Synergistic Antitumorigenic Activity of Calcitriol with Curcumin or Resveratrol is Mediated by Angiogenesis Inhibition in Triple Negative Breast Cancer Xenografts

 ,
,  , ,
, ,  , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Characterization of Cell Lines and Tumors
2.1.1. MBCDF-T and EA.hy926 Cells Expressed VDR and RXR in Different Cell Compartments
2.1.2. MBCDF-T Tumors were ERα, PR, and HER2 Negative with a Mesenchymal-Like Phenotype
2.2. The Phytochemicals and Calcitriol Differentially Regulated the Proliferation of MBCDF-T and EA.hy926 Cells In Vitro
2.3. Depending on the Cell Line, the Experimental Compounds Differentially Regulated the Cell Cycle Profile; particularly, Resveratrol Significantly Induced Endothelial Cell Death
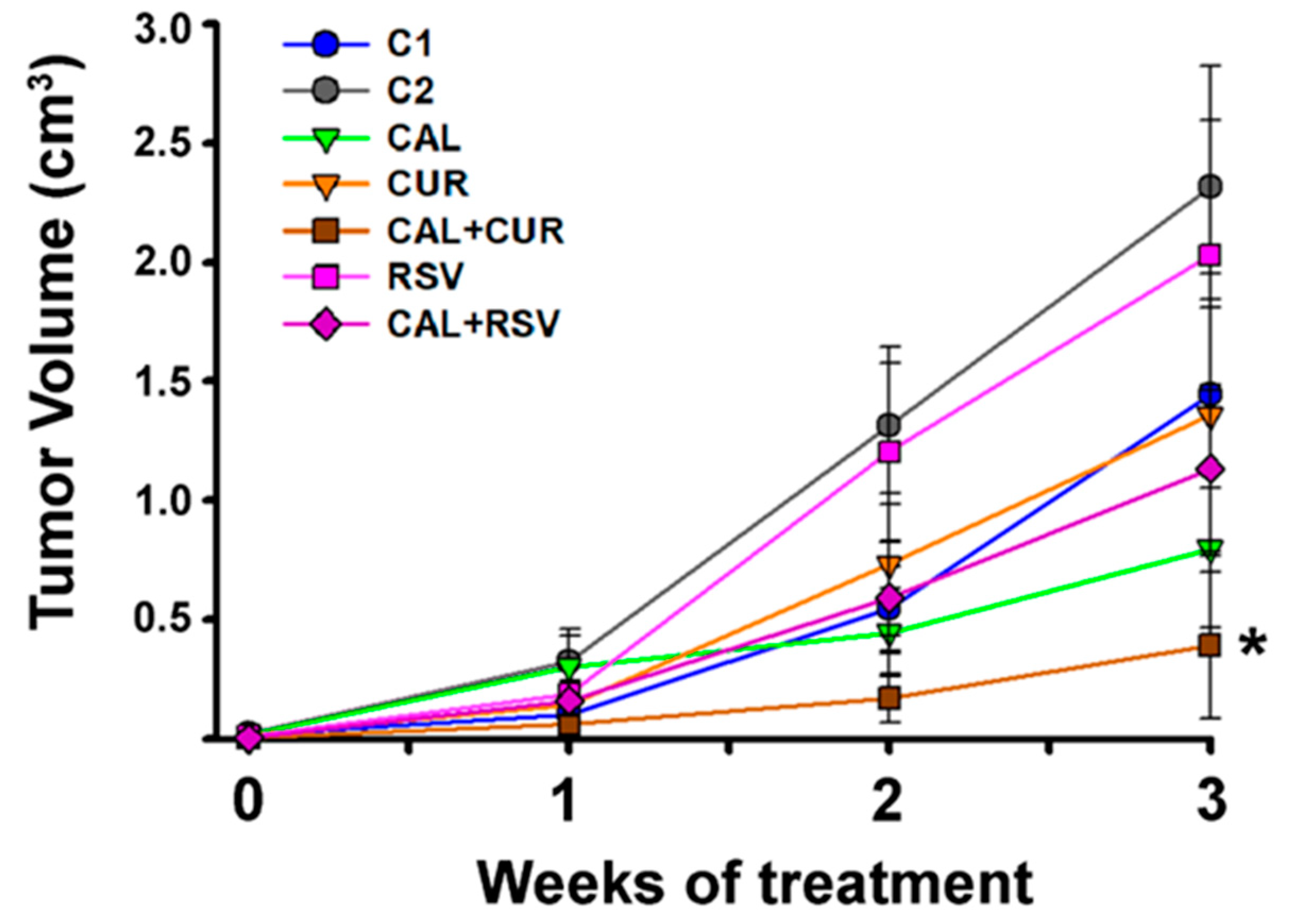
2.4. The In Vivo Combination of Calcitriol and Curcumin Reduced the Number and Size of Breast Cancer Tumors
2.5. Tumors in the Cal+Cur and Cal+Rsv Groups Had a Reduced Intratumor Angiogenic Network Compared to Controls
2.6. Tumor Uptake of Integrin-Targeted Biosensors Was Proportional to the Tumor Size
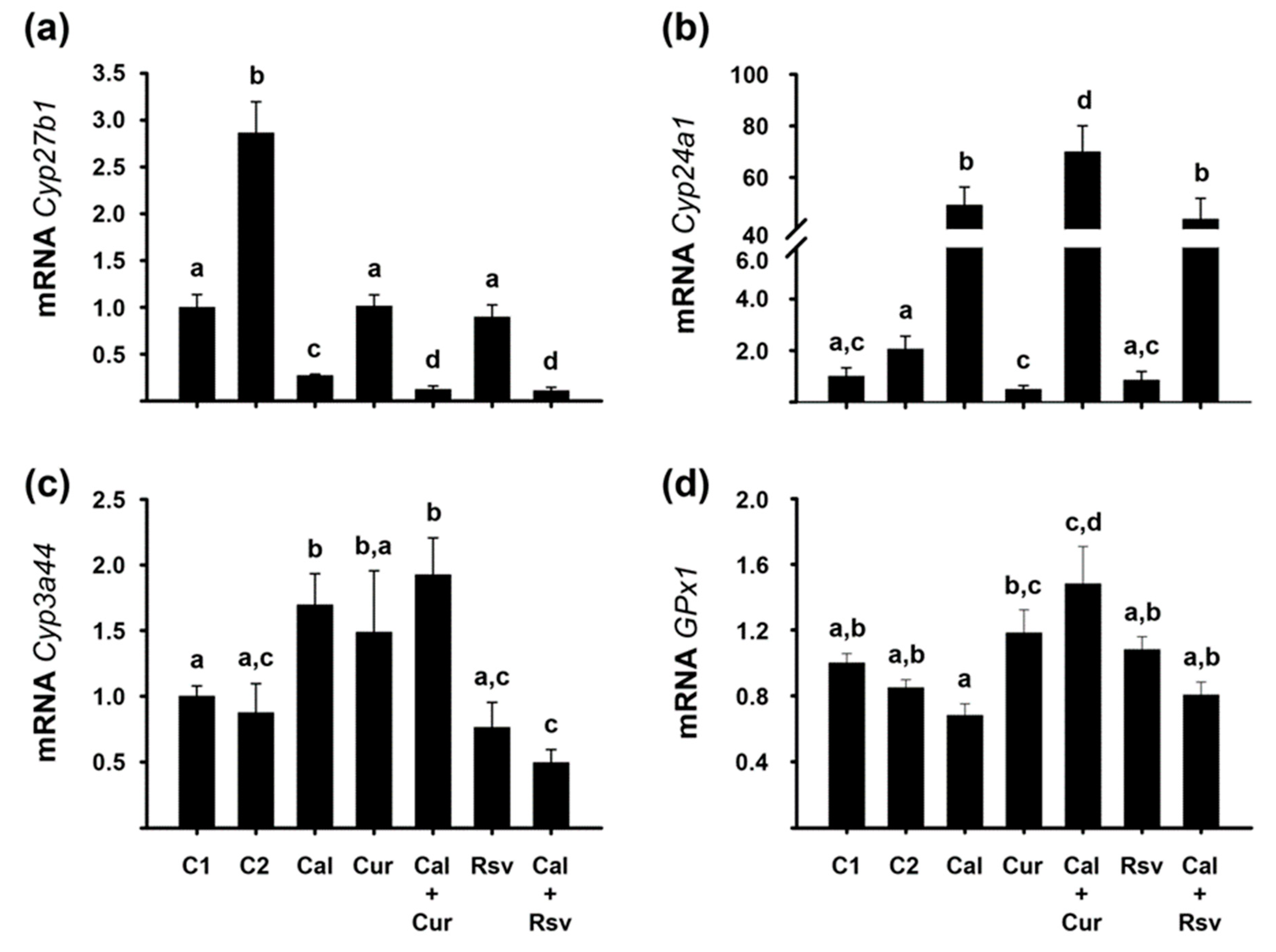
2.7. In Vivo Bioactivity of the Experimental Compounds Was Corroborated by the Modification of Renal Cyp24a1 and Cyp2b1, and Hepatic Cyp3a44 and GPx1 Gene Expression
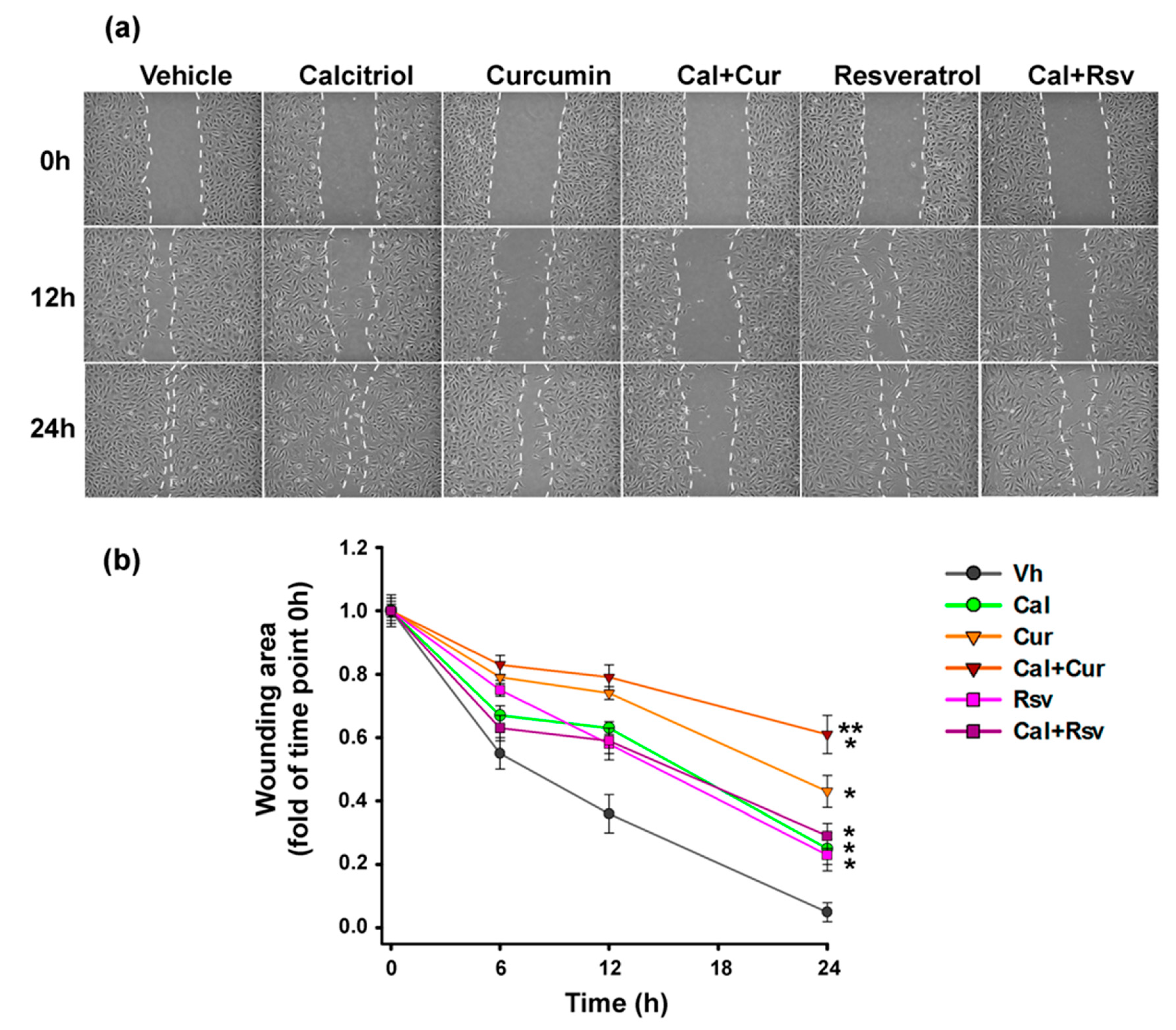
2.8. Curcumin Combined with Calcitriol Was the Most Efficient Strategy to Prevent Endothelial Cells Migration
3. Discussion
3.1. The Phytochemicals Curcumin and Resveratrol Enhanced the Antineoplastic In Vivo Effect of Calcitriol by Inhibiting the Adequate Formation of a Tumor Vessel Network
3.2. The Differential Renal and Hepatic Expression of P450 Cythochromes and GPx1 among the Experimental Groups Reflected the Compounds Interaction and Bioactivity In Vivo
3.3. In Vivo, the Combination of Cal+Cur Prevented Tumor Unset and Progression by Blocking Integrin Signalization in Both Tumor and Endothelial Cells, in which their Migratory Capacity Was also Blocked
3.4. In Vitro Mechanistic Insights and Clinical Implications
3.5. Highlights and Limitations of This Study
4. Materials and Methods
4.1. Reagents
4.2. Ethical Approval
4.3. Cell Culture
4.4. Proliferation Studies
4.5. Cell Cycle Analysis
4.6. Xenotransplantation and Therapeutic Protocol
4.7. In Vivo Imaging Studies of Activated Endothelium
4.7.1. Biosensors/Pharmaceuticals Assessed
4.7.2. Animal Preparation for Imaging
4.7.3. Imaging
Fluorescence and X-Ray Imaging
SPECT/CT Imaging
4.8. Tumor Phenotype Characterization and Immunocytochemical Analysis
4.9. Intratumor Vessel Density Evaluation
4.10. Real Time PCR Amplifications
4.11. In Vitro Wound Healing Assay
4.12. Statistical Analysis
5. Conclusions
- Curcumin potentiated calcitriol antitumorigenic in vivo effects in TNBC by preventing intratumor vessel formation and favoring an antioxidant state.
- Curcumin antiangiogenic effect was mediated by the inhibition of endothelial cells proliferation and integrin expression.
- Resveratrol induced endothelial cell death in vitro, while in vivo inhibited angiogenesis in TNBC tumors only when combined with calcitriol.
- Curcumin solubilization in ethanol promoted its dilution in water, facilitating its administration and improving its absorption and bioavailability.
- Calcitriol coadministered with curcumin represents a promising adjuvant therapy for TNBC by inhibiting tumor growth and angiogenesis.
- Given that calcitriol and its combination with curcumin or resveratrol differentially regulated hepatic Cyp3a44 gene expression, potential pharmacokinetic interactions should be considered when administering these compounds with CYP3A4 substrates in the clinic (including drugs and dietary compounds).
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brem, S.; Cotran, R.; Folkman, J. Tumor angiogenesis: A quantitative method for histologic grading. J. Natl. Cancer Inst. 1972, 48, 347–356. [Google Scholar]
- Kraby, M.R.; Opdahl, S.; Akslen, L.A.; Bofin, A.M. Quantifying tumour vascularity in non-luminal breast cancers. J. Clin. Pathol. 2017, 70, 766–774. [Google Scholar] [CrossRef] [Green Version]
- Niu, S.; Zhu, Q.; Jiang, Y.; Zhu, J.; Xiao, M.; You, S.; Zhou, W.; Xiao, Y. Correlations Among Ultrasound-Guided Diffuse Optical Tomography, Microvessel Density, and Breast Cancer Prognosis. J. Ultrasound Med. 2018, 37, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Weidner, N. Intratumor microvessel density as a prognostic factor in cancer. Am. J. Pathol. 1995, 147, 9–19. [Google Scholar] [PubMed]
- Muindi, J.R.; Johnson, C.S.; Trump, D.L.; Christy, R.; Engler, K.L.; Fakih, M.G. A phase I and pharmacokinetics study of intravenous calcitriol in combination with oral dexamethasone and gefitinib in patients with advanced solid tumors. Cancer Chemother. Pharmacol. 2009, 65, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trump, D.L. Calcitriol and cancer therapy: A missed opportunity. Bone Rep. 2018, 9, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Trump, D.L.; Hershberger, P.A.; Bernardi, R.J.; Ahmed, S.; Muindi, J.; Fakih, M.; Yu, W.D.; Johnson, C.S. Anti-tumor activity of calcitriol: Pre-Clinical and clinical studies. J. Steroid Biochem. Mol. Biol. 2004, 89–90, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Hu, P.; Xie, D.; Qin, Y.; Wang, F.; Wang, H. Meta-analysis of vitamin D, calcium and the prevention of breast cancer. Breast Cancer Res. Treat. 2010, 121, 469–477. [Google Scholar] [CrossRef]
- Berger, U.; McClelland, R.A.; Wilson, P.; Greene, G.L.; Haussler, M.R.; Pike, J.W.; Colston, K.; Easton, D.; Coombes, R.C. Immunocytochemical determination of estrogen receptor, progesterone receptor, and 1,25-dihydroxyvitamin D3 receptor in breast cancer and relationship to prognosis. Cancer Res. 1991, 51, 239–244. [Google Scholar] [PubMed]
- Diaz, L.; Diaz-Munoz, M.; Garcia-Gaytan, A.C.; Mendez, I. Mechanistic Effects of Calcitriol in Cancer Biology. Nutrients 2015, 7, 5020–5050. [Google Scholar] [CrossRef] [Green Version]
- Shan, N.L.; Wahler, J.; Lee, H.J.; Bak, M.J.; Gupta, S.D.; Maehr, H.; Suh, N. Vitamin D compounds inhibit cancer stem-like cells and induce differentiation in triple negative breast cancer. J. Steroid Biochem. Mol. Biol. 2017, 173, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, R.J.; Johnson, C.S.; Modzelewski, R.A.; Trump, D.L. Antiproliferative effects of 1alpha,25-dihydroxyvitamin D(3) and vitamin D analogs on tumor-derived endothelial cells. Endocrinology 2002, 143, 2508–2514. [Google Scholar] [CrossRef] [PubMed]
- Mantell, D.J.; Owens, P.E.; Bundred, N.J.; Mawer, E.B.; Canfield, A.E. 1 alpha,25-dihydroxyvitamin D(3) inhibits angiogenesis in vitro and in vivo. Circ. Res. 2000, 87, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Garcia, N.I.; Palmer, H.G.; Garcia, M.; Gonzalez-Martin, A.; del Rio, M.; Barettino, D.; Volpert, O.; Munoz, A.; Jimenez, B. 1alpha,25-Dihydroxyvitamin D3 regulates the expression of Id1 and Id2 genes and the angiogenic phenotype of human colon carcinoma cells. Oncogene 2005, 24, 6533–6544. [Google Scholar] [CrossRef] [PubMed]
- Grundmann, M.; Haidar, M.; Placzko, S.; Niendorf, R.; Darashchonak, N.; Hubel, C.A.; von Versen-Hoynck, F. Vitamin D improves the angiogenic properties of endothelial progenitor cells. Am. J. Physiol. Cell Physiol. 2012, 303, C954–C962. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Amizuka, N.; Sasaki, T.; Aarts, M.M.; Ozawa, H.; Goltzman, D.; Henderson, J.E.; White, J.H. 1Alpha,25-dihydroxyvitamin D3 promotes vascularization of the chondro-osseous junction by stimulating expression of vascular endothelial growth factor and matrix metalloproteinase 9. J. Bone Miner. Res. 2002, 17, 1604–1612. [Google Scholar] [CrossRef]
- Yamamoto, T.; Kozawa, O.; Tanabe, K.; Akamatsu, S.; Matsuno, H.; Dohi, S.; Hirose, H.; Uematsu, T. 1,25-dihydroxyvitamin D3 stimulates vascular endothelial growth factor release in aortic smooth muscle cells: Role of p38 mitogen-activated protein kinase. Arch. Biochem. Biophys. 2002, 398, 1–6. [Google Scholar] [CrossRef]
- Garcia-Quiroz, J.; Garcia-Becerra, R.; Santos-Martinez, N.; Avila, E.; Larrea, F.; Diaz, L. Calcitriol stimulates gene expression of cathelicidin antimicrobial peptide in breast cancer cells with different phenotype. J. Biomed. Sci. 2016, 23, 78. [Google Scholar] [CrossRef]
- Garcia-Quiroz, J.; Rivas-Suarez, M.; Garcia-Becerra, R.; Barrera, D.; Martinez-Reza, I.; Ordaz-Rosado, D.; Santos-Martinez, N.; Villanueva, O.; Santos-Cuevas, C.L.; Avila, E.; et al. Calcitriol reduces thrombospondin-1 and increases vascular endothelial growth factor in breast cancer cells: Implications for tumor angiogenesis. J. Steroid Biochem. Mol. Biol. 2014, 144 Pt A, 215–222. [Google Scholar] [CrossRef]
- Anisiewicz, A.; Pawlik, A.; Filip-Psurska, B.; Turlej, E.; Dzimira, S.; Milczarek, M.; Gdesz, K.; Papiernik, D.; Jarosz, J.; Klopotowska, D.; et al. Unfavorable effect of calcitriol and its low-calcemic analogs on metastasis of 4T1 mouse mammary gland cancer. Int. J. Oncol. 2018, 52, 103–126. [Google Scholar] [CrossRef]
- Hong, R.L.; Spohn, W.H.; Hung, M.C. Curcumin inhibits tyrosine kinase activity of p185neu and also depletes p185neu. Clin. Cancer Res. 1999, 5, 1884–1891. [Google Scholar] [PubMed]
- Wang, J.; Jiang, Y.F. Natural compounds as anticancer agents: Experimental evidence. World J. Exp. Med. 2012, 2, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Varoni, E.M.; Lo Faro, A.F.; Sharifi-Rad, J.; Iriti, M. Anticancer Molecular Mechanisms of Resveratrol. Front. Nutr. 2016, 3, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arbiser, J.L.; Klauber, N.; Rohan, R.; van Leeuwen, R.; Huang, M.T.; Fisher, C.; Flynn, E.; Byers, H.R. Curcumin is an in vivo inhibitor of angiogenesis. Mol. Med. 1998, 4, 376–383. [Google Scholar] [CrossRef]
- Cao, Y.; Fu, Z.D.; Wang, F.; Liu, H.Y.; Han, R. Anti-angiogenic activity of resveratrol, a natural compound from medicinal plants. J. Asian Nat. Prod. Res. 2005, 7, 205–213. [Google Scholar] [CrossRef]
- Dampf Stone, A.; Batie, S.F.; Sabir, M.S.; Jacobs, E.T.; Lee, J.H.; Whitfield, G.K.; Haussler, M.R.; Jurutka, P.W. Resveratrol potentiates vitamin D and nuclear receptor signaling. J. Cell. Biochem. 2015, 116, 1130–1143. [Google Scholar] [CrossRef]
- Liu, Y.; Chang, R.L.; Cui, X.X.; Newmark, H.L.; Conney, A.H. Synergistic effects of curcumin on all-trans retinoic acid- and 1 alpha,25-dihydroxyvitamin D3-induced differentiation in human promyelocytic leukemia HL-60 cells. Oncol. Res. 1997, 9, 19–29. [Google Scholar]
- Bartik, L.; Whitfield, G.K.; Kaczmarska, M.; Lowmiller, C.L.; Moffet, E.W.; Furmick, J.K.; Hernandez, Z.; Haussler, C.A.; Haussler, M.R.; Jurutka, P.W. Curcumin: A novel nutritionally derived ligand of the vitamin D receptor with implications for colon cancer chemoprevention. J. Nutr. Biochem. 2010, 21, 1153–1161. [Google Scholar] [CrossRef]
- Sokoloski, J.A.; Shyam, K.; Sartorelli, A.C. Induction of the differentiation of HL-60 promyelocytic leukemia cells by curcumin in combination with low levels of vitamin D3. Oncol. Res. 1997, 9, 31–39. [Google Scholar]
- Basheer, L.; Kerem, Z. Interactions between CYP3A4 and Dietary Polyphenols. Oxidative Med. Cell. Longev. 2015, 2015, 854015. [Google Scholar] [CrossRef]
- Deb, S.; Pandey, M.; Adomat, H.; Guns, E.S. Cytochrome P450 3A-mediated microsomal biotransformation of 1alpha,25-dihydroxyvitamin D3 in mouse and human liver: Drug-related induction and inhibition of catabolism. Drug Metab. Dispos. 2012, 40, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Hashizume, T.; Shuhart, M.C.; Davis, C.L.; Nelson, W.L.; Sakaki, T.; Kalhorn, T.F.; Watkins, P.B.; Schuetz, E.G.; Thummel, K.E. Intestinal and hepatic CYP3A4 catalyze hydroxylation of 1alpha,25-dihydroxyvitamin D(3): Implications for drug-induced osteomalacia. Mol. Pharmacol. 2006, 69, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Vuoriluoto, K.; Haugen, H.; Kiviluoto, S.; Mpindi, J.P.; Nevo, J.; Gjerdrum, C.; Tiron, C.; Lorens, J.B.; Ivaska, J. Vimentin regulates EMT induction by Slug and oncogenic H-Ras and migration by governing Axl expression in breast cancer. Oncogene 2011, 30, 1436–1448. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, F.; Chen, X. Integrin alpha(v)beta(3)-Targeted Cancer Therapy. Drug Dev. Res. 2008, 69, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Thompson, P.D.; Jurutka, P.W.; Whitfield, G.K.; Myskowski, S.M.; Eichhorst, K.R.; Dominguez, C.E.; Haussler, C.A.; Haussler, M.R. Liganded VDR induces CYP3A4 in small intestinal and colon cancer cells via DR3 and ER6 vitamin D responsive elements. Biochem. Biophys. Res. Commun. 2002, 299, 730–738. [Google Scholar] [CrossRef]
- Anakk, S.; Huang, W.; Staudinger, J.L.; Tan, K.; Cole, T.J.; Moore, D.D.; Strobel, H.W. Gender dictates the nuclear receptor-mediated regulation of CYP3A44. Drug Metab. Dispos. 2007, 35, 36–42. [Google Scholar] [CrossRef]
- Das, L.; Vinayak, M. Anti-carcinogenic action of curcumin by activation of antioxidant defence system and inhibition of NF-kappaB signalling in lymphoma-bearing mice. Biosci. Rep. 2012, 32, 161–170. [Google Scholar] [CrossRef]
- Deng, R.; Xu, C.; Chen, X.; Chen, P.; Wang, Y.; Zhou, X.; Jin, J.; Niu, L.; Ying, M.; Huang, M.; et al. Resveratrol suppresses the inducible expression of CYP3A4 through the pregnane X receptor. J. Pharmacol. Sci. 2014, 126, 146–154. [Google Scholar] [CrossRef]
- Shindikar, A.; Singh, A.; Nobre, M.; Kirolikar, S. Curcumin and Resveratrol as Promising Natural Remedies with Nanomedicine Approach for the Effective Treatment of Triple Negative Breast Cancer. J. Oncol. 2016, 2016, 9750785. [Google Scholar] [CrossRef]
- Nagao, N.; Nakayama, T.; Etoh, T.; Saiki, I.; Miwa, N. Tumor invasion is inhibited by phosphorylated ascorbate via enrichment of intracellular vitamin C and decreasing of oxidative stress. J. Cancer Res. Clin. Oncol. 2000, 126, 511–518. [Google Scholar] [CrossRef]
- van Rossen, M.E.; Sluiter, W.; Bonthuis, F.; Jeekel, H.; Marquet, R.L.; van Eijck, C.H. Scavenging of reactive oxygen species leads to diminished peritoneal tumor recurrence. Cancer Res. 2000, 60, 5625–5629. [Google Scholar] [PubMed]
- Baliga, M.S.; Wang, H.; Zhuo, P.; Schwartz, J.L.; Diamond, A.M. Selenium and GPx-1 overexpression protect mammalian cells against UV-induced DNA damage. Biol. Trace Elem. Res. 2007, 115, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Gardaneh, M.; Shojaei, S.; Rahimi Shamabadi, A.; Akbari, P. Breast Cancer Cell Apoptosis is Synergistically Induced by Curcumin, Trastuzumab, and Glutathione Peroxidase-1 but Robustly Inhibited by Glial Cell Line-Derived Neurotrophic Factor. Nutr. Cancer 2018, 70, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Quiroz, J.; Garcia-Becerra, R.; Lara-Sotelo, G.; Avila, E.; Lopez, S.; Santos-Martinez, N.; Halhali, A.; Ordaz-Rosado, D.; Barrera, D.; Olmos-Ortiz, A.; et al. Chronic moderate ethanol intake differentially regulates vitamin D hydroxylases gene expression in kidneys and xenografted breast cancer cells in female mice. J. Steroid Biochem. Mol. Biol. 2017, 173, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Javadi, S.; Rostamizadeh, K.; Hejazi, J.; Parsa, M.; Fathi, M. Curcumin mediated down-regulation of alphaV beta3 integrin and up-regulation of pyruvate dehydrogenase kinase 4 (PDK4) in Erlotinib resistant SW480 colon cancer cells. Phytother. Res. 2018, 32, 355–364. [Google Scholar] [CrossRef]
- Simmons, K.M.; Beaudin, S.G.; Narvaez, C.J.; Welsh, J. Gene Signatures of 1,25-Dihydroxyvitamin D3 Exposure in Normal and Transformed Mammary Cells. J. Cell. Biochem. 2015, 116, 1693–1711. [Google Scholar] [CrossRef]
- Desgrosellier, J.S.; Cheresh, D.A. Integrins in cancer: Biological implications and therapeutic opportunities. Nat. Rev. Cancer 2010, 10, 9–22. [Google Scholar] [CrossRef]
- Robinson, S.D.; Hodivala-Dilke, K.M. The role of beta3-integrins in tumor angiogenesis: Context is everything. Curr. Opin. Cell Biol. 2011, 23, 630–637. [Google Scholar] [CrossRef]
- Perry, M.C.; Demeule, M.; Regina, A.; Moumdjian, R.; Beliveau, R. Curcumin inhibits tumor growth and angiogenesis in glioblastoma xenografts. Mol. Nutr. Food Res. 2010, 54, 1192–1201. [Google Scholar] [CrossRef]
- Barsony, J.; Renyi, I.; McKoy, W. Subcellular distribution of normal and mutant vitamin D receptors in living cells. Studies with a novel fluorescent ligand. J. Biol. Chem. 1997, 272, 5774–5782. [Google Scholar] [CrossRef]
- Kim, Y.S.; MacDonald, P.N.; Dedhar, S.; Hruska, K.A. Association of 1 alpha,25-dihydroxyvitamin D3-occupied vitamin D receptors with cellular membrane acceptance sites. Endocrinology 1996, 137, 3649–3658. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, D.G.; Horsford, J.; Michalak, M.; White, J.H.; Hendy, G.N. Calreticulin inhibits vitamin D3 signal transduction. Nucleic Acids Res. 1995, 23, 3268–3274. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, R.; Fox, J. Biology and disease of mice. In Laboratory Animal Medicine; Fox, J., Cohen, B.J., Loew, F.M., Quimby, F.W., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1984; pp. 31–89. [Google Scholar]
- Esparza-Lopez, J.; Medina-Franco, H.; Escobar-Arriaga, E.; Leon-Rodriguez, E.; Zentella-Dehesa, A.; Ibarra-Sanchez, M.J. Doxorubicin induces atypical NF-kappaB activation through c-Abl kinase activity in breast cancer cells. J. Cancer Res. Clin. Oncol. 2013, 139, 1625–1635. [Google Scholar] [CrossRef] [PubMed]
- Esparza-Lopez, J.; Ramos-Elias, P.A.; Castro-Sanchez, A.; Rocha-Zavaleta, L.; Escobar-Arriaga, E.; Zentella-Dehesa, A.; Leon-Rodriguez, E.; Medina-Franco, H.; Ibarra-Sanchez Mde, J. Primary breast cancer cell culture yields intra-tumor heterogeneous subpopulations expressing exclusive patterns of receptor tyrosine kinases. BMC Cancer 2016, 16, 740. [Google Scholar] [CrossRef]
- Weidner, N.; Semple, J.P.; Welch, W.R.; Folkman, J. Tumor angiogenesis and metastasis—Correlation in invasive breast carcinoma. N. Engl. J. Med. 1991, 324, 1–8. [Google Scholar] [CrossRef]
- Grada, A.; Otero-Vinas, M.; Prieto-Castrillo, F.; Obagi, Z.; Falanga, V. Research Techniques Made Simple: Analysis of Collective Cell Migration Using the Wound Healing Assay. J. Investig. Dermatol. 2017, 137, e11–e16. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | MBCDF-T | EA.hy926 |
|---|---|---|
| Cal | 1.04 nM | NA |
| Cur | 7.96 μM | 8.42 μM |
| Rsv | 7.83 μM | 8.74 μM |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Quiroz, J.; García-Becerra, R.; Santos-Cuevas, C.; Ramírez-Nava, G.J.; Morales-Guadarrama, G.; Cárdenas-Ochoa, N.; Segovia-Mendoza, M.; Prado-Garcia, H.; Ordaz-Rosado, D.; Avila, E.; et al. Synergistic Antitumorigenic Activity of Calcitriol with Curcumin or Resveratrol is Mediated by Angiogenesis Inhibition in Triple Negative Breast Cancer Xenografts. Cancers 2019, 11, 1739. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11111739
García-Quiroz J, García-Becerra R, Santos-Cuevas C, Ramírez-Nava GJ, Morales-Guadarrama G, Cárdenas-Ochoa N, Segovia-Mendoza M, Prado-Garcia H, Ordaz-Rosado D, Avila E, et al. Synergistic Antitumorigenic Activity of Calcitriol with Curcumin or Resveratrol is Mediated by Angiogenesis Inhibition in Triple Negative Breast Cancer Xenografts. Cancers. 2019; 11(11):1739. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11111739
Chicago/Turabian StyleGarcía-Quiroz, Janice, Rocío García-Becerra, Clara Santos-Cuevas, Gerardo J. Ramírez-Nava, Gabriela Morales-Guadarrama, Nohemí Cárdenas-Ochoa, Mariana Segovia-Mendoza, Heriberto Prado-Garcia, David Ordaz-Rosado, Euclides Avila, and et al. 2019. "Synergistic Antitumorigenic Activity of Calcitriol with Curcumin or Resveratrol is Mediated by Angiogenesis Inhibition in Triple Negative Breast Cancer Xenografts" Cancers 11, no. 11: 1739. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11111739