Lipopolysaccharide from the Cyanobacterium Geitlerinema sp. Induces Neutrophil Infiltration and Lung Inflammation

 , , and
, , and 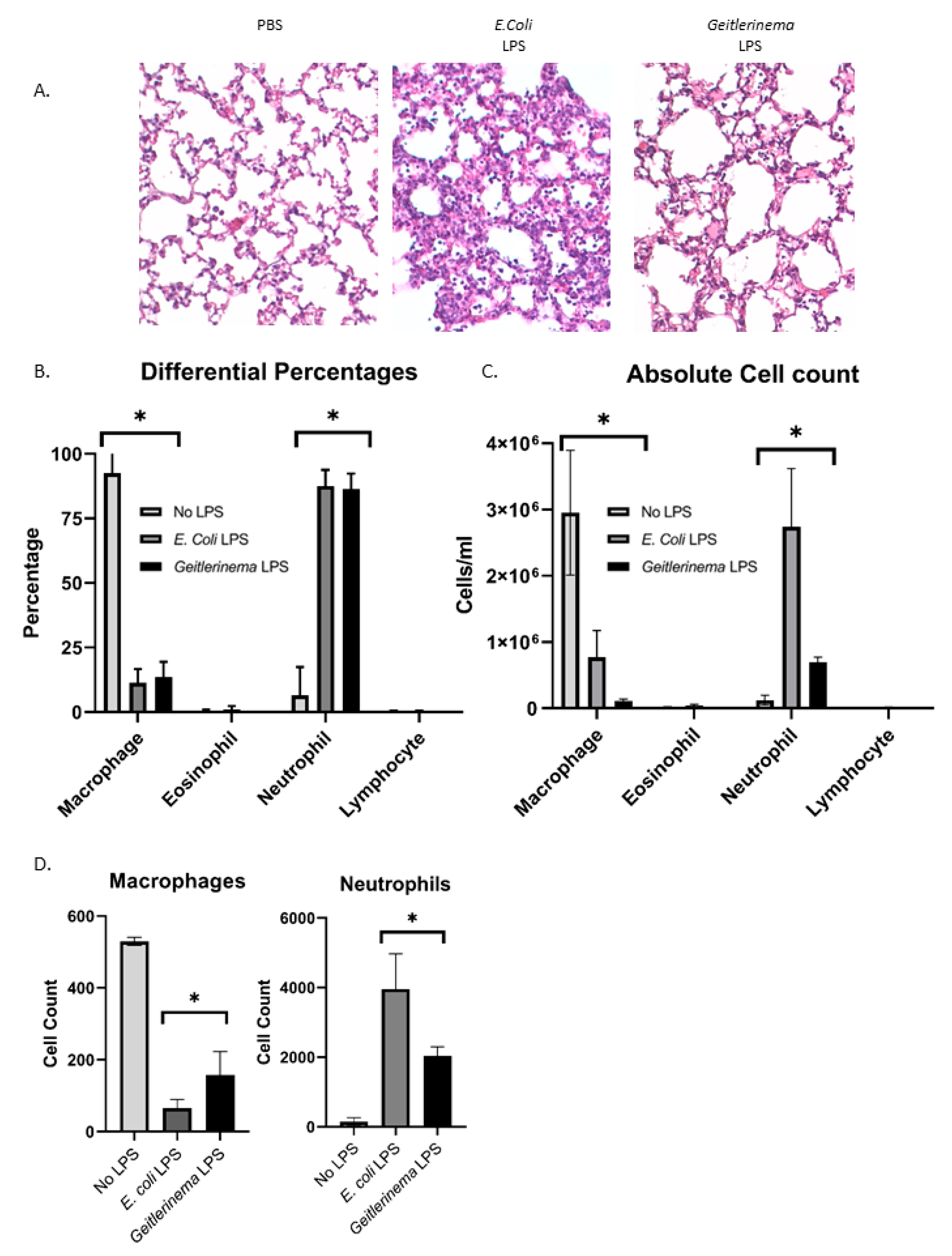{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Aerosolized Geitlerinema sp. LPS Induces Lung Inflammation In Vivo
2.2. Geitlerinema sp. LPS Induces Cytokine Production in the Lungs of LPS-Treated Mice In Vivo
2.3. Geitlerinema sp. LPS Induces ICAM-1 Levels in the Lungs of LPS-Treated Mice In Vivo and Human Airway Epithelial Cells In Vitro
2.4. Geitlerinema sp. LPS Induces IL-6 and IL-8 in an NF-kB-Dependent Manner In Vitro
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Lipopolysaccharide Isolation from HCC1097
4.3. In Vivo LPS Exposure
4.4. Flow Cytometry
4.5. MULTIPLEX Assay for Cytokines and Chemokines
4.6. Hematoxylin and Eosin Staining
4.7. Immunohistochemistry
4.8. Immunofluorescence Staining
4.9. ELISA
4.10. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marshall, C.L.; Hasani, K.; Mookherjee, N. Immunobiology of Steroid-Unresponsive Severe Asthma. Front. Allergy 2021, 2. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y. Heterogeneity of Glucocorticoid Resistance in Patients with Bronchial Asthma. Int. J. Biomed. Sci. IJBS 2010, 6, 158–166. [Google Scholar] [PubMed]
- Wadhwa, R.; Dua, K.; Adcock, I.M.; Horvat, J.C.; Kim, R.Y.; Hansbro, P.M. Cellular Mechanisms Underlying Steroid-Resistant Asthma. Eur. Respir. Rev. 2019, 28, 190096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, P.J.; Adcock, I.M. Glucocorticoid Resistance in Inflammatory Diseases. Lancet 2009, 373, 1905–1917. [Google Scholar] [CrossRef]
- Nunes, C.; Pereira, A.M.; Morais-Almeida, M. Asthma Costs and Social Impact. Asthma Res. Pract. 2017, 3, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; He, X.Y.; Baines, K.; Gunawardhana, L.P.; Simpson, J.L.; Li, F.; Gibson, P.G. Different Inflammatory Phenotypes in Adults and Children with Acute Asthma. Eur. Respir. J. 2011, 38, 567–574. [Google Scholar] [CrossRef] [Green Version]
- Nabe, T. Steroid-Resistant Asthma and Neutrophils. Biol. Pharm. Bull. 2020, 43, 31–35. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishnan, R.K.; Al Heialy, S.; Hamid, Q. Role of IL-17 in Asthma Pathogenesis and Its Implications for the Clinic. Expert Rev. Respir. Med. 2019, 13, 1057–1068. [Google Scholar] [CrossRef]
- Wiśniewska, K.; Lewandowska, A.; Śliwińska-Wilczewska, S. The Importance of Cyanobacteria and Microalgae Present in Aerosols to Human Health and the Environment—Review Study. Environ. Int. 2019, 131, 104964. [Google Scholar] [CrossRef]
- Yu, Q.; Chen, Z. Establishment of Different Experimental Asthma Models in Mice. Exp. Ther. Med. 2018, 15, 2492–2498. [Google Scholar] [CrossRef] [Green Version]
- Goleva, E.; Hauk, P.J.; Hall, C.F.; Liu, A.H.; Riches, D.W.; Martin, R.J.; Leung, D.Y. Corticosteroid-Resistant Asthma Is Associated with Classical Antimicrobial Activation of Airway Macrophages. J. Allergy Clin. Immunol. 2008, 122, 550–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matute-Bello, G.; Charles, W.; Frevert, C.W.; Martin, T.R. Animal Models of Acute Lung Injury. Am. J. Physiol. 2008, 295, L379–L399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moosová, Z.; Šindlerová, L.; Ambrůzová, B.; Ambrožová, G.; Vašíček, O.; Velki, M.; Babica, P.; Kubala, L. Lipopolysaccharides from Microcystis Cyanobacteria-Dominated Water Bloom and from Laboratory Cultures Trigger Human Immune Innate Response. Toxins 2019, 11, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, I.; Schluter, P.J.; Shaw, G.R. Cyanobacterial Lipopolysaccharides and Human Health—A Review. Environ. Health 2006, 5, 7. [Google Scholar] [CrossRef] [Green Version]
- Plaas, H.E.; Paerl, H.W. Toxic Cyanobacteria: A Growing Threat to Water and Air Quality. Environ. Sci. Technol. 2020, 55, 44–64. [Google Scholar] [CrossRef]
- Backer, L.C.; Manassaram-Baptiste, D.; LePrell, R.; Bolton, B. Cyanobacteria and Algae Blooms: Review of Health and Environmental Data from the Harmful Algal Bloom-Related Illness Surveillance System (HABISS) 2007–2011. Toxins 2015, 7, 1048–1064. [Google Scholar] [CrossRef] [Green Version]
- Genitsaris, S.; Kanstantinos, A.K.; Moutaka-Gouni, M. Airbone Algae and Cyanobacteria: Occurrence and Related Health Effects. Front. Biosci. 2011, 3, 772–787. [Google Scholar]
- Chu, W.-L.; Tneh, S.-Y.; Ambu, S. A Survey of Airborne Algae and Cyanobacteria within the Indoor Environment of an Office Building in Kuala Lumpur, Malaysia. Grana 2013, 52, 207–220. [Google Scholar] [CrossRef]
- Annadotter, H.; Cronberg, G.; Nystrand, R.; Rylander, R. Endotoxins from Cyanobacteria and Gram-negative Bacteria as the Cause of an Acute Influenza-like Reaction after Inhalation of Aerosols. EcoHealth 2005, 2, 209–221. [Google Scholar] [CrossRef]
- Facciponte, D.N.; Bough, M.W.; Seidler, D.; Carroll, J.L.; Ashare, A.; Andrew, A.S.; Tsongalis, G.J.; Vaickus, L.J.; Henegan, P.L.; Butt, T.H.; et al. Identifying Aerosolized Cyanobacteria in the Human Respiratory Tract: A Proposed Mechanism for Cyanotoxin-Associated Diseases. Sci. Total Environ. 2018, 645, 1003–1013. [Google Scholar] [CrossRef]
- Lang-Yona, N.; Lehahn, Y.; Herut, B.; Burshtein, N.; Rudich, Y. Marine Aerosol as a Possible Source for Endotoxins in Coastal Areas. Sci. Total Environ. 2014, 499, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Slatkin, D.N.; Stoner, R.D.; Adams, W.H.; Kycia, J.H.; Siegelman, H.W. Atypical Pulmonary Thrombosis Caused by a Toxic Cyanobacterial Peptide. Science 1983, 220, 1383–1385. [Google Scholar] [CrossRef] [PubMed]
- Picanço, M.; Soares, R.; Cagido, V.; Azevedo, S.; Rocco, P.; Zin, W. Toxicity of a Cyanobacterial Extract Containing Microcystins to Mouse Lungs. Braz. J. Med. Biol. Res. 2004, 37, 1225–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohkouchi, Y.; Tajima, S.; Nomura, M.; Itoh, S. Inflammatory Responses and Potencies of Various Lipopolysaccharides from Bacteria and Cyanobacteria in Aquatic Environments and Water Supply Systems. Toxicon 2015, 97, 23–31. [Google Scholar] [CrossRef]
- Dantas, E.W.; Moura, A.N.; Bittencourt-Oliveira, M.D.C. Cyanobacterial Blooms in Stratified and Destratified Eutrophic Reservoirs in Semi-arid Region of Brazil. Biol. Sci. 2011, 83, 1327–1338. [Google Scholar] [CrossRef] [Green Version]
- Hur, M.; Lee, I.; Tak, B.-M.; Lee, H.J.; Yu, J.J.; Cheon, S.U.; Kim, B.-S. Temporal Shifts in Cyanobacterial Communities at Different Sites on the Nakdong River in Korea. Water Res. 2013, 47, 6973–6982. [Google Scholar] [CrossRef]
- Willame, R.; Boutte, C.; Grubisic, S.; Wilmotte, A.; Komárek, J.; Hoffmann, L. Morphological and Molecular Characterization of Planktonic Cyanobacteria from Belgium and Luxembourg. J. Phycol. 2006, 42, 1312–1332. [Google Scholar] [CrossRef]
- Swanson-Mungerson, M.; Incrocci, R.; Subramaniam, V.; Williams, P.; Hall, M.L.; Mayer, A. Effects of Cyanobacteria Oscillatoria sp. Lipopolysaccharide on B Cell Activation and Toll-like Receptor 4 Signaling. Toxicol. Lett. 2017, 275, 101–107. [Google Scholar] [CrossRef]
- Swanson-Mungerson, M.; Williams, P.G.; Gurr, J.R.; Incrocci, R.; Subramaniam, V.; Radowska, R.; Hall, M.L.; Mayer, A.M.S. Biochemical and Functional Analysis of Cyanobacterium Geitlerinema sp. LPS on Human Monocytes. Toxicol. Sci. 2019, 171, 421–430. [Google Scholar] [CrossRef]
- Dhaliwal, K.; Scholefield, E.; Ferenbach, D.; Gibbons, M.; Duffin, R.; Dorward, D.A.; Morris, A.C.; Humphries, D.; MacKinnon, A.; Wilkinson, T.S.; et al. Monocytes Control Second-Phase Neutrophil Emigration in Established Lipopolysaccharide-induced Murine Lung Injury. Am. J. Respir. Crit. Care Med. 2012, 186, 514–524. [Google Scholar] [CrossRef] [Green Version]
- Swartzendruber, J.A.; Incrocci, R.W.; Wolf, S.A.; Jung, A.; Knight, K.L. Bacillus Subtilis Exopolysaccharide Prevents Allergic Eosinophilia. Allergy 2018, 74, 819–821. [Google Scholar] [CrossRef] [PubMed]
- Baudiss, K.; Vieira, R.d.P.; Cicko, S.; Ayata, K.; Hossfeld, M.; Ehrat, N.; Gómez-Muñoz, A.; Eltzschig, H.K.; Idzko, M. C1P Attenuates Lipopolysaccharide-Induced Acute Lung Injury by Preventing NF-kappaB Activation in Neutrophils. J. Immunol. 2016, 196, 2319–2326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, C.J.; Finkelstein, J.N.; Gelein, R.; Oberdörster, G. Pulmonary Cytokine and Chemokine mRNA Levels after Onhalation of Lipopolysaccharide in C57BL/6 Mice. Toxicol. Sci. 1998, 46, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Hol, J.; Wilhelmsen, L.; Haraldsen, G. The Murine IL-8 Homologues KC, MIP-2, and LIX are Found in Endothelial Cytoplasmic Granules but Not in Weibel-Palade Bodies. J. Leukoc. Biol. 2010, 87, 501–508. [Google Scholar] [CrossRef]
- Hoffmann, E.; Dittrich-Breiholz, O.; Holtmann, H.; Kracht, M. Multiple Control of Interleukin-8 Gene Expression. J. Leukoc. Biol. 2002, 72, 847–855. [Google Scholar]
- Libermann, T.A.; Baltimore, D. Activation of Interleukin-6 Gene Expression through the NF-kappa B Transcription Factor. Mol. Cell. Biol. 1990, 10, 2327–2334. [Google Scholar] [CrossRef]
- Stewart, I.; Webb, P.M.; Schluter, P.J.; Fleming, L.E.; Burns, J.W., Jr.; Gantar, M.; Backer, L.C.; Shaw, G.R. Epidemiology of Recreational Exposure to Freshwater Cyanobacteria—An International Prospective Cohort Study. BMC Public Health 2006, 6, 93. [Google Scholar] [CrossRef] [Green Version]
- Omidi, A.; Pflugmacher, S.; Kaplan, A.; Kim, Y.; Esterhuizen, M. Reviewing Interspecies Interactions as a Driving Force Affecting the Community Structure in Lakes via Cyanotoxins. Microorganisms 2021, 9, 1583. [Google Scholar] [CrossRef]
- Wang, Z.; Akbar, S.; Sun, Y.; Gu, L.; Zhang, L.; Lyu, K.; Huang, Y.; Yang, Z. Cyanobacterial Dominance and Succession: Factors, Mechanisms, Predictions, and Managements. J. Environ. Manag. 2021, 297, 113281. [Google Scholar] [CrossRef]
- Borges, H.L.F.; Branco, L.H.Z.; Martins, M.D.; Lima, C.; Barbosa, P.T.; Lira, G.A.S.T.; Bittencourt-Oliveira, M.C.; Molica, R.J.R. Cyanotoxin Production and Phylogeny of Benthic Cyanobacterial Strains Isolated from the Northeast of Brazil. Harmful Algae 2015, 43, 46–57. [Google Scholar] [CrossRef]
- Richardson, L.L.; Sekar, R.; Myers, J.L.; Gantar, M.; Voss, J.D.; Kaczmarsky, L.; Remily, E.R.; Boyer, G.L.; Zimba, P.V. The Presence of the Cyanobacterial Toxin Microcystin in Black Band Disease of Corals. FEMS Microbiol. Lett. 2007, 272, 182–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samylina, O.S.; Sapozhnikov, F.V.; Gainanova, O.Y.; Ryabova, A.V.; Nikitin, M.A.; Sorokin, D.Y. Algo-Bacterial Communities of the Kulunda Steppe (Altai Region, Russia) Soda Lakes. Microbiology 2014, 83, 849–860. [Google Scholar] [CrossRef]
- Andrianasolo, E.H.; Goeger, D.; Gerwick, W.H. Mitsoamide: A Cytotoxic Linear Lipopeptide from the Madagascar Marine Cyanobacterium Geitlerinema sp. Pure Appl. Chem. 2007, 79, 593–602. [Google Scholar] [CrossRef]
- Lankoff, A.; Carmichael, W.W.; Grasman, K.A.; Yuan, M. The Uptake Kinetics and Immunotoxic Effects of Microcystin-LR in Human and Chicken Peripheral Blood Lymphocytes in Vitro. Toxicology 2004, 204, 23–40. [Google Scholar] [CrossRef]
- Teneva, I.; Mladenov, R.; Popov, N.; Dzhambazov, B. Cytotoxicity and Apoptotic Effects of Microcystin-LR and Anatoxin-a in Mouse Lymphocytes. Folia Biol. 2005, 51, 62–77. [Google Scholar]
- Chen, T.; Zhao, X.; Liu, Y.; Shi, Q.; Hua, Z.; Shen, P. Analysis of Immunomodulating Nitric Oxide, iNOS and Cytokines mRNA in Mouse Macrophages Induced by Microcystin-LR. Toxicology 2004, 197, 67–77. [Google Scholar] [CrossRef]
- Diez-Quijada, L.; Benítez-González, M.D.M.; Puerto, M.; Jos, A.; Cameán, A.M. Immunotoxic Effects Induced by Microcystins and Cylindrospermopsin: A Review. Toxins 2021, 13, 711. [Google Scholar] [CrossRef]
- Rocha, M.F.G.; Sidrim, J.J.C.; Soares, A.M.; Jimenez, G.C.; Guerrant, R.L.; Ribeiro, R.A.; Lima, A.A.M. Supernatants from Macrophages Stimulated with Microcystin-LR Induce Electrogenic Intestinal Response in Rabbit Ileum. Pharmacol. Toxicol. 2000, 87, 46–51. [Google Scholar] [CrossRef]
- Pahan, K.; Sheikh, F.G.; Namboodiri, A.M.; Singh, I. Inhibitors of Protein Phosphatase 1 and 2A Differentially Regulate the Expression of Inducible Nitric-oxide Synthase in Rat Astrocytes and Macrophages. J. Biol. Chem. 1998, 273, 12219–12226. [Google Scholar] [CrossRef] [Green Version]
- Sharma, N.K.; Rai, A.K. Allergenicity of Airborne Cyanobacteria Phormidium Fragile and Nostoc Muscorum. Ecotoxicol. Environ. Saf. 2008, 69, 158–162. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Murphy, J.; MacAdam, D.; Osterbauer, C.; Baseer, I.; Hall, M.L.; Feher, D.; Williams, P. Classical and Alternative Activation of Cyanobacterium Oscillatoria sp. Lipopolysaccharide-Treated Rat Microglia in vitro. Toxicol. Sci. 2016, 149, 484–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Boer, J.D.; Roelofs, J.J.T.H.; De Vos, A.F.; De Beer, R.; Schouten, M.; Hommes, T.J.; Hoogendijk, A.J.; de Boer, O.; Stroo, I.; Van Der Zee, J.S.; et al. Lipopolysaccharide Inhibits Th2 Lung Inflammation Induced by House Dust Mite Allergens in Mice. Am. J. Respir. Cell Mol. Biol. 2013, 48, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Chorus, I.; Falconer, I.R.; Salas, H.J.; Bartram, J. Health Risks Caused by Freshwater Cyanobacteria in Recreational Waters. J. Toxicol. Environ. Health Part B 2000, 3, 323–347. [Google Scholar] [CrossRef]
- Codd, G.A.; Morrison, L.F.; Metcalf, J. Cyanobacterial Toxins: Risk Management for Health Protection. Toxicol. Appl. Pharmacol. 2005, 203, 264–272. [Google Scholar] [CrossRef]
- May, N.W.; Olson, N.E.; Panas, M.G.; Axson, J.L.; Tirella, P.S.; Kirpes, R.M.; Craig, R.L.; Gunsch, M.J.; China, S.; Laskin, A.; et al. Aerosol Emissions from Great Lakes Harmful Algal Blooms. Environ. Sci. Technol. 2018, 52, 397–405. [Google Scholar] [CrossRef]
- Wiśniewska, K.A.; Śliwińska-Wilczewska, S.; Lewandowska, A.U. The First Characterization of Airborne Cyanobacteria and Microalgae in the Adriatic Sea Region. PLoS ONE 2020, 15, e0238808. [Google Scholar] [CrossRef]
- Kline, J.N.; Cowden, J.D.; Hunninghake, G.W.; Schutte, B.C.; Watt, J.L.; Wohlford-Lenane, C.L.; Powers, L.S.; Jones, M.P.; Schwartz, D.A. Variable Airway Responsiveness to Inhaled Lipopolysaccharide. Am. J. Respir. Crit. Care Med. 1999, 160, 297–303. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Swartzendruber, J.A.; Del Toro, R.M.; Incrocci, R.; Seangmany, N.; Gurr, J.R.; Mayer, A.M.S.; Williams, P.G.; Swanson-Mungerson, M. Lipopolysaccharide from the Cyanobacterium Geitlerinema sp. Induces Neutrophil Infiltration and Lung Inflammation. Toxins 2022, 14, 267. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14040267
Swartzendruber JA, Del Toro RM, Incrocci R, Seangmany N, Gurr JR, Mayer AMS, Williams PG, Swanson-Mungerson M. Lipopolysaccharide from the Cyanobacterium Geitlerinema sp. Induces Neutrophil Infiltration and Lung Inflammation. Toxins. 2022; 14(4):267. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14040267
Chicago/Turabian StyleSwartzendruber, Julie A., Rosalinda Monroy Del Toro, Ryan Incrocci, Nessa Seangmany, Joshua R. Gurr, Alejandro M. S. Mayer, Philip G. Williams, and Michelle Swanson-Mungerson. 2022. "Lipopolysaccharide from the Cyanobacterium Geitlerinema sp. Induces Neutrophil Infiltration and Lung Inflammation" Toxins 14, no. 4: 267. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14040267