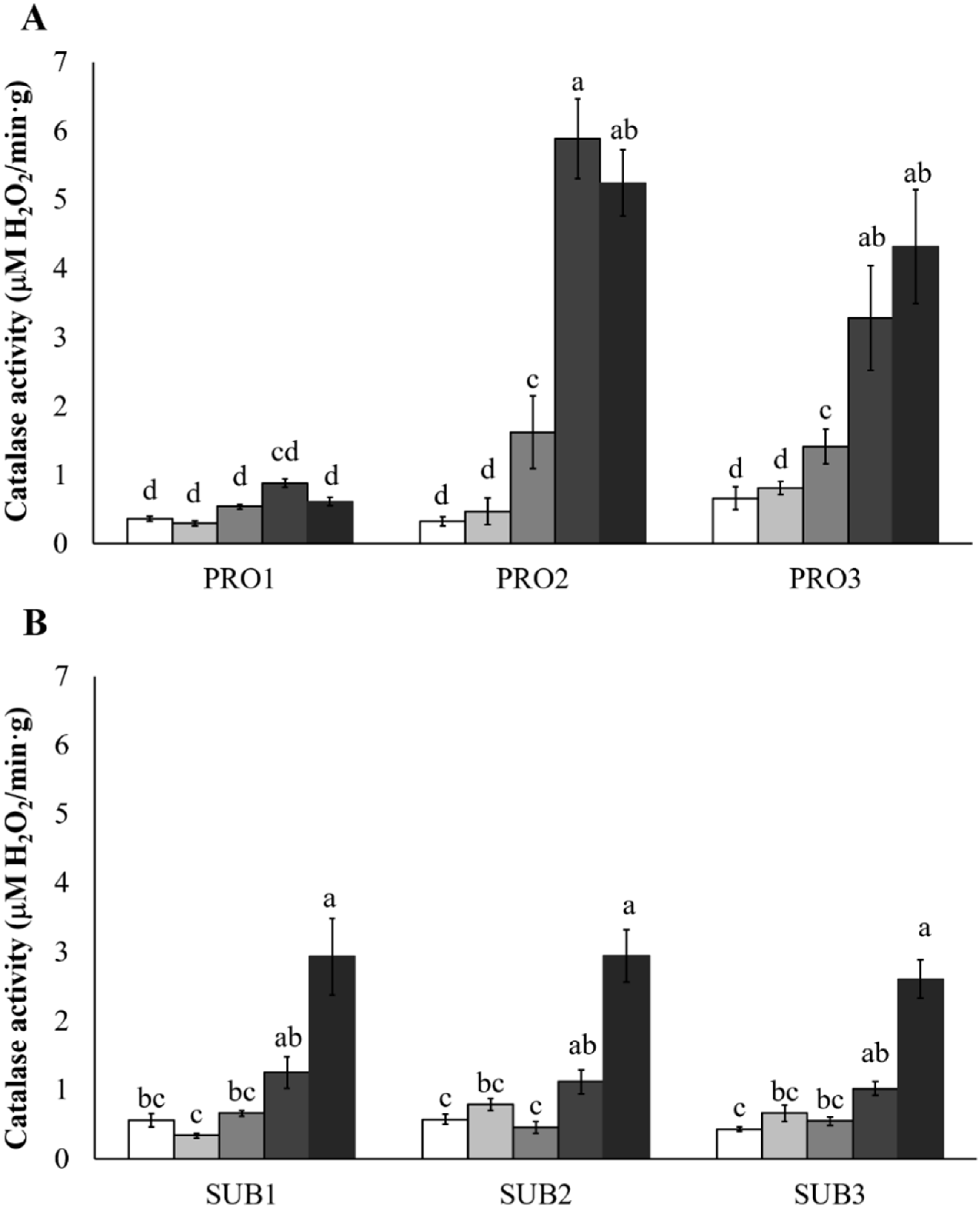
2.3. Effect of Hydrogen Peroxide Treatments on Catalase Activity
2.4. Effect of 1 mM Hydrogen Peroxide on MON and FBs in a Certified Maize Reference Material and in a Multi-Mycotoxin Analytical Standard Solution
3. Discussion and Conclusions
4. Materials and Methods
4.1. Chemicals
4.2. Fusarium Isolates Identification and Inoculum Preparation
4.3. Effect of Temperature on Fusarium Growth
4.4. Effect of Hydrogen Peroxide Treatments on Fusarium Biomass and Mycotoxin Production
4.5. Effect of Hydrogen Peroxide Treatments on Fusarium Catalase Activity
4.6. Effect of Hydrogen Peroxide on MON and FBs in a Certified Maize Reference Material and in a Multi-Mycotoxin Analytical Standard Solution
- -
- in a certified reference material of maize containing certified concentrations of FBs (FBs = FB1 + FB2 = 2600 ± 278 µg/kg) and measured concentration of MON (Trilogy® Analytical Laboratory, Washington, MO, USA)
- -
- in a multi-mycotoxin analytical standard solution containing a comparable concentration of FBs and MON to that of the certified reference material of maize.
- -
- Five grams of the certified reference material of maize were treated with 5 mL of H2O2 in a conical flask and incubated for 72h at 30 °C. After this incubation period the weight was checked and the sample was extracted with 15 mL of extracting solution CH3CN/H2O/CH3COOH (79/20/1, v/v/v) and subsequently underwent to the other extraction and dilution steps.
- -
- the multi-mycotoxin analytical standard solution with a comparable concentration of FBs and MON to that of the certified reference material of maize was prepared in CH3CN/H2O, 50/50 (v/v) containing 1 mM H2O2, not acidified with CH3COOH, in order to preserve the oxidizing capacity of H2O2, which is essential to assess the effect of H2O2 as a presumable degrading agent on MON and FBs.
4.7. LC-MS/MS Multi-Mycotoxin Analysis for the Determination of MON and FB Content
4.8. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Frisvad, J. A critical review of producers of small lactone mycotoxins: Patulin, penicillic acid and moniliformin. World Mycotoxin J. 2018, 11, 73–100. [Google Scholar] [CrossRef]
- Marasas, W.F.O.; Thiel, P.G.; Rabie, C.J.; Nelson, P.E.; Toussoun, T.A. Moniliformin Production in Fusarium Section Liseola. Mycologia 1986, 78, 242. [Google Scholar] [CrossRef]
- Scarpino, V.; Reyneri, A.; Vanara, F.; Scopel, C.; Causin, R.; Blandino, M. Relationship between European Corn Borer injury, Fusarium proliferatum and F. subglutinans infection and moniliformin contamination in maize. Field Crops Res. 2015, 183, 69–78. [Google Scholar] [CrossRef]
- Gutema, T.; Munimbazi, C.; Bullerman, L.B. Occurrence of Fumonisins and Moniliformin in Corn and Corn-Based Food Products of U.S. Origin†. J. Food Prot. 2000, 63, 1732–1737. [Google Scholar] [CrossRef]
- Logrieco, A.F.; Mule, G.; Moretti, A.; Bottalico, A. Toxigenic Fusarium Species and Mycotoxins Associated with Maize Ear Rot in Europe. Eur. J. Plant Pathol. 2002, 108, 597–609. [Google Scholar] [CrossRef]
- Fremy, J.-M.; Alassane-Kpembi, I.; Oswald, I.; Cottrill, B.; Van Egmond, H. A review on combined effects of moniliformin and co-occurring Fusarium toxins in farm animals. World Mycotoxin J. 2019, 12, 281–291. [Google Scholar] [CrossRef]
- Jestoi, M.N.; Paavanen-Huhtala, S.; Parikka, P.; Yli-Mattila, T. In vitroandin vivomycotoxin production ofFusariumspecies isolated from Finnish grains. Arch. Phytopathol. Plant Prot. 2008, 41, 545–558. [Google Scholar] [CrossRef]
- Jonsson, M.; Atosuo, J.; Jestoi, M.; Nathanail, A.V.; Kokkonen, U.-M.; Anttila, M.; Koivisto, P.; Lilius, E.-M.; Peltonen, K. Repeated dose 28-day oral toxicity study of moniliformin in rats. Toxicol. Lett. 2015, 233, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Akbar, A.; Baazeem, A.; Rodriguez, A.; Magan, N. Climate change, food security and mycotoxins: Do we know enough? Fungal Biol. Rev. 2017, 31, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Moretti, A.; Pascale, M.; Logrieco, A.F. Mycotoxin risks under a climate change scenario in Europe. Trends Food Sci. Technol. 2019, 84, 38–40. [Google Scholar] [CrossRef]
- Cendoya, E.; Monge, M.D.P.; Chiacchiera, S.M.; Farnochi, M.C.; Ramirez, M.L. Influence of water activity and temperature on growth and fumonisin production by Fusarium proliferatum strains on irradiated wheat grains. Int. J. Food Microbiol. 2018, 266, 158–166. [Google Scholar] [CrossRef]
- Doohan, F.; Brennan, J.; Cooke, B. Influence of Climatic Factors on Fusarium Species Pathogenic to Cereals. Eur. J. Plant Pathol. 2003, 109, 755–768. [Google Scholar] [CrossRef]
- Magan, N.; Medina, A. Integrating Gene Expression, Ecology and Mycotoxin Production by Fusarium and Aspergillus Species in Relation to Interacting Environmental Factors. World Mycotoxin J. 2016, 9, 673–684. [Google Scholar] [CrossRef] [Green Version]
- Montibus, M.; Pinson-Gadais, L.; Richard-Forget, F.; Barreau, C.; Ponts, N. Coupling of transcriptional response to oxidative stress and secondary metabolism regulation in filamentous fungi. Crit. Rev. Microbiol. 2013, 41, 295–308. [Google Scholar] [CrossRef]
- García-Cela, E.; Verheecke, C.; Magan, N.; Medina, A. The ‘‘-omics’’ contributions to the understanding of mycotoxin production under diverse environmental conditions. Curr. Opin. Food Sci. 2018, 23, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Medina, A.; Schmidt-Heydt, M.; Cárdenas-Chávez, D.L.; Parra, R.; Geisen, R.; Magan, N. Integrating toxin gene expression, growth and fumonisin B 1 and B 2 production by a strain of Fusarium verticillioides under different environmental factors. J. R. Soc. Interface 2013, 10, 20130320. [Google Scholar] [CrossRef] [Green Version]
- Picot, A.; Barreau, C.; Pinson-Gadais, L.; Caron, D.; Lannou, C.; Richard-Forget, F. Factors of theFusarium verticillioides-maize environment modulating fumonisin production. Crit. Rev. Microbiol. 2010, 36, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Kebede, H.; Abbas, H.K.; Fisher, D.K.; Bellaloui, N. Relationship between Aflatoxin Contamination and Physiological Responses of Corn Plants under Drought and Heat Stress. Toxins 2012, 4, 1385–1403. [Google Scholar] [CrossRef] [PubMed]
- Parsons, M.; Munkvold, G. Associations of planting date, drought stress, and insects withFusariumear rot and fumonisin B1contamination in California maize. Food Addit. Contam. Part A 2010, 27, 591–607. [Google Scholar] [CrossRef] [PubMed]
- Esaxena, I.; Srikanth, S.; Echen, Z. Cross Talk between H2O2 and Interacting Signal Molecules under Plant Stress Response. Front. Plant Sci. 2016, 7, 570. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Petrov, V.; Van Breusegem, F. Hydrogen peroxide—A central hub for information flow in plant cells. AoB Plants 2012, 2012, pls014. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.-Y.; Roze, L.V.; Linz, J.E. Oxidative Stress-Related Transcription Factors in the Regulation of Secondary Metabolism. Toxins 2013, 5, 683–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audenaert, K.; Callewaert, E.; Höfte, M.; De Saeger, S.; Haesaert, G. Hydrogen peroxide induced by the fungicide prothioconazole triggers deoxynivalenol (DON) production by Fusarium graminearum. BMC Microbiol. 2010, 10, 112. [Google Scholar] [CrossRef] [Green Version]
- Heller, J.; Tudzynski, P. Reactive Oxygen Species in Phytopathogenic Fungi: Signaling, Development, and Disease. Annu. Rev. Phytopathol. 2011, 49, 369–390. [Google Scholar] [CrossRef]
- Kulik, T.; Łojko, M.; Jestoi, M.; Perkowski, J. Sublethal concentrations of azoles induce tri transcript levels and trichothecene production in Fusarium graminearum. FEMS Microbiol. Lett. 2012, 335, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Schmidt-Heydt, M.; Magan, N.; Geisen, R. Stress induction of mycotoxin biosynthesis genes by abiotic factors. FEMS Microbiol. Lett. 2008, 284, 142–149. [Google Scholar] [CrossRef] [Green Version]
- Fountain, J.C.; Bajaj, P.; Pandey, M.; Nayak, S.N.; Yang, L.; Kumar, V.; Jayale, A.S.; Chitikineni, A.; Zhuang, W.; Scully, B.T.; et al. Oxidative stress and carbon metabolism influence Aspergillus flavus transcriptome composition and secondary metabolite production. Sci. Rep. 2016, 6, 38747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponts, N.; Couedelo, L.; Pinson-Gadais, L.; Verdal-Bonnin, M.-N.; Barreau, C.; Richard-Forget, F.; Verdal-Bonnin, M.-N. Fusariumresponse to oxidative stress by H2O2 is trichothecene chemotype-dependent. FEMS Microbiol. Lett. 2009, 293, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Ferrigo, D.; Raiola, A.; Bogialli, S.; Bortolini, C.; Tapparo, A.; Causin, R. In Vitro Production of Fumonisins by Fusarium verticillioides under Oxidative Stress Induced by H2O2. J. Agric. Food Chem. 2015, 63, 4879–4885. [Google Scholar] [CrossRef] [PubMed]
- Grintzalis, K.; Vernardis, S.; Klapa, M.I.; Georgiou, C.D. Role of Oxidative Stress in Sclerotial Differentiation and Aflatoxin B1 Biosynthesis in Aspergillus flavus. Appl. Environ. Microbiol. 2014, 80, 5561–5571. [Google Scholar] [CrossRef] [Green Version]
- Angelova, M.B.; Pashova, S.B.; Spasova, B.K.; Vassilev, S.V.; Slokoska, L.S. Oxidative stress response of filamentous fungi induced by hydrogen peroxide and paraquat. Mycol. Res. 2005, 109, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, D.; Sakai, K.; Suzuki, S.; Umemura, M.; Nogawa, T.; Kato, N.; Osada, H.; Watanabe, A.; Kawamoto, S.; Gonoi, T.; et al. Temperature during conidiation affects stress tolerance, pigmentation, and trypacidin accumulation in the conidia of the airborne pathogen Aspergillus fumigatus. PLoS ONE 2017, 12, e0177050. [Google Scholar] [CrossRef]
- Kreiner, M.; Harvey, L.M.; McNeil, B. Oxidative stress response of a recombinant Aspergillus niger to exogenous menadione and H2O2 addition. Enzym. Microb. Technol. 2002, 30, 346–353. [Google Scholar] [CrossRef]
- Nagygyörgy, E.; Kovács, B.; Leiter, E.; Miskei, M.; Pócsi, I.; Hornok, L.; Ádám, A. Toxicity of abiotic stressors to Fusarium species: Differences in hydrogen peroxide and fungicide tolerance. Acta Microbiol. Immunol. Hung. 2014, 61, 189–208. [Google Scholar] [CrossRef]
- Karlovsky, P.; Suman, M.; Berthiller, F.; De Meester, J.; Eisenbrand, G.; Perrin, I.; Oswald, I.; Speijers, G.; Chiodini, A.; Recker, T.; et al. Impact of food processing and detoxification treatments on mycotoxin contamination. Mycotoxin Res. 2016, 32, 179–205. [Google Scholar] [CrossRef]
- Prudente, A.D., Jr.; King, J.M. Efficacy and Safety Evaluation of Ozonation to Degrade Aflatoxin in Corn. J. Food Sci. 2002, 67, 2866–2872. [Google Scholar] [CrossRef]
- Aiko, V.; Mehta, A. Occurrence, detection and detoxification of mycotoxins. J. Biosci. 2015, 40, 943–954. [Google Scholar] [CrossRef] [PubMed]
- Freitas-Silva, O.; Venâncio, A. Ozone applications to prevent and degrade mycotoxins: A review. Drug Metab. Rev. 2010, 42, 612–620. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, J. Detoxifying Moniliformin in Grains and Water. J. Environ. Sci. 1997, 9, 215–220. [Google Scholar]
- Giles, S.S.; Batinić-Haberle, I.; Perfect, J.R.; Cox, G.M. Cryptococcus neoformans Mitochondrial Superoxide Dismutase: An Essential Link between Antioxidant Function and High-Temperature Growth. Eukaryot. Cell 2005, 4, 46–54. [Google Scholar] [CrossRef] [Green Version]
- Papapostolou, I.; Sideri, M.; Georgiou, C.D. Cell proliferating and differentiating role of H2O2 in Sclerotium rolfsii and Sclerotinia sclerotiorum. Microbiol. Res. 2014, 169, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Hydrogen peroxide as a central redox signaling molecule in physiological oxidative stress: Oxidative eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef]
- Cheeseman, J.M. Hydrogen Peroxide and Plant Stress: A Challenging Relationship. Plant Stress 2007, 1, 4–15. [Google Scholar]
- De Pinto, M.C.; Paradiso, A.; Leonetti, P.; De Gara, L. Hydrogen peroxide, nitric oxide and cytosolic ascorbate peroxidase at the crossroad between defence and cell death. Plant J. 2006, 48, 784–795. [Google Scholar] [CrossRef]
- Shalaby, S.; Horwitz, B.A. Plant phenolic compounds and oxidative stress: Integrated signals in fungal–plant interactions. Curr. Genet. 2014, 61, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Jayashree, T.; Subramanyam, C. Oxidative stress as a prerequisite for aflatoxin production by Aspergillus parasiticus. Free. Radic. Biol. Med. 2000, 29, 981–985. [Google Scholar] [CrossRef]
- Eponts, N. Mycotoxins are a component of Fusarium graminearum stress-response system. Front. Microbiol. 2015, 6, 1234. [Google Scholar] [CrossRef] [Green Version]
- Castellá, G.; Munkvold, G.P.; Imerman, P.; Hyde, W.G. Effects of temperature, incubation period and substrate on production of fusaproliferin by Fusarium subglutinans ITEM 2404. Nat. Toxins 1999, 7, 129–132. [Google Scholar] [CrossRef]
- Marin, S.; Sanchis, V.; Vinas, I.; Canela, R.; Magan, N. Effect of water activity and temperature on growth and fumonisin B1 and B2 production by Fusarium proliferatum and F. moniliforme on maize grain. Lett. Appl. Microbiol. 1995, 21, 298–301. [Google Scholar] [CrossRef]
- Covarelli, L.; Beccari, G.; Salvi, S. Infection by mycotoxigenic fungal species and mycotoxin contamination of maize grain in Umbria, central Italy. Food Chem. Toxicol. 2011, 49, 2365–2369. [Google Scholar] [CrossRef]
- Abrashev, R.; Stoitsova, S.; Krumova, E.; Pashova, S.; Paunova-Krasteva, T.; Vassilev, S.; Dolashka-Angelova, P.; Angelova, M. Temperature-stress tolerance of the fungal strain Aspergillus niger 26: Physiological and ultrastructural changes. World J. Microbiol. Biotechnol. 2013, 30, 1661–1668. [Google Scholar] [CrossRef]
- Wang, Z.-L.; Zhang, L.-B.; Ying, S.-H.; Feng, M.-G. Catalases play differentiated roles in the adaptation of a fungal entomopathogen to environmental stresses. Environ. Microbiol. 2012, 15, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Stępień, L.; Koczyk, G.; Waśkiewicz, A. Genetic and phenotypic variation of Fusarium proliferatum isolates from different host species. J. Appl. Genet. 2011, 52, 487–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostecki, M.; Wiśniewska, H.; Perrone, G.; Ritieni, A.; Jerzy, P.G.; Chelkowski, J.; Logrieco, A.F. The effects of cereal substrate and temperature on production of beauvericin, moniliformin and fusaproliferin by Fusarium subglutinans ITEM-1434. Food Addit. Contam. 1999, 16, 361–365. [Google Scholar] [CrossRef]
- Zhong, Z.; A McDonald, B.; Palma-Guerrero, J. Tolerance to oxidative stress is associated with both oxidative stress response and inherent growth in a fungal wheat pathogen. Genetics 2020, 217, iyaa022. [Google Scholar] [CrossRef] [PubMed]
- Croll, D.; McDonald, B.A. The genetic basis of local adaptation for pathogenic fungi in agricultural ecosystems. Mol. Ecol. 2016, 26, 2027–2040. [Google Scholar] [CrossRef] [PubMed]
- Stefansson, T.S.; McDonald, B.; Willi, Y. Local adaptation and evolutionary potential along a temperature gradient in the fungal pathogenRhynchosporium commune. Evol. Appl. 2013, 6, 524–534. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, R.; Li, Y.; Li, X.; You, L.; Kulikouskaya, V.; Hileuskaya, K. Degradation of polysaccharides from Sargassum fusiforme using UV/H2O2 and its effects on structural characteristics. Carbohydr. Polym. 2020, 230, 115647. [Google Scholar] [CrossRef]
- Tian, F.; Liu, Y.; Hu, K.; Zhao, B. The depolymerization mechanism of chitosan by hydrogen peroxide. J. Mater. Sci. 2003, 38, 4709–4712. [Google Scholar] [CrossRef]
- Chung, K.-R. Stress Response and Pathogenicity of the Necrotrophic Fungal PathogenAlternaria alternata. Scientifica 2012, 2012, 635431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.-Q.; Jiang, H.-F.; Wang, S.-Y.; Lei, Y.; Liao, B.-S. The Metabolic Responses of Aspergillus flavus to N-Acetylcysteine, Ascorbate, and H2O2. Agric. Sci. China 2008, 7, 74–81. [Google Scholar] [CrossRef]
- Aguirre, J.; Rios-Momberg, M.; Hewitt, D.; Hansberg, W. Reactive oxygen species and development in microbial eukaryotes. Trends Microbiol. 2005, 13, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Medentsev, A.G.; Arinbasarova, A.Y.; Akimenko, V.K. Adaptation of the Phytopathogenic Fungus Fusarium decemcellulare to Oxidative Stress. Microbiology 2001, 70, 26–30. [Google Scholar] [CrossRef]
- Aguirre, J.; Hansberg, W.; Navarro, R. Fungal responses to reactive oxygen species. Med Mycol. 2006, 44, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Segal, L.; Wilson, R.A. Reactive oxygen species metabolism and plant-fungal interactions. Fungal Genet. Biol. 2018, 110, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt-Heydt, M.; Stoll, D.; Schütz, P.; Geisen, R. Oxidative stress induces the biosynthesis of citrinin by Penicillium verrucosum at the expense of ochratoxin. Int. J. Food Microbiol. 2015, 192, 1–6. [Google Scholar] [CrossRef]
- Yun, Y.; Lu, Z.; Yang, J.; Liang, T.; Xiao, G.; Qiao, Y.; Liu, Y. Electrochemical analysis of specific catalase activity during development of Aspergillus flavus and its correlation with aflatoxin B1 production. Food Chem. 2020, 337, 127978. [Google Scholar] [CrossRef]
- McKenzie, K.; Sarr, A.; Mayura, K.; Bailey, R.; Miller, D.; Rogers, T.; Norred, W.; Voss, K.; Plattner, R.; Kubena, L.; et al. Oxidative degradation and detoxification of mycotoxins using a novel source of ozone. Food Chem. Toxicol. 1997, 35, 807–820. [Google Scholar] [CrossRef]
- Crespo-Sempere, A.; Selma-Lázaro, C.; Palumbo, J.D.; González-Candelas, L.; Martínez-Culebras, P.V. Effect of oxidant stressors and phenolic antioxidants on the ochratoxigenic fungusAspergillus carbonarius. J. Sci. Food Agric. 2015, 96, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Ferrigo, D.; Mondin, M.; Scopel, C.; Maso, E.D.; Stefenatti, M.; Raiola, A.; Causin, R. Effects of a prothioconazole- and tebuconazole-based fungicide on Aspergillus flavus development under laboratory and field conditions. Eur. J. Plant Pathol. 2019, 155, 151–161. [Google Scholar] [CrossRef]
- Furukawa, T.; Sakuda, S. Inhibition of Aflatoxin Production by Paraquat and External Superoxide Dismutase in Aspergillus flavus. Toxins 2019, 11, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateo, E.M.; Gómez, J.V.; Gimeno-Adelantado, J.V.; Romera, D.; Mateo-Castro, R.; Jiménez, M. Assessment of azole fungicides as a tool to control growth of Aspergillus flavus and aflatoxin B1 and B2 production in maize. Food Addit. Contam. Part A 2017, 34, 1039–1051. [Google Scholar] [CrossRef]
- Ponts, N.; Pinson-Gadais, L.; Verdal-Bonnin, M.-N.; Barreau, C.; Richard-Forget, F. Accumulation of deoxynivalenol and its 15-acetylated form is significantly modulated by oxidative stress in liquid cultures of Fusarium graminearum. FEMS Microbiol. Lett. 2006, 258, 102–107. [Google Scholar] [CrossRef] [Green Version]
- Mylona, K.; Kogkaki, E.; Sulyok, M.; Magan, N. Efficacy of gaseous ozone treatment on spore germination, growth and fumonisin production by Fusarium verticillioides in vitro and in situ in maize. J. Stored Prod. Res. 2014, 59, 178–184. [Google Scholar] [CrossRef]
- Shen, M.-H.; Singh, R.K. Detoxification of aflatoxins in foods by ultraviolet irradiation, hydrogen peroxide, and their combination—A review. LWT 2021, 142, 110986. [Google Scholar] [CrossRef]
- Fouler, S.G.; Trivedi, A.B.; Kitabatake, N. Detoxification of Citrinin and Ochratoxin A by Hydrogen Peroxide. J. AOAC Int. 1994, 77, 631–637. [Google Scholar] [CrossRef]
- Alla, E.S.A. Zearalenone: Incidence, toxigenic fungi and chemical decontamination in Egyptian cereals. Food/Nahrung 1997, 41, 362–365. [Google Scholar] [CrossRef]
- Afsah-Hejri, L.; Hajeb, P.; Ehsani, R.J. Application of ozone for degradation of mycotoxins in food: A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1777–1808. [Google Scholar] [CrossRef] [PubMed]
- Scarpino, V.; Reyneri, A.; Blandino, M. Development and Comparison of Two Multiresidue Methods for the Determination of 17 Aspergillus and Fusarium Mycotoxins in Cereals Using HPLC-ESI-TQ-MS/MS. Front. Microbiol. 2019, 10, 361. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; John Wiley & Sons: Hoboken, NJ, USA, 2008; ISBN 0470276460. [Google Scholar]
- Tuite, J. Plant Pathological Methods. Fungi and Bacteria; CABI: Wallingford, UK, 1969; p. 239. [Google Scholar]
- Mulè, G.; Susca, A.; Stea, G.; Moretti, A. A Species-Specific PCR Assay Based on the Calmodulin Partial Gene for Identification of Fusarium Verticillioides, F. Proliferatum and F. Subglutinans. Eur. J. Plant Pathol. 2004, 110, 495–502. [Google Scholar] [CrossRef]
- Van Der Fels-Klerx, H.; Liu, C.; Battilani, P. Modelling climate change impacts on mycotoxin contamination. World Mycotoxin J. 2016, 9, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Nakamoto, S. Germ Tube Formation of Candida albicans in Corn Meal Broth Using the Non-Slip Slide Glass Incubation Method. Yonago Acta Med. 1998, 41, 65–72. [Google Scholar]
- Klimek-Ochab, M.; Brzezińska-Rodak, M.; Żymańczyk-Duda, E.; Lejczak, B.; Kafarski, P. Comparative study of fungal cell disruption—Scope and limitations of the methods. Folia Microbiol. 2011, 56, 469–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadwan, M.H.; Abed, H.N. Data supporting the spectrophotometric method for the estimation of catalase activity. Data Brief 2015, 6, 194–199. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| PRO1 | PRO2 | PRO3 | ||||
|---|---|---|---|---|---|---|
| H2O2 Concentration | Biomass (mg ± SE) | Relative Yield% | Biomass (mg ± SE) | Relative Yield% | Biomass (mg ± SE) | Relative Yield% |
| Control | 1298 ± 153 a | 100 | 860 ± 80 a | 100 | 1701 ± 88 bc | 100 |
| 1 mM | 1275 ± 74 a | 98 | 809 ± 47 a | 94 | 1616 ± 55 c | 95 |
| 2 mM | 1362 ± 27 a | 105 | 832 ± 63 a | 97 | 2265 ± 107 a | 133 |
| 5 mM | 1246 ± 52 a | 96 | 920 ± 79 a | 107 | 2460 ± 312 a | 145 |
| 10 mM | 1220 ± 132 a | 94 | 958 ± 64 a | 111 | 1841 ± 181 ab | 108 |
| SUB1 | SUB2 | SUB3 | ||||
| H2O2 Concentration | Biomass (mg ± SE) | Relative Yield % | Biomass (mg ± SE) | Relative Yield% | Biomass (mg ± SE) | Relative Yield% |
| Control | 528 ± 27 a | 100 | 1047 ± 16 a | 100 | 1008 ± 71 ab | 100 |
| 1 mM | 385 ± 53 b | 73 | 635 ± 73 ab | 60 | 970 ± 73 ab | 96 |
| 2 mM | 417 ± 89 ab | 79 | 693 ± 22 ab | 66 | 1210 ± 112 a | 120 |
| 5 mM | 370 ± 52 bc | 70 | 534 ± 43 b | 51 | 1330 ± 96 a | 132 |
| 10 mM | 280 ± 33 c | 53 | 440 ± 81 b | 42 | 443 ± 39 c | 44 |
| Isolate | MON a | FBs a | ||||||
|---|---|---|---|---|---|---|---|---|
| ng/mg ± SE | p-Value b | Percent Variance (%) c | ng/mg ± SE | p-Value b | Percent Variance (%) c | |||
| Control | H2O2 | Control | H2O2 | |||||
| PRO1 | 7.843 ± 0.971 | 0.010 ± 0.002 | ** | −99.9 | 937.584 ± 34.583 | 337.061 ± 30.139 | ** | −64.1 |
| PRO2 | 0.018 ± 0.002 | 0.019 ± 0.001 | ns | 5.1 | 288.672 ± 118.101 | 112.375 ± 36.661 | ns | −61.1 |
| PRO3 | 0.299 ± 0.290 | 0,009 ± 0.001 | ns | −96.9 | 448.426 ± 45.825 | 517.013 ± 24.130 | ns | 15.3 |
| SUB1 | 0.029 ± 0.002 | 0.040 ± 0.005 | ns | 41.2 | 7.120 ± 6.195 | 0.282 ± 0.034 | ns | −96.0 |
| SUB2 | 49.654 ± 9.950 | 0.097 ± 0.073 | ** | −99.8 | 2.625 ± 2.525 | 0.170 ± 0.020 | ns | −93.5 |
| SUB3 | 144.156 ± 41.304 | 3.681 ± 3.665 | * | −97.4 | 6.128 ± 3.021 | 0.108 ± 0.001 | ns | −98.2 |
| Sample | MON | FBs | ||||||
|---|---|---|---|---|---|---|---|---|
| ng/mg ± SE | p-Value a | Percent Variance (%) b | ng/mg ± SE | p-Value a | Percent Variance (%) b | |||
| Control | H2O2 | Control | H2O2 | |||||
| Multi-mycotoxin standard | 1.763 ± 0.069 | <LOD c | *** | −100.0 | 2.639 ± 0.054 | 1.164 ± 0.025 | *** | −55.9 |
| Certified maize Reference Material | 1.755 ± 0.021 | 0.505 ± 0.014 | *** | −71.2 | 2.720 ± 0.088 | 0.645 ± 0.051 | *** | −76.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrigo, D.; Scarpino, V.; Vanara, F.; Causin, R.; Raiola, A.; Blandino, M. Influence of H2O2-Induced Oxidative Stress on In Vitro Growth and Moniliformin and Fumonisins Accumulation by Fusarium proliferatum and Fusarium subglutinans. Toxins 2021, 13, 653. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090653
Ferrigo D, Scarpino V, Vanara F, Causin R, Raiola A, Blandino M. Influence of H2O2-Induced Oxidative Stress on In Vitro Growth and Moniliformin and Fumonisins Accumulation by Fusarium proliferatum and Fusarium subglutinans. Toxins. 2021; 13(9):653. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090653
Chicago/Turabian StyleFerrigo, Davide, Valentina Scarpino, Francesca Vanara, Roberto Causin, Alessandro Raiola, and Massimo Blandino. 2021. "Influence of H2O2-Induced Oxidative Stress on In Vitro Growth and Moniliformin and Fumonisins Accumulation by Fusarium proliferatum and Fusarium subglutinans" Toxins 13, no. 9: 653. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090653