
Expression of K1 Toxin Derivatives in Saccharomyces cerevisiae Mimics Treatment with Exogenous Toxin and Provides a Useful Tool for Elucidating K1 Mechanisms of Action and Immunity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
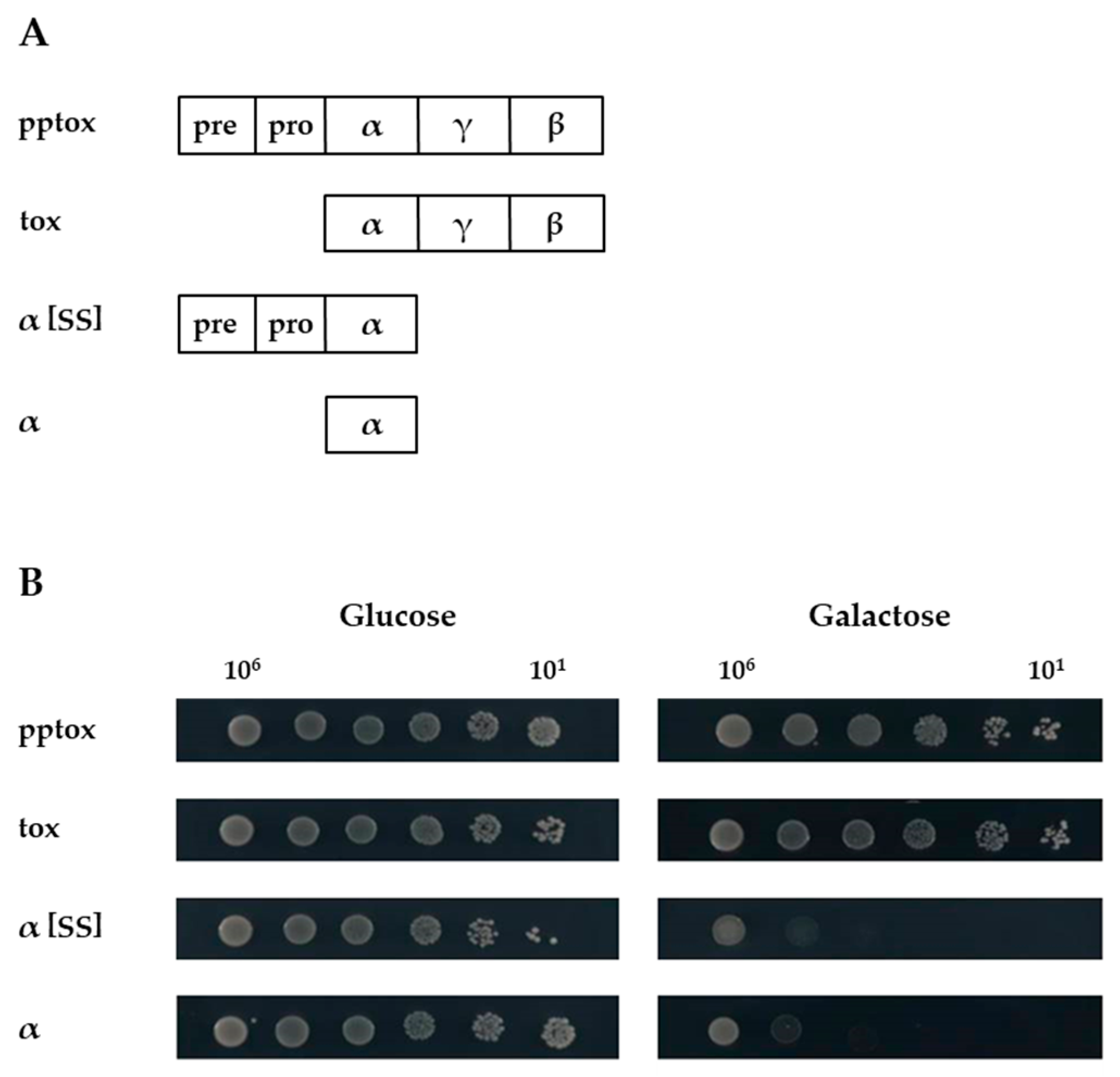
2.1. Expression of K1α Derivatives Mimics Natural Toxin Action
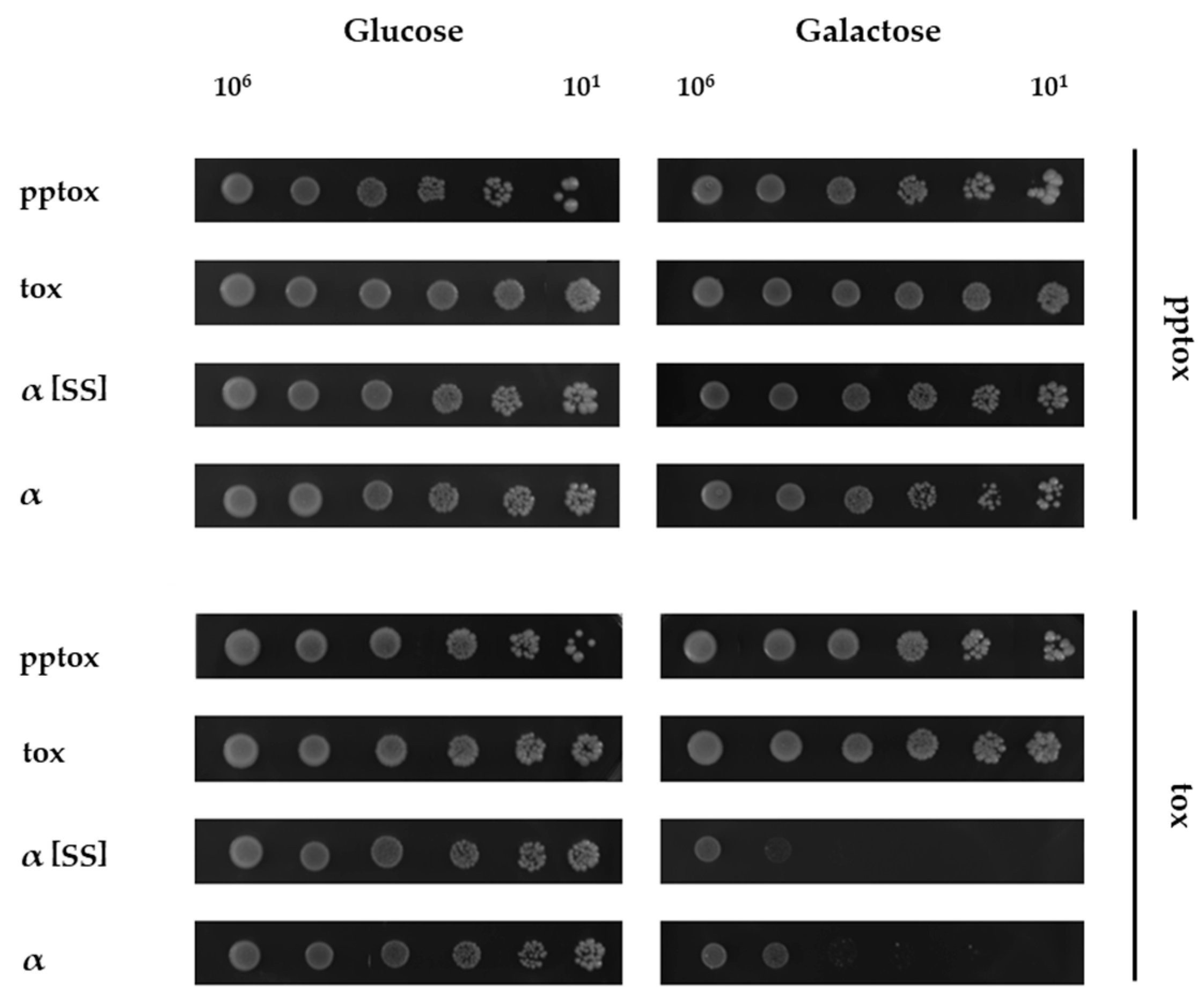
2.2. Co-Expression of K1 Toxin Precursor Abolishes the Suicidal Phenotype of K1α Derivatives
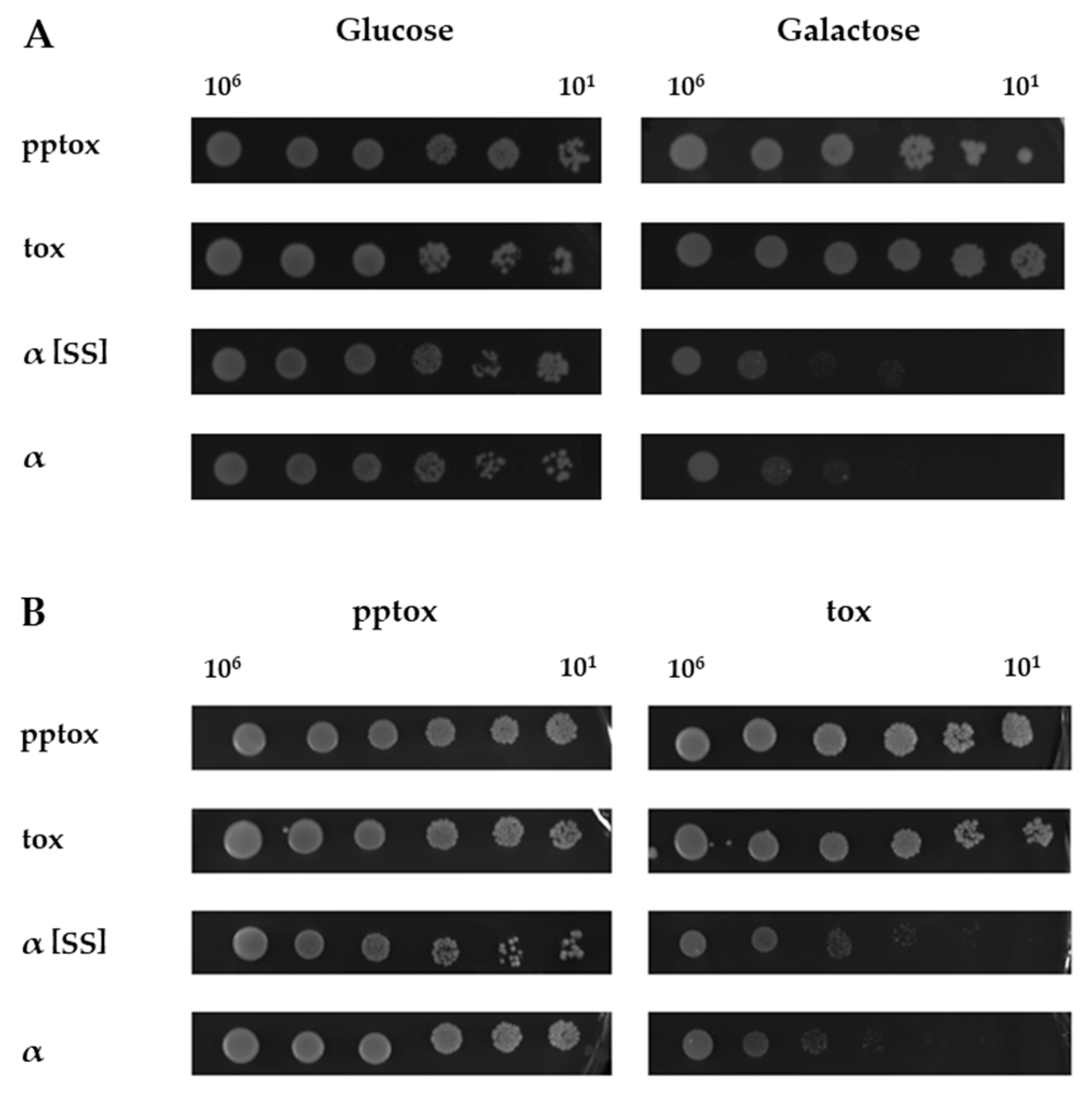
2.3. Kre1p Is Involved in Neither K1 Lethality nor Immunity
3. Discussion
4. Materials and Methods
4.1. Strains, Plasmids, and DNA Manipulation
4.2. Yeast Transformation and Selection
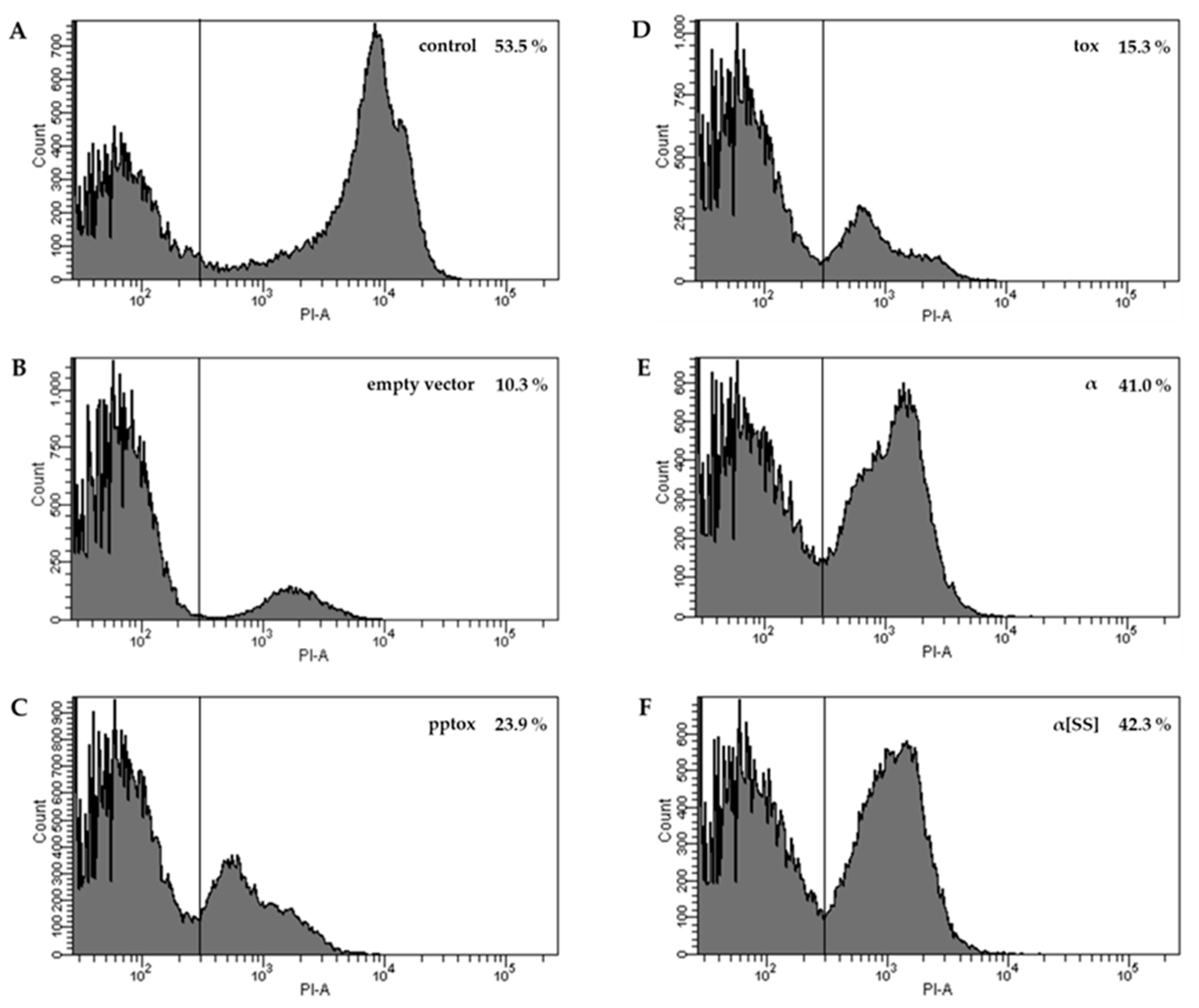
4.3. Characterization of Suicidal Phenotype
4.4. Flow Cytometry
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bevan, E.A.; Makower, M. The physiological basis of the killer-character in yeast. In Genetics Today. Proceedings of the XI International Congress of Genetics; Geerts, S.J., Ed.; Pergamon Press: Oxford, UK, 1963; Volume 1, pp. 202–203. [Google Scholar]
- Schmitt, M.J.; Breinig, F. The viral killer system in yeast: From molecular biology to application. FEMS Microbiol. Rev. 2002, 26, 257–276. [Google Scholar] [CrossRef] [PubMed]
- Magliani, W.; Conti, S.; Gerloni, M.; Bertolotti, D.; Polonelli, L. Yeast killer systems. Clin. Microbiol. Rev. 1997, 10, 369–400. [Google Scholar] [PubMed]
- Schmitt, M.J.; Breinig, F. Yeast viral killer toxins: Lethality and self-protection. Nat. Rev. Microbiol. 2006, 4, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Tipper, D.J.; Schmitt, M.J. Yeast dsRNA viruses: Replication and killer phenotypes. Mol. Microbiol. 1991, 5, 2331–2338. [Google Scholar] [CrossRef] [PubMed]
- Bostian, K.A.; Elliott, Q.; Bussey, H.; Burn, V.; Smith, A.; Tipper, D.J. Sequence of the preprotoxin dsRNA gene of type 1 killer yeast: Multiple processing events produce a two-component toxin. Cell 1984, 36, 741–751. [Google Scholar] [CrossRef]
- Schmitt, M.J.; Tipper, D.J. Sequence of the m28 dsRNA: Preprotoxin is processed to an alpha/beta heterodimeric protein toxin. Virology 1995, 213, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Eisfeld, K.; Riffer, F.; Mentges, J.; Schmitt, M.J. Endocytotic uptake and retrograde transport of a virally encoded killer toxin in yeast. Mol. Microbiol. 2000, 37, 926–940. [Google Scholar] [CrossRef] [PubMed]
- Heiligenstein, S.; Eisfeld, K.; Sendzik, T.; Jimenez-Becker, N.; Breinig, F.; Schmitt, M.J. Retrotranslocation of a viral a/b toxin from the yeast endoplasmic reticulum is independent of ubiquitination and ERAD. EMBO J. 2006, 25, 4717–4727. [Google Scholar] [CrossRef] [PubMed]
- Martinac, B.; Zhu, H.; Kubalski, A.; Zhou, X.L.; Culbertson, M.; Bussey, H.; Kung, C. Yeast K1 killer toxin forms ion channels in sensitive yeast spheroplasts and in artificial liposomes. Proc. Natl. Acad. Sci. USA 1990, 87, 6228–6232. [Google Scholar] [CrossRef] [PubMed]
- Breinig, F.; Tipper, D.J.; Schmitt, M.J. Kre1p, the plasma membrane receptor for the yeast K1 viral toxin. Cell 2002, 108, 395–405. [Google Scholar] [CrossRef]
- Pagé, N.; Gerard-Vincent, M.; Menard, P.; Beaulieu, M.; Azuma, M.; Dijkgraaf, G.J.; Li, H.; Marcoux, J.; Nguyen, T.; Dowse, T.; et al. A Saccharomyces cerevisiae genome-wide mutant screen for altered sensitivity to K1 killer toxin. Genetics 2003, 163, 875–894. [Google Scholar] [PubMed]
- Zhu, H.; Bussey, H. Mutational analysis of the functional domains of yeast K1 killer toxin. Mol. Cell. Biol. 1991, 11, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Valis, K.; Masek, T.; Novotna, D.; Pospisek, M.; Janderova, B. Immunity to killer toxin K1 is connected with the golgi-to-vacuole protein degradation pathway. Folia Microbiol. 2006, 51, 196–202. [Google Scholar] [CrossRef]
- Boone, C.; Bussey, H.; Greene, D.; Thomas, D.Y.; Vernet, T. Yeast killer toxin: Site-directed mutations implicate the precursor protein as the immunity component. Cell 1986, 46, 105–113. [Google Scholar] [CrossRef]
- Breinig, F.; Schleinkofer, K.; Schmitt, M.J. Yeast Kre1p is GPI-anchored and involved in both cell wall assembly and architecture. Microbiology 2004, 150, 3209–3218. [Google Scholar] [CrossRef] [PubMed]
- Roemer, T.; Bussey, H. Yeast Kre1p is a cell surface O-glycoprotein. Mol. Gen. Genet. 1995, 249, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Breinig, F.; Sendzik, T.; Eisfeld, K.; Schmitt, M.J. Dissecting toxin immunity in virus-infected killer yeast uncovers an intrinsic strategy of self-protection. Proc. Natl. Acad. Sci. USA 2006, 103, 3810–3815. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Sesti, F.; Ilan, N.; Shih, T.M.; Sturley, S.L.; Goldstein, S.A. A molecular target for viral killer toxin: Tok1 potassium channels. Cell 1999, 99, 283–291. [Google Scholar] [CrossRef]
- Sesti, F.; Shih, T.M.; Nikolaeva, N.; Goldstein, S.A. Immunity to K1 killer toxin: Internal Tok1 blockade. Cell 2001, 105, 637–644. [Google Scholar] [CrossRef]
- Bertl, A.; Ramos, J.; Ludwig, J.; Lichtenberg-Frate, H.; Reid, J.; Bihler, H.; Calero, F.; Martinez, P.; Ljungdahl, P.O. Characterization of potassium transport in wild-type and isogenic yeast strains carrying all combinations of trk1, trk2 and tok1 null mutations. Mol. Microbiol. 2003, 47, 767–780. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.S.; Kane, J.; Zhang, X.Y.; Zhang, M.; Tipper, D.J. Role of the gamma component of preprotoxin in expression of the yeast K1 killer phenotype. Yeast 1993, 9, 251–266. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Fukuda, Y.; Murata, K.; Kimura, A. Transformation of intact yeast cells treated with alkali cations. J. Bacteriol. 1983, 153, 163–168. [Google Scholar] [PubMed]




© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gier, S.; Schmitt, M.J.; Breinig, F. Expression of K1 Toxin Derivatives in Saccharomyces cerevisiae Mimics Treatment with Exogenous Toxin and Provides a Useful Tool for Elucidating K1 Mechanisms of Action and Immunity. Toxins 2017, 9, 345. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins9110345
Gier S, Schmitt MJ, Breinig F. Expression of K1 Toxin Derivatives in Saccharomyces cerevisiae Mimics Treatment with Exogenous Toxin and Provides a Useful Tool for Elucidating K1 Mechanisms of Action and Immunity. Toxins. 2017; 9(11):345. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins9110345
Chicago/Turabian StyleGier, Stefanie, Manfred J. Schmitt, and Frank Breinig. 2017. "Expression of K1 Toxin Derivatives in Saccharomyces cerevisiae Mimics Treatment with Exogenous Toxin and Provides a Useful Tool for Elucidating K1 Mechanisms of Action and Immunity" Toxins 9, no. 11: 345. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins9110345