Human Bocavirus Infection of Permanent Cells Differentiated to Air-Liquid Interface Cultures Activates Transcription of Pathways Involved in Tumorigenesis

 and
and
Abstract
:1. Introduction
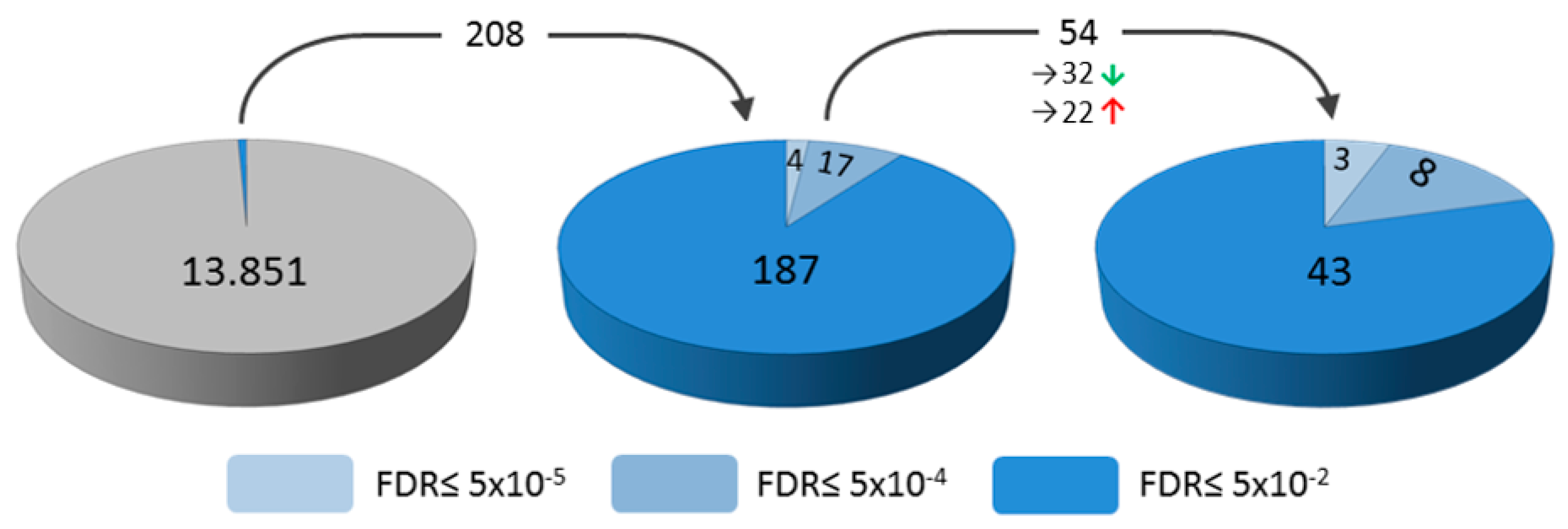
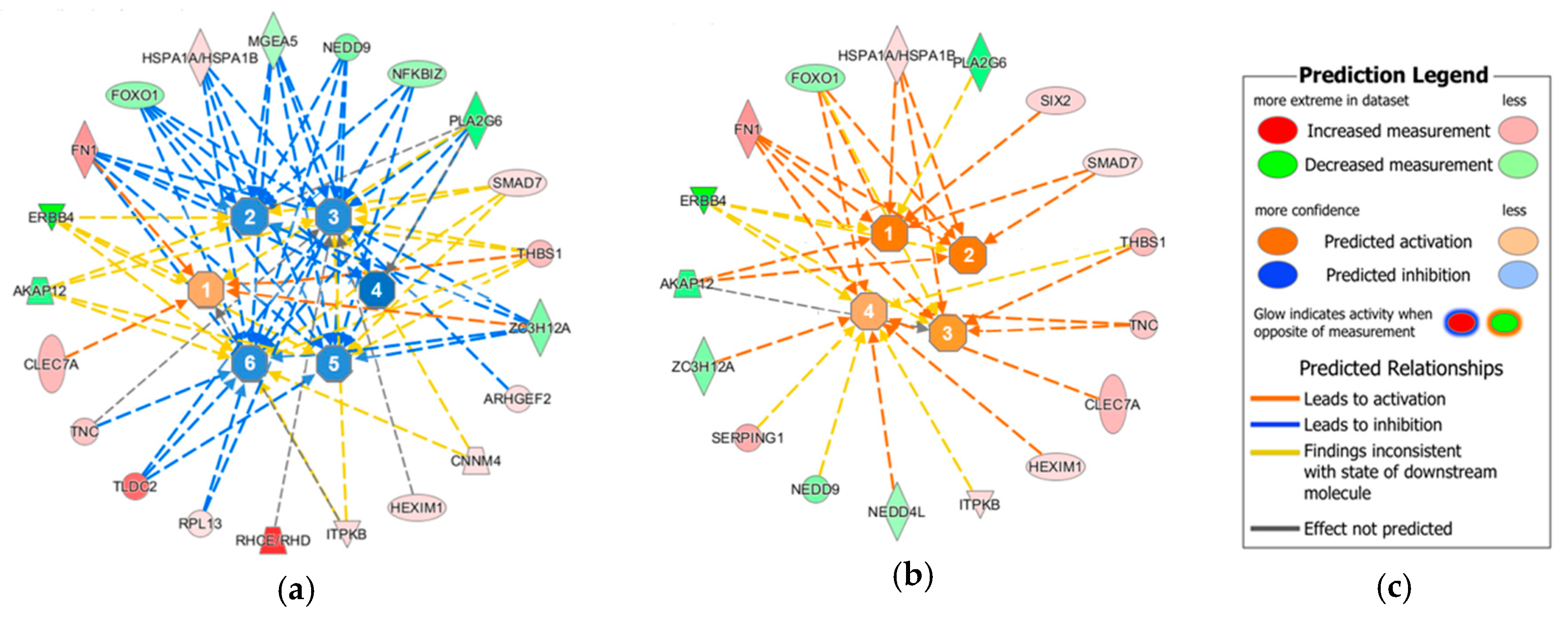
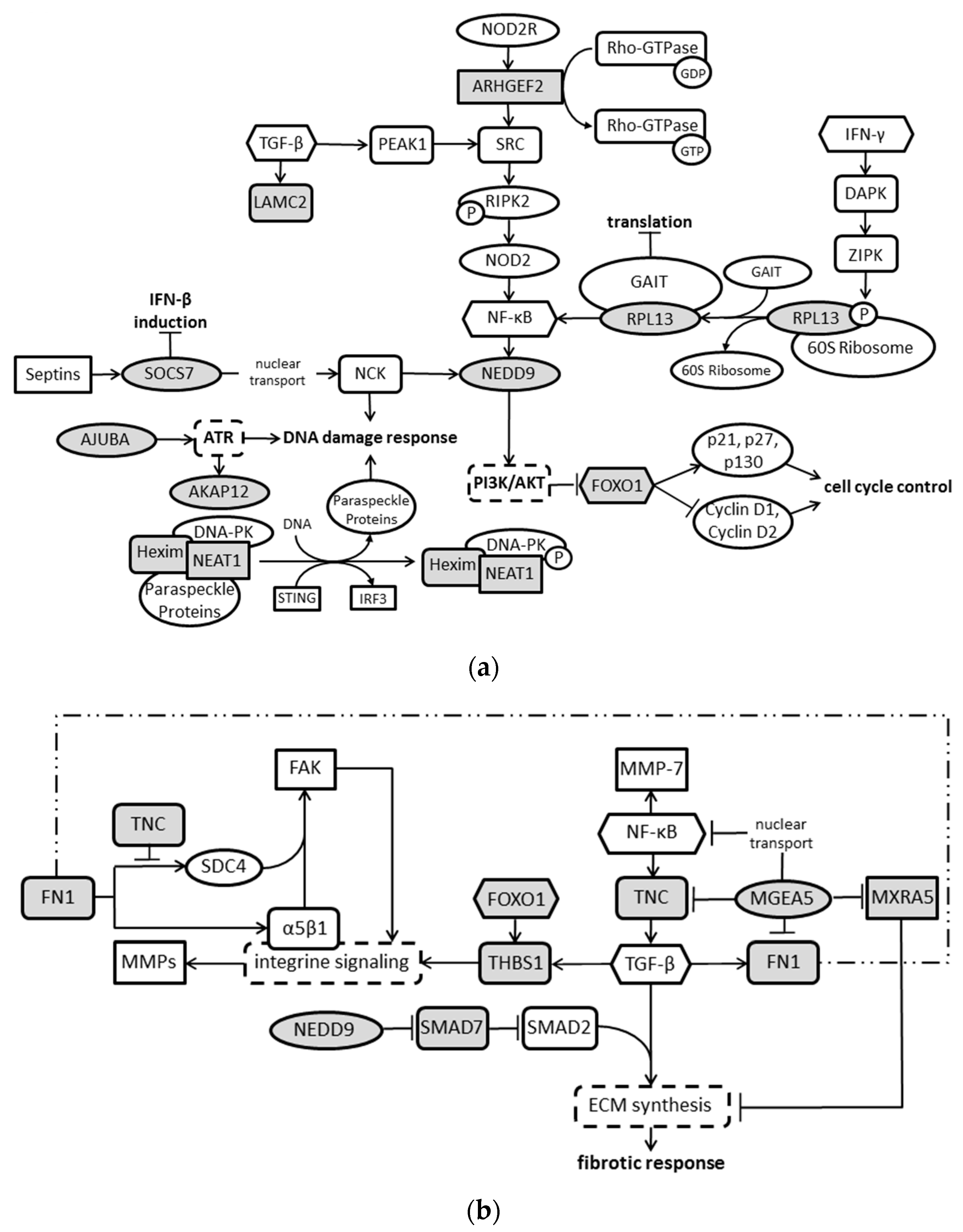
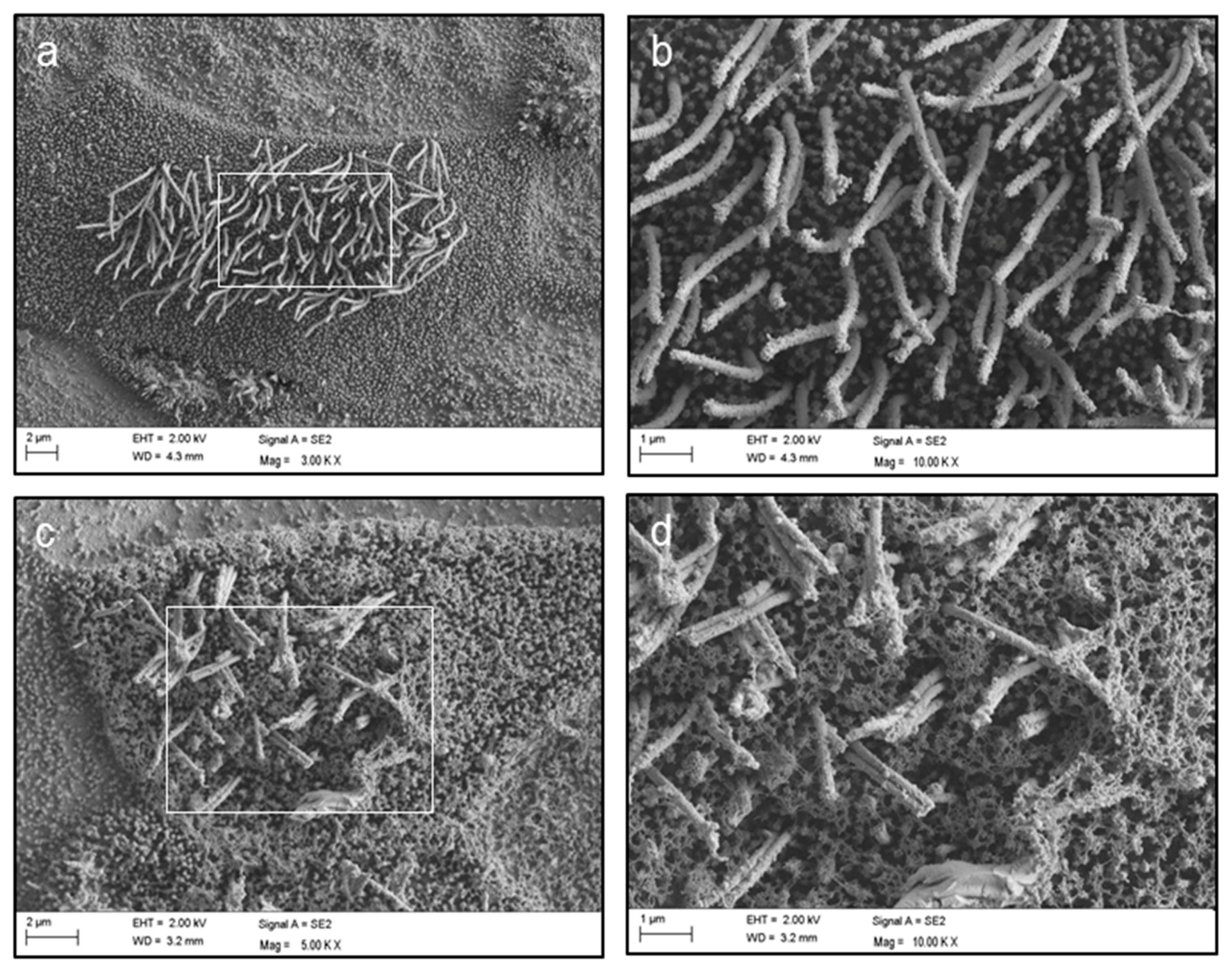
2. Results
3. Discussion
4. Materials and Methods
4.1. Cells and Infections
4.2. RNA Extractions and Whole Transcriptome Analyses
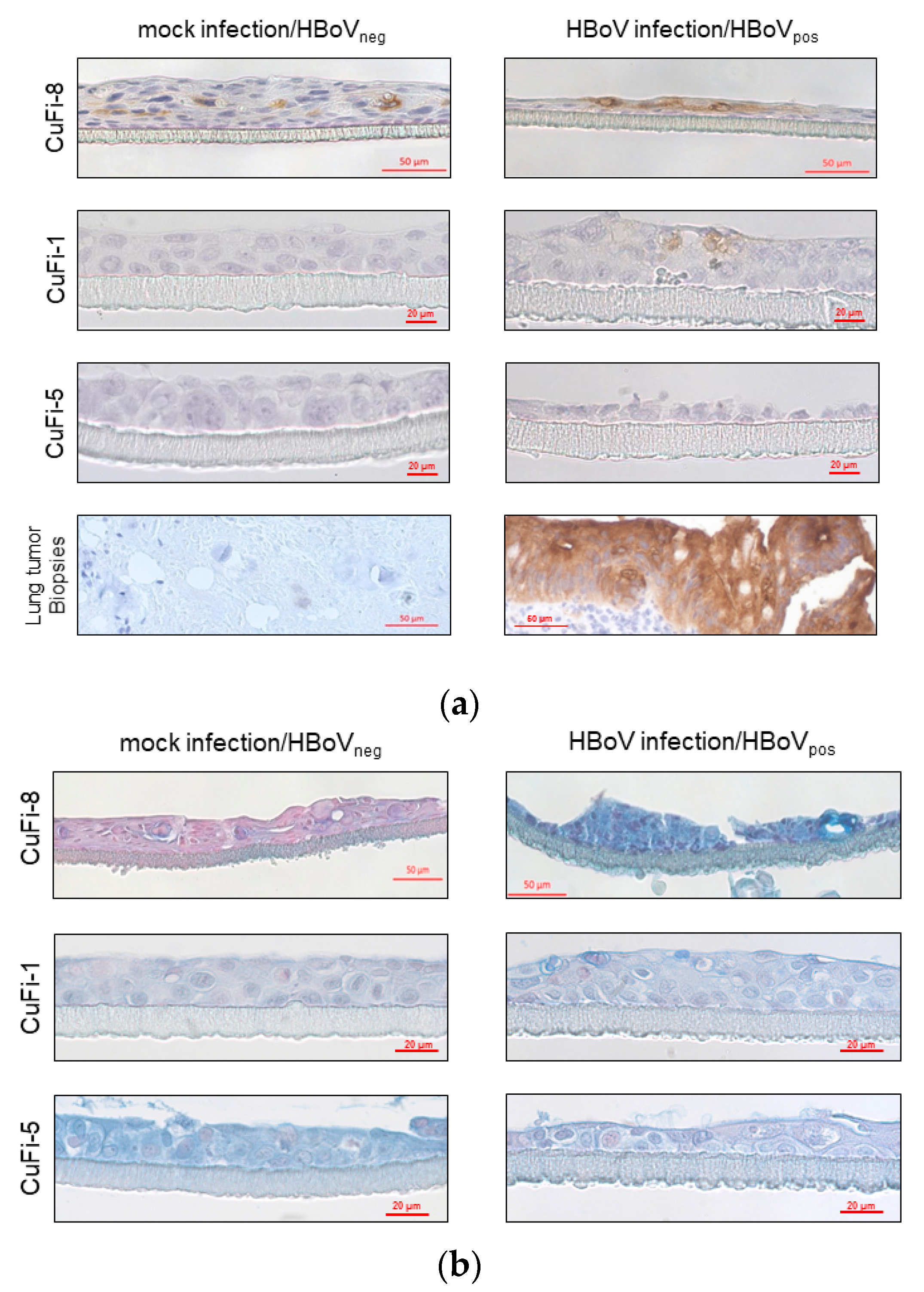
4.3. Immunohistochemical Analyses of CuFi air–Liquid-Interface Cultures
4.4. Scanning Electron Microscopy
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Schildgen, V.; Malecki, M.; Tillmann, R.L.; Brockmann, M.; Schildgen, O. The human bocavirus is associated with some lung and colorectal cancers and persists in solid tumors. PLoS ONE 2013, 8, e68020. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Moneim, A.S.; El-Fol, H.A.; Kamel, M.M.; Soliman, A.S.; Mahdi, E.A.; El-Gammal, A.S.; Mahran, T.Z. Screening of human bocavirus in surgically excised cancer specimens. Arch. Virol. 2016, 161, 2095–2102. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chen, X.Z.; Waterboer, T.; Castro, F.A.; Brenner, H. Viral infections and colorectal cancer: A systematic review of epidemiological studies. Int. J. Cancer 2015, 137, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Dong, Y.; Jiang, J.; Yang, Y.; Liu, K. High prevelance of human parvovirus infection in patients with malignant tumors. Oncol. Lett. 2012, 3, 635–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guido, M.; Tumolo, M.R.; Verri, T.; Romano, A.; Serio, F.; De Giorgi, M.; De Donno, A.; Bagordo, F.; Zizza, A. Human bocavirus: Current knowledge and future challenges. World J. Gastroenterol. 2016, 22, 8684–8697. [Google Scholar] [CrossRef] [PubMed]
- Broccolo, F.; Falcone, V.; Esposito, S.; Toniolo, A. Human bocaviruses: Possible etiologic role in respiratory infection. J. Clin. Virol. 2015, 72, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Lusebrink, J.; Schildgen, V.; Tillmann, R.L.; Wittleben, F.; Bohmer, A.; Muller, A.; Schildgen, O. Detection of head-to-tail DNA sequences of human bocavirus in clinical samples. PLoS ONE 2011, 6, e19457. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.C.; Pyles, R.B.; Miller, A.L.; Nokso-Koivisto, J.; Loeffelholz, M.J.; Chonmaitree, T. Determining persistence of bocavirus DNA in the respiratory tract of children by pyrosequencing. Pediatr. Infect. Dis. J. 2016, 35, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Garcia, M.L.; Calvo, C.; Pozo, F.; Perez-Brena, P.; Quevedo, S.; Bracamonte, T.; Casas, I. Human bocavirus detection in nasopharyngeal aspirates of children without clinical symptoms of respiratory infection. Pediatr. Infect. Dis. J. 2008, 27, 358–360. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J.; Schildgen, V.; Tillmann, R.; Hardt, A.-L.; Lüsebrink, J.; Windisch, W.; Schildgen, O. Low copy number detection of hbov DNA in bal of asymptomatic adult patients. Future Virol. 2014, 9, 715–720. [Google Scholar] [CrossRef]
- Chonmaitree, T.; Alvarez-Fernandez, P.; Jennings, K.; Trujillo, R.; Marom, T.; Loeffelholz, M.J.; Miller, A.L.; McCormick, D.P.; Patel, J.A.; Pyles, R.B. Symptomatic and asymptomatic respiratory viral infections in the first year of life: Association with acute otitis media development. Clin. Infect. Dis. 2015, 60, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Windisch, W.; Pieper, M.; Ziemele, I.; Rockstroh, J.; Brockmann, M.; Schildgen, O.; Schildgen, V. Latent infection of human bocavirus accompanied by flare of chronic cough, fatigue and episodes of viral replication in an immunocompetent adult patient, cologne, germany. JMM Case Rep. 2016, 3, e005052. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, A.; Hornig, M.; Asokan, A.; Williams, B.; Henriquez, J.A.; Lipkin, W.I. Bocavirus episome in infected human tissue contains non-identical termini. PLoS ONE 2011, 6, e21362. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Pesavento, P.A.; Leutenegger, C.M.; Estrada, M.; Coffey, L.L.; Naccache, S.N.; Samayoa, E.; Chiu, C.; Qiu, J.; Wang, C.; et al. A novel bocavirus in canine liver. Virol. J. 2013, 10, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Zhao, L.; Sun, Y.; Qian, Y.; Liu, L.; Jia, L.; Zhang, Y.; Dong, H. Detection of a bocavirus circular genome in fecal specimens from children with acute diarrhea in beijing, china. PLoS ONE 2012, 7, e48980. [Google Scholar] [CrossRef] [PubMed]
- Schildgen, V.; Khalfaoui, S.; Schildgen, O. Human bocavirus: From common cold to cancer? Speculations on the importance of an episomal genomic form of human bocavirus. Rev. Med. Microbiol. 2014, 25, 113–118. [Google Scholar] [CrossRef]
- Khalfaoui, S.; Eichhorn, V.; Karagiannidis, C.; Bayh, I.; Brockmann, M.; Pieper, M.; Windisch, W.; Schildgen, O.; Schildgen, V. Lung infection by human bocavirus induces the release of profibrotic mediator cytokines in vivo and in vitro. PLoS ONE 2016, 11, e0147010. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Deng, X.; Yan, Z.; Cheng, F.; Luo, Y.; Shen, W.; Lei-Butters, D.C.; Chen, A.Y.; Li, Y.; Tang, L.; et al. Establishment of a reverse genetics system for studying human bocavirus in human airway epithelia. PLoS Pathog. 2012, 8, e1002899. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Yan, Z.; Cheng, F.; Engelhardt, J.F.; Qiu, J. Replication of an autonomous human parvovirus in non-dividing human airway epithelium is facilitated through the DNA damage and repair pathways. PLoS Pathog. 2016, 12, e1005399. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Xu, P.; Zou, W.; Shen, W.; Peng, J.; Liu, K.; Engelhardt, J.F.; Yan, Z.; Qiu, J. DNA damage signaling is required for replication of human bocavirus 1 DNA in dividing hek293 cells. J. Virol. 2017, 91, e01831-16. [Google Scholar] [CrossRef] [PubMed]
- Schildgen, V.; Mai, S.; Khalfaoui, S.; Lusebrink, J.; Pieper, M.; Tillmann, R.L.; Brockmann, M.; Schildgen, O. Pneumocystis jirovecii can be productively cultured in differentiated cufi-8 airway cells. mBio 2014, 5, e01186-14. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in ingenuity pathway analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Errington, T.M.; Macara, I.G. Depletion of the adaptor protein nck increases uv-induced p53 phosphorylation and promotes apoptosis. PLoS ONE 2013, 8, e76204. [Google Scholar] [CrossRef] [PubMed]
- Kremer, B.E.; Adang, L.A.; Macara, I.G. Septins regulate actin organization and cell-cycle arrest through nuclear accumulation of nck mediated by socs7. Cell 2007, 130, 837–850. [Google Scholar] [CrossRef] [PubMed]
- Kalan, S.; Matveyenko, A.; Loayza, D. Lim protein ajuba participates in the repression of the atr-mediated DNA damage response. Front. Genet. 2013, 4, 95. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Zou, W.; Xiong, M.; Wang, Z.; Engelhardt, J.F.; Ye, S.Q.; Yan, Z.; Qiu, J. Human parvovirus infection of human airway epithelia induces pyroptotic cell death by inhibiting apoptosis. J. Virol. 2017, 91, e01533-17. [Google Scholar] [CrossRef] [PubMed]
- Jarrett, S.G.; Wolf Horrell, E.M.; D’Orazio, J.A. Akap12 mediates pka-induced phosphorylation of atr to enhance nucleotide excision repair. Nucleic Acids Res. 2016, 44, 10711–10726. [Google Scholar] [CrossRef] [PubMed]
- Garber, M.E.; Troyanskaya, O.G.; Schluens, K.; Petersen, S.; Thaesler, Z.; Pacyna-Gengelbach, M.; van de Rijn, M.; Rosen, G.D.; Perou, C.M.; Whyte, R.I.; et al. Diversity of gene expression in adenocarcinoma of the lung. Proc. Natl. Acad. Sci. USA 2001, 98, 13784–13789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tessema, M.; Willink, R.; Do, K.; Yu, Y.Y.; Yu, W.; Machida, E.O.; Brock, M.; Van Neste, L.; Stidley, C.A.; Baylin, S.B.; et al. Promoter methylation of genes in and around the candidate lung cancer susceptibility locus 6q23-25. Cancer Res. 2008, 68, 1707–1714. [Google Scholar] [CrossRef] [PubMed]
- Morchikh, M.; Cribier, A.; Raffel, R.; Amraoui, S.; Cau, J.; Severac, D.; Dubois, E.; Schwartz, O.; Bennasser, Y.; Benkirane, M. Hexim1 and neat1 long non-coding rna form a multi-subunit complex that regulates DNA-mediated innate immune response. Mol. Cell 2017, 67, 387–399.e5. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, C.; Baker, D.K.; Pierce, A.J.; Pittman, D.L. The splicing-factor related protein sfpq/psf interacts with rad51d and is necessary for homology-directed repair and sister chromatid cohesion. Nucleic Acids Res. 2011, 39, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Salton, M.; Lerenthal, Y.; Wang, S.Y.; Chen, D.J.; Shiloh, Y. Involvement of matrin 3 and sfpq/nono in the DNA damage response. Cell Cycle 2010, 9, 1568–1576. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.L.; Carmichael, G.G. Altered nuclear retention of mrnas containing inverted repeats in human embryonic stem cells: Functional role of a nuclear noncoding rna. Mol. Cell 2009, 35, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Reinertsen, T.; Halgunset, J.; Viset, T.; Flatberg, A.; Haugsmoen, L.L.; Skogseth, H. Gene expressional changes in prostate fibroblasts from cancerous tissue. Apmis 2012, 120, 558–571. [Google Scholar] [CrossRef] [PubMed]
- Midwood, K.S.; Orend, G. The role of tenascin-c in tissue injury and tumorigenesis. J. Cell Commun. Signal. 2009, 3, 287–310. [Google Scholar] [CrossRef] [PubMed]
- Midwood, K.; Sacre, S.; Piccinini, A.M.; Inglis, J.; Trebaul, A.; Chan, E.; Drexler, S.; Sofat, N.; Kashiwagi, M.; Orend, G.; et al. Tenascin-c is an endogenous activator of toll-like receptor 4 that is essential for maintaining inflammation in arthritic joint disease. Nat. Med. 2009, 15, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Wang, W.; Morales-Nebreda, L.; Feng, G.; Wu, M.; Zhou, X.; Lafyatis, R.; Lee, J.; Hinchcliff, M.; Feghali-Bostwick, C.; et al. Tenascin-c drives persistence of organ fibrosis. Nat. Commun. 2016, 7, 11703. [Google Scholar] [CrossRef] [PubMed]
- Tavazoie, S.F.; Alarcon, C.; Oskarsson, T.; Padua, D.; Wang, Q.; Bos, P.D.; Gerald, W.L.; Massague, J. Endogenous human micrornas that suppress breast cancer metastasis. Nature 2008, 451, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Ramos, J.M.; Ruiz, A.; Colen, R.; Lopez, I.D.; Grossman, L.; Matta, J.L. DNA repair and breast carcinoma susceptibility in women. Cancer 2004, 100, 1352–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pugacheva, E.N.; Jablonski, S.A.; Hartman, T.R.; Henske, E.P.; Golemis, E.A. Hef1-dependent aurora a activation induces disassembly of the primary cilium. Cell 2007, 129, 1351–1363. [Google Scholar] [CrossRef] [PubMed]
- Pugacheva, E.N.; Golemis, E.A. The focal adhesion scaffolding protein hef1 regulates activation of the aurora-a and nek2 kinases at the centrosome. Nat. Cell Biol. 2005, 7, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Pugacheva, E.N.; Golemis, E.A. Hef1-aurora a interactions: Points of dialog between the cell cycle and cell attachment signaling networks. Cell Cycle 2006, 5, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Stefanovic, L.; Longo, L.; Zhang, Y.; Stefanovic, B. Characterization of binding of larp6 to the 5’ stem-loop of collagen mrnas: Implications for synthesis of type i collagen. RNA Biol. 2014, 11, 1386–1401. [Google Scholar] [CrossRef] [PubMed]
- Fahim, A.; Crooks, M.G.; Wilmot, R.; Campbell, A.P.; Morice, A.H.; Hart, S.P. Serum carcinoembryonic antigen correlates with severity of idiopathic pulmonary fibrosis. Respirology 2012, 17, 1247–1252. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, L. Neonatal Viral Infections of Pups: Canine Herpesvirus and Minute Virus of Canines (Canine Parvovirus-1); International Veterinary Information Service, Ithaca: New York, NY, USA, 1999. [Google Scholar]
- Storz, J.; Young, S.; Carroll, E.J.; Bates, R.C.; Bowen, R.A.; Keney, D.A. Parvovirus infection of the bovine fetus: Distribution of infection, antibody response, and age-related susceptibility. Am. J. Vet. Res. 1978, 39, 1099–1102. [Google Scholar] [PubMed]
- Windisch, W.; Schildgen, V.; Malecki, M.; Lenz, J.; Brockmann, M.; Karagiannidis, C.; Schildgen, O. Detection of hbov DNA in idiopathic lung fibrosis, cologne, germany. J. Clin. Virol. 2013, 58, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarty, S.; Tobon, A.; Varani, J.; Brattain, M.G. Induction of carcinoembryonic antigen secretion and modulation of protein secretion/expression and fibronectin/laminin expression in human colon carcinoma cells by transforming growth factor-beta. Cancer Res. 1988, 48, 4059–4064. [Google Scholar] [PubMed]
- Jain, P.; Mondal, S.K.; Sinha, S.K.; Mukhopadhyay, M.; Chakraborty, I. Diagnostic and prognostic significance of different mucin expression, preoperative cea, and ca-125 in colorectal carcinoma: A clinicopathological study. J. Nat. Sci. Biol. Med. 2014, 5, 404–408. [Google Scholar] [PubMed]
- Pai, P.; Rachagani, S.; Dhawan, P.; Batra, S.K. Mucins and wnt/beta-catenin signaling in gastrointestinal cancers: An unholy nexus. Carcinogenesis 2016, 37, 223–232. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Upregulated | Downregulated | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Target RNA | Characterization | Target RNA | Characterization | Target RNA | Characterization | Target RNA | Characterization | Target RNA | Characterization |
| HEXIM1 (I) | transcription regulator | TAGLN (XII) | calcium binding protein | SRSF5 (1) | splicing factor | PHLDB2 (12) | microtubule-anchoring factor | ADGRF4 (23) | G protein-coupled receptor |
| FN1 (II) | ECM glycoprotein | IFITM1 (XIII) | transmembrane protein | NEAT1 (2) | lnc RNA | PLA2G4F (13) | Phospholipase A | LINC0113 8 (24) | lncRNA |
| RHCE/RHD (III) | membrane protein | SOCS7 (XIV) | SSI protein nucleocytoplasmic shuttling protein | ERBB4 (3) | receptor tyrosine kinase | NEDD4L (14) | ubiquitin ligase | SNORD80 (25) | C/D box snoRNA |
| AJUBA (IV) | transcription regulator | ITPKB (XV) | kinase | AKAP12 (4) | scaffold protein | ZNF587B (15) | transcriptional inhibitor | LINC00365 (26) | lncRNA |
| THBS1 (V) | ECM glycoprotein | CNNM4 (XVI) | metal cation transport mediator | SNHG3 (5) | lncRNA | LINC01451 (16) | lncRNA | ANO9 (27) | membrane channel |
| HSPA1A/HSPA1B (VI) | chaperon | LAMC2 (XVII) | ECM glycoprotein | S100A3 (6) | Calcium binding protein | TNNI2 (17) | inhibitory subunit of the troponin complex | NFKBIZ (28) | transcription regulator |
| ARHGEF2 (VII) | guanine nucleotide exchange factor | CLEC7A (XVIII) | membrane receptor | NEDD9 (7) | scaffold protein | PLA2G6 (18) | phospholipase A | MIR5047 (29) | miRNA |
| ANKLE1 (VIII) | endonuclease | LARP6 (XIX) | translational regulator | LOC105374476 (8) | uncharacterized, affiliated with ncRNA class | THUMPD3-AS1 (19) | lncRNA | MXRA5 (30) | matrix-remodelling associated protein |
| SERPING1 (IX) | protease inhibitor | SIX2 (XX) | transcription regulator | ZC3H12A (9) | transcriptional activator | MGEA5 (20) | O-GlcNAcase & Acetyltransferase | CAPN8 (31) | cysteine peptidase |
| TNC (X) | ECM glycoprotein | RPL13 (XXI) | translational regulator | RIMKLB (10) | N-Acetylaspartyl-Glutamate Synthetase | PSG8 (21) | glycoprotein | LENG8 (32) | Member of leukocyte receptor cluster |
| TLDC2 (XI) | OXR1 protein | SMAD7 (XXII) | transcription regulator | FOXO1 (11) | transcription regulator | SH3D21 (22) | nuclear protein | ||
| Disease Pattern (Σ Involved Molecules) | Involved Molecules |
|---|---|
| Cancer in general (Σ 43) | I, II, III, IV, V, VI, VII, VIII, IX, X, XI, XII, XIII, XV, XVI, XVII, XVIII, XIX, XX, XXI, XXII, 1, 2, 3, 4, 6, 7, 9, 10, 11, 12, 13, 14, 17, 18, 20, 21, 22, 27, 28, 30, 31, 32 |
| Lung cancer (Σ 9) | II, IV, V, X, XVIII, 3, 4, 7, 30 |
| Gastrointestinal cancer (Σ 40) | I, II, III, IV, V, VI, VII, VIII, IX, X, XI, XII, XIII, XV, XVI, XVII, XIX, XX, XXII, 1, 2, 3, 4, 7, 9, 10, 11, 12, 13, 14, 17, 18, 20, 21, 22, 27, 28, 30, 31, 32 |
| Head and neck cancer (Σ 19) | IV, 4, 27, VII, 3, II, 11, XV, XVII, 30, 18, 21, IX, XX, XXII, XII, V, X, 9 |
| Neoplasia of epithelial tissue (Σ 41) | I, II, III, IV, V, VI, VII, VIII, IX, X, XI, XII, XIII, XV, XVI, XVII, XVIII, XIX, XX, XXII, 1, 3, 4, 6, 7, 9, 10, 11, 12, 13, 14, 17, 18, 20, 21, 22, 27, 28, 30, 31, 32 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schildgen, V.; Pieper, M.; Khalfaoui, S.; Arnold, W.H.; Schildgen, O. Human Bocavirus Infection of Permanent Cells Differentiated to Air-Liquid Interface Cultures Activates Transcription of Pathways Involved in Tumorigenesis. Cancers 2018, 10, 410. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers10110410
Schildgen V, Pieper M, Khalfaoui S, Arnold WH, Schildgen O. Human Bocavirus Infection of Permanent Cells Differentiated to Air-Liquid Interface Cultures Activates Transcription of Pathways Involved in Tumorigenesis. Cancers. 2018; 10(11):410. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers10110410
Chicago/Turabian StyleSchildgen, Verena, Monika Pieper, Soumaya Khalfaoui, Wolfgang H. Arnold, and Oliver Schildgen. 2018. "Human Bocavirus Infection of Permanent Cells Differentiated to Air-Liquid Interface Cultures Activates Transcription of Pathways Involved in Tumorigenesis" Cancers 10, no. 11: 410. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers10110410