Multiple Year Influences of the Aflatoxin Biocontrol Product AF-X1 on the A. flavus Communities Associated with Maize Production in Italy

 ,
, 
 and
and
Abstract
:1. Introduction
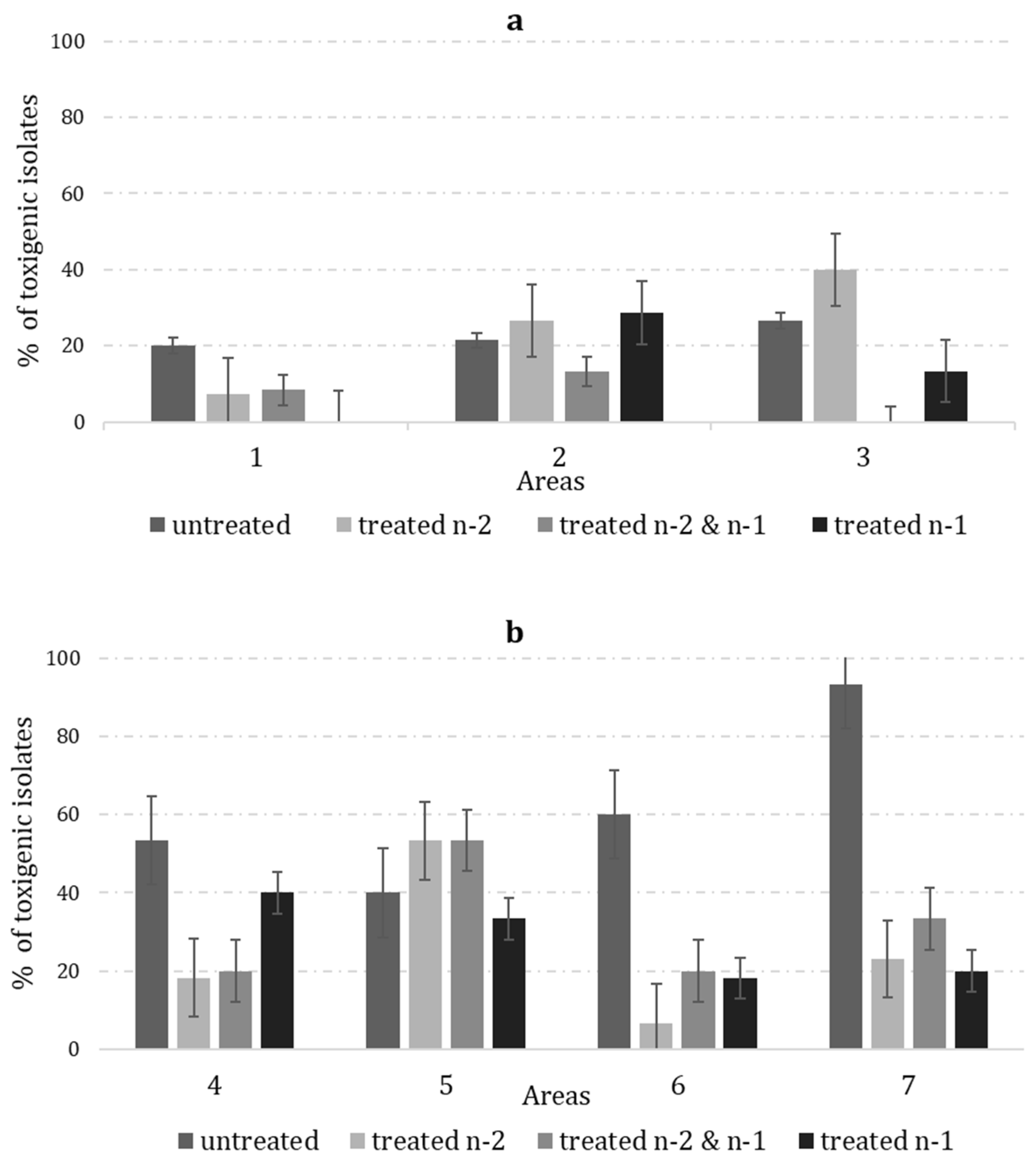
2. Results
2.1. Cropping Systems of the Surveyed Fields
2.2. Soil Fungal Populations
2.3. Frequency of Non-Alfatoxigenic and Toxigenic A. flavus
2.4. Frequency of Isolates Belonging to VCG IT006
2.5. Impact of Cropping System on the Soil Fungal Population
3. Discussion
4. Material and Methods
4.1. Soil Sampling and Filed Data Collection
4.2. Aspergillus flavus Isolation
4.3. DNA Extraction
4.4. qPCR Conditions
4.5. Vegetative Compatibility Analysis (VCA)
4.6. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cotty, P.J.; Bayman, D.S.; Egel, D.S.; Elias, K.S. Agriculture, Aflatoxins and Aspergillus. In The Genus Aspergillus; Powell, K., Ed.; Plenum Press: New York, NY, USA, 1994; pp. 1–2. [Google Scholar]
- Piva, G.; Battilani, P.; Pietri, A. Emerging Issues in Southern Europe: Aflatoxins in Italy. In Emerging issues in southern Europe: Aflatoxins in Italy; Barug, D., Bhatnagar, D., Van Egmond, H.P., Van der Kamp, J.W., Van Ossenbruggen, W.A., Visconti, A., Eds.; Wageningen Academic Publishers: Wgeningen, The Netherland, 2006; pp. 135–139. [Google Scholar]
- Perrone, G.; Gallo, A.; Logrieco, A.F. Biodiversity of Aspergillus section Flavi in Europe in relation to the management of aflatoxin risk. Front. Microbiol. 2014, 5, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bock, C.H.; Mackey, B.; Cotty, P.J. Population dynamics of Aspergillus flavus in the air of an intensively cultivated region of South-West Arizona. Plant Pathol. 2004, 5, 422–433. [Google Scholar] [CrossRef]
- Zuluaga-montero, A.; Rami, L.; Rauscher, J.; Bayman, P. Marine isolates of Aspergillus flavus: Denizens of the deep or lost at sea? Fungal Ecol. 2010, 3, 5–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization; International Agency for Research on Cancer. Aflatoxins. In IARC Monographs on the Evaluation of Carcinogenc Risks to Humans; IARC Press: Lyon, France, 2002; Volume 82, pp. 171–300. [Google Scholar]
- Marchese, S.; Polo, A.; Ariano, A.; Velotto, S.; Costantini, S.; Severino, L. Aflatoxin B1 and M1: Biological properties and their involvement in cancer development. Toxins 2018, 10, 214. [Google Scholar] [CrossRef] [Green Version]
- Ayalew, A.; Kimanya, M.; Matumba, L.; Bandyopadhayay, R.; Menkir, A.; Cotty, P. Controlling aflatoxins in maize in Africa: Strategies, challenges and opportunities for improvement. In Achieving Sustainable Cultivation of Maize Cultivation Techniques, Pest, and Disease Control. Volume 2; Watson, D., Ed.; Burleigh Dodds Science: Cambridge, UK, 2017; pp. 1–24. [Google Scholar]
- Singh, P.; Cotty, P.J. Aflatoxin contamination of dried red chilies: Contrasts between the United States and Nigeria, two markets differing in regulation enforcement. Food Control 2017, 80, 374–379. [Google Scholar] [CrossRef]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25%. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef]
- Hedayati, M.T.; Pasqualotto, A.C.; Warn, P.A.; Bowyer, P.; Denning, D.W. Aspergillus flavus: Human pathogen, allergen and mycotoxin producer. Microbiology 2007, 153, 1677–1692. [Google Scholar] [CrossRef] [Green Version]
- Sepahvand, A.; Shams-ghahfarokhi, M. A survey on distribution and toxigenicity of Aspergillus flavus from indoor and outdoor hospital environments. Folia Microbiol. 2011, 56, 527–534. [Google Scholar] [CrossRef]
- Horn, B.W. Ecology and population biology of aflatoxigenic fungi in soil. J. Toxicol.—Toxin Rev. 2003, 22, 351–379. [Google Scholar] [CrossRef]
- Ojiambo, P.S.; Battilani, P.; Cary, J.W.; Blum, B.H.; Carbone, I. Cultural and genetic approaches to manage aflatoxin contamination: Recent insights provide opportunities for improved control. Phytopathology 2018, 108, 1024–1037. [Google Scholar] [CrossRef] [Green Version]
- Mehl, H.L.; Jaime-Garcia, R.; Callicott, K.A.; Probst, C.; Garber, N.P.; Ortega-Beltran, A.; Grubisha, L.C.; Cotty, P.J. Aspergillus flavus diversity on crops and in the environment can be exploited to reduce aflatoxin exposure and improve health. Ann. N. Y. Acad. Sci. 2012, 1273, 7–17. [Google Scholar] [CrossRef]
- Cotty, P.J. Virulence and cultural characteristics of two Aspergillus flavus strains pathogenic on cotton. Phytopathology 1989, 79, 808–814. [Google Scholar] [CrossRef] [Green Version]
- Grubisha, L.C.; Cotty, P.J. Genetic analysis of the Aspergillus flavus vegetative compatibility group to which a biological control agent that limits aflatoxin contamination in U.S. crops belongs. Appl. Environ. Microbiol. 2015, 81, 5889–5899. [Google Scholar] [CrossRef] [Green Version]
- Leslie, J.F. Fungal Vegetative Compatibility. Annu. Rev. Phytopathol. 1993, 31, 127–150. [Google Scholar] [CrossRef]
- Adhikari, B.N.; Bandyopadhyay, R.; Cotty, P.J. Degeneration of aflatoxin gene clusters in Aspergillus flavus from Africa and North America. AMB Express 2016, 6, 771–789. [Google Scholar] [CrossRef] [Green Version]
- Cotty, P.J.; Bayman, P. Competitive exculusion of a toxigenic strain of Aspergillus flavus by and atoxigenic strain. Postharvest Pathol. Mycotoxins 1993, 83, 1283–1287. [Google Scholar]
- Cotty, P.J. Biocompetitive exclusion of toxigenic fungi. In The Mycotoxin Factbook: Food and Feed Topics; Barug, D., Bhatnagar, D., van Egmond, H.P., van der Kamp, J.W., van Osenbruggen, W.A., Visconti, A., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2006; pp. 179–197. [Google Scholar]
- Cotty, P.J.; Probst, C.; Jaime-Garcia, R. Etiology and Management of Aflatoxin Contamination. In Mycotoxins: Detection Methods, Management, Public Health and Agricultural Trade; Leslie, J.F., Bandyopadhyay, R., Visconti, A., Eds.; CABI Publisher: Wallingford, UK, 2008; pp. 287–299. [Google Scholar]
- Weaver, M.A.; Abbas, H.K. Field displacement of aflatoxigenic Aspergillus flavus strains through repeated biological control applications. Front. Microbiol. 2019, 10, 1788. [Google Scholar] [CrossRef] [Green Version]
- Atehnkeng, J.; Ojiambo, P.; Ortega-Beltran, A.; Augusto, J.; Cotty, P.J.; Bandyopadhyay, R. Impact of frequency of application on the long-term efficacy of the biocontrol product aflasafe in reducing aflatoxin contamination in maize. Front. Microbiol. 2022, 13, 1049013. [Google Scholar] [CrossRef]
- Khan, R.; Ghazali, F.M.; Mahyudin, N.A.; Samsudin, N.I.P. Aflatoxin biosynthesis, genetic regulation, toxicity, and control strategies: A Review. J. Fungi 2021, 7, 606. [Google Scholar] [CrossRef]
- Moral, J.; Garcia-Lopez, M.T.; Camiletti, B.X.; Jaime-Garcia, R.; Michailides, T.J.; Bandyopadhyay, R.; Ortega-Beltran, A. Present status and perspective on the future use of aflatoxin biocontrol products. Agronomy 2020, 10, 491. [Google Scholar] [CrossRef] [Green Version]
- Mauro, A.; Battilani, P.; Cotty, P.J. Atoxigenic Aspergillus flavus endemic to Italy for biocontrol of aflatoxins in maize. BioControl 2015, 60, 125–134. [Google Scholar] [CrossRef]
- Mauro, A.; Garcia-Cela, E.; Pietri, A.; Cotty, P.J.; Battilani, P. Biological control products for aflatoxin prevention in Italy: Commercial field evaluation of atoxigenic Aspergillus flavus active ingredients. Toxins 2018, 10, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrone, G.; Ferrara, M.; Medina, A.; Pascale, M.; Magan, N. Toxigenic fungi and mycotoxins in a climate change scenario: Ecology, genomics, distribution, prediction and prevention of the risk. Microorganisms 2020, 8, 1496. [Google Scholar] [CrossRef] [PubMed]
- Anfossi, L.; Baggiani, C.; Giovannoli, C.; D’Arco, G.; Passini, C.; Giraudi, G. Occurrence of aflatoxin M 1 in Italian cheese: Results of a survey conducted in 2010 and correlation with manufacturing, production season, milking animals, and maturation of cheese. Food Control 2012, 25, 125–130. [Google Scholar] [CrossRef]
- Battilani, P.; Toscano, P.; Van Der Fels-Klerx, H.J.; Moretti, A.; Camardo Leggieri, M.; Brera, C.; Rortais, A.; Goumperis, T.; Robinson, T. Aflatoxin B1 contamination in maize in Europe increases due to climate change. Sci. Rep. 2016, 6, 24328. [Google Scholar] [CrossRef] [Green Version]
- Girolami, F.; Barbarossa, A.; Badino, P.; Ghadiri, S.; Cavallini, D.; Zaghini, A.; Nebbia, C. Effects of turmeric powder on aflatoxin M1 and aflatoxicol excretion in milk from dairy cows exposed to aflatoxin B1 at the EU maximum tolerable levels. Toxins 2022, 14, 430. [Google Scholar] [CrossRef]
- Walte, H.-G.; Knappstein, K.; Maul, R.; Steinberg, P. Re-Evaluation of Aflatoxin M1 transfer into milk of high-yielding cows considering ration composition. J. Anim. Feed Sci. 2022, 31, 343–351. [Google Scholar] [CrossRef]
- Anonymous Biocontrollo Aflatossine, La Soluzione Contro Le Aflatossine. Available online: https://www.corteva.it/content/dam/dpagco/corteva/eu/it/it/files/cp/brochures/DF-AF-X1_2022-volantino.pdf (accessed on 17 December 2022).
- Anonymous Agente Di Biocontrollo per La Riduzione Delle Aflatossine Nel Mais. Available online: https://www.granapadano.it/it-it/af-x1-2022-agente-di-biocontrollo-per-la-riduzione-delle-aflatossine-nel-mais (accessed on 17 December 2022).
- Mehl, H.L.; Cotty, P.J. Nutrient environments influence competition among Aspergillus flavus genotypes. Appl. Environ. Microbiol. 2013, 79, 1473–1480. [Google Scholar] [CrossRef] [Green Version]
- Moore, G.G.; Lebar, M.D.; Carter-Wientjes, C.H.; Gilbert, M.K. The potential role of fungal volatile organic compounds in Aspergillus flavus biocontrol efficacy. Biol. Control 2021, 160, 104686. [Google Scholar] [CrossRef]
- Moore, G.G.; Lebar, M.D.; Carter-Wientjes, C.H. The role of extrolites secreted by nonaflatoxigenic Aspergillus flavus in biocontrol efficacy. J. Appl. Microbiol. 2019, 126, 1257–1264. [Google Scholar] [CrossRef]
- Agbetiameh, D.; Ortega-Beltran, A.; Awuah, R.; Atehnkeng, J.; Elzein, A.; Cotty, P.; Bandyopadhyay, R. Field efficacy of two atoxigenic biocontrol products for mitigation of aflatoxin contamination in maize and groundnut in Ghana. Biol. Control 2020, 150, 104351. [Google Scholar] [CrossRef]
- Bandyopadhyay, R.; Ortega-Beltran, A.; Konlambigue, M.; Kaptoge, L.; Falade, T.D.O.; Cotty, P.J. Development and scale-up of bioprotectants to keep staple foods safe from aflatoxin contamination in Africa. In Microbial Bioprotectants for Plant Disease Management; Kohl, J., Ravensverg, W.J., Eds.; Burleigh Dodds Science Publishing: Cambridge, UK, 2022; pp. 587–628. [Google Scholar]
- Mahuku, G.; Mauro, A.; Pallangyo, B.; Nsami, E.; Boni, S.B.; Koyano, E.; Mponda, O.; Ortega-Beltran, A.; Atehnkeng, J.; Aquiline, F.; et al. Atoxigenic-based technology for biocontrol of aflatoxin in maize and groundnuts for Tanzania. World Mycotoxin J. 2022, 10, 59–73. [Google Scholar] [CrossRef]
- Cotty, P.J.; Mellon, J.E. Ecology of aflatoxin producing fungi and biocontrol of aflatoxin contamination. Mycotoxin Res. 2006, 22, 110–117. [Google Scholar] [CrossRef]
- Doster, M.A.; Cotty, P.J.; Michailides, T.J. Evaluation of the atoxigenic Aspergillus flavus strain AF36 in pistachio orchards. Plant Dis. 2014, 98, 948–956. [Google Scholar] [CrossRef] [Green Version]
- Bandyopadhyay, R.; Ortega-Beltran, A.; Akande, A.; Mutegi, C.; Atehnkeng, J.; Kaptoge, L.; Senghor, A.L.; Adhikari, B.N.; Cotty, P.J. Biological control of aflatoxins in Africa: Current status and potential challenges in the face of climate change. World Mycotoxin J. 2016, 9, 771–789. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Beltran, A.; Kaptoge, L.; Senghor, A.L.; Aikore, M.O.S.; Jarju, P.; Momanyi, H.; Konlambigue, M.; Falade, T.D.O.; Bandyopadhyay, R. Can it be all more simple? Manufacturing aflatoxin biocontrol products using dry spores of atoxigenic isolates of Aspergillus flavus as active ingredients. Microb. Biotechnol. 2021, 15, 901–914. [Google Scholar] [CrossRef]
- Cotty, P.J.; Jaime-Garcia, R. Influences of climate on aflatoxin producing fungi and aflatoxin contamination. Int. J. Food Microbiol. 2007, 119, 109–115. [Google Scholar] [CrossRef]
- Camardo Leggieri, M.; Toscano, P.; Battilani, P. Predicted aflatoxin B1 increase in Europe due to climate change: Actions and Reactions at global level. Toxins 2021, 13, 292. [Google Scholar] [CrossRef]
- Abbas, H.K.; Accinelli, C.; Thomas Shier, W. biological control of aflatoxin contamination in U.S. crops and the use of bioplastic formulations of Aspergillus flavus biocontrol strains to optimize application strategies. J. Agric. Food Chem. 2017, 65, 7081–7087. [Google Scholar] [CrossRef]
- Ching’anda, C.; Atehnkeng, J.; Bandyopadhyay, R.; Callicott, K.A.; Orbach, M.J.; Mehl, H.L.; Cotty, P.J. Temperature influences on interactions among aflatoxigenic species of Aspergillus section Flavi during maize colonization. Front. Fungal Biol. 2021, 2, 720276. [Google Scholar] [CrossRef]
- Atehnkeng, J.; Ojiambo, P.S.; Cotty, P.J.; Bandyopadhyay, R. Field efficacy of a mixture of atoxigenic Aspergillus flavus Link: Fr Vegetative Compatibility Groups in preventing aflatoxin contamination in maize (Zea mays L.). Biol. Control 2014, 72, 62–70. [Google Scholar] [CrossRef]
- Das, M.K.; Ehrlich, K.C.; Cotty, P.J. Use of pyrosequencing to quantify incidence of a specific Aspergillus flavus strain within complex fungal communities associated with commercial cotton crops. Phytopathology 2008, 98, 282–288. [Google Scholar] [CrossRef] [Green Version]
- Grubisha, L.C.; Cotty, P.J. Twenty-Four microsatellite markers for the aflatoxin-producing fungus Aspergillus flavus. Mol. Ecol. Resour. 2009, 9, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Callicott, K.A.; Mutegi, C.; Bandyopadhyay, R.; Cotty, P.J. Distribution of active ingredients of a commercial aflatoxin biocontrol product in naturally occurring fungal communities across Kenya. Microb. Biotechnol. 2021, 14, 1331–1342. [Google Scholar] [CrossRef] [PubMed]
- Ortega, S.F.; Siciliano, I.; Prencipe, S.; Gullino, M.L.; Spadaro, D. Development of PCR, LAMP and QPCR assays for the detection of aflatoxigenic strains of Aspergillus flavus and A. parasiticus in hazelnut. Toxins 2020, 12, 757. [Google Scholar] [CrossRef]
- Rodríguez, A.; Rodríguez, M.; Luque, M.I.; Martín, A.; Córdoba, J.J. Real-Time PCR assays for detection and quantification of aflatoxin-producing molds in foods. Food Microbiol. 2012, 31, 89–99. [Google Scholar] [CrossRef]
- Shapira, R.; Paster, N.; Eyal, O.; Menasherov, M.; Mett, A.; Salomon, R. Detection of aflatoxigenic molds in grains by PCR. Appl. Environ. Microbiol. 1996, 62, 3270–3273. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Lopez, M.T.; Luo, Y.; Ortega-Beltran, A.; Jaime-Garcia, R.; Moral, J.; Michailides, T.J. Quantification of the aflatoxin biocontrol strain Aspergillus flavus AF36 in soil and in nuts and leaves of pistachio by real-time PCR. Plant Dis. 2021, 105, 1657–1665. [Google Scholar] [CrossRef]
- Callicott, K.A.; Cotty, P.J. Method for monitoring deletions in the aflatoxin biosynthesis gene cluster of Aspergillus flavus with multiplex PCR. Lett. Appl. Microbiol. 2015, 60, 60–65. [Google Scholar] [CrossRef]
- Mauro, A.; Battilani, P.; Callicott, K.A.; Giorni, P.; Pietri, A.; Cotty, P.J. Structure of an Aspergillus flavus population from maize kernels in Northern Italy. Int. J. Food Microbiol. 2013, 162, 1–7. [Google Scholar] [CrossRef]
- Cotty, P.J.; Antilla, L.; Wakelyn, P.J. Competitive exclusion of aflatoxin producers: Farmer-driven research and development. In Biological Control: A Global Perspective; Vincent, C., Goettel, M.S., Lazarovits, G., Eds.; CAB International: Cambridge, MA, USA, 2007; pp. 241–253. [Google Scholar]
- Molo, M.S.; White, J.B.; Cornish, V.; Gell, R.M.; Baars, O.; Singh, R.; Carbone, M.A.; Isakeit, T.; Wise, K.A.; Woloshuk, C.P.; et al. Asymmetrical lineage introgression and recombination in populations of Aspergillus flavus: Implications for biological control. PLoS ONE 2022, 10, e0276556. [Google Scholar] [CrossRef] [PubMed]
- Dorner, J.W. Biological control of aflatoxin contamination of crops. J. Toxicol.—Toxin Rev. 2004, 23, 425–450. [Google Scholar] [CrossRef]
- Dowd, P.F. Insect Management to Facilitate Preharvest Mycotoxin Management. Toxin Rev. 2003, 22, 327–350. [Google Scholar] [CrossRef]
- Abbas, H.K.; Zablotowicz, R.M.; Locke, M.A. Spatial variability of Aspergillus flavus soil populations under different crops and corn grain colonization and aflatoxins. Can. J. Bot. 2004, 82, 1768–1775. [Google Scholar] [CrossRef] [Green Version]
- Jaime-Garcia, R.; Cotty, P.J. Spatial distribution of Aspergillus flavus and its toxigenic strains on commercial cottonseed from south texas and its relationship to aflatoxin contamination. Plant Pathol. 2006, 55, 358–366. [Google Scholar] [CrossRef]
- Jaime-Garcia, R.; Cotty, P.J. Crop rotation and soil temperature influence the community structure of Aspergillus flavus in soil. Soil Biol. Biochem. 2010, 42, 1842–1847. [Google Scholar] [CrossRef]
- Abbas, H.K.; Accinelli, C.; Zablotowicz, R.M.; Abel, C.A.; Bruns, H.A.; Dong, Y.; Shier, W.T. Dynamics of mycotoxin and Aspergillus flavus levels in aging Bt and non-Bt corn residues under mississippi no-till conditions. J. Agric. Food Chem. 2008, 56, 7578–7585. [Google Scholar] [CrossRef]
- Donner, M.; Lichtemberg, P.S.F.; Doster, M.; Picot, A.; Cotty, P.J.; Puckett, R.D.; Michailides, T.J. Community structure of Aspergillus flavus and A. parasiticus in major almond-producing areas of California, United States. Plant Dis. 2015, 99, 1161–1169. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, K.B.; Longing, S.D.; West, C.P. Soil microbial communities in corn fields treated with atoxigenic Aspergillus flavus. Soil Syst. 2020, 4, 35. [Google Scholar] [CrossRef]
- Reis, T.A.; Oliveira, T.D.; Zorzete, P.; Faria, P.; Corrêa, B. A Non-Toxigenic Aspergillus flavus strain prevents the spreading of Fusarium verticillioides and fumonisins in maize. Toxicon 2020, 181, 6–8. [Google Scholar] [CrossRef]
- Cotty, P.J. Influence of field application of an atoxigenic strain of Aspergillus flavus on the population of A.flavus infecting cotton bolls and on the aflatoxin content of cottonseed. Biol. Control 1994, 84, 1270–1277. [Google Scholar]
- Jaime-Garcia, R.; Leisner, L.; Antilla, L.; Mehl, H.L.; Cotty, P.J. Area-wide programs for aflatoxin mitigation: Treatments to cotton can be cost effective. In Proceedings of the Beltwide Cotton Production Research Conference, Dallas, TX, USA, 4–6 January 2017; pp. 327–350. [Google Scholar]
- Meier, U. Growth Stage of Mono- and Dicoyle-Donous Plants. BBCH Monograph. In Growth Stage of Mono- and Dicoyle-Donous Plants. BBCH Monograph; Meier, U., Ed.; Julius Kuhn-Institute (JKI): Quedlinburg, Germany, 2018; p. 204. [Google Scholar]
- Cotty, P.J. Aflatoxin-producing potential of communities of Aspergillus Section Flavi from cotton producing areas in the United States. Mycol. Res. 1997, 101, 698–704. [Google Scholar] [CrossRef] [Green Version]
- Senghor, L.A.; Ortega-Beltran, A.; Atehnkeng, J.; Callicott, K.A.; Cotty, P.J.; Bandyopadhyay, R. The Atoxigenic biocontrol product aflasafe SN01 is a valuable tool to mitigate aflatoxin contamination of both maize and groundnut cultivated in Senegal. Plant Dis. 2019, 104, 510–520. [Google Scholar] [CrossRef]
- Cotty, P.J. Comparison of four media for the isolation of Aspergillus flavus group fungi. Mycopathologia 1994, 125, 157–162. [Google Scholar] [CrossRef]
- Klich, M.A.; Pitt, J. Differentiation of Aspergillus flavus from A. parasiticus and other closely related species. Trans. Br. Mycol. Soc. 1988, 91, 99–108. [Google Scholar] [CrossRef]
- Probst, C.; Bandyopadhyay, R.; Price, L.E.; Cotty, P.J. Identification of atoxigenic Aspergillus flavus isolates to reduce aflatoxin contamination of maize in Kenya. Plant Dis. 2011, 95, 212–218. [Google Scholar] [CrossRef] [Green Version]
- Probst, C.; Cotty, P.J. Relationships between in vivo and in vitro aflatoxin production: Reliable prediction of fungal ability to contaminate maize with aflatoxins. Fungal Biol. 2012, 116, 503–510. [Google Scholar] [CrossRef]
- Bayman, P.; Cotty, P.J. Vegetative Compatibility and Genetic Diversity in the Aspergillus flavus Population of a Single Field. Can. J. Bot. 1991, 69, 1707–1711. [Google Scholar] [CrossRef]
- Cotty, P.J.; Taylor, D.R. Influence of complementation medium composition on vegetative compatibility analyses of Aspergillus flavus. In Proceedings of the American Phytopathological Society Annual Meeting, Charlotte, NC, USA, 9–13 August 2003; p. 18. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Sampling Year | Area N 1 | District 2 | Municipality 3 | Latitude | Longitude | Treatment 4 | Soil Type | Rotation 1 5 | Rotation 2 6 | Tillage | Stalk Burial |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2020 | 1 | Rovigo | Occhiobello | 44.9297388 | 11.62801768 | Untreated | Silt | Maize | Tomato | Till | No |
| 2020 | 1 | Rovigo | Occhiobello | 44.943139 | 11.567212 | n-2 | Sand | Maize | Maize | Till | No |
| 2020 | 1 | Rovigo | Occhiobello | 44.951586 | 11.547809 | n-1&n-2 | Silt | Maize | Maize | Till | No |
| 2020 | 1 | Rovigo | Occhiobello | 44.955789 | 11.554532 | n-1 | Silt | Maize | Tomato | Till | No |
| 2020 | 2 | Rovigo | Occhiobello | 44.969438 | 11.70443 | Untreated | Silt | Wheat | Pea | No till | No |
| 2020 | 2 | Rovigo | Occhiobello | 44.93645466 | 11.62733917 | n-2 | Clay | Wheat | Maize | No till | No |
| 2020 | 2 | Rovigo | Fiesso Umbertino | 44.955438 | 11.589537 | n-1&n-2 | Silt | Maize | Maize | No till | Yes |
| 2020 | 2 | Rovigo | Occhiobello | 44.94879764 | 11.60269669 | n-1 | Silt | Maize | Wheat | Till | Yes |
| 2020 | 3 | Rovigo | Occhiobello | 44.94469842 | 11.49092054 | Untreated | Clay | Wheat | Wheat | Till | No |
| 2020 | 3 | Rovigo | Occhiobello | 44.94834217 | 11.59027712 | n-2 | Sand | Wheat | Maize | Till | Yes |
| 2020 | 3 | Rovigo | Fiesso Umbertino | 44.973695 | 11.630536 | n-1&n-2 | Silt | Maize | Maize | Till | Yes |
| 2020 | 3 | Rovigo | Occhiobello | 44.96235125 | 11.6690985 | n-1 | Silt | Maize | Soybean | Till | Yes |
| 2021 | 4 | Modena | Finale Emilia | 44.81483 | 11.217337 | Untreated | Clay | Wheat | Maize | Till | No |
| 2021 | 4 | Modena | Finale Emilia | 44.829477 | 11.094888 | n-2 | Silt | Wheat | Maize | Till | No |
| 2021 | 4 | Modena | Finale Emilia | 44.835933 | 11.27449 | n-1&n-2 | Sand | Maize | Maize | Till | No |
| 2021 | 4 | Modena | Finale Emilia | 44.86689 | 11.174496 | n-1 | Silt | Maize | Wheat | Till | No |
| 2021 | 5 | Rovigo | Occhiobello | 44.98778 | 11.698756 | Untreated | Clay | Soybean | Wheat | Till | No |
| 2021 | 5 | Rovigo | Occhiobello | 44.931735 | 11.629024 | n-2 | Silt | Soybean | Maize | Till | No |
| 2021 | 5 | Rovigo | Occhiobello | 44.958125 | 11.627436 | n-1&n-2 | Silt | Maize | Maize | Till | Yes |
| 2021 | 5 | Rovigo | Occhiobello | 44.971765 | 11.721178 | n-1 | Silt | Maize | Wheat | Till | Yes |
| 2021 | 6 | Padova | Noale | 45.54925 | 12.052739 | Untreated | Silt | Soybean | Soybean | Till | Yes |
| 2021 | 6 | Padova | Noale | 45.549233 | 12.05297 | n-2 | Silt | Soybean | Maize | Till | Yes |
| 2021 | 6 | Padova | Noale | 45.553166 | 12.045033 | n-1&n-2 | Clay | Maize | Maize | Till | Yes |
| 2021 | 6 | Padova | Noale | 45.552759 | 12.048152 | n-1 | Clay | Maize | Soybean | Till | Yes |
| 2021 | 7 | Venezia | Scorzè | 45.563136 | 12.10695 | Untreated | Silt | Soybean | Soybean | Till | Yes |
| 2021 | 7 | Venezia | Scorzè | 45.569842 | 12.099649 | n-2 | Silt | Soybean | Maize | Till | Yes |
| 2021 | 7 | Venezia | Scorzè | 45.565978 | 12.09769 | n-1&n-2 | Silt | Maize | Maize | Till | No |
| 2021 | 7 | Venezia | Scorzè | 45.56353 | 12.10323 | n-1 | Silt | Maize | Soybean | Till | No |
| Sample Year | Treatment Year | N. of Isolates | Replicates 1 | Total Fungi (CFU/g) | A. flavus (CFU/g) | % AF- 2 | IT006 on A. flavus % 3 | IT006 on A. flavus Atox % 3 |
|---|---|---|---|---|---|---|---|---|
| 2020 | * | * | NS | NS | NS | |||
| Untreated | 39 | 3 | 702 b | 51 c | 77 | 37 | 48 | |
| 2018 | 44 | 3 | 704 b | 163 ab | 75 | 41 | 56 | |
| 2018 + 2019 | 42 | 3 | 778 b | 190 a | 93 | 72 | 77 | |
| 2019 | 44 | 3 | 2688 a | 86 b | 86 | 77 | 88 | |
| 2021 | NS | NS | NS | ** | ** | |||
| Untreated | 60 | 4 | 1229 | 96 | 38 | 12 b | 23 b | |
| 2019 | 54 | 4 | 964 | 63 | 75 | 62 a | 86 a | |
| 2019 + 2020 | 60 | 4 | 863 | 84 | 68 | 57 a | 79 a | |
| 2020 | 56 | 4 | 943 | 138 | 72 | 54 a | 74 ab | |
| 2020, 2021 Combined | NS | * | * | ** | ** | |||
| Untreated | 99 | 7 | 1003 | 77 b | 55 b | 23 b | 34 b | |
| Treated 1 yr | 198 | 14 | 1271 | 111 a | 77 ab | 58 a | 76 a | |
| Treated 2 yrs | 102 | 7 | 826 | 130 a | 79 a | 63 a | 78 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouadhene, M.A.; Ortega-Beltran, A.; Sanna, M.; Cotty, P.J.; Battilani, P. Multiple Year Influences of the Aflatoxin Biocontrol Product AF-X1 on the A. flavus Communities Associated with Maize Production in Italy. Toxins 2023, 15, 184. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins15030184
Ouadhene MA, Ortega-Beltran A, Sanna M, Cotty PJ, Battilani P. Multiple Year Influences of the Aflatoxin Biocontrol Product AF-X1 on the A. flavus Communities Associated with Maize Production in Italy. Toxins. 2023; 15(3):184. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins15030184
Chicago/Turabian StyleOuadhene, Mohamed Ali, Alejandro Ortega-Beltran, Martina Sanna, Peter J. Cotty, and Paola Battilani. 2023. "Multiple Year Influences of the Aflatoxin Biocontrol Product AF-X1 on the A. flavus Communities Associated with Maize Production in Italy" Toxins 15, no. 3: 184. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins15030184