Stemness, Pluripotentiality, and Wnt Antagonism: sFRP4, a Wnt antagonist Mediates Pluripotency and Stemness in Glioblastoma

 ,
,  and
and 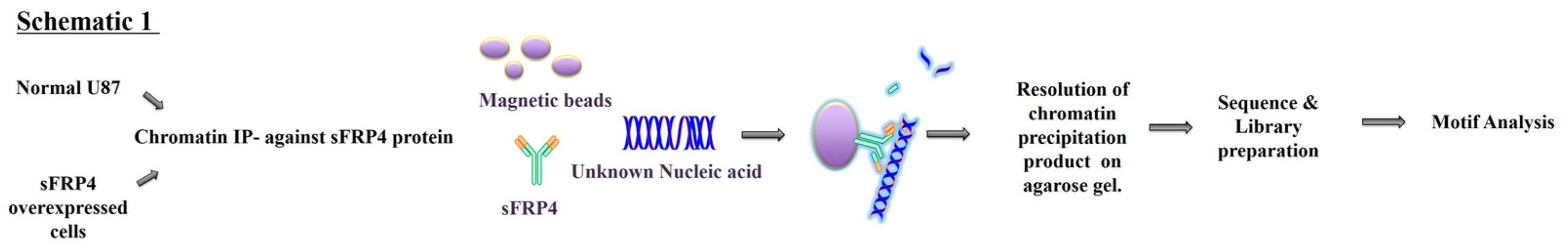{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. sFRP4 Overexpression
2.3. sFRP4 siRNA Silencing
2.4. Cell Viability, Proliferation, Reactive Oxygen Species (ROS), and Caspase Assays
2.5. Secondary Sphere Forming Assay
2.6. Chromatin Immunoprecipitation (ChIP) Sequencing
2.7. MicroRNA 885 Analysis
2.8. RNA Sequencing
2.9. Bioinformatics Analysis
2.10. sFRP4 RNA Interference
2.11. Conversion of WJMSCs into ESC-Like Cells
2.12. Real-Time Quantitative PCR
2.13. Immunocytochemistry
2.14. Western Blotting
2.15. Statistical Analysis
3. Results
3.1. sFRP4 Overexpression has an Anti-Proliferative Effect in Glioma Cell Lines
3.2. sFRP4 OE Cells Induces Apoptosis, Inhibits Cancer Stemness
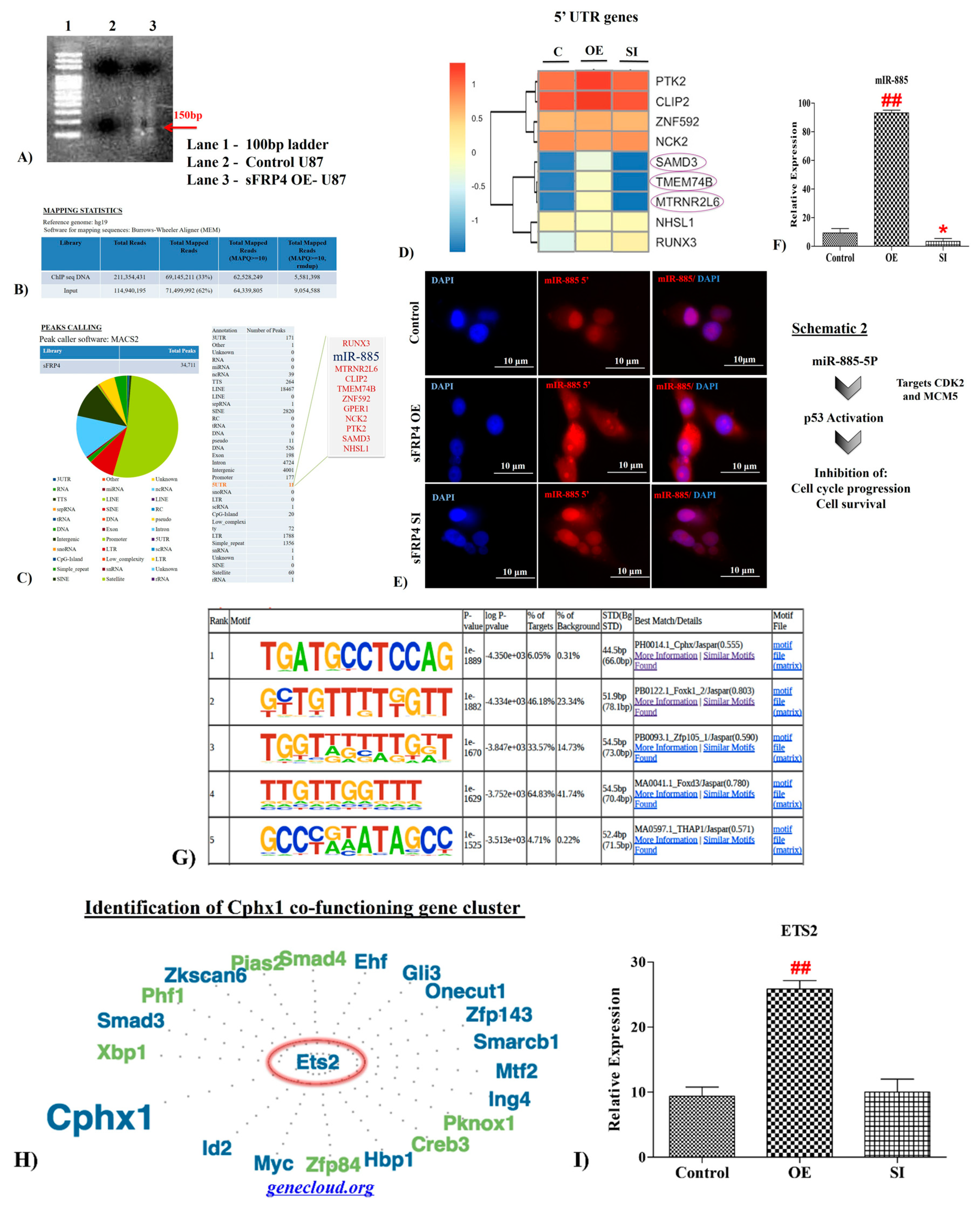
3.3. sFRP4 Localized in the Nucleus with a DNA Binding Ability
3.4. Senescence Related Genes were Identified in sFRP4 ChIP Sequencing Analysis of sFRP4 OE Cells
3.4.1. UTR Analysis
3.4.2. MicroRNA885 Upregulation in sFRP4 OE
3.5. Motif Analysis
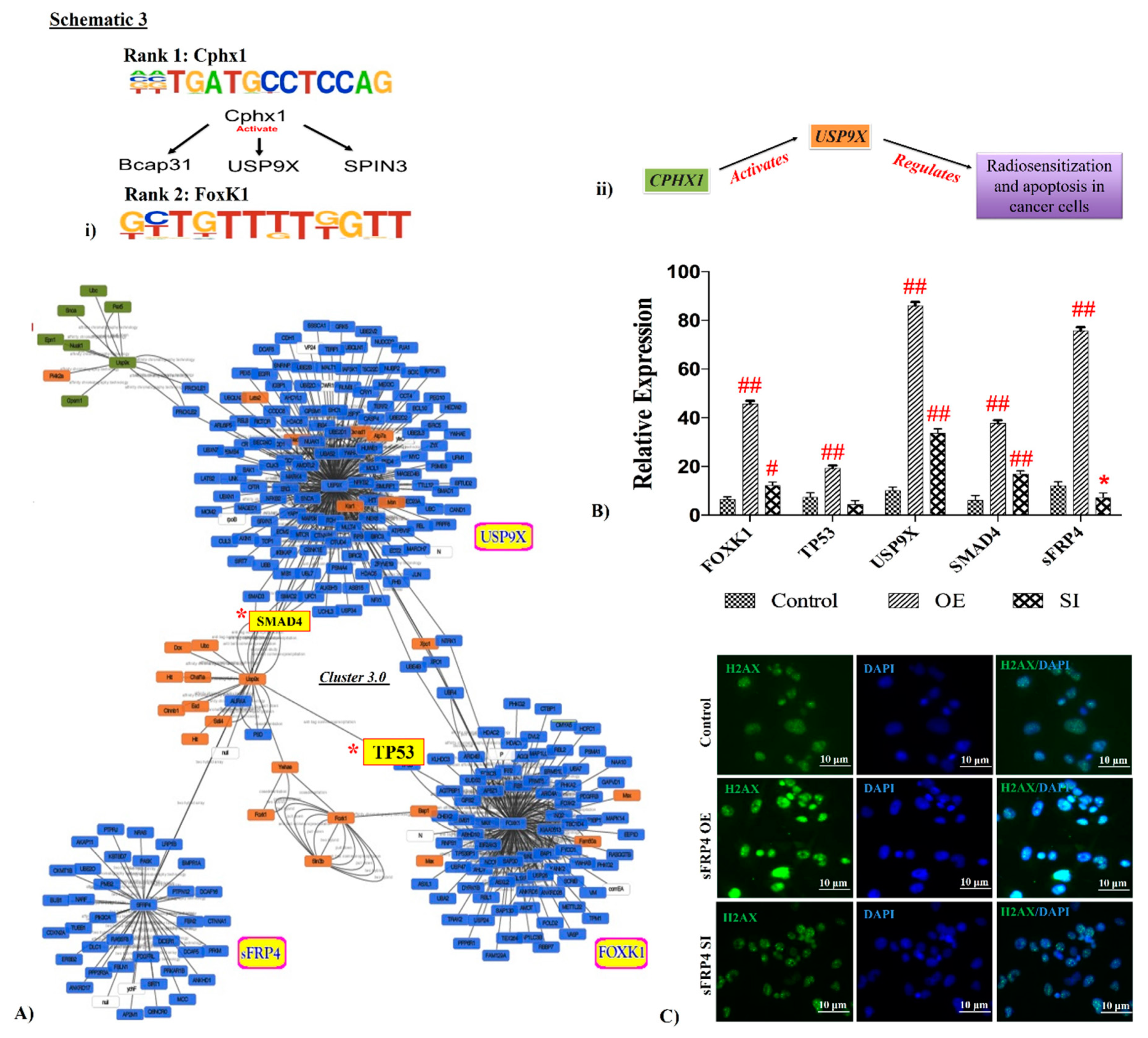
3.6. Interlinking Analysis of sFRP4 with ChIP Motifs
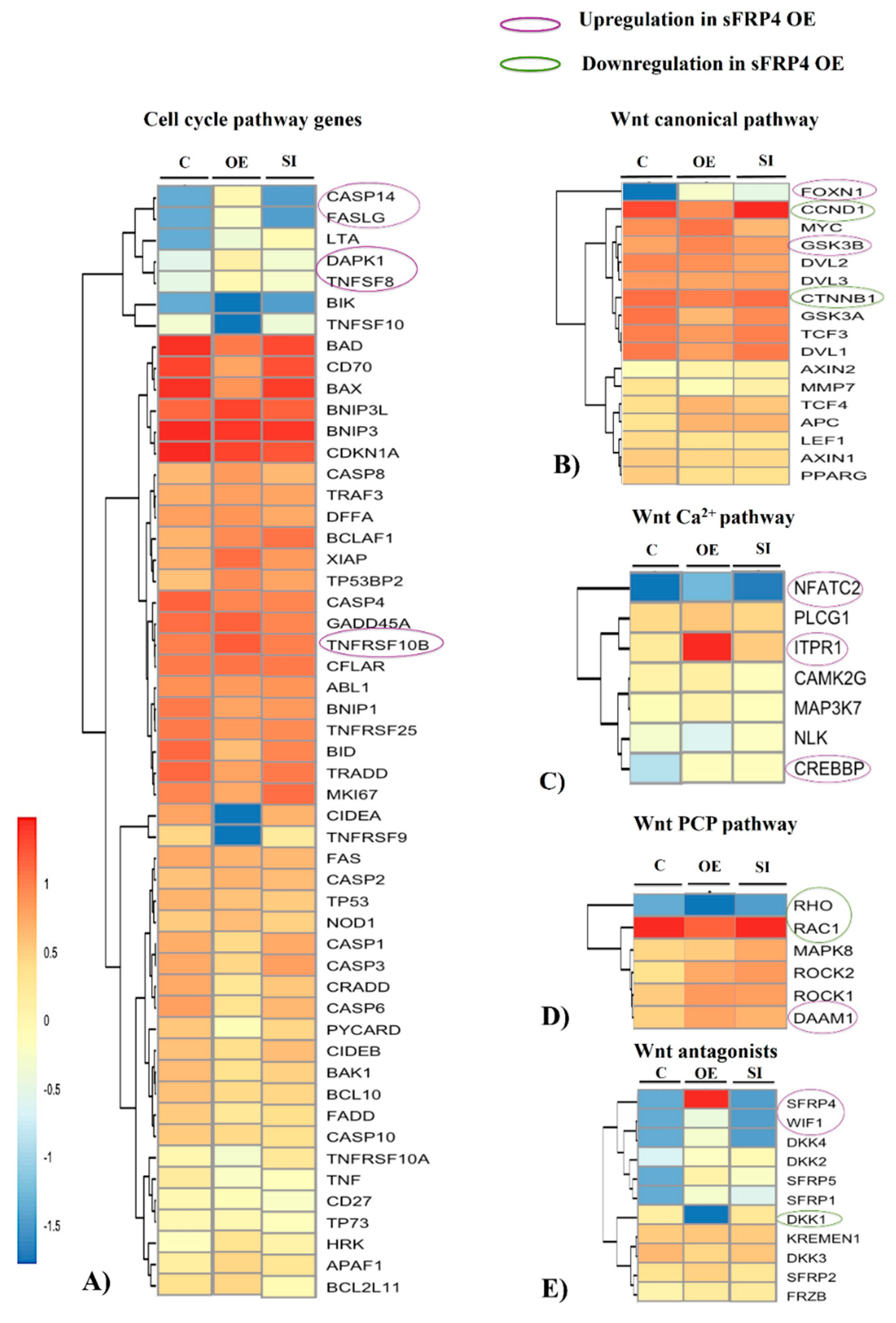
3.7. Whole Transcriptome Analysis of sFRP4 OE and sFRP4 SI
3.7.1. Cell Cycle Pathway Genes
3.7.2. Wnt Canonical Pathway
3.7.3. Wnt Non-Canonical Pathways
3.7.4. Wnt Antagonists
3.7.5. Wnt Receptors and Co-Receptors
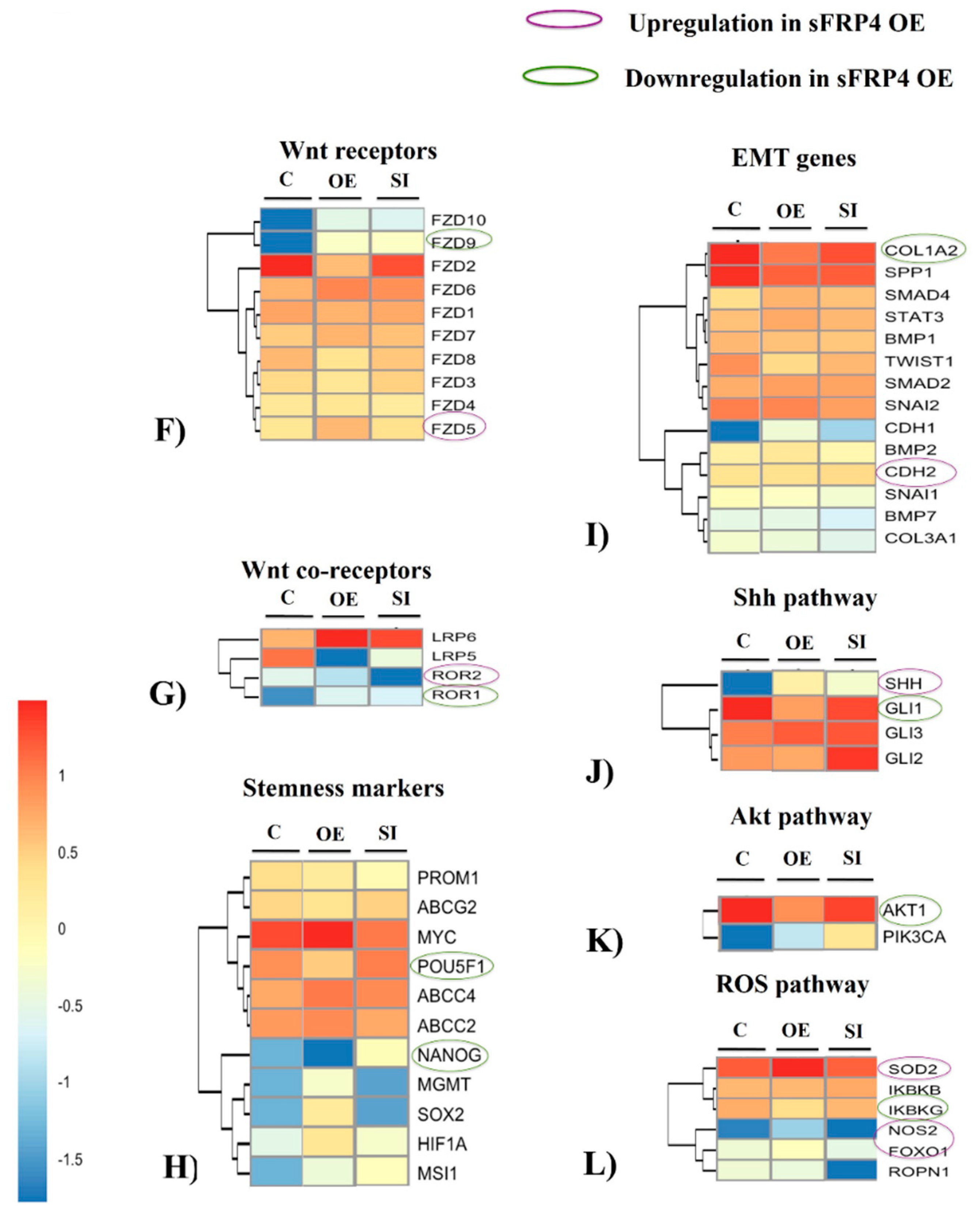
3.7.6. Stemness and Epithelial-Mesenchymal Transition (EMT) Markers
3.7.7. Sonic Hedgehog (Shh), pI3K-AKT, and ROS Pathways
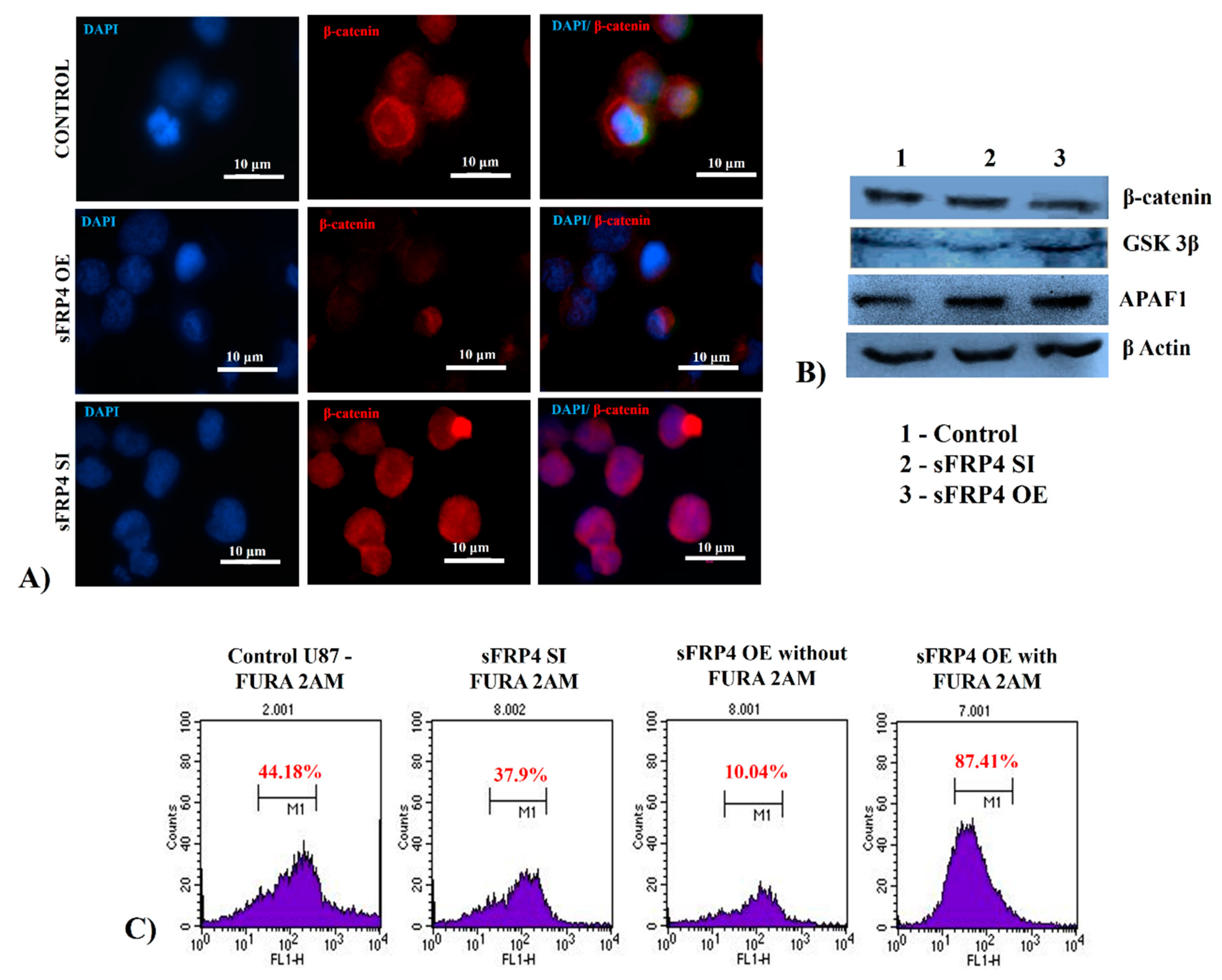
3.8. Quantification of Wnt Pathway Proteins and Functional Activity
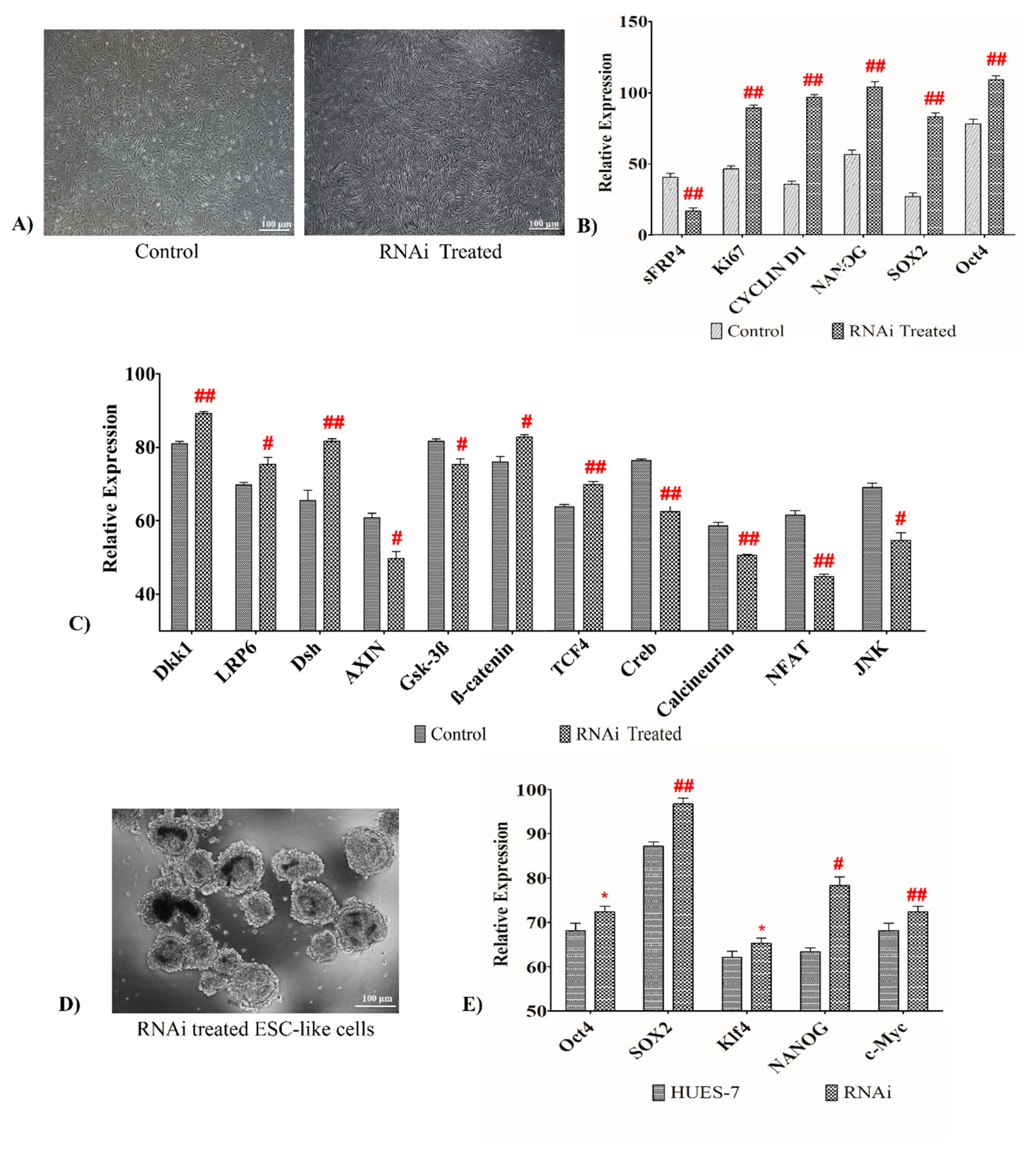
3.9. sFRP4 Silencing Increases Pluripotent Property by Activating Wnt Canonical Pathway in Human Wharton’s Jelly Mesenchymal Stem Cells
3.10. sFRP4 Knockdown Elicits Embryonic Stem Cell-Like Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Singh, S.K.; Clarke, I.D.; Terasaki, M.; Bonn, V.E.; Hawkins, C.; Squire, J.; Dirks, P.B. Identification of a cancer stem cell in human brain tumors. Cancer Res. 2003, 63, 5821–5828. [Google Scholar] [PubMed]
- Altaner, C. Glioblastoma and stem cells. Neoplasma 2008, 55, 369–374. [Google Scholar] [PubMed]
- Yuan, X.; Curtin, J.; Xiong, Y.; Liu, G.; Waschsmann-Hogiu, S.; Farkas, D.L.; Black, K.L.; John, S.Y. Isolation of cancer stem cells from adult glioblastoma multiforme. Oncogene 2004, 23, 9392. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Guo, Y.; Wang, X.; Zhao, H.; Ji, Z.; Cheng, C.; Li, L.; Fang, Y.; Xu, D.; Zhu, H.H. WNT/β-catenin directs self-renewal symmetric cell division of hTERThigh prostate cancer stem cells. Cancer Res. 2017, 77, 2534–2547. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.-B.; Kim, J.-Y.; Cho, S.-D.; Park, K.-S.; Jung, J.-Y.; Lee, H.-Y.; Hong, I.-S.; Nam, J.-S. Blockade of Wnt/β-catenin signaling suppresses breast cancer metastasis by inhibiting CSC-like phenotype. Sci. Rep. 2015, 5, 12465. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Bergevoet, S.M.; Gilissen, C.; de Witte, T.; Jansen, J.H.; van der Reijden, B.A.; Raymakers, R.A. Hematopoietic stem cells exhibit a specific ABC transporter gene expression profile clearly distinct from other stem cells. BMC Pharmacol. 2010, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H.; Sasaki, S.; Suzuki, H.; Toyota, M.; Maruyama, R.; Nojima, M.; Yamamoto, H.; Omata, M.; Tokino, T.; Imai, K. Frequent epigenetic inactivation of SFRP genes in hepatocellular carcinoma. J. Gastroenterol. 2008, 43, 378–389. [Google Scholar] [CrossRef] [Green Version]
- Pannone, G.; Bufo, P.; Santoro, A.; Franco, R.; Aquino, G.; Longo, F.; Botti, G.; Serpico, R.; Cafarelli, B.; Abbruzzese, A. WNT pathway in oral cancer: Epigenetic inactivation of WNT-inhibitors. Oncol. Rep. 2010, 24, 1035–1041. [Google Scholar] [Green Version]
- Schiefer, L.; Visweswaran, M.; Perumal, V.; Arfuso, F.; Groth, D.; Newsholme, P.; Warrier, S.; Dharmarajan, A. Epigenetic regulation of the secreted frizzled-related protein family in human glioblastoma multiforme. Cancer Gene Ther. 2014, 21, 297. [Google Scholar] [CrossRef]
- Warrier, S.; Balu, S.K.; Kumar, A.P.; Millward, M.; Dharmarajan, A. Wnt antagonist, secreted frizzled-related protein 4 (sFRP4), increases chemotherapeutic response of glioma stem-like cells. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2014, 21, 93–102. [Google Scholar] [CrossRef]
- Bhuvanalakshmi, G.; Arfuso, F.; Millward, M.; Dharmarajan, A.; Warrier, S. Secreted frizzled-related protein 4 inhibits glioma stem-like cells by reversing epithelial to mesenchymal transition, inducing apoptosis and decreasing cancer stem cell properties. PLoS ONE 2015, 10, e0127517. [Google Scholar]
- Warrier, S.; Bhuvanalakshmi, G.; Arfuso, F.; Rajan, G.; Millward, M.; Dharmarajan, A. Cancer stem-like cells from head and neck cancers are chemosensitized by the Wnt antagonist, sFRP4, by inducing apoptosis, decreasing stemness, drug resistance and epithelial to mesenchymal transition. Cancer Gene Ther. 2014, 21, 381. [Google Scholar] [CrossRef]
- Bhuvanalakshmi, G.; Rangappa, K.S.; Dharmarajan, A.; Sethi, G.; Kumar, A.P.; Warrier, S. Breast Cancer Stem-Like Cells Are Inhibited by Diosgenin, a Steroidal Saponin, by the Attenuation of the Wnt β-Catenin Signaling via the Wnt Antagonist Secreted Frizzled Related Protein-4. Front. Pharmacol. 2017, 8, 124. [Google Scholar] [CrossRef] [PubMed]
- Bhuvanalakshmi, G.; Arfuso, F.; Kumar, A.P.; Dharmarajan, A.; Warrier, S. Epigenetic reprogramming converts human Wharton’s jelly mesenchymal stem cells into functional cardiomyocytes by differential regulation of Wnt mediators. Stem Cell Res. Ther. 2017, 8, 185. [Google Scholar] [CrossRef] [PubMed]
- Raap, A.K.; van de Rijke, F.M.; Dirks, R.W. mRNA in situ hybridization to in vitro cultured cells. In In Situ Hybridization Protocols; Humana Press: Totowa, NJ, USA, 1994; Volume 33, pp. 293–300. [Google Scholar]
- Szilágyi, A.; Skolnick, J. Efficient prediction of nucleic acid binding function from low-resolution protein structures. J. Mol. Biol. 2006, 358, 922–933. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Zhang, Y.; Liu, T.; Meyer, C.A.; Eeckhoute, J.; Johnson, D.S.; Bernstein, B.E.; Nusbaum, C.; Myers, R.M.; Brown, M.; Li, W. Model-based analysis of ChIP-Seq (MACS). Genome Biol. 2008, 9, R137. [Google Scholar] [CrossRef]
- Khomtchouk, B.B.; Van Booven, D.J.; Wahlestedt, C. HeatmapGenerator: High performance RNAseq and microarray visualization software suite to examine differential gene expression levels using an R and C++ hybrid computational pipeline. Source Code Biol. Med. 2014, 9, 30. [Google Scholar] [CrossRef]
- Bhaskaran, M.; Kolliputi, N.; Wang, Y.; Gou, D.; Chintagari, N.R.; Liu, L. Trans-differentiation of alveolar epithelial type II cells to type I cells involves autocrine signaling by transforming growth factor β1 through the Smad pathway. J. Biol. Chem. 2007, 282, 3968–3976. [Google Scholar] [CrossRef]
- André, E.; Beckerandre, M. Expression of an N-terminally truncated form of human focal adhesion kinase in brain. Biochem. Biophys. Res. Commun. 1993, 190, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Afanasyeva, E.A.; Mestdagh, P.; Kumps, C.; Vandesompele, J.; Ehemann, V.; Theissen, J.; Fischer, M.; Zapatka, M.; Brors, B.; Savelyeva, L. MicroRNA miR-885-5p targets CDK2 and MCM5, activates p53 and inhibits proliferation and survival. Cell Death Differ. 2011, 18, 974. [Google Scholar] [CrossRef] [PubMed]
- Madissoon, E.; Jouhilahti, E.-M.; Vesterlund, L.; Töhönen, V.; Krjutškov, K.; Petropoulos, S.; Einarsdottir, E.; Linnarsson, S.; Lanner, F.; Månsson, R. Characterization and target genes of nine human PRD-like homeobox domain genes expressed exclusively in early embryos. Sci. Rep. 2016, 6, 28995. [Google Scholar] [CrossRef]
- Wolvetang, E.; Bradfield, O.; Hatzistavrou, T.; Crack, P.; Busciglio, J.; Kola, I.; Hertzog, P. Overexpression of the chromosome 21 transcription factor Ets2 induces neuronal apoptosis. Neurobiol. Dis. 2003, 14, 349–356. [Google Scholar] [CrossRef]
- Atsumi, Y.; Inase, A.; Osawa, T.; Sugihara, E.; Sakasai, R.; Fujimori, H.; Teraoka, H.; Saya, H.; Kanno, M.; Tashiro, F. The Arf/p53 protein module, which induces apoptosis, down-regulates histone H2AX to allow normal cells to survive in the presence of anti-cancer drugs. J. Biol. Chem. 2013, 288, 13269–13277. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, M.K.; Shao, C.; Wang, J.; Wei, Q.; Wang, X.; Collier, Z.; Tang, S.; Liu, H.; Zhang, F.; Huang, J. Wnt/β-catenin signaling plays an ever-expanding role in stem cell self-renewal, tumorigenesis and cancer chemoresistance. Genes Dis. 2016, 3, 11–40. [Google Scholar] [CrossRef]
- Trowbridge, J.J.; Xenocostas, A.; Moon, R.T.; Bhatia, M. Glycogen synthase kinase-3 is an in vivo regulator of hematopoietic stem cell repopulation. Nat. Med. 2006, 12, 89. [Google Scholar] [CrossRef] [PubMed]
- Balciunaite, G.; Keller, M.P.; Balciunaite, E.; Piali, L.; Zuklys, S.; Mathieu, Y.D.; Gill, J.; Boyd, R.; Sussman, D.J.; Holländer, G.A. Wnt glycoproteins regulate the expression of FoxN1, the gene defective in nude mice. Nat. Immunol. 2002, 3, 1102. [Google Scholar] [CrossRef]
- Berridge, M.J. Inositol trisphosphate and calcium signalling. Nature 1993, 361, 315. [Google Scholar] [CrossRef]
- Hirota, J.; Hideaki, A.; Hamada, K.; Mikoshiba, K. Carbonic anhydrase-related protein is a novel binding protein for inositol 1, 4, 5-trisphosphate receptor type 1. Biochem. J. 2003, 372, 435–441. [Google Scholar] [CrossRef]
- Pinson, K.I.; Brennan, J.; Monkley, S.; Avery, B.J.; Skarnes, W.C. An LDL-receptor-related protein mediates Wnt signalling in mice. Nature 2000, 407, 535. [Google Scholar] [CrossRef]
- Gray, J.D.; Kholmanskikh, S.; Castaldo, B.S.; Hansler, A.; Chung, H.; Klotz, B.; Singh, S.; Brown, A.M.; Ross, M.E. LRP6 exerts non-canonical effects on Wnt signaling during neural tube closure. Hum. Mol. Genet. 2013, 22, 4267–4281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryja, V.; Andersson, E.R.; Schambony, A.; Esner, M.; Bryjová, L.; Biris, K.K.; Hall, A.C.; Kraft, B.; Cajanek, L.; Yamaguchi, T.P. The extracellular domain of Lrp5/6 inhibits noncanonical Wnt signaling in vivo. Mol. Biol. Cell 2009, 20, 924–936. [Google Scholar] [CrossRef] [PubMed]
- Moskalev, E.A.; Luckert, K.; Vorobjev, I.A.; Mastitsky, S.E.; Gladkikh, A.A.; Stephan, A.; Schrenk, M.; Kaplanov, K.D.; Kalashnikova, O.B.; Pötz, O. Concurrent epigenetic silencing of wnt/β-catenin pathway inhibitor genes in B cell chronic lymphocytic leukaemia. BMC Cancer 2012, 12, 213. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F.; Ukegjini, K.; Nixdorf, S.; Ford, C.E.; Olivier, J.; Caduff, R.; Scurry, J.P.; Guertler, R.; Hornung, D.; Mueller, R. Loss of secreted frizzled-related protein 4 correlates with an aggressive phenotype and predicts poor outcome in ovarian cancer patients. PLoS ONE 2012, 7, e31885. [Google Scholar] [CrossRef] [PubMed]
- Finch, P.W.; He, X.; Kelley, M.J.; Üren, A.; Schaudies, R.P.; Popescu, N.C.; Rudikoff, S.; Aaronson, S.A.; Varmus, H.E.; Rubin, J.S. Purification and molecular cloning of a secreted, Frizzled-related antagonist of Wnt action. Proc. Natl. Acad. Sci. USA 1997, 94, 6770–6775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melkonyan, H.S.; Chang, W.C.; Shapiro, J.P.; Mahadevappa, M.; Fitzpatrick, P.A.; Kiefer, M.C.; Tomei, L.D.; Umansky, S.R. SARPs: A family of secreted apoptosis-related proteins. Proc. Natl. Acad. Sci. USA 1997, 94, 13636–13641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, V.; Ke, G.; Dharmarajan, A.; Bielke, W.; Artuso, L.; Saurer, S.; Friis, R. DDC-4, an apoptosis-associated gene, is a secreted frizzled relative. FEBS Lett. 1997, 417, 385–389. [Google Scholar] [CrossRef] [Green Version]
- Leyns, L.; Bouwmeester, T.; Kim, S.-H.; Piccolo, S.; De Robertis, E.M. Frzb-1 is a secreted antagonist of Wnt signaling expressed in the Spemann organizer. Cell 1997, 88, 747–756. [Google Scholar] [CrossRef]
- Üren, A.; Reichsman, F.; Anest, V.; Taylor, W.G.; Muraiso, K.; Bottaro, D.P.; Cumberledge, S.; Rubin, J.S. Secreted frizzled-related protein-1 binds directly to Wingless and is a biphasic modulator of Wnt signaling. J. Biol. Chem. 2000, 275, 4374–4382. [Google Scholar] [CrossRef]
- Waring, M.; Britten, R.J. Nucleotide sequence repetition: A rapidly reassociating fraction of mouse DNA. Science 1966, 154, 791–794. [Google Scholar] [CrossRef]
- Britten, R.J.; Kohne, D.E. Repeated sequences in DNA. Science 1968, 161, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Zelivianski, S.; Cooley, A.; Kall, R.; Jeruss, J.S. CDK4-Mediated Phosphorylation Inhibits Smad3 Activity in Cyclin D Overexpressing Breast Cancer Cells. Mol. Cancer Res. 2010. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Jia, X.; Zhao, J.; Cui, R.; Yan, M. Substance P promotes hepatic stellate cell proliferation and activation via the TGF-β1/Smad-3 signaling pathway. Toxicol. Appl. Pharmacol. 2017, 329, 293–300. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Peng, Z.; Luo, Y.; Wang, L.; Yu, P.; Deng, W.; An, Y.; Shi, T.; Ma, D. High-throughput functional screening for autophagy-related genes and identification of TM9SF1 as an autophagosome-inducing gene. Autophagy 2009, 5, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Bodzioch, M.; Lapicka-Bodzioch, K.; Zapala, B.; Kamysz, W.; Kiec-Wilk, B.; Dembinska-Kiec, A. Evidence for potential functionality of nuclearly-encoded humanin isoforms. Genomics 2009, 94, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, Q.-L.; Sun, W.; Chandrasekharan, P.; Cheng, H.S.; Ying, Z.; Lakshmanan, M.; Raju, A.; Tenen, D.G.; Cheng, S.-Y. Non-canonical NF-κB signalling and ETS1/2 cooperatively drive C250T mutant TERT promoter activation. Nat. Cell Biol. 2015, 17, 1327. [Google Scholar] [CrossRef] [PubMed]
- Capper, D.; Gaiser, T.; Hartmann, C.; Habel, A.; Mueller, W.; Herold-Mende, C.; von Deimling, A.; Siegelin, M.D. Stem-cell-like glioma cells are resistant to TRAIL/Apo2L and exhibit down-regulation of caspase-8 by promoter methylation. Acta Neuropathol. 2009, 117, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Owen-Schaub, L.B.; Zhang, W.; Cusack, J.C.; Angelo, L.S.; Santee, S.M.; Fujiwara, T.; Roth, J.A.; Deisseroth, A.B.; Zhang, W.-W.; Kruzel, E. Wild-type human p53 and a temperature-sensitive mutant induce Fas/APO-1 expression. Mol. Cell. Biol. 1995, 15, 3032–3040. [Google Scholar] [CrossRef]
- Koster, R.; Timmer-Bosscha, H.; Bischoff, R.; Gietema, J.A.; de Jong, S. Disruption of the MDM2–p53 interaction strongly potentiates p53-dependent apoptosis in cisplatin-resistant human testicular carcinoma cells via the Fas/FasL pathway. Cell Death Dis. 2011, 2, e148. [Google Scholar] [CrossRef]
- Wozniak, A.L.; Wang, X.; Stieren, E.S.; Scarbrough, S.G.; Elferink, C.J.; Boehning, D. Requirement of biphasic calcium release from the endoplasmic reticulum for Fas-mediated apoptosis. J. Cell Biol. 2006, 175, 709–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinmann, C.; Landsverk, M.L.; Barral, J.M.; Boehning, D. Requirement of inositol 1, 4, 5-trisphosphate receptors for tumor-mediated lymphocyte apoptosis. J. Biol. Chem. 2008, 283, 13506–13509. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.; Lin, Z.; Leung, P. Role of reactive oxygen species in brucein D-mediated p38-mitogen-activated protein kinase and nuclear factor-κB signalling pathways in human pancreatic adenocarcinoma cells. Br. J. Cancer 2010, 102, 583. [Google Scholar] [CrossRef] [PubMed]
- Fruehauf, J.P.; Meyskens, F.L. Reactive oxygen species: A breath of life or death? Clin. Cancer Res. 2007, 13, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Pias, E.K.; Ekshyyan, O.Y.; Rhoads, C.A.; Fuseler, J.; Harrison, L.; Aw, T.Y. Differential effects of superoxide dismutase isoform expression on hydroperoxide-induced apoptosis in PC-12 cells. J. Biol. Chem. 2003, 278, 13294–13301. [Google Scholar] [CrossRef] [PubMed]
- Vo, B.T.; Morton, D., Jr.; Komaragiri, S.; Millena, A.C.; Leath, C.; Khan, S.A. TGF-β effects on prostate cancer cell migration and invasion are mediated by PGE2 through activation of PI3K/AKT/mTOR pathway. Endocrinology 2013, 154, 1768–1779. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Graham, P.; Hao, J.; Ni, J.; Bucci, J.; Cozzi, P.; Kearsley, J.; Li, Y. PI3K/Akt/mTOR pathway inhibitors enhance radiosensitivity in radioresistant prostate cancer cells through inducing apoptosis, reducing autophagy, suppressing NHEJ and HR repair pathways. Cell Death Dis. 2014, 5, e1437. [Google Scholar] [CrossRef]
- Jones, M.B.; Chu, C.H.; Pendleton, J.C.; Betenbaugh, M.J.; Shiloach, J.; Baljinnyam, B.; Rubin, J.S.; Shamblott, M.J. Proliferation and pluripotency of human embryonic stem cells maintained on type I collagen. Stem Cells Dev. 2010, 19, 1923–1935. [Google Scholar] [CrossRef]
- He, R.; Xhabija, B.; Al-Qanber, B.; Kidder, B.L. OCT4 supports extended LIF-independent self-renewal and maintenance of transcriptional and epigenetic networks in embryonic stem cells. Sci. Rep. 2017, 7, 16360. [Google Scholar] [CrossRef]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhuvanalakshmi, G.; Gamit, N.; Patil, M.; Arfuso, F.; Sethi, G.; Dharmarajan, A.; Prem Kumar, A.; Warrier, S. Stemness, Pluripotentiality, and Wnt Antagonism: sFRP4, a Wnt antagonist Mediates Pluripotency and Stemness in Glioblastoma. Cancers 2019, 11, 25. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11010025
Bhuvanalakshmi G, Gamit N, Patil M, Arfuso F, Sethi G, Dharmarajan A, Prem Kumar A, Warrier S. Stemness, Pluripotentiality, and Wnt Antagonism: sFRP4, a Wnt antagonist Mediates Pluripotency and Stemness in Glioblastoma. Cancers. 2019; 11(1):25. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11010025
Chicago/Turabian StyleBhuvanalakshmi, Gurubharathi, Naisarg Gamit, Manasi Patil, Frank Arfuso, Gautam Sethi, Arun Dharmarajan, Alan Prem Kumar, and Sudha Warrier. 2019. "Stemness, Pluripotentiality, and Wnt Antagonism: sFRP4, a Wnt antagonist Mediates Pluripotency and Stemness in Glioblastoma" Cancers 11, no. 1: 25. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11010025