Potential Biotechnological Applications of Venoms from the Viperidae Family in Central America for Thrombosis

 , ,
, ,  , , and
, , and 
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genera | Described Species | Principal Protein Families Percentages (%) | Reference | |||
|---|---|---|---|---|---|---|
| SVMPs | PLA2s | SVSPs | LAAOs | |||
| Agkistrodon | A. bilineatus | 21–33 | 32–42 | 9–23 | 0.8–9 | [12,13,14,15,16] |
| Atropoides | A. picadoi | 66 | 9.5 | 13.5 | 2.2 | [17,18,19,20] |
| Bothriechis | B. schlegelii | 17.7 | 43.8 | 5.8 | 8.9 | [21,22,23] |
| Bothrops | B. asper | 41 | 29 | 18 | 9 | [24,25] |
| Cerrophidion | C. godmani | 32.8 | 23.4 | 19.1 | <10 | [26,27] |
| Crotalus | C. simus | 28 | 22 | 30 | 17 | [10] |
| Lachesis | L. melanocephala | 49.2 | 13.4 | 21 | 3.6 | [28,29] |
| Metlapilcoatlus | M. mexicanus | 18.2 | 36.5 | 22 | 9.1 | [20,30] |
| Porthidium | P. nasutum | 52.1 | 11.6 | 9.6 | 9.8 | [31] |
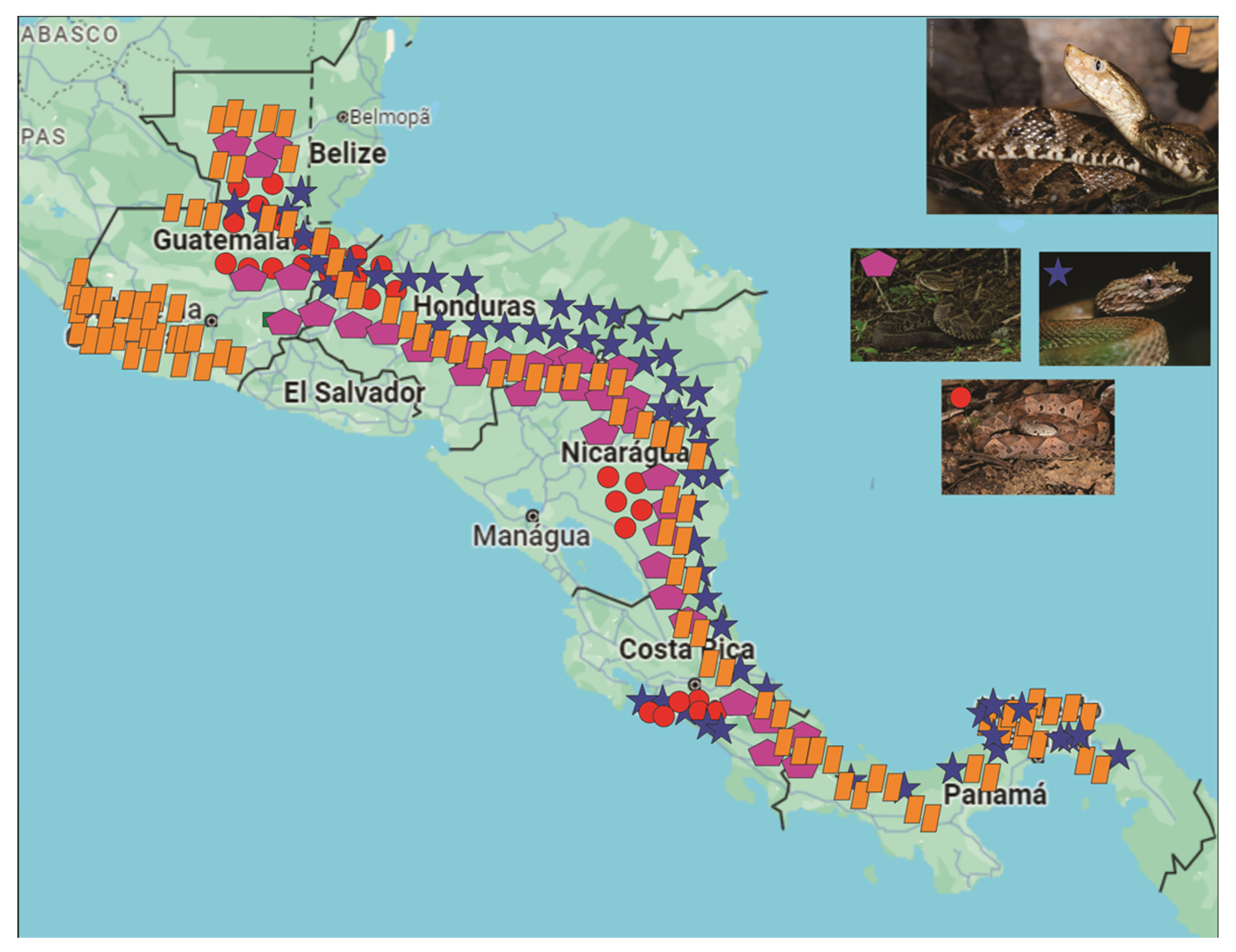
2. Viperidae Family
2.1. Agkistrodon
Agkistrodon bilineatus
2.2. Atropoides
Atropoides picadoi
2.3. Bothriechis
2.3.1. Bothriechis aurifer
2.3.2. Bothriechis bicolor
2.3.3. Bothriechis thalassinus
2.3.4. Bothriechis schlegelii
2.4. Bothrops
Bothrops asper
2.5. Cerrophidion
Cerrophidion godmani
2.6. Crotalus
2.6.1. Crotalus simus
2.6.2. Crotalus tzabcan
2.7. Lachesis
2.7.1. Lachesis stenhophrys
2.7.2. Lachesis melanocephala
2.7.3. Lachesis acrochorda
2.8. Metlapilcoatlus
Metlapilcoatlus mexicanus
2.9. Porthidium
2.9.1. Porthidium nasutum
2.9.2. Porthidium ophryomegas
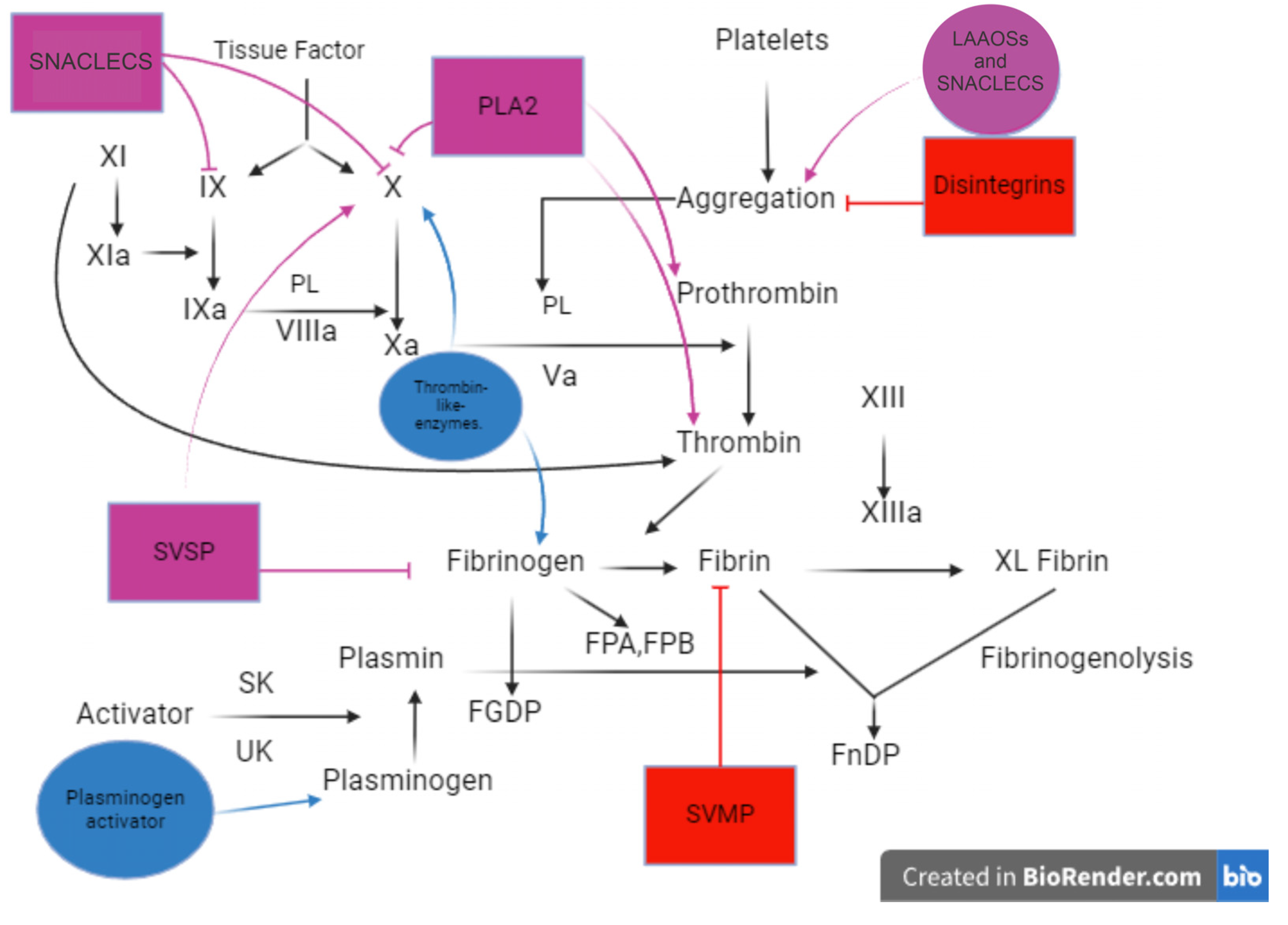
3. Potential Biotechnological Applications of Venom Components from Viperidae for Thrombosis
3.1. Thrombosis
3.2. Venom Components
3.2.1. Metalloproteases and Disintegrins
Jararhagin
Crotalin
B. asper Venom Metall Oproteinases That Interfere with Blood Compounds
Tzabcanin
Jararacin/Jarastatin
3.2.2. Phospholipases A2 (PLA2s)
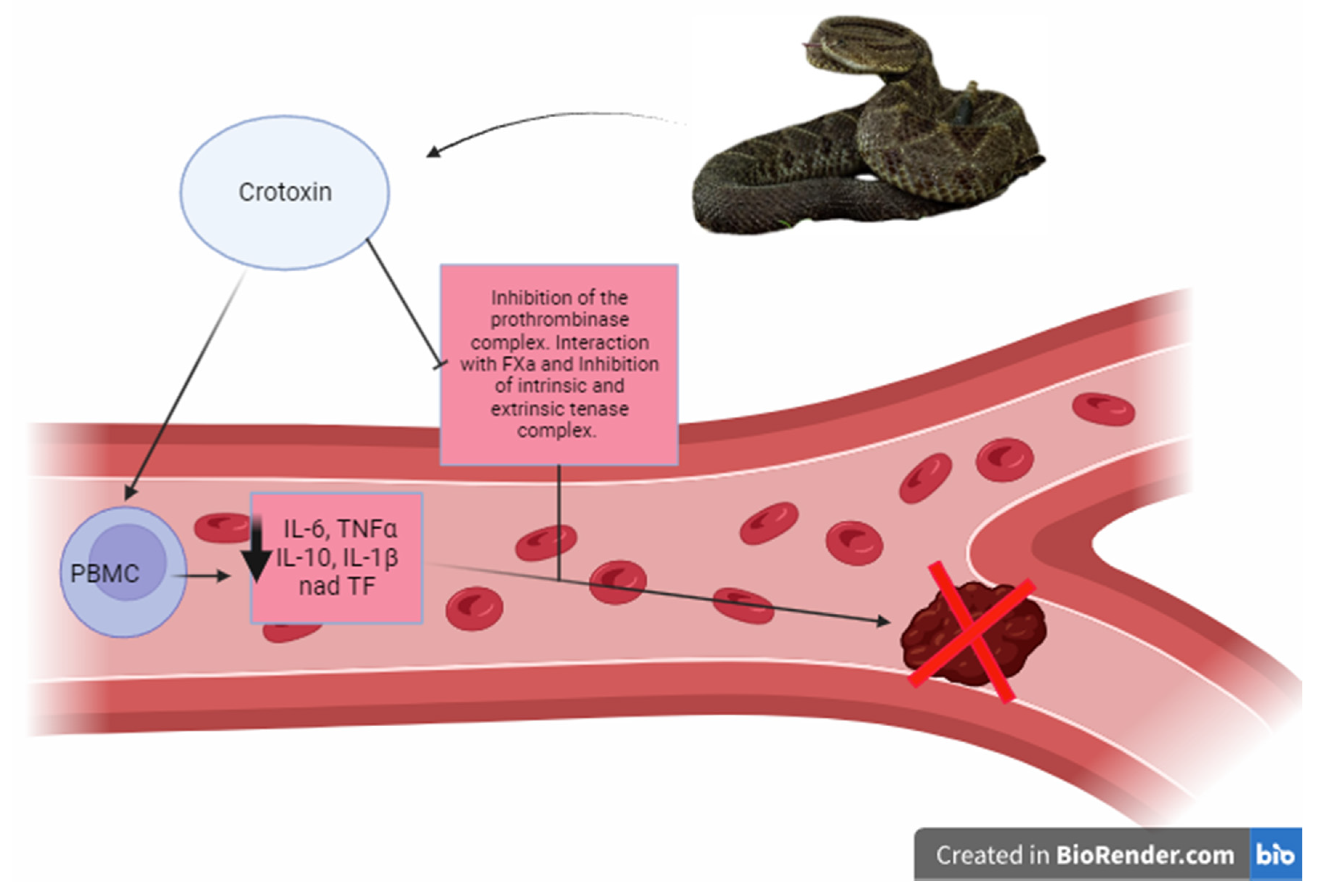
Crotoxin
3.2.3. Serine Proteases
Batroxobin
Plasminogen Activators
3.2.4. L-Amino-Acid Oxidases
3.2.5. Snaclecs (Snake C-Type Lectin-Like Proteins)
Bothrojaracin
IX/X-bp (Factors IX- and X-Binding Protein)
Rhodocetin
Glycoprotein Ib-Binding Protein
3.2.6. Bioactive Peptides
Crotamine
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morrone, J.J. Biogeographic Areas and Transition Zones of Latin America and the Caribbean Islands Based on Panbiogeographic and Cladistic Analyses of the Entomofauna. Annu. Rev. Entomol. 2006, 51, 467–494. [Google Scholar] [CrossRef]
- Bolanos, R. Las Serpientes Venenosas de Centroamerica y El Problema Del Ofidismo.Primera Parte. Aspectos Zoologicos, Epidemiologicos y Biomedicos. Rev. Costarric. Cienc. Méd. 1982, 3, 165–185. [Google Scholar]
- Campbell, J.; Lamar, W. The Venomous Reptiles of the Western Hemisphere, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 2004. [Google Scholar]
- Lomeo, R.D.S.; Gonçalves, A.P.D.F.; Da Silva, C.N.; De Paula, A.T.; Costa Santos, D.O.; Fortes-Dias, C.L.; Gomes, D.A.; De Lima, M.E. Crotoxin from Crotalus Durissus Terrificus Snake Venom Induces the Release of Glutamate from Cerebrocortical Synaptosomes via N and P/Q Calcium Channels. Toxicon 2014, 85, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Sartim, M.A.; Menaldo, D.L.; Sampaio, S.V. Immunotherapeutic Potential of Crotoxin: Anti-Inflammatory and Immunosuppressive Properties. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 1–13. [Google Scholar] [CrossRef] [PubMed]
- De Araújo Pimenta, L.; de Almeida, M.E.S.; Bretones, M.L.; Cirillo, M.C.; Curi, R.; Sampaio, S.C. Crotoxin Promotes Macrophage Reprogramming towards an Antiangiogenic Phenotype. Sci. Rep. 2019, 9, 4281. [Google Scholar] [CrossRef] [PubMed]
- Saviola, A.J.; Burns, P.D.; Mukherjee, A.K.; Mackessy, S.P. The Disintegrin Tzabcanin Inhibits Adhesion and Migration in Melanoma and Lung Cancer Cells. Int. J. Biol. Macromol. 2016, 88, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Saviola, A.J.; Modahl, C.M.; Mackessy, S.P. Disintegrins of Crotalus Simus Tzabcan Venom: Isolation, Characterization and Evaluation of the Cytotoxic and Anti-Adhesion Activities of Tzabcanin, a New RGD Disintegrin. Biochimie 2015, 116, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Vu, T.T.; Stafford, A.R.; Leslie, B.A.; Kim, P.Y.; Fredenburgh, J.C.; Weitz, J.I. Batroxobin Binds Fibrin with Higher Affinity and Promotes Clot Expansion to a Greater Extent than Thrombin. J. Biol. Chem. 2013, 288, 16862–16871. [Google Scholar] [CrossRef] [PubMed]
- Neri-Castro, E.; Bénard-Valle, M.; Paniagua, D.; Boyer, L.V.; Possani, L.D.; López-Casillas, F.; Olvera, A.; Romero, C.; Zamudio, F.; Alagón, A. Neotropical Rattlesnake (Crotalus Simus) Venom Pharmacokinetics in Lymph and Blood Using an Ovine Model. Toxins 2020, 12, 455. [Google Scholar] [CrossRef]
- Nielsen, V.G.; Frank, N.; Afshar, S. De Novo Assessment and Review of Pan-American Pit Viper Anticoagulant and Procoagulant Venom Activities via Kinetomic Analyses. Toxins 2019, 11, 94. [Google Scholar] [CrossRef]
- Gloyd, H.K.; Conant, R. Snakes of the Agkistrodon Complex—A Monographic Review. Amphib.-Reptil. 1990, 12, 1–614. [Google Scholar]
- Porras, L.; Wilson, L.; Schuett, G.; Reiserer, R. A Taxonomic Reevaluation and Conservation Assessment of the Common Cantil, Agkistrodon Bilineatus (Squamata: Viperidae): A Race against Time. Amphib. Reptile Conserv. 2013, 7, 48–73. [Google Scholar]
- Smetsers, P. Agkistrodon Bilineatus—Gunther, the Tropical Moccasin. Litteratura Serpentium Engl. Ed. 1993, 13, 38–40. [Google Scholar]
- Mérida, J.E.; Cruz, G.A. Ampliación de Ámbito de La Serpiente Agkistrodon Bilineatus (Serpentes: Viperidae), En El Sureste de Honduras. UNED Res. J. 2012, 4, 41–43. [Google Scholar] [CrossRef]
- Lomonte, B.; Tsaia, W.-C.; Ureña-Diaz, J.M.; Sanz, L.; Mora Obando, D.; Sánchez, E.E.; Bryan, G.F.; Gutiérrez, J.M.; Gibb, L.; Sovic, G.; et al. Venomics of New World Pit Vipers: Genus-Wide Comparisons of Venom Proteomes across Agkistrodon. J. Proteom. 2015, 96, 103–116. [Google Scholar] [CrossRef]
- Wüster, W.; Peppin, L.; Pook, C.E.; Walker, D.E. A Nesting of Vipers: Phylogeny and Historical Biogeography of the Viperidae (Squamata: Serpentes). Mol. Phylogenet. Evol. 2008, 49, 445–459. [Google Scholar] [CrossRef]
- Antúnez, J.; Fernández, J.; Lomonte, B.; Angulo, Y.; Sanz, L.; Pérez, A.; Calvete, J.J.; Gutiérrez, J.M. Antivenomics of Atropoides Mexicanus and Atropoides Picadoi Snake Venoms: Relationship to the Neutralization of Toxic and Enzymatic Activities. J. Venom. Res. 2010, 1, 8–17. [Google Scholar]
- Jadin, R.C.; Gutberlet, R.L.; Smith, E.N. Phylogeny, Evolutionary Morphology, and Hemipenis Descriptions of the Middle American Jumping Pitvipers (Serpentes: Crotalinae: Atropoides). J. Zool. Syst. Evol. Res. 2010, 48, 360–365. [Google Scholar] [CrossRef]
- Angulo, Y.; Escolano, J.; Lomonte, B.; Gutiérrez, J.M.; Sanz, L.; Calvete, J.J. Snake Venomics of Central American Pitvipers: Clues for Rationalizing the Distinct Envenomation Profiles of Atropoides Nummifer and Atropoides Picadoi. J. Proteome Res. 2008, 7, 708–719. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.A.; Smith, E.N. A New Species of Arboreal Pitviper from the Atlantic Versant of Northern Central America. Rev. Biol. Trop. 2000, 48, 1001–1013. [Google Scholar] [PubMed]
- Pla, D.; Sanz, L.; Sasa, M.; Acevedo, M.E.; Dwyer, Q.; Durban, J.; Pérez, A.; Rodriguez, Y.; Lomonte, B.; Calvete, J.J. Proteomic Analysis of Venom Variability and Ontogeny across the Arboreal Palm-Pitvipers (Genus Bothriechis). J. Proteomics 2017, 152, 1–12. [Google Scholar] [CrossRef]
- Lomonte, B.; Escolano, J.; Fernández, J.; Sanz, L.; Angulo, Y.; Gutiérrez, J.M.; Calvete, J.J. Snake Venomics and Antivenomics of the Arboreal Neotropical Pitvipers Bothriechis Lateralis and Bothriechis Schlegelii. J. Proteome Res. 2008, 7, 2445–2457. [Google Scholar] [CrossRef]
- Otero-Patiño, R. Epidemiological, Clinical and Therapeutic Aspects of Bothrops Asper Bites. Toxicon 2009, 54, 998–1011. [Google Scholar] [CrossRef]
- Mora-Obando, D.; Lomonte, B.; Pla, D.; Guerrero-Vargas, J.A.; Ayerbe-González, S.; Gutiérrez, J.M.; Sasa, M.; Calvete, J.J. Half a Century of Research on Bothrops Asper Venom Variation: Biological and Biomedical Implications. Toxicon. 2023, 221, 106983. [Google Scholar] [CrossRef]
- Jadin, R.C.; Townsend, J.H.; Castoe, T.A.; Campbell, J.A. Cryptic Diversity in Disjunct Populations of Middle American Montane Pitvipers: A Systematic Reassessment of Cerrophidion Godmani. Zool. Scr. 2012, 41, 455–470. [Google Scholar] [CrossRef]
- Durban, J.; Juárez, P.; Angulo, Y.; Lomonte, B.; Flores-Diaz, M.; Alape-Girón, A.; Sasa, M.; Sanz, L.; Gutiérrez, J.M.; Dopazo, J.; et al. Profiling the Venom Gland Transcriptomes of Costa Rican Snakes by 454 Pyrosequencing. BMC Genom. 2011, 12, 259. [Google Scholar] [CrossRef] [PubMed]
- Aragón Ortiz, F. Isolation and Partial Characterization of Lachesis Muta Melanocephala Coagulant Proteinase: Biochemical Parameters of the Venom. Rev. Biol. Trop. 1988, 36, 387–392. [Google Scholar] [PubMed]
- Madrigal, M.; Sanz, L.; Flores-Díaz, M.; Sasa, M.; Núñez, V.; Alape-Girón, A.; Calvete, J.J. Snake Venomics across Genus Lachesis. Ontogenetic Changes in the Venom Composition of Lachesis Stenophrys and Comparative Proteomics of the Venoms of Adult Lachesis Melanocephala and Lachesis Acrochorda. J. Proteom. 2012, 77, 280–297. [Google Scholar] [CrossRef]
- García-Osorio, B.; Lomonte, B.; Bénard-Valle, M.; López de León, J.; Román-Domínguez, L.; Mejía-Domínguez, N.R.; Lara-Hernández, F.; Alagón, A.; Neri-Castro, E. Ontogenetic Changes in the Venom of Metlapilcoatlus Nummifer, the Mexican Jumping Viper. Toxicon. 2020, 184, 204–214. [Google Scholar] [CrossRef]
- Bonilla-Porras, A.R.; Vargas, L.J.; Jimenez-Del-Rio, M.; Nuñez, V.; Velez-Pardo, C. Purification of Nasulysin-1: A New Toxin from Porthidium Nasutum Snake Venom That Specifically Induces Apoptosis in Leukemia Cell Model through Caspase-3 and Apoptosis-Inducing Factor Activation. Toxicon 2016, 120, 166–174. [Google Scholar] [CrossRef]
- Castoe, T.A.; Parkinson, C.L. Bayesian Mixed Models and the Phylogeny of Pitvipers (Viperidae: Serpentes). Mol. Phylogenet. Evol. 2006, 39, 91–110. [Google Scholar] [CrossRef]
- Arnold, S.J. Snakes, the Evolution of Mystery in Nature. Integr. Biol. Issues News Rev. 1998, 1, 76–77. [Google Scholar] [CrossRef]
- Pyron, R.A.; Burbrink, F.T.; Wiens, J.J. A Phylogeny and Revised Classification of Squamata, Including 4161 Species of Lizards and Snakes A Phylogeny and Revised Classification of Squamata, Including 4161 Species of Lizards and Snakes. BMC Evol. Biol. 2013, 13, 1–54. [Google Scholar] [CrossRef]
- Hargreaves, A.; Swain, M.; Logan, D.; Mulley, J. Testing the Toxicofera: Comparative Transcriptomics Casts Doubt on the Single, Early Evolution of the Reptile Venom System. Toxicon 2014, 92, 140–156. [Google Scholar] [CrossRef]
- Sachett, J.A.G.; da Silva, I.M.; Alves, E.C.; Oliveira, S.S.; Sampaio, V.S.; do Vale, F.F.; Romero, G.A.S.; dos Santos, M.C.; Marques, H.O.; Colombini, M.; et al. Poor Efficacy of Preemptive Amoxicillin Clavulanate for Preventing Secondary Infection from Bothrops Snakebites in the Brazilian Amazon: A Randomized Controlled Clinical Trial. PLoS Negl. Trop. Dis. 2017, 11, 1–21. [Google Scholar] [CrossRef]
- Bernal, J.C.C.; Bisneto, P.F.; Pereira, J.P.T.; Ibiapina, H.N.d.S.; Sarraff, L.K.S.; Monteiro-Júnior, C.; da Silva Pereira, H.; Santos, B.; de Moura, V.M.; de Oliveira, S.S.; et al. “Bad Things Come in Small Packages”: Predicting Venom-Induced Coagulopathy in Bothrops Atrox Bites Using Snake Ontogenetic Parameters. Clin. Toxicol. 2019, 58, 388–396. [Google Scholar] [CrossRef]
- Silva de Oliveira, S.; Campos Alves, E.; dos Santos Santos, A.; Freitas Nascimento, E.; Tavares Pereira, J.P.; Mendonça da Silva, I.; Sachett, J.; dos Santos Ibiapina, H.N.; Santos Sarraf, L.K.; Contreras Bernal, J.C.; et al. Bothrops Snakebites in the Amazon: Recovery from Hemostatic Disorders after Brazilian Antivenom Therapy. Clin. Toxicol. 2019, 58, 266–274. [Google Scholar] [CrossRef]
- Hui Wen, F.; Monteiro, W.M.; Moura da Silva, A.M.; Tambourgi, D.V.; Mendonça da Silva, I.; Sampaio, V.S.; dos Santos, M.C.; Sachett, J.; Ferreira, L.C.L.; Kalil, J.; et al. Snakebites and Scorpion Stings in the Brazilian Amazon: Identifying Research Priorities for a Largely Neglected Problem. PLoS Negl. Trop. Dis. 2015, 9, e0003701. [Google Scholar] [CrossRef] [PubMed]
- Toshiaki, N.; Yumiko, K.; Akihito, O.; Satoru, Y.; Yasushi, O.; Hisayoshi, S. Characterization and Amino-Terminal Sequence of Phospholipase A2-II from the Venom of Agkistrodon Bilineatus (Common Cantil). Int. J. Biochem. 1994, 26, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Román-Domínguez, L.; Neri-Castro, E.; Vázquez López, H.; García-Osorio, B.; Archundia, I.G.; Ortiz-Medina, J.A.; Petricevich, V.L.; Alagón, A.; Bénard-Valle, M. Biochemical and Immunochemical Characterization of Venoms from Snakes of the Genus Agkistrodon. Toxicon X 2019, 4, 100013. [Google Scholar] [CrossRef] [PubMed]
- Rojas, G.; Gutiérrez, J.M.; Gené, J.A.; Gómez, M.; Cerdas, L. Neutralization of Toxic and Enzyme Activities of 4 Venoms from Snakes of Guatemala and Honduras by the Polyvalent Antivenin Produced in Costa Rica. Rev. Biol. Trop. 1987, 35, 59–67. [Google Scholar] [CrossRef]
- Komori, Y.; Nikai, T.; Ohara, A.; Yagihashi, S.; Sugihara, H. Effect of Bilineobin, a Thrombin-like Proteinase from the Venom of Common Cantil (Agkistrodon Bilineatus). Toxicon 1993, 31, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Nikai, T.; Akihito, O.; Yumiko, K.; Fox, J.W.; Sugihara, H. Primary Structure of a Coagulant Enzyme, Bilineobin, from Agkistrodon Bilineatus Venom. Arch. Biochem. Biophys. 1995, 318, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Nakagaki, T.; Kazimll, A.L.; Kisiel, W. Isolation and Characterization of a Protein C Activator from Tropical Moccasin Venom. Thromb. Res. 1990, 58, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Moore, G.W. Snake Venoms in Diagnostic Hemostasis and Thrombosis. Semin. Thromb. Hemost. 2022, 48, 145–160. [Google Scholar] [CrossRef]
- Raymond, T. Hoser A New Genus of Jumping Pitviper from Middle America. Aus. J. Herp. 2012, 5698, 33–34. [Google Scholar]
- Jones, L.; Waite, C.; Neri-Castro, E.; Fry, B.G. Comparative Analysis of Alpha-1 Orthosteric-Site Binding by a Clade of Central American Pit Vipers (Genera Atropoides, Cerrophidion, Metlapilcoatlus, and Porthidium). Toxins 2023, 15, 487. [Google Scholar] [CrossRef]
- Camacho, E.; Villalobos, E.; Sanz, L.; Pérez, A.; Escalante, T.; Lomonte, B.; Calvete, J.J.; Gutiérrez, J.M.; Rucavado, A. Understanding Structural and Functional Aspects of PII Snake Venom Metalloproteinases: Characterization of BlatH1, a Hemorrhagic Dimeric Enzyme from the Venom of Bothriechis Lateralis. Biochimie 2014, 101, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Fernández, J.; Gutiérrez, J.M.; Calvete, J.J.; Sanz, L.; Lomonte, B. Characterization of a Novel Snake Venom Component: Kazal-Type Inhibitor-like Protein from the Arboreal Pitviper Bothriechis Schlegelii. Biochimie 2016, 125, 83–90. [Google Scholar] [CrossRef]
- Gómez, C.; Buitrago-González, W. Bothriechis Schlegelii (Berthold 1846). Catálogo Anfibios y Reptil. Colomb. 2017, 3, 1–11. [Google Scholar]
- Youngman, N.J.; Peng, Y.; Harris, R.; Lee, J.; Llinas, J.; Haworth, M.; Gillett, A.; Fry, B.G. Differential Coagulotoxic and Neurotoxic Venom Activity from Species of the Arboreal Viperid Snake Genus Bothriechis (Palm-Pitvipers). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2022, 256, 109326. [Google Scholar] [CrossRef] [PubMed]
- McCranie, J.R. The Snakes of Honduras. Systematics, Distribution, and Conservation. Amphib.-Reptil. 2011, 32, 569. [Google Scholar] [CrossRef]
- Townsend, J.H.; Medina-Flores, M.; Wilson, L.D.; Jadin, R.C.; Austin, J.D. A Relict Lineage and New Species of Green Palm-Pitviper (Squamata, Viperidae, Bothriechis) from the Chortís Highlands of Mesoamerica. Zookeys 2013, 298, 77–106. [Google Scholar] [CrossRef] [PubMed]
- Meza, P.; Almendáriz, A.; Yánez, M. Datos Sobre La Dieta de Bothriechis Schlegelii (Berthold, 1846)(Serpentes-Viperidae) En El Occidente Del Ecuador. Boletín Técnico Ser. Zoológica 2010, 9, 15–18. [Google Scholar]
- Angulo, Y.; Lomonte, B. Differential Susceptibility of C2C12 Myoblasts and Myotubes to Group II Phospholipase A2 Myotoxins from Crotalid Snake Venoms. Cell Biochem. Funct. 2005, 23, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Alape-Girón, A.; Flores-Díaz, M.; Sanz, L.; Madrigal, M.; Escolano, J.; Sasa, M.; Calvete, J.J. Studies on the Venom Proteome of Bothrops Asper: Perspectives and Applications. Toxicon 2009, 54, 938–948. [Google Scholar] [CrossRef]
- Larréché, S.; Chippaux, J.P.; Chevillard, L.; Mathé, S.; Résière, D.; Siguret, V.; Mégarbane, B. Bleeding and Thrombosis: Insights into Pathophysiology of Bothrops Venom-Related Hemostasis Disorders. Int. J. Mol. Sci. 2021, 22, 9643. [Google Scholar] [CrossRef]
- Benvenuti, L.A.; França, F.O.S.; Barbaro, K.C.; Nunes, J.R.; Cardoso, J.L.C. Pulmonary Haemorrhage Causing Rapid Death after Bothrops Jararacussu Snakebite: A Case Report. Toxicon 2003, 42, 331–334. [Google Scholar] [CrossRef]
- Malaque, C.M.S.; Duayer, I.F.; Santoro, M.L. Acute Kidney Injury Induced by Thrombotic Microangiopathy in Two Cases of Bothrops Envenomation. Clin. Toxicol. 2019, 57, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Wasko, D.K.; Sasa, M. Habitat Selection of the Terciopelo (Serpentes: Viperidae: Bothrops Asper) in a Lowland Rainforest in Costa Rica. Herpetologica 2010, 66, 148–158. [Google Scholar] [CrossRef]
- Vega-coto, J.; Ramírez-Arce, D.; Baaijen, W.; Artavia-león, A.; ZúñiGa, A. Bothrops Asper. Arboreal Behavior. Mesoam. Herpetology. 2015, 2, 199–201. [Google Scholar]
- Gutiérrez, J.M.; Escalante, T.; Rucavado, A. Experimental Pathophysiology of Systemic Alterations Induced by Bothrops Asper Snake Venom. Toxicon 2009, 54, 976–987. [Google Scholar] [CrossRef] [PubMed]
- Rucavado, A.; Soto, M.; Kamiguti, A.S.; Theakston, R.D.G.; Fox, J.W.; Escalante, T.; Gutiérrez, J.M. Characterization of Aspercetin, a Platelet Aggregating Component from the Venom of the Snake Bothrops Asper Which Induces Thrombocytopenia and Potentiates Metalloproteinase-Induced Hemorrhage. Thromb. Haemost. 2001, 85, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Borkow, G.; Gutiérrez, J.; Ovadia, M. In Vitro Activity of BaH1, the Main Hemorrhagic Toxin of Bothrops Asper Snake Venom on Bovine Endothelial Cells. Toxicon 1995, 33, 1387–1391. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Rey-Suárez, P.; Tsai, W.C.; Angulo, Y.; Sasa, M.; Gutiérrez, J.M.; Calvete, J.J. Snake Venomics of the Pit Vipers Porthidium Nasutum, Porthidium Ophryomegas, and Cerrophidion Godmani from Costa Rica: Toxicological and Taxonomical Insights. J. Proteomics 2012, 75, 1675–1689. [Google Scholar] [CrossRef] [PubMed]
- Schramer, T.D.; de la Torre-Loranca, M.A.; Salazar-Saavedra, M.; Kalki, Y.; Wylie, D.B. Additional Diet Information for Cerrophidion Godmani (Günther, 1863) and a Dietary Synopsis of the Genus Cerrophidion Campbell and Lamar, 1992. Rev. Latinoam. Herpetol. 2018, 1, 35–43. [Google Scholar] [CrossRef]
- Russell, F.E.; Walter, F.G.; Bey, T.A.; Fernandez, M.C. Snakes and Snakebite in Central America. Toxicon 1997, 35, 1469–1522. [Google Scholar] [CrossRef] [PubMed]
- Chaves, G.; Lamar, W.; Porras, L.W.; Solórzano, A.; Townsend, J.H.; Wilson, L.D. Cerrophidion Godmani, Godman’s Montane Pit Viper. IUCN Red List Threat. Species. Available online: https://www.iucnredlist.org/species/197466/217775108 (accessed on 29 February 2024).
- Arni, R.K.; Fontes, M.R.M.; Barberato, C.; Gutiérrez, J.M.; Díaz, C.; Ward, R.J. Crystal Structure of Myotoxin II, a Monomeric Lys49-Phospholipase A2 Homologue Isolated from the Venom of Cerrophidion (Bothrops) Godmani. Arch. Biochem. Biophys. 1999, 366, 177–182. [Google Scholar] [CrossRef]
- Díaz, C.; Gutiérrez, J.; Lomonte, B. Isolation and Characterization of Basic Myotoxic Phospholipases A2 from Bothrops Godmani (Godman’s Pit Viper) Snake Venom. Arch. Biochem. Biophys. 1992, 298, 135–142. [Google Scholar] [CrossRef]
- De Sousa, M.V.; Morhy, L.; Arni, R.K.; Ward, R.J.; Díaz, C.; Gutiérrez, J.M. Amino Acid Sequence of a Myotoxic Lys49-Phospholipase A2 Homologue from the Venom of Cerrophidion (Bothrops) Godmani. Biochim. Biophys. Acta—Protein Struct. Mol. Enzymol. 1998, 1384, 204–208. [Google Scholar] [CrossRef]
- Angulo, Y.; Castro, A.; Lomonte, B.; Rucavado, A.; Fernández, J.; Calvete, J.J.; Gutiérrez, J.M. Isolation and Characterization of Four Medium-Size Disintegrins from the Venoms of Central American Viperid Snakes of the Genera Atropoides, Bothrops, Cerrophidion and Crotalus. Biochimie 2014, 107, 376–384. [Google Scholar] [CrossRef]
- Castro, E.N.; Lomonte, B.; del Carmen Gutiérrez, M.; Alagón, A.; Gutiérrez, J.M. Intraspecies Variation in the Venom of the Rattlesnake Crotalus Simus from Mexico: Different Expression of Crotoxin Results in Highly Variable Toxicity in the Venoms of Three Subspecies. J. Proteomics 2013, 87, 103–121. [Google Scholar] [CrossRef]
- Calvete, J.J.; Sanz, L.; Angulo, Y.; Lomonte, B.; Gutiérrez, J.M. Venoms, Venomics, Antivenomics. FEBS Lett. 2009, 583, 1736–1743. [Google Scholar] [CrossRef]
- Gutierrez, J. Envenenamientos Por Mordeduras de Serpientes En América Latina y El Caribe. Boletín Mariología Y Salud Ambient. 2011, 5I, 1–16. [Google Scholar]
- De Andrade, C.M.; Rey, F.M.; Bianchini, F.J.; Sampaio, S.V.; Torqueti, M.R. Crotoxin, a Neurotoxin from Crotalus Durissus Terrificus Snake Venom, as a Potential Tool against Thrombosis Development. Int. J. Biol. Macromol. 2019, 134, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Carbajal-Márquez, R.A.; Cedeño-Vázquez, J.R.; Martins, M.; Köhler, G. Life History, Activity Pattern, and Morphology of Crotalus Tzabcan Klauber, 1952 (Serpentes: Viperidae). Herpetol. Conserv. Biology. 2020, 15, 228–237. [Google Scholar]
- De Assis, E.B.; Estevão-Costa, M.I.; Do Carmo Valentim, A.; Silva-Neto, A.; Agostini Cotta, G.; Alvarenga Mudado, M.; Richardson, M.; Fortes-Dias, C.L. Purification and Complete Primary Structure of the First PLA2 from Lachesis Stenophrys (the Central American Bushmaster) Snake Venom. Protein J. 2008, 27, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Gómez, A.; Osorno-Valencia, D.; Gómez-Díaz, M.; Bolívar-García, W.; Jiménez-Charris, E. Proteomic and Functional Analyses of Lachesis Acrochorda Snake Venom from the Valle Del Cauca Department of Colombia. Acta Trop. 2023, 241, 106895. [Google Scholar] [CrossRef] [PubMed]
- Franco-Vásquez, A.M.; Lazcano-Pérez, F.; Mejía-Sánchez, M.A.; Corzo, G.; Zamudio, F.; Carbajal-Saucedo, A.; Román-González, S.A.; Gómez-Manzo, S.; Arreguín-Espinosa, R. Structural, Biochemical and Immunochemical Characterization of an Acidic Phospholipase A2 from Lachesis Acrochorda (Viperidae: Crotalinae) Venom. Toxicon 2024, 237, 107528. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.A.; Frost, D.R.; Castoe, T.A. New Generic Name for Jumping Pitvipers (Serpentes: Viperidae). Rev. Latinoam. Herpetol. 2019, 2, 52–53. [Google Scholar] [CrossRef]
- Castoe, T.A.; Chippindale, P.T.; Campbell, J.A.; Ammerman, L.K.; Parkinson, C.L. Molecular Systematics of the Middle American Jumping Pitvipers (Genus Atropoides) and Phylogeography of the Atropoides Nummifer Complex. Herpetologica 2003, 59, 420–431. [Google Scholar] [CrossRef]
- Jones, L.; Youngman, N.J.; Neri-Castro, E.; Guadarrama-Martínez, A.; Lewin, M.R.; Carter, R.; Frank, N.; Fry, B.G. Differential Antivenom and Small-Molecule Inhibition of Novel Coagulotoxic Variations in Atropoides, Cerrophidion, Metlapilcoatlus, and Porthidium American Viperid Snake Venoms. Toxins 2022, 14, 511. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J. Snake Venomics: From the Inventory of Toxins to Biology. Toxicon 2013, 75, 44–62. [Google Scholar] [CrossRef] [PubMed]
- Lamar, W.W.; Sasa, M. A New Species of Hognose Pitviper, Genus Porthidium, from the Southwestern Pacific of Costa Rica (Serpentes: Viperidae). Rev. Biol. Trop. 2003, 51, 797–804. [Google Scholar] [PubMed]
- Cisneros-Heredia, D.F.; Yánez-Muñoz, M. Reptilia, Viperidae, Crotalinae, Porthidium Nasutum: Distribution Extension and Remarks on Its Range and Records. Check List. 2005, 1, 16–17. [Google Scholar] [CrossRef]
- Montealegre-Sánchez, L.; Gimenes, S.N.C.; Lopes, D.S.; Teixeira, S.C.; Solano-Redondo, L.; de Melo Rodrigues, V.; Jiménez-Charris, E. Antitumoral Potential of Lansbermin-I, a Novel Disintegrin from Porthidium Lansbergii Lansbergii Venom on Breast Cancer Cells. Curr. Top. Med. Chem. 2019, 19, 2069–2078. [Google Scholar] [CrossRef]
- Vargas, L.J.; Londoño, M.; Quintana, J.C.; Rua, C.; Segura, C.; Lomonte, B.; Núñez, V. An Acidic Phospholipase A2 with Antibacterial Activity from Porthidium Nasutum Snake Venom. Comp. Biochem. Physiol.—B Biochem. Mol. Biol. 2012, 161, 341–347. [Google Scholar] [CrossRef]
- Vindas, J.; Carrera, Y.; Lomonte, B.; Gutiérrez, J.M.; Calvete, J.J.; Sanz, L.; Fernández, J. A Novel Pentameric Phospholipase A2 Myotoxin (PophPLA2) from the Venom of the Pit Viper Porthidium Ophryomegas. Int. J. Biol. Macromol. 2018, 118, 1–8. [Google Scholar] [CrossRef]
- Næss, I.A.; Christiansen, S.C.; Romundstad, P.; Cannegieter, S.C.; Rosendaal, F.R.; Hammerstrøm, J. Incidence and Mortality of Venous Thrombosis: A Population-Based Study. J. Thromb. Haemost. 2007, 5, 692–699. [Google Scholar] [CrossRef]
- Mackman, N. Triggers, Targets and Treatments for Thrombosis. Nature 2008, 451, 914–918. [Google Scholar] [CrossRef] [PubMed]
- Savage, B.; Almus-Jacobs, F.; Ruggeri, Z.M. Specific Synergy of Multiple Substrate-Receptor Interactions in Platelet Thrombus Formation under Flow. Cell 1998, 94, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Adler, M.; Lazarus, R.A.; Dennis, M.S.; Wagner, G. Solution Structure of Kistrin, a Potent Platelet Aggregation Inhibitor and GP IIb-IIIa Antagonist Author. Science 1991, 253, 445–448. [Google Scholar] [CrossRef] [PubMed]
- Coughlin, S.R. Protease-Activated Receptors in Hemostasis, Thrombosis and Vascular Biology. J. Thromb. Haemost. 2005, 3, 1800–1814. [Google Scholar] [CrossRef]
- Marmur, J.; Singanallore, V.; Fyfe, B.; Guha, A.; Sharma, S.; Ambrose, J.; Fallon, J.; Nemerson, Y.; Taubman, M. Identification of Active Tissue Fctor in Human Coronary Atheroma. Circulation 1996, 94, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Tremoli, E.; Camera, M.; Toschi, V.; Colli, S. Tissue Factor in Atherosclerosis. Atherosclerosis 1999, 144, 273–283. [Google Scholar] [CrossRef]
- Meadows, T.A.; Bhatt, D.L. Clinical Aspects of Platelet Inhibitors and Thrombus Formation. Circ. Res. 2007, 100, 1261–1275. [Google Scholar] [CrossRef]
- Lippi, G.; Montagnana, M.; Danese, E.; Favaloro, E.J.; Franchini, M. Glycoprotein IIb/IIIa Inhibitors: An Update on the Mechanism of Action and Use of Functional Testing Methods to Assess Antiplatelet Efficacy. Biomark. Med. 2011, 5, 63–70. [Google Scholar] [CrossRef]
- Stone, J.; Hangge, P.; Albadawi, H.; Wallace, A.; Shamoun, F.; Knuttien, M.G.; Naidu, S.; Oklu, R. Deep Vein Thrombosis: Pathogenesis, Diagnosis, and Medical Management. Cardiovasc. Diagn. Ther. 2017, 7, S276–S284. [Google Scholar] [CrossRef]
- Lord, S.T. Fibrinogen and Fibrin: Scaffold Proteins in Hemostasis. Curr. Opin. Hematol. 2007, 14, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Mackman, N. New Insights into the Mechanisms of Venous Thrombosis. J. Clin. Invest. 2012, 122, 2331–2336. [Google Scholar] [CrossRef] [PubMed]
- Boldrini-França, J.; Cologna, C.T.; Pucca, M.B.; de Bordon, K.C.F.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cerni, F.A.; Pinheiro-Junior, E.L.; et al. Minor Snake Venom Proteins: Structure, Function and Potential Applications. Biochim. Biophys. Acta—Gen. Subj. 2017, 1861, 824–838. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.K.; Saikia, D.; Thakur, R. Medical and Diagnostic Applications of Snake Venom Proteomes. J. Proteins Proteomics 2011, 2, 31–40. [Google Scholar]
- Fernandes Pedrosa, M.; Félix-Silva, J.; Menezes, Y.A.S. Toxins from Venomous Animals: Gene Cloning, Protein Expression and Biotechnological Applications. In An Integrated View of the Molecular Recognition and Toxinology—From Analytical Procedures to Biomedical Applications; InTech: Atyrau, Republic of Kazakhstan, 2013. [Google Scholar]
- Chen, N.; Xu, S.; Zhang, Y.; Wang, F. Animal Protein Toxins: Origins and Therapeutic Applications. Biophys. Rep. 2018, 4, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J.; Juárez, P.; Sanz, L. Snake Venomics. Strategy and Applications. J. Mass. Spectrom. 2007, 42, 1405–1414. [Google Scholar] [CrossRef]
- Moura-da-Silva, A.; Butera, D.; Tanjoni, I. Importance of Snake Venom Metalloproteinases in Cell Biology: Effects on Platelets, Inflammatory and Endothelial Cells. Curr. Pharm. Des. 2007, 13, 2893–2905. [Google Scholar] [CrossRef]
- Tanjoni, I.; Weinlich, R.; Della-Casa, M.S.; Clissa, P.B.; Saldanha-Gama, R.F.; De Freitas, M.S.; Barja-Fidalgo, C.; Amarante-Mendes, G.P.; Moura-Da-Silva, A.M. Jararhagin, a Snake Venom Metalloproteinase, Induces a Specialized Form of Apoptosis (Anoikis) Selective to Endothelial Cells. Apoptosis 2005, 10, 851–861. [Google Scholar] [CrossRef]
- Maria, A.A.; da Silva, L.G.L.; Correia, C.C.; Ruiz, G.R.G. Antiproliferative Effect of the Jararhagin Toxin on B16F10 Murine Melanoma. BMC Complement. Altern. Med. 2014, 14, 446. [Google Scholar] [CrossRef]
- Matsui, T.; Fujimura, Y.; Titani, K. Snake Venom Proteases Affecting Hemostasis and Thrombosis. Biochim. Biophys. Acta—Protein Struct. Mol. Enzymol. 2000, 1477, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Markland, F.S.; Swenson, S. Snake Venom Metalloproteinases. Toxicon 2013, 62, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Takeya, H.; Iwanaga, S. Snake Venom Metalloproteinases: Structure, Function and Relevance to the Mammalian ADAM/ADAMTS Family Proteins. Biochim. Biophys. Acta—Proteins Proteom. 2012, 1824, 164–176. [Google Scholar] [CrossRef]
- Da Silva Araújo, L.; Conceição, A.S.M.M.; de Souza Cunha, D.M.; de Morais, G.B.; de Moraes Silveira, J.A.; Xavier Júnior, F.A.F.; da Silva Macambira, K.D.; Araújo, S.L.; Pessoa, N.O.; Evangelista, J.S.A.M. Crotalus Durissus Venom: Biological Effects and Relevant Applications. A Review. Rev. Bras. Hig. E Sanid. Anim. 2016, 10, 9–21. [Google Scholar] [CrossRef]
- Fonseca, F.V.; Antunes, E.; Morganti, R.P.; Monteiro, H.S.A.; Martins, A.M.C.; Toyama, D.O.; Marangoni, S.; Toyama, M.H. Characterization of a New Platelet Aggregating Factor from Crotoxin Crotalus Durissus Cascavella Venom. Protein J. 2006, 25, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Serrano, S.M.T.; Maroun, R.C. Snake Venom Serine Proteinases: Sequence Homology vs. Substrate Specificity, a Paradox to Be Solved. Toxicon 2005, 45, 1115–1132. [Google Scholar] [CrossRef]
- Sakurai, Y.; Shima, M.; Matsumoto, T.; Takatsuka, H.; Nishiya, K.; Kasuda, S.; Fujimura, Y.; Yoshioka, A. Anticoagulant Activity of M-LAO, L-Amino Acid Oxidase Purified from Agkistrodon Halys Blomhoffii, through Selective Inhibition of Factor IX. Biochim. Biophys. Acta—Proteins Proteom. 2003, 1649, 51–57. [Google Scholar] [CrossRef]
- Sakurai, Y.; Takatsuka, H.; Yoshioka, A.; Matsui, T.; Suzuki, M.; Titani, K.; Fujimura, Y. Inhibition of Human Platelet Aggregation by L-Amino Acid Oxidase Purified from Naja Naja Kaouthia Venom. Toxicon 2001, 39, 1827–1833. [Google Scholar] [CrossRef] [PubMed]
- Paloschi, M.V.; Pontes, A.S.A.M.S.; Zuliani, J.P. An Update on Potential Molecular Mechanisms Underlying the Actions of Snake Venom L-Amino Acid Oxidases (LAAOs). Curr. Med. Chem. 2018, 25, 2520–2530. [Google Scholar] [CrossRef]
- Nathan, I.; Dvilansky, A.; Yirmiyahu, T.; Aharon, M.; Livne, A. Impairment of Platelet Aggregation by Echis Colorata Venom Mediated by L-Amino Acid Oxidase or H2O2. Thromb. Haemost. 1982, 48, 277–282. [Google Scholar] [CrossRef]
- Momic, T.; Cohen, G.; Reich, R.; Arlinghaus, F.T.; Eble, J.A.; Marcinkiewicz, C.; Lazarovici, P. Vixapatin (VP12), a C-Type Lectin-Protein from Vipera Xantina Palestinae Venom: Characterization as a Novel Anti-Angiogenic Compound. Toxins 2012, 4, 862–877. [Google Scholar] [CrossRef]
- Ogawa, T.; Chijiwa, T.; Oda-Ueda, N.; Ohno, M. Molecular Diversity and Accelerated Evolution of C-Type Lectin-like Proteins from Snake Venom. Toxicon 2005, 45, 1–14. [Google Scholar] [CrossRef]
- Mizuno, H.; Fujimoto, Z.; Koizumi, M.; Kano, H.; Atoda, H.; Morita, T. Crystal Structure of Coagulation Factor IX-Binding Protein from Habu Snake Venom at 2.6 Å: Implication of Central Loop Swapping Based on Deletion in the Linker Region. J. Mol. Biol. 1999, 289, 103–112. [Google Scholar] [CrossRef]
- Zingali, R.B.; Jandrot-Perrus, M.; Guillin, M.C.; Bon, C. Bothrojaracin, a New Thrombin Inhibitor Isolated from Bothrops Jararaca Venom: Characterization and Mechanism of Thrombin Inhibition. Biochemistry 1993, 32, 10794–10802. [Google Scholar] [CrossRef]
- Coronado, M.A.; Gabdulkhakov, A.; Georgieva, D.; Sankaran, B.; Murakami, M.T.; Raghuvir, A.K.; Betzelb, C. A Multi Structure of the Polypeptide Crotamine from the Brazilian Rattlesnake Crotalus Durissus Terrificus Research Papers. Acta Crystallogr. Sect. D 2013, 1958–1964. [Google Scholar] [CrossRef]
- Radis-Baptista, G.; Kerkis, I. Crotamine, a Small Basic Polypeptide Myotoxin from Rattlesnake Venom with Cell-Penetrating Properties. Curr. Pharm. Des. 2011, 17, 4351–4361. [Google Scholar] [CrossRef]
- Andrews, R.K.; Gardiner, E.E.; Shen, Y.; Berndt, M.C. Structure-Activity Relationships of Snake Toxins Targeting Platelet Receptors, Glycoprotein Ib-IX-V and Glycoprotein VI. Curr. Med. Chem. Cardiovasc. Hematol. Agents 2003, 1, 143–149. [Google Scholar] [CrossRef]
- Camargo, A.C.M.; Ianzer, D.; Guerreiro, J.R.; Serrano, S.M.T. Bradykinin-Potentiating Peptides: Beyond Captopril. Toxicon 2012, 59, 516–523. [Google Scholar] [CrossRef]
- Ferreira, S.H. A Bradykinin-Potentiating Factor (Bpf) Present in the Venom of Bothrops Jararaca. Br. J. Pharmacol. Chemother. 1965, 24, 163–169. [Google Scholar] [CrossRef]
- Oliveira, A.L.; Viegas, M.F.; da Silva, S.L.; Soares, A.M.; Ramos, M.J.; Fernandes, P.A. The Chemistry of Snake Venom and Its Medicinal Potential. Nat. Rev. Chem. 2022, 6, 451–469. [Google Scholar] [CrossRef] [PubMed]
- Siebler, M.; Hennerici, M.G.; Schneider, D.; Von Reutern, G.M.; Seitz, R.J.; Röther, J.; Witte, O.W.; Hamann, G.; Junghans, U.; Villringer, A.; et al. Safety of Tirofiban in Acute Ischemic Stroke: The SaTIS Trial. Stroke 2011, 42, 2388–2392. [Google Scholar] [CrossRef] [PubMed]
- Gan, Z.R.; Gould, R.J.; Jacobs, J.W.; Friedman, P.A.; Polokoff, M.A. Echistatin. A Potent Platelet Aggregation Inhibitor from the Venom of the Viper, Echis Carinatus. J. Biol. Chem. 1988, 263, 19827–19832. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.R.; Scarborough, R.M. Clinical Pharmacology of Eptifibatide. Am. J. Cardiol. 1997, 80, 11B–20B. [Google Scholar] [CrossRef] [PubMed]
- Scarborough, R.M.; Rose, J.W.; Hsu, M.A.; Phillips, D.R.; Fried, V.A.; Campbell, A.M.; Nannizzi, L.; Charo, I.F. Barbourin: A GPIIb-IIIa-Specific Integrin Antagonist from the Venom of Sistrurus, M. Barbouri. J. Biol. Chem. 1991, 266, 9359–9362. [Google Scholar] [CrossRef] [PubMed]
- Karapetian, H. Reptilase Time (RT). Methods Mol. Biol. 2013, 992, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wang, Y.; Zhao, X.; Li, X.; Wang, Q.; Zhong, W.; Mequanint, K.; Zhan, R.; Xing, M.; Luo, G. Snake Extract-Laden Hemostatic Bioadhesive Gel Cross-Linked by Visible Light. Sci. Adv. 2021, 7, 9635. [Google Scholar] [CrossRef] [PubMed]
- Castro, H.C.; Zingali, R.B.; Albuquerque, M.G.; Pujol-Luz, M.; Rodrigues, C.R. Snake Venom Thrombin-like Enzymes: From Reptilase to Now. Cell. Mol. Life Sci. 2004, 61, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Stocker, K.; Fischera, H.; Meier, J.; Brogli, M.; Svendsen, L. Characterization of the Protein C Activator Protac® from the Venom of the Southern Copperhead (Agkistrodon Contortrix) Snake. Toxicon 1987, 25, 239–252. [Google Scholar] [CrossRef]
- Gempeler-Messina, P.M.; Volz, K.; Bühler, B.; Müller, C.; Gempeler-Messina, P.M. Anticoagulant Proteins and Diagnostic Agents Protein C Activators from Snake Venoms and Their Diagnostic Use. Pathophysiol. Haemost. Thromb. 2002, 31, 266–272. [Google Scholar] [CrossRef]
- Brass, L.F.; Zhu, L.; Stalker, T.J. Minding the Gaps to Promote Thrombus Growth and Stability. J. Clin. Investig. 2005, 115, 3385–3392. [Google Scholar] [CrossRef]
- Rivas-Mercado, E.A.; Garza-Ocañas, L. Disintegrins Obtained from Snake Venom and Their Pharmacological Potential. Med. Univ. 2017, 19, 32–37. [Google Scholar] [CrossRef]
- Laing, G.D.; Moura-da-Silva, A.M. Jararhagin and Its Multiple Effects on Hemostasis. Toxicon 2005, 45, 987–996. [Google Scholar] [CrossRef]
- Muniz, J.R.C.; Ambrosio, A.L.B.; Selistre-de-Araujo, H.S.; Cominetti, M.R.; Moura-da-Silva, A.M.; Oliva, G.; Garratt, R.C.; Souza, D.H.F. The Three-Dimensional Structure of Bothropasin, the Main Hemorrhagic Factor from Bothrops Jararaca Venom: Insights for a New Classification of Snake Venom Metalloprotease Subgroups. Toxicon 2008, 52, 807–816. [Google Scholar] [CrossRef]
- Kamiguti, A.S.; Hay, C.R.M.; Theakston, R.D.G. Insights into the Mechanism of Haemorrhage Caused by Snake Venom Metalloproteinases. Toxicon 1996, 34, 627–642. [Google Scholar] [CrossRef]
- Kamiguti, A.S.; Theakston, R.D.G.; Watson, S.P.; Bon, C.; Laing, G.D.; Zuzel, M. Distinct Contributions of Glycoprotein VI and A2β1 Integrin to the Induction of Platelet Protein Tyrosine Phosphorylation and Aggregation. Arch. Biochem. Biophys. 2000, 374, 356–362. [Google Scholar] [CrossRef]
- Wu, W.B.; Peng, H.C.; Huang, T.F. Crotalin, a VWF and GP Ib Cleaving Metalloproteinase from Venom of Crotalus Atrox. Thromb. Haemost. 2001, 86, 1501–1511. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.-C.; Lin, H.-K.; Peng, H.-C.; Huang, T.-F. Antithrombotic Effect of Crotalin, a Platelet Membrane Glycoprotein Ib Antagonist From Venom of Crotalus Atrox. Blood 1998, 91, 1582–1589. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.; Romero, M.; Díaz, C.; Borkow, G.; Ovadia, M. Isolation and Characterization of a Metalloproteinase with Weak Hemorrhagic Activity from the Venom of the Snake Bothrops Asper (Terciopelo). Toxicon 1995, 33, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, A.; Rucavado, A.; Mora, N.; Gutiérrez, J.M. Purification and Characterization of BaH4, a Hemorrhagic Metalloproteinase from the Venom of the Snake Bothrops Asper. Toxicon 2000, 38, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Loría, G.D.; Rucavado, A.; Kamiguti, A.S.; Theakston, R.D.G.; Fox, J.W.; Alape, A.; Gutiérrez, J.M. Characterization of “basparin A,” a Prothrombin-Activating Metalloproteinase, from the Venom of the Snake Bothrops Asper That Inhibits Platelet Aggregation and Induces Defibrination and Thrombosis. Arch. Biochem. Biophys. 2003, 418, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, A.A.; Estrada, J.C.; David, V.; Wermelinger, L.S.; Almeida, F.C.L.; Zingali, R.B. Structure-Function Relationship of the Disintegrin Family: Sequence Signature and Integrin Interaction. Front. Mol. Biosci. 2021, 8, 783301. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, A.A.; Estrada, J.C.; Caruso, I.P.; Kurtenbach, E.; Zingali, R.B.; Almeida, F.C.L. Toward the Mechanism of Jarastatin (RJast) Inhibition of the Integrin AVβ3. Int. J. Biol. Macromol. 2024, 255, 128078. [Google Scholar] [CrossRef]
- Vasconcelos, A.A.; Succar, B.B.; di Piero, L.B.; Kurtenbach, E.; Zingali, R.B.; Almeida, F.C.L. 15N, 13C, and 1H Resonance Assignments of Jarastatin: A Disintegrin of Bothrops Jararaca. Biomol. NMR Assign. 2022, 16, 37–40. [Google Scholar] [CrossRef]
- Xiong, J.; Stehle, T.; Diefenbach, B.; Zhang, R.; Scott, D.L.; Joachimiak, A.; Goodman, S.L.; Amin, M. Crystal. Struttura Cristallina. Science 2010, 294, 339–345. [Google Scholar] [CrossRef]
- Succar, B.B.; Saldanha-Gama, R.F.G.; Valle, A.S.; Wermelinger, L.S.; Barja-Fidalgo, C.; Kurtenbach, E.; Zingali, R.B. The Recombinant Disintegrin, Jarastatin, Inhibits Platelet Adhesion and Endothelial Cell Migration. Toxicon 2022, 217, 87–95. [Google Scholar] [CrossRef] [PubMed]
- David, V.; Wermelinger, L.S.; Frattani, F.S.; Lima, A.G.F.; Santos, Y.F.S.; de Mourão, P.A.S.; Almeida, F.C.L.; Eleonora Kurtenbach, C.R.B.Z. rJararacin, a Recombinant Disintegrin from Bothrops Jararaca Venom: Exploring Its Effects on Hemostasis and Thrombosis. Arch. Biochem. Biophys. 2023, 738, 109557. [Google Scholar] [CrossRef]
- Wermelinger, L.S.; Geraldo, R.B.; Frattani, F.S.; Rodrigues, C.R.; Juliano, M.A.; Castro, H.C.; Zingali, R.B. Integrin Inhibitors from Snake Venom: Exploring the Relationship between the Structure and Activity of RGD-Peptides. Arch. Biochem. Biophys. 2009, 482, 25–32. [Google Scholar] [CrossRef] [PubMed]
- López-Dávila, A.J.; Weber, N.; Kraft, T.; Matinmehr, F.; Arias-Hidalgo, M.; Fernández, J.; Lomonte, B.; Gutiérrez, J.M. Cytotoxicity of Snake Venom Lys49 PLA2-like Myotoxin on Rat Cardiomyocytes Ex Vivo Does Not Involve a Direct Action on the Contractile Apparatus. Sci. Rep. 2021, 11, 19452. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Fernández, J.; Sanz, L.; Angulo, Y.; Sasa, M.; Gutiérrez, J.M.; Calvete, J.J. Venomous Snakes of Costa Rica: Biological and Medical Implications of Their Venom Proteomic Profiles Analyzed through the Strategy of Snake Venomics. J. Proteom. 2014, 105, 323–339. [Google Scholar] [CrossRef] [PubMed]
- Salvador, G.H.M.; Dos Santos, J.I.; Borges, R.J.; Fontes, M.R.M. Structural Evidence for a Fatty Acid-Independent Myotoxic Mechanism for a Phospholipase A2-like Toxin. Biochim. Biophys. Acta Proteins Proteom. 2018, 1866, 473–481. [Google Scholar] [CrossRef]
- Tan, P.; Tan, P.T.J.; Khan, A.M.; Brusic, V. Bioinformatics for Venom and Toxin Sciences. Br. Bioinform. 2003, 4, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M.; Evans, H.J. Structure-Function Relationships of Phospholipases. The Anticoagulant Region of Phospholipases A2. J. Biol. Chem. 1987, 262, 14402–14407. [Google Scholar] [CrossRef]
- Kini, R.M. Excitement Ahead: Structure, Function and Mechanism of Snake Venom Phospholipase A2 Enzymes. Toxicon 2003, 42, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Almeida, C.D.S.; Andrade-Oliveira, V.; Câmara, N.O.S.; Jacysyn, J.F.; Faquim-Mauro, E.L. Crotoxin from Crotalus Durissus Terrificus Is Able to Down-Modulate the Acute Intestinal Inflammation in Mice. PLoS ONE 2015, 10, e0121427. [Google Scholar] [CrossRef]
- Sarray, S.; Luis, J.; El, M.; Marrakchi, N. Snake Venom Peptides: Promising Molecules with Anti-Tumor Effects. In Bioactive Food Peptides in Health and Disease; InTech: Atyrau, Republic of Kazakhstan, 2013; Volume I, p. 13. [Google Scholar] [CrossRef]
- Gimenez, B.T.; Cezarette, G.N.; De Sousa Bomfim, A.; Monteiro, W.M.; De Sousa Russo, E.M.; Frantz, F.G.; Sampaio, S.V.; Sartim, M.A. Role of Crotoxin in Coagulation: Novel Insights into Anticoagulant Mechanisms and Impairment of Inflammation-Induced Coagulation. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Prezoto, B.C.; Tanaka-Azevedo, A.M.; Marcelino, J.R.; Tashima, A.K.; Nishiduka, E.S.; Kapronezai, J.; Mota, J.O.; Rocha, M.M.T.; Serino-Silva, C.; Oguiura, N. A Functional and Thromboelastometric-Based Micromethod for Assessing Crotoxin Anticoagulant Activity and Antiserum Relative Potency against Crotalus Durissus Terrificus Venom. Toxicon 2018, 148, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Sousa, I.D.L.; Barbosa, A.R.; Salvador, G.H.M.; Frihling, B.E.F.; Santa-Rita, P.H.; Soares, A.M.; Pessôa, H.L.F.; Marchi-Salvador, D.P. Secondary Hemostasis Studies of Crude Venom and Isolated Proteins from the Snake Crotalus Durissus Terrificus. Int. J. Biol. Macromol. 2019, 131, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Faure, G.; Gowda, V.T.; Maroun, R.C. Characterization of a Human Coagulation Factor Xa-Binding Site on Viperidae Snake Venom Phospholipases A2 by Affinity Binding Studies and Molecular Bioinformatics. BMC Struct. Biol. 2007, 7, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M. Serine Proteases Affecting Blood Coagulation and Fibrinolysis from Snake Venoms. Pathophysiol. Haemost. Thromb. 2006, 34, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Latinović, Z.; Leonardi, A.; Koh, C.Y.; Kini, R.M.; Bakija, A.T.; Pungerčar, J.; Križaj, I. The Procoagulant Snake Venom Serine Protease Potentially Having a Dual, Blood Coagulation Factor v and X-Activating Activity. Toxins 2020, 12, 358. [Google Scholar] [CrossRef]
- Bakker, H.M.; Tans, G.; Yukelson, L.Y.; Janssen-Claessen, T.W.; Bertina, R.M.; Hemker, H.C.; Rosing, J. Protein C Activation by an Activator Purified from the Venom of Agkistrodon Halys Halys. Blood Coagul. Fibrinolysis 1993, 44, 605–614. [Google Scholar] [CrossRef]
- Au, L.C.; Lin, S.B.; Chou, J.S.; Teh, G.W.; Chang, K.J.; Shih, C.M. Molecular Cloning and Sequence Analysis of the CDNA for Ancrod, a Thrombin-like Enzyme from the Venom of Calloselasma Rhodostoma. Biochem. J. 1993, 294, 387–390. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, Y.Q.; Liu, F.; Li, M.H.; Zhao, J.G.; Tan, H.Q.; Wang, J.B.; Cheng, Y.S.; Zhang, P.L. Batroxobin for Prevention of Restenosis in Diabetic Patients after Infrapopliteal Arterial Angioplasty: A Small Randomized Pilot Trial. Ann. Vasc. Surg. 2010, 24, 876–884. [Google Scholar] [CrossRef]
- Xu, G.; Liu, X.; Zhu, W.; Yin, Q.; Zhang, R.; Fan, X. Feasibility of Treating Hyperfibrinogenemia with Intermittently Administered Batroxobin in Patients with Ischemic Stroke/Transient Ischemic Attack for Secondary Prevention. Blood Coagul. Fibrinolysis 2007, 18, 193–197. [Google Scholar] [CrossRef]
- Braud, S.; Le Bonniec, B.F.; Bon, C.; Wisner, A. The Stratagem Utilized by the Plasminogen Activator from the Snake Trimeresurus Stejnegeri to Escape Serpins. Biochemistry 2002, 41, 8478–8484. [Google Scholar] [CrossRef]
- Zhang, Y.; Wisner, A.; Xiong, Y.; Bon, C. A Novel Plasminogen Activator from Snake Venom: Purification, Characterization, and Molecular Cloning. J. Biol. Chem. 1995, 270, 10246–10255. [Google Scholar] [CrossRef]
- Sanchez, E.F.; Felicori, L.F.; Chavez-Olortegui, C.; Magalhaes, H.B.P.; Hermogenes, A.L.; Diniz, M.V.; de Junqueira-de-Azevedo, I.L.M.; Magalhaes, A.; Richardson, M. Biochemical Characterization and Molecular Cloning of a Plasminogen Activator Proteinase (LV-PA) from Bushmaster Snake Venom. Biochim. Biophys. Acta—Gen. Subj. 2006, 1760, 1762–1771. [Google Scholar] [CrossRef] [PubMed]
- Hermogenes, A.L.; Richardson, M.; Magalhaes, A.; Yarleque, A.; Rodriguez, E.; Sanchez, E.F. Interaction of a Plasminogen Activator Proteinase, LV-PA with Human A2-Macroglobulin. Toxicon 2006, 47, 490–494. [Google Scholar] [CrossRef]
- Liu, S.; Sun, M.Z.; Greenaway, F.T. A Novel Plasminogen Activator from Agkistrodon Blomhoffii Ussurensis Venom (ABUSV-PA): Purification and Characterization. Biochem. Biophys. Res. Commun. 2006, 348, 1279–1287. [Google Scholar] [CrossRef] [PubMed]
- Morita, T. Structures and Functions of Snake Venom CLPs (C-Type Lectin-like Proteins) with Anticoagulant-, Procoagulant-, and Platelet-Modulating Activities. Toxicon 2005, 45, 1099–1114. [Google Scholar] [CrossRef]
- Eble, J.A. Structurally Robust and Functionally Highly Versatile—C-Type Lectin (-Related) Proteins in Snake Venoms. Toxins 2019, 11, 136. [Google Scholar] [CrossRef] [PubMed]
- Marcinkiewicz, C.; Lobb, R.R.; Marcinkiewicz, M.M.; Daniel, J.L.; Smith, J.B.; Dangelmaier, C.; Weinreb, P.H.; Beacham, D.A.; Niewiarowski, S. Isolation and Characterization of EMS16, a C-Lectin Type Protein from Echis Multisquamatus Venom, a Potent and Selective Inhibitor of the A2β1 Integrin. Biochemistry 2000, 39, 9859–9867. [Google Scholar] [CrossRef]
- Atoda, H.; Kaneko, H.; Mizuno, H.; Morita, T. Calcium-Binding Analysis and Molecular Modeling Reveal Echis Coagulation Factor IX/Factor X-Binding Protein Has the Ca-Binding Properties and Ca Ion-Independent Folding of Other C-Type Lectin-like Proteins. FEBS Lett. 2002, 531, 229–234. [Google Scholar] [CrossRef]
- Atoda, H.; Morita, T. Arrangement of the Disulfide Bridges in a Blood Coagulation Factor IX/Factor X-Binding Protein from the Venom of Trimeresurus Flavoviridis. J. Biochem. 1993, 113, 159–163. [Google Scholar] [CrossRef]
- Assafim, M.; Frattani, F.S.; Ferreira, M.S.; Silva, D.M.; Monteiro, R.Q.; Zingali, R.B. Exploiting the Antithrombotic Effect of the (pro)Thrombin Inhibitor Bothrojaracin. Toxicon 2016, 119, 46–51. [Google Scholar] [CrossRef]
- Monteiro, R.Q.; Rapôso, J.G.; Wisner, A.; Guimarães, J.A.; Bon, C.; Zingali, R.B. Allosteric Changes of Thrombin Catalytic Site Induced by Interaction of Bothrojaracin with Anion-Binding Exosites I and II. Biochem. Biophys. Res. Commun. 1999, 262, 819–822. [Google Scholar] [CrossRef] [PubMed]
- Arocas, V.; Zingali, R.B.; Guillin, M.-C.; Bon, C.; Jandrot-Perrus, M. Bothrojaracin: A Potent Two-Site-Directed Thrombin Inhibitor. Biochemistry 1996, 35, 9083–9089. [Google Scholar] [CrossRef] [PubMed]
- Zingali, R.B.; Bianconi, M.L.; Monteiro, R.Q. Interaction of Bothrojaracin with Prothrombin. Haemostasis 2001, 31, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.Q.; Zingali, R.B. Inhibition of Prothrombin Activation by Bothrojaracin, a C-Type Lectin from Bothrops Jararaca Venom. Arch. Biochem. Biophys. 2000, 382, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Sekiya, F.; Atoda, H.; Morita, T. Isolation and Characterization of an Anticoagulant Protein Homologous to Botrocetin from the Venom of Bothrops Jararaca. Biochemistry 1993, 32, 6892–6897. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, R.; Yoshiara, E.; Yamada, M.; Shima, K.; Atoda, H.; Morita, T. CDNA Cloning of IX/X-BP, a Heterogeneous Two-Chain Anticoagulant Protein from Snake Venom. Biochem. Biophys. Res. Commun. 1996, 220, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Kumashiro, M.; Yamazaki, Y.; Atoda, H.; Morita, T. Anticoagulant Mechanism of Factor IX/Factor X-Binding Protein Isolated from the Venom of Trimeresurus Flavoviridis. J. Biochem. 2009, 145, 123–128. [Google Scholar] [CrossRef]
- Sekiya, F.; Yamashita, T.; Morita, T. Role of Calcium(II) Ions in the Recognition of Coagulation Factors IX and X by IX/X-Bp, an Anticoagulant from Snake Venom. Biochemistry 1995, 34, 10043–10047. [Google Scholar] [CrossRef]
- Cheng, X.; Qian, Y.; Liu, Q.; Li, B.X.Y.; Zhang, M.; Liu, J. Purification, Characterization, and CDNA Cloning of a New Fibrinogenlytic Venom Protein, Agkisacutacin, from Agkistrodon Acutus Venom. Biochem. Biophys. Res. Commun. 1999, 265, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Eble, J.A. Titration Elisa as a Method to Determine the Dissociation Constant of Receptor Ligand Interaction. J. Vis. Exp. 2018, 2018, 57334. [Google Scholar] [CrossRef]
- Eble, J.A.; McDougall, M.; Orriss, G.L.; Niland, S.; Johanningmeier, B.; Pohlentz, G.; Meier, M.; Karrasch, S.; Estevão-Costa, M.I.; Martins Lima, A.; et al. Dramatic and Concerted Conformational Changes Enable Rhodocetin to Block A2β1 Integrin Selectively. PLoS Biol. 2017, 15, 1492. [Google Scholar] [CrossRef] [PubMed]
- Eble, J.A.; Niland, S.; Bracht, T.; Mormann, M.; Peter-Katalinic, J.; Pohlentz, G.; Stetefeld, J. The A2β1 Integrin-specific Antagonist Rhodocetin Is a Cruciform, Heterotetrameric Molecule. FASEB J. 2009, 23, 2917–2927. [Google Scholar] [CrossRef]
- Watson, A.A.; Eble, J.A.; O’Callaghan, C.A. Crystal Structure of Rhodocytin, a Ligand for the Platelet-activating Receptor CLEC-2. Protein Sci. 2008, 17, 1611–1616. [Google Scholar] [CrossRef]
- May, F.; Hagedorn, I.; Pleines, I.; Bender, M.; Vögtle, T.; Eble, J.; Elvers, M.; Nieswandt, B. CLEC-2 Is an Essential Platelet-Activating Receptor in Hemostasis and Thrombosis. Blood 2009, 114, 3464–3472. [Google Scholar] [CrossRef]
- Fujimura, Y.; Ikeda, Y.; Miura, S.; Yoshida, E.; Shima, H.; Nishida, S.; Suzuki, M.; Titani, K.; Taniuchi, Y.; Kawasaki, T. Isolation and Characterization of Jararaca GPIb-BP, a Snake Venom Antagonist Specific to Platelet Glycoprotein Ib. Thromb. Haemost. 1995, 74, 743–750. [Google Scholar] [PubMed]
- Clemetson, K.J.; Lu, Q.; Clemetson, J.M. Snake C-Type Lectin-like Proteins and Platelet Receptors. Pathophysiol. Haemost. Thromb. 2006, 34, 150–155. [Google Scholar] [CrossRef]
- Munawar, A.; Ali, S.A.; Akrem, A.; Betzel, C. Snake Venom Peptides: Tools of Biodiscovery. Toxins 2018, 10, 474. [Google Scholar] [CrossRef]
- Kerkis, I.; Hayashi, M.A.F.; Prieto da Silva, A.R.B.; Pereira, A.; De Sá Júnior, P.L.; Zaharenko, A.J.; Rádis-Baptista, G.; Kerkis, A.; Yamane, T. State of the Art in the Studies on Crotamine, a Cell Penetrating Peptide from South American Rattlesnake. Biomed. Res. Int. 2014, 2014, 675985. [Google Scholar] [CrossRef] [PubMed]
- Mas, C.D.; Rossato, L.; Oliveira, E.B.; Pedro, I.; Junior, S.; Meis, J.F.; Colombo, A.L.; Hayashi, M.A.F. Effects of the Natural Peptide Crotamine from a South American Rattlesnake on Candida Auris, an Emergent Multidrug Antifungal Resistant Human Pathogen. Biomolecules 2019, 9, 10. [Google Scholar]
- Batista da Cunha, D.; Pupo Silvestrini, A.V.; Gomes da Silva, A.C.; Maria de Paula Estevam, D.; Pollettini, F.L.; de Oliveira Navarro, J.; Alves, A.A.; Remédio Zeni Beretta, A.L.; Annichino Bizzacchi, J.M.; Pereira, L.C.; et al. Mechanistic Insights into Functional Characteristics of Native Crotamine. Toxicon 2018, 146, 1–12. [Google Scholar] [CrossRef] [PubMed]




| Protein Family | Biotechnological Potential for Thrombosis | Reference |
|---|---|---|
| Metalloproteases and disintegrins | Inhibition of clot formation in relation to the consumption of fibrinogen; cleavage of platelet receptors; inhibition of platelet aggregation | [7,31,108,109,110,111,112,113] |
| Phospholipases A2 (PLA2s) | Strong anti-coagulant proteins that display inhibition of Factors Xa and X at lower concentrations by an extrinsic tenase complex and activation of prothrombin to thrombin. | [114,115] |
| Weak anti-coagulant proteins that exhibit inhibition of Factors Xa and X. | [90,114,115] | |
| Serine proteases | Cleaves fibrinogen plasminogen, Factor Va/VIIIa, and protein C in plasma | [111,116] |
| L-amino-acid oxidases | Isolation of proteins with anti-coagulant activity on factors of the coagulation cascade | [117,118,119,120] |
| Snake C-type lectin-like proteins (snaclecs) | Inhibition of clotting factors (thrombin, Factor X, and Factor IX); inhibition of platelet receptors | [58,64,121,122,123,124] |
| Bioactive peptides | Compounds with high affinity and specificity of receptors associated with different biological activities | [125,126] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang Estrada, J.E.; Guerrero, T.N.; Reyes-Enríquez, D.F.; Nardy, E.S.; Guimarães Ferreira, R.; Ruiz Calderón, C.J.; Wellmann, I.A.; Monteiro Espíndola, K.M.; do Prado, A.F.; Soares, A.M.; et al. Potential Biotechnological Applications of Venoms from the Viperidae Family in Central America for Thrombosis. Toxins 2024, 16, 142. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins16030142
Chang Estrada JE, Guerrero TN, Reyes-Enríquez DF, Nardy ES, Guimarães Ferreira R, Ruiz Calderón CJ, Wellmann IA, Monteiro Espíndola KM, do Prado AF, Soares AM, et al. Potential Biotechnological Applications of Venoms from the Viperidae Family in Central America for Thrombosis. Toxins. 2024; 16(3):142. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins16030142
Chicago/Turabian StyleChang Estrada, Jorge Eduardo, Taissa Nunes Guerrero, Daniel Fernando Reyes-Enríquez, Erica Santos Nardy, Roseane Guimarães Ferreira, Cristian José Ruiz Calderón, Irmgardt A. Wellmann, Kaio Murilo Monteiro Espíndola, Alejandro Ferraz do Prado, Andreimar Martins Soares, and et al. 2024. "Potential Biotechnological Applications of Venoms from the Viperidae Family in Central America for Thrombosis" Toxins 16, no. 3: 142. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins16030142