POLQ Overexpression Is Associated with an Increased Somatic Mutation Load and PLK4 Overexpression in Lung Adenocarcinoma

 ,
,
Abstract
:1. Introduction
2. Results
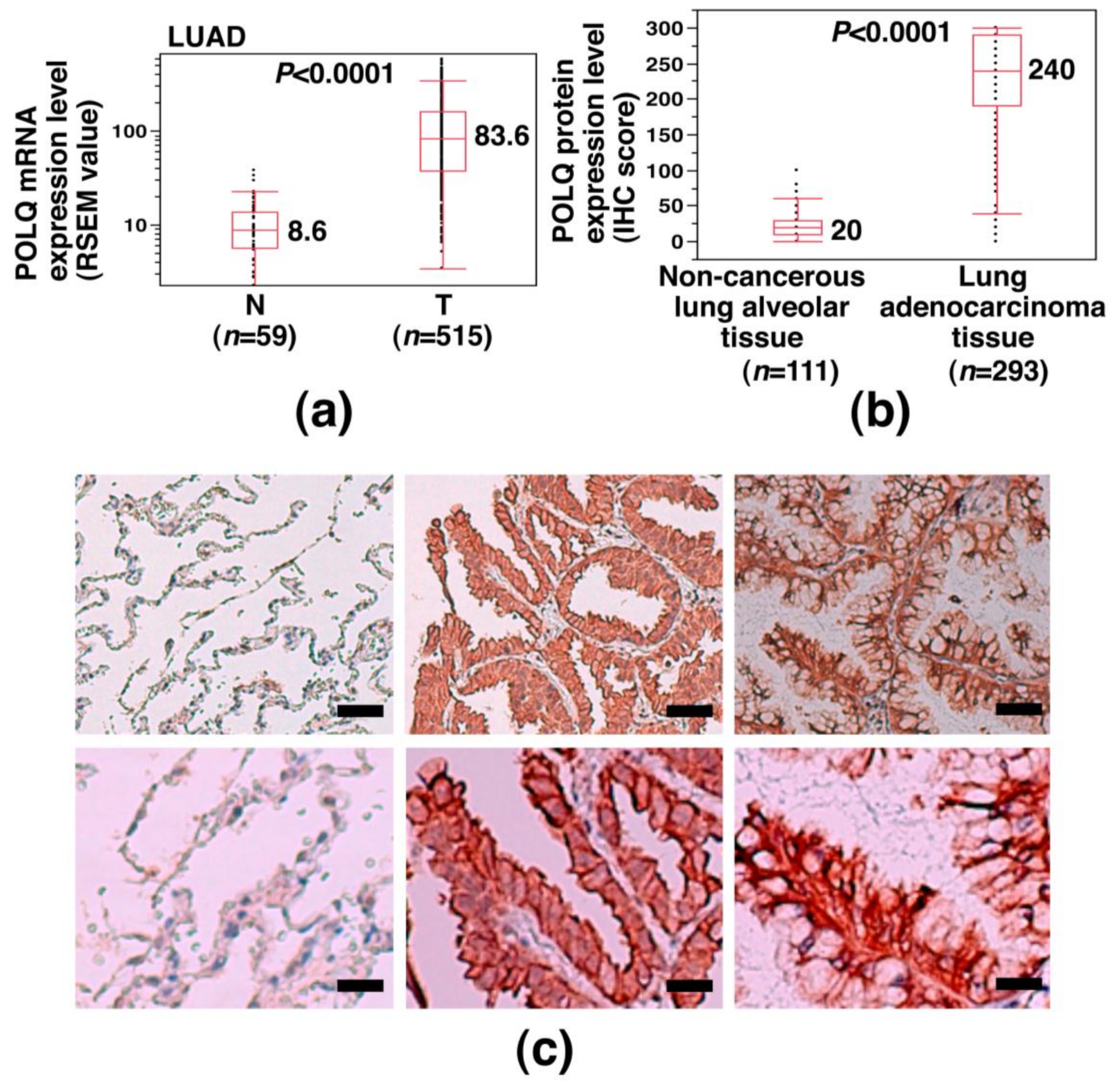
2.1. Overexpression of POLQ mRNA and Protein in LAC
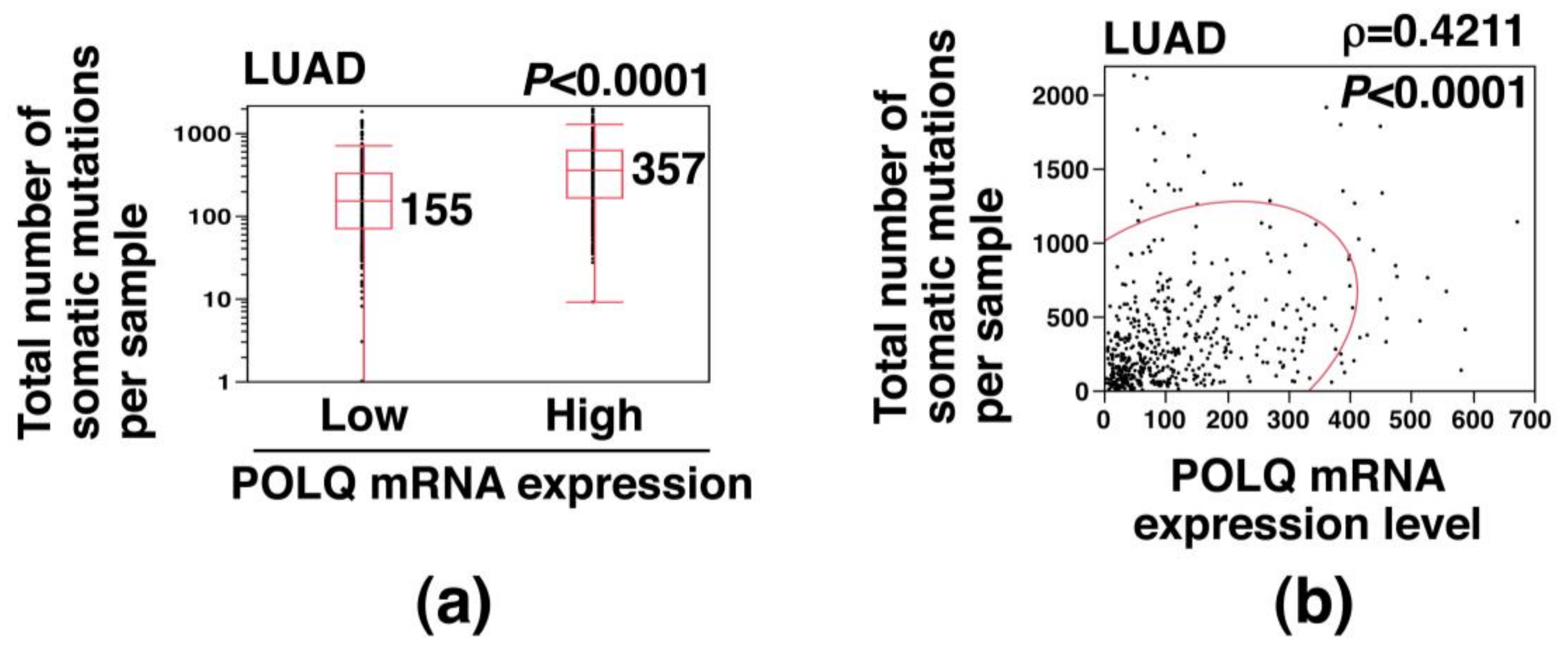
2.2. Association of Increased POLQ Expression with Increased Somatic Mutation Load in LAC
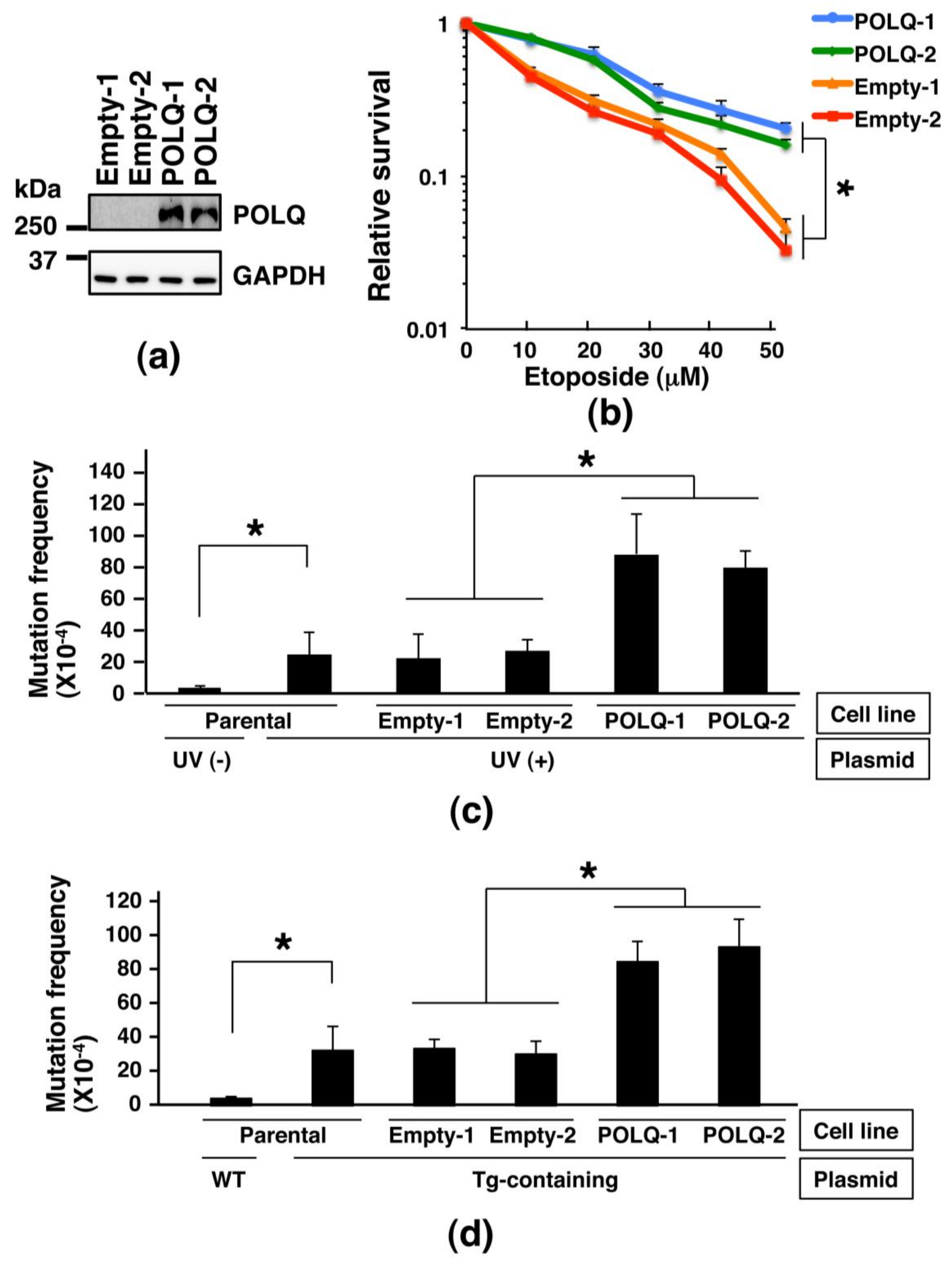
2.3. Comparison of the Sensitivity to DNA-Damaging Agent and Frequency of Mutations among Lung Cancer Cells Showing Different Expression Levels of POLQ
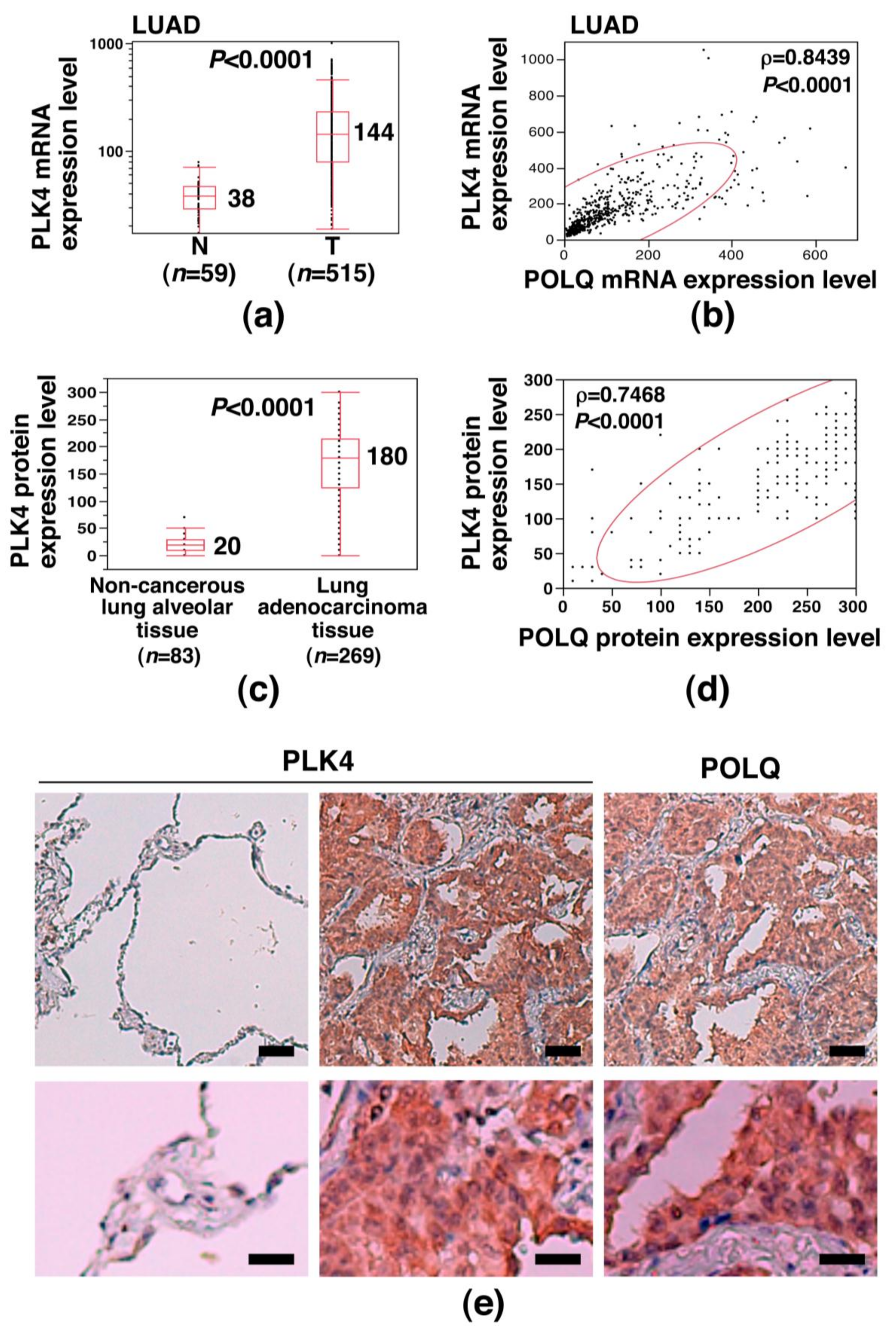
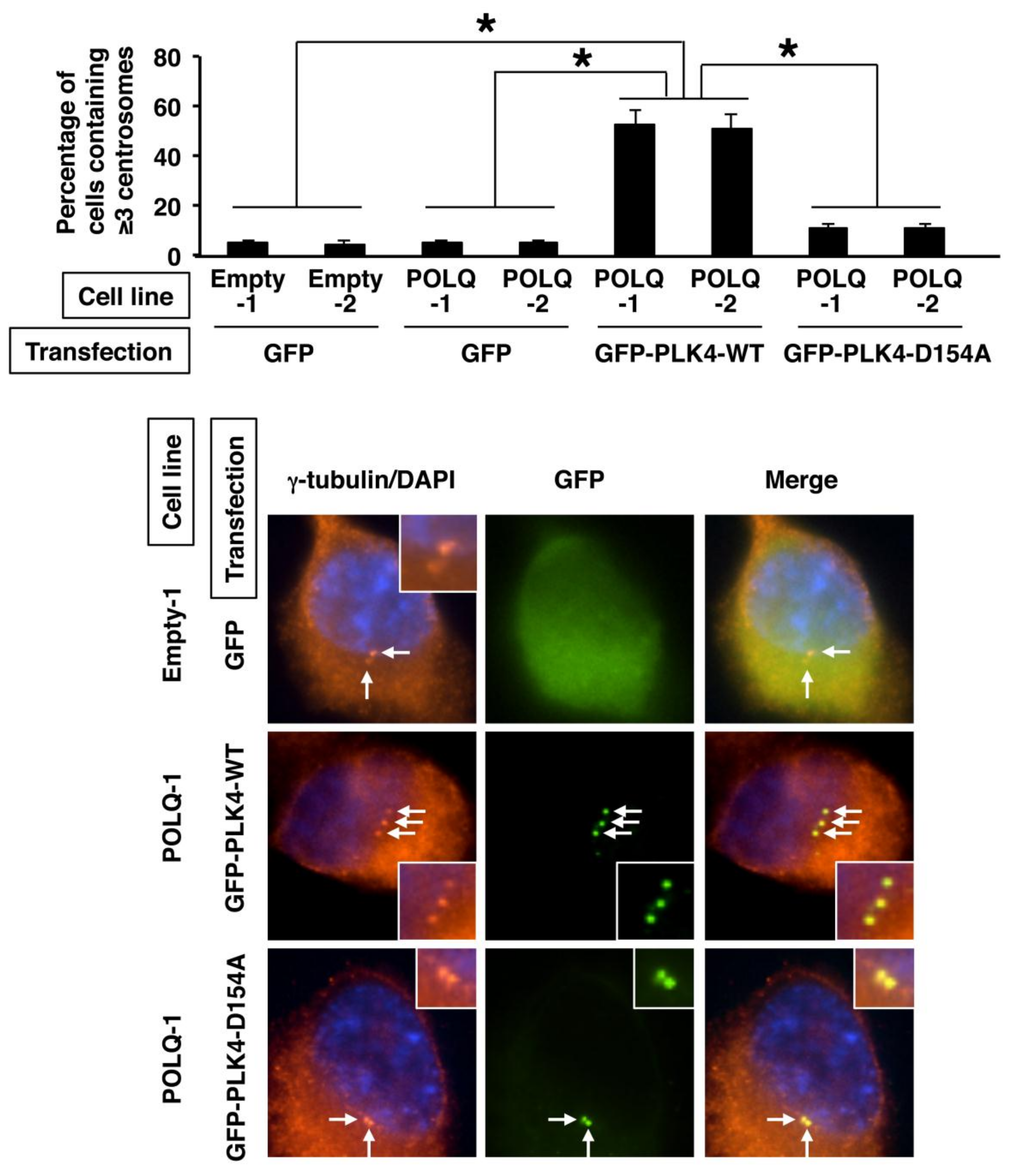
2.4. Concurrent POLQ and PLK4 Overexpression in LAC and Induction of Centrosome Amplification
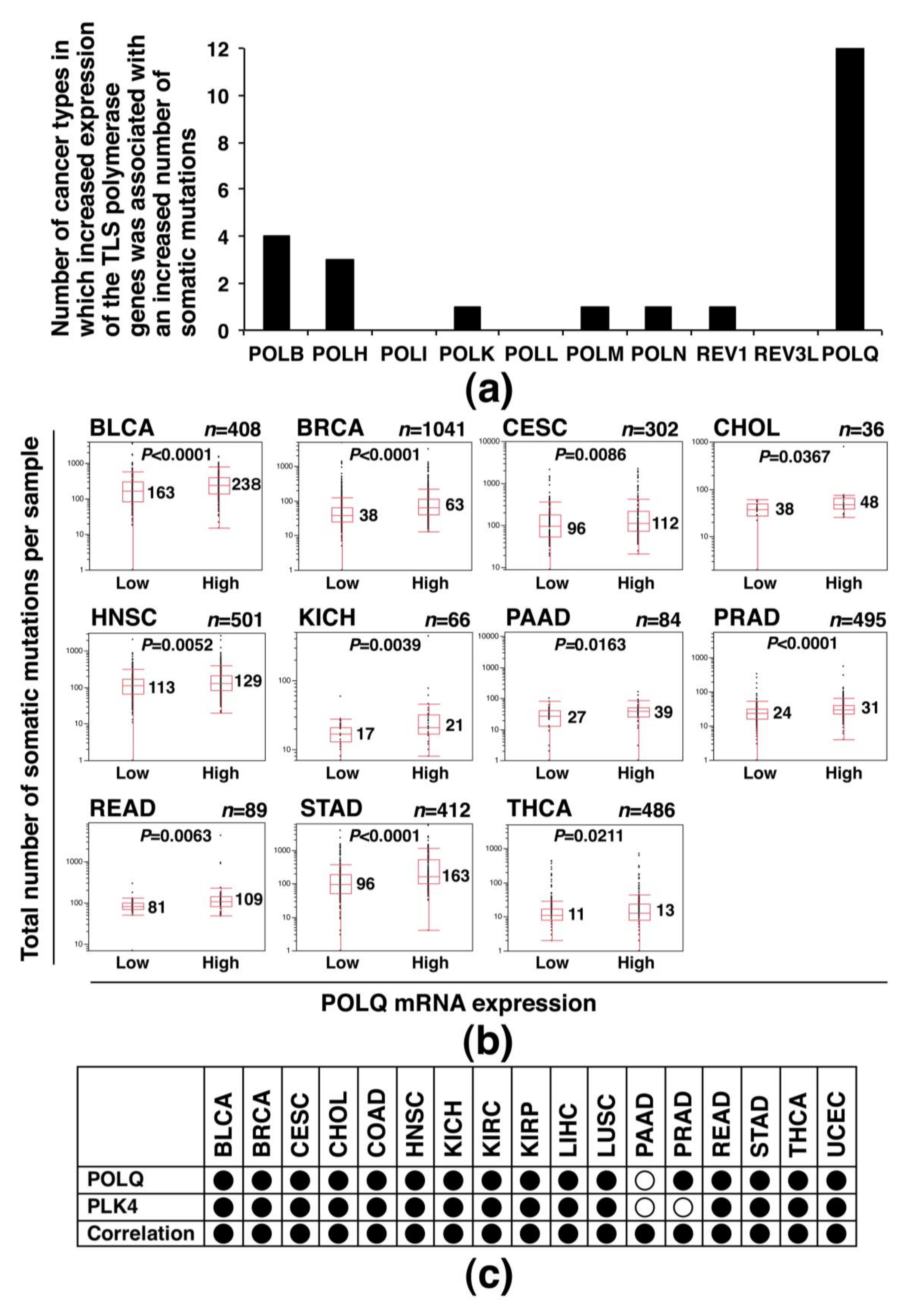
2.5. Association of Increased POLQ Expression with an Increased Somatic Mutation Load and PLK4 Overexpression in Diverse Human Cancers
3. Discussion
4. Materials and Methods
4.1. Collection of Publicly Available Data
4.2. Primary LAC Specimens
4.3. IHC Analysis
4.4. Cell Cultures
4.5. Establishment of Stable Inducible Cell Lines
4.6. Western Blot Analysis
4.7. Clonogenic Survival Assay
4.8. Preparation of Shuttle Vector Plasmids
4.9. SupF Forward Mutation Assay
4.10. Indirect Immunofluorescence Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Galateau-Salle, F.; Churg, A.; Roggli, V.; Travis, W.D. The 2015 World Health Organization Classification of Tumors of the Pleura: Advances since the 2004 Classification. J. Thorac. Oncol. 2016, 11, 142–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inamura, K. Update on immunohistochemistry for the diagnosis of lung cancer. Cancers 2018, 10, 72. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Research Network. Comprehensive molecular profiling of lung adenocarcinoma. Nature 2014, 511, 543–550. [Google Scholar] [CrossRef]
- Alexandrov, L.B.; Ju, Y.S.; Haase, K.; Van Loo, P.; Martincorena, I.; Nik-Zainal, S.; Totoki, Y.; Fujimoto, A.; Nakagawa, H.; Shibata, T.; et al. Mutational signatures associated with tobacco smoking in human cancer. Science 2016, 354, 618–622. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Hua, X.; Zhu, B.; Ravichandran, S.; Wang, M.; Nguyen, C.; Brodie, S.A.; Palleschi, A.; Alloisio, M.; Pariscenti, G.; et al. Somatic genomics and clinical features of lung adenocarcinoma: A retrospective study. PLoS Med. 2016, 13, e1002162. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wei, Y.; Zhang, R.; Su, L.; Gogarten, S.M.; Liu, G.; Brennan, P.; Field, J.K.; McKay, J.D.; Lissowska, J.; et al. Multi-omics analysis reveals a HIF network and hub gene EPAS1 associated with lung adenocarcinoma. EBioMedicine 2018, 32, 93–101. [Google Scholar] [CrossRef]
- Kundu, S.T.; Grzeskowiak, C.L.; Fradette, J.J.; Gibson, L.A.; Rodriguez, L.B.; Creighton, C.J.; Scott, K.L.; Gibbons, D.L. TMEM106B drives lung cancer metastasis by inducing TFEB-dependent lysosome synthesis and secretion of cathepsins. Nat. Commun. 2018, 9, 2731. [Google Scholar] [CrossRef]
- Wood, R.D.; Doublié, S. DNA polymerase 0 (POLQ), double-strand break repair, and cancer. DNA Repair (Amst.) 2016, 44, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Sallmyr, A.; Tomkinson, A.E. Repair of DNA double-strand breaks by mammalian alternative end-joining pathways. J. Biol. Chem. 2018, 293, 10536–10546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seki, M.; Marini, F.; Wood, R.D. POLQ (Pol theta), a DNA polymerase and DNA-dependent ATPase in human cells. Nucleic Acids Res. 2003, 31, 6117–6126. [Google Scholar] [CrossRef] [PubMed]
- Seki, M.; Masutani, C.; Yang, L.W.; Schuffert, A.; Iwai, S.; Bahar, I.; Wood, R.D. High-efficiency bypass of DNA damage by human DNA polymerase Q. EMBO J. 2004, 23, 4484–4494. [Google Scholar] [CrossRef] [Green Version]
- Arana, M.E.; Seki, M.; Wood, R.D.; Rogozin, I.B.; Kunkel, T.A. Low-fidelity DNA synthesis by human DNA polymerase theta. Nucleic Acids Res. 2008, 36, 3847–3856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roerink, S.F.; van Schendel, R.; Tijsterman, M. Polymerase theta-mediated end joining of replication-associated DNA breaks in C. elegans. Genome Res. 2014, 24, 954–962. [Google Scholar] [CrossRef]
- Yousefzadeh, M.J.; Wyatt, D.W.; Takata, K.; Mu, Y.; Hensley, S.C.; Tomida, J.; Bylund, G.O.; Doublié, S.; Johansson, E.; Ramsden, D.A.; et al. Mechanism of suppression of chromosomal instability by DNA polymerase POLQ. PLoS Genet. 2014, 10, e1004654. [Google Scholar] [CrossRef]
- Koole, W.; van Schendel, R.; Karambelas, A.E.; van Heteren, J.T.; Okihara, K.L.; Tijsterman, M. A Polymerase Theta-dependent repair pathway suppresses extensive genomic instability at endogenous G4 DNA sites. Nat. Commun. 2014, 5, 3216. [Google Scholar] [CrossRef] [Green Version]
- Ceccaldi, R.; Liu, J.C.; Amunugama, R.; Hajdu, I.; Primack, B.; Petalcorin, M.I.; O’Connor, K.W.; Konstantinopoulos, P.A.; Elledge, S.J.; Boulton, S.J.; et al. Homologous-recombination-deficient tumours are dependent on Polθ-mediated repair. Nature 2015, 518, 258–262. [Google Scholar] [CrossRef]
- Mateos-Gomez, P.A.; Gong, F.; Nair, N.; Miller, K.M.; Lazzerini-Denchi, E.; Sfeir, A. Mammalian polymerase θ promotes alternative NHEJ and suppresses recombination. Nature 2015, 518, 254–257. [Google Scholar] [CrossRef]
- Ranjha, L.; Howard, S.M.; Cejka, P. Main steps in DNA double-strand break repair: An introduction to homologous recombination and related processes. Chromosoma 2018, 127, 187–214. [Google Scholar] [CrossRef]
- Kawamura, K.; Bahar, R.; Seimiya, M.; Chiyo, M.; Wada, A.; Okada, S.; Hatano, M.; Tokuhisa, T.; Kimura, H.; Watanabe, S.; et al. DNA polymerase theta is preferentially expressed in lymphoid tissues and upregulated in human cancers. Int. J. Cancer 2004, 109, 9–16. [Google Scholar] [CrossRef]
- Lemée, F.; Bergoglio, V.; Fernandez-Vidal, A.; Machado-Silva, A.; Pillaire, M.J.; Bieth, A.; Gentil, C.; Baker, L.; Martin, A.L.; Leduc, C.; et al. DNA polymerase theta up-regulation is associated with poor survival in breast cancer, perturbs DNA replication, and promotes genetic instability. Proc. Natl. Acad. Sci. USA 2010, 107, 13390–13395. [Google Scholar] [CrossRef]
- Higgins, G.S.; Harris, A.L.; Prevo, R.; Helleday, T.; McKenna, W.G.; Buffa, F.M. Overexpression of POLQ confers a poor prognosis in early breast cancer patients. Oncotarget 2010, 1, 175–184. [Google Scholar]
- Pillaire, M.J.; Selves, J.; Gordien, K.; Gourraud, P.A.; Gentil, C.; Danjoux, M.; Do, C.; Negre, V.; Bieth, A.; Guimbaud, R.; et al. A ‘DNA replication’ signature of progression and negative outcome in colorectal cancer. Oncogene 2010, 29, 876–887. [Google Scholar] [CrossRef]
- Allera-Moreau, C.; Rouquette, I.; Lepage, B.; Oumouhou, N.; Walschaerts, M.; Leconte, E.; Schilling, V.; Gordien, K.; Brouchet, L.; Delisle, M.B.; et al. DNA replication stress response involving PLK1, CDC6, POLQ, RAD51 and CLASPIN upregulation prognoses the outcome of early/mid-stage non-small cell lung cancer patients. Oncogenesis 2012, 1, e30. [Google Scholar] [CrossRef]
- Lessa, R.C.; Campos, A.H.; Freitas, C.E.; Silva, F.R.; Kowalski, L.P.; Carvalho, A.L.; Vettore, A.L. Identification of upregulated genes in oral squamous cell carcinomas. Head Neck 2013, 35, 1475–1481. [Google Scholar] [CrossRef]
- Seki, M.; Wood, R.D. DNA polymerase theta (POLQ) can extend from mismatches and from bases opposite a (6–4) photoproduct. DNA Repair (Amst.) 2008, 7, 119–127. [Google Scholar] [CrossRef]
- Habedanck, R.; Stierhof, Y.D.; Wilkinson, C.J.; Nigg, E.A. The Polo kinase Plk4 functions in centriole duplication. Nat. Cell Biol. 2005, 7, 1140–1146. [Google Scholar] [CrossRef]
- Arquint, C.; Nigg, E.A. The PLK4-STIL-SAS-6 module at the core of centriole duplication. Biochem. Soc. Trans. 2016, 44, 1253–1263. [Google Scholar] [CrossRef]
- Joshi, H.C. Microtubule organizing centers and gamma-tubulin. Curr. Opin. Cell Biol. 1994, 6, 54–62. [Google Scholar] [CrossRef]
- Sale, J.E. Translesion DNA synthesis and mutagenesis in eukaryotes. Cold Spring Harb. Perspect. Biol. 2013, 5, a012708. [Google Scholar] [CrossRef]
- Roberts, S.A.; Lawrence, M.S.; Klimczak, L.J.; Grimm, S.A.; Fargo, D.; Stojanov, P.; Kiezun, A.; Kryukov, G.V.; Carter, S.L.; Saksena, G.; et al. An APOBEC cytidine deaminase mutagenesis pattern is widespread in human cancers. Nat. Genet. 2013, 45, 970–976. [Google Scholar] [CrossRef]
- Burns, M.B.; Temiz, N.A.; Harris, R.S. Evidence for APOBEC3B mutagenesis in multiple human cancers. Nat. Genet. 2013, 45, 977–983. [Google Scholar] [CrossRef] [Green Version]
- Alexandrov, L.B.; Nik-Zainal, S.; Wedge, D.C.; Aparicio, S.A.; Behjati, S.; Biankin, A.V.; Bignell, G.R.; Bolli, N.; Borg, A.; Børresen-Dale, A.L.; et al. Signatures of mutational processes in human cancer. Nature 2013, 500, 415–421. [Google Scholar] [CrossRef] [Green Version]
- Shinmura, K.; Kato, H.; Kawanishi, Y.; Igarashi, H.; Goto, M.; Tao, H.; Inoue, Y.; Nakamura, S.; Misawa, K.; Mineta, H.; et al. Abnormal Expressions of DNA Glycosylase Genes NEIL1, NEIL2, and NEIL3 Are associated with somatic mutation loads in human cancer. Oxid. Med. Cell. Longev. 2016, 2016, 1546392. [Google Scholar] [CrossRef]
- Shinmura, K.; Kato, H.; Kawanishi, Y.; Yoshimura, K.; Igarashi, H.; Goto, M.; Tao, H.; Inoue, Y.; Sugiyama, T.; Furuse, H.; et al. Reduced expression of the DNA glycosylase gene MUTYH is associated with an increased number of somatic mutations via a reduction in the DNA repair capacity in prostate adenocarcinoma. Mol. Carcinog. 2017, 56, 781–788. [Google Scholar] [CrossRef]
- Braun, D.A.; Burke, K.P.; Van Allen, E.M. Genomic Approaches to Understanding Response and Resistance to Immunotherapy. Clin. Cancer Res. 2016, 22, 5642–5650. [Google Scholar] [CrossRef] [Green Version]
- Kawakami, M.; Mustachio, L.M.; Zheng, L.; Chen, Y.; Rodriguez-Canales, J.; Mino, B.; Kurie, J.M.; Roszik, J.; Villalobos, P.A.; Thu, K.L.; et al. Polo-like kinase 4 inhibition produces polyploidy and apoptotic death of lung cancers. Proc. Natl. Acad. Sci. USA 2018, 115, 1913–1918. [Google Scholar] [CrossRef] [Green Version]
- Koutsami, M.K.; Tsantoulis, P.K.; Kouloukoussa, M.; Apostolopoulou, K.; Pateras, I.S.; Spartinou, Z.; Drougou, A.; Evangelou, K.; Kittas, C.; Bartkova, J.; et al. Centrosome abnormalities are frequently observed in non-small-cell lung cancer and are associated with aneuploidy and cyclin E overexpression. J. Pathol. 2006, 209, 512–521. [Google Scholar] [CrossRef]
- Shinmura, K.; Iwaizumi, M.; Igarashi, H.; Nagura, K.; Yamada, H.; Suzuki, M.; Fukasawa, K.; Sugimura, H. Induction of centrosome amplification and chromosome instability in p53-deficient lung cancer cells exposed to benzo[a]pyrene diol epoxide (B[a]PDE). J. Pathol. 2008, 216, 365–374. [Google Scholar] [CrossRef]
- Ganem, N.J.; Godinho, S.A.; Pellman, D. A mechanism linking extra centrosomes to chromosomal instability. Nature 2009, 460, 278–282. [Google Scholar] [CrossRef] [Green Version]
- Cosenza, M.R.; Krämer, A. Centrosome amplification, chromosomal instability and cancer: Mechanistic, clinical and therapeutic issues. Chromosome Res. 2016, 24, 105–126. [Google Scholar] [CrossRef] [PubMed]
- Nigg, E.A.; Holland, A.J. Once and only once: Mechanisms of centriole duplication and their deregulation in disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 297–312. [Google Scholar] [CrossRef] [PubMed]
- Godinho, S.A.; Picone, R.; Burute, M.; Dagher, R.; Su, Y.; Leung, C.T.; Polyak, K.; Brugge, J.S.; Théry, M.; Pellman, D. Oncogene-like induction of cellular invasion from centrosome amplification. Nature 2014, 510, 167–171. [Google Scholar] [CrossRef] [Green Version]
- Kleylein-Sohn, J.; Westendorf, J.; Le Clech, M.; Habedanck, R.; Stierhof, Y.D.; Nigg, E.A. Plk4-induced centriole biogenesis in human cells. Dev. Cell 2007, 13, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Shinmura, K.; Kurabe, N.; Goto, M.; Yamada, H.; Natsume, H.; Konno, H.; Sugimura, H. PLK4 overexpression and its effect on centrosome regulation and chromosome stability in human gastric cancer. Mol. Biol. Rep. 2014, 41, 6635–6644. [Google Scholar] [CrossRef]
- Ling, H.; Hanashiro, K.; Luong, T.H.; Benavides, L.; Fukasawa, K. Functional relationship among PLK2, PLK4 and ROCK2 to induce centrosome amplification. Cell Cycle 2015, 14, 544–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirsch, F.R.; Suda, K.; Wiens, J.; Bunn, P.A., Jr. New and emerging targeted treatments in advanced non-small-cell lung cancer. Lancet 2016, 388, 1012–1024. [Google Scholar] [CrossRef]
- Wang, Y.; Blandino, G.; Givol, D. Induced p21waf expression in H1299 cell line promotes cell senescence and protects against cytotoxic effect of radiation and doxorubicin. Oncogene 1999, 18, 2643–2649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boggaram, V. Thyroid transcription factor-1 (TTF-1/Nkx2.1/TITF1) gene regulation in the lung. Clin. Sci. (Lond.) 2009, 116, 27–35. [Google Scholar] [CrossRef]
- Yatabe, Y.; Mitsudomi, T.; Takahashi, T. TTF-1 expression in pulmonary adenocarcinomas. Am. J. Surg. Pathol. 2002, 26, 767–773. [Google Scholar] [CrossRef]
- Qian, H.H.; Xu, T.S.; Cai, X.Q.; Ji, T.L.; Guo, H.X. Prognostic value of TTF-1 expression in patients with non-small cell lung cancer: A meta-analysis. Clin. Chim. Acta 2015, 451, 208–214. [Google Scholar] [CrossRef]
- cBioPortal. Available online: https://www.cbioportal.org/ (accessed on 8 May 2019).
- Khozin, S.; Blumenthal, G.M.; Jiang, X.; He, K.; Boyd, K.; Murgo, A.; Justice, R.; Keegan, P.; Pazdur, R.U.S. Food and Drug Administration approval summary: Erlotinib for the first-line treatment of metastatic non-small cell lung cancer with epidermal growth factor receptor exon 19 deletions or exon 21 (L858R) substitution mutations. Oncologist 2014, 19, 774–779. [Google Scholar] [CrossRef]
- Piva, S.; Ganzinelli, M.; Garassino, M.C.; Caiola, E.; Farina, G.; Broggini, M.; Marabese, M. Across the universe of K-RAS mutations in non-small-cell-lung cancer. Curr. Pharm. Des. 2014, 20, 3933–3943. [Google Scholar] [CrossRef]
- Nguyen-Ngoc, T.; Bouchaab, H.; Adjei, A.A.; Peters, S. BRAF Alterations as therapeutic targets in non-small-cell lung cancer. J. Thorac. Oncol. 2015, 10, 1396–1403. [Google Scholar] [CrossRef]
- Matsuda, T.; Yagi, T.; Kawanishi, M.; Matsui, S.; Takebe, H. Molecular analysis of mutations induced by 2-chloroacetaldehyde, the ultimate carcinogenic form of vinyl chloride, in human cells using shuttle vectors. Carcinogenesis 1995, 16, 2389–2394. [Google Scholar] [CrossRef]
- Kawanishi, M.; Matsuda, T.; Sasaki, G.; Yagi, T.; Matsui, S.; Takebe, H. A spectrum of mutations induced by crotonaldehyde in shuttle vector plasmids propagated in human cells. Carcinogenesis 1998, 19, 69–72. [Google Scholar] [CrossRef]
- Shinmura, K.; Goto, M.; Suzuki, M.; Tao, H.; Yamada, H.; Igarashi, H.; Matsuura, S.; Maeda, M.; Konno, H.; Matsuda, T.; et al. Reduced expression of MUTYH with suppressive activity against mutations caused by 8-hydroxyguanine is a novel predictor of a poor prognosis in human gastric cancer. J. Pathol. 2011, 225, 414–423. [Google Scholar] [CrossRef]
- Shinmura, K.; Kato, H.; Kawanishi, Y.; Goto, M.; Tao, H.; Yoshimura, K.; Nakamura, S.; Misawa, K.; Sugimura, H. Defective repair capacity of variant proteins of the DNA glycosylase NTHL1 for 5-hydroxyuracil, an oxidation product of cytosine. Free Radic. Biol. Med. 2019, 131, 264–273. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | No. of Cases | POLQ Protein Expression Level § | p-Value † | |
|---|---|---|---|---|
| Low (n = 56) | High (n = 237) | |||
| Age (y) | ||||
| <60 | 82 | 19 (23.2%) | 63 (76.8%) | 0.3206 |
| ≥60 | 211 | 37 (17.5%) | 174 (82.5%) | |
| Sex | ||||
| Female | 123 | 23 (18.7%) | 100 (81.3%) | 1.0000 |
| Male | 170 | 33 (19.4%) | 137 (80.6%) | |
| Smoking | ||||
| Non-smoker | 99 | 21 (21.2%) | 78 (78.8%) | 0.1510 |
| Smoker | 128 | 17 (13.3%) | 111 (86.7%) | |
| pT | ||||
| pT1/pT2 | 258 | 51 (19.8%) | 207 (80.2%) | 0.6463 |
| pT3/pT4 | 35 | 5 (14.3%) | 30 (85.7%) | |
| pN | ||||
| pN0 | 218 | 50 (22.9%) | 168 (77.1%) | 0.0012 |
| pN1-pN3 | 69 | 4 (5.8%) | 65 (94.2%) | |
| TNM Stage | ||||
| I/II | 240 | 54 (22.5%) | 186 (77.5%) | 0.0008 |
| III/IV | 53 | 2 (3.8%) | 51 (96.2%) | |
| Factor | No. of Cases | POLQ mRNA Expression Level | p-Value † | |
|---|---|---|---|---|
| Low (n = 49) | High (n = 181) | |||
| EGFR | ||||
| WT | 212 | 40 (18.9%) | 172 (81.1%) | 0.0047 |
| Mutation | 18 | 9 (50.0%) | 9 (50.0%) | |
| KRAS | ||||
| WT | 156 | 31 (19.9%) | 125 (80.1%) | 0.4915 |
| Mutation | 74 | 18 (24.3%) | 56 (75.7%) | |
| BRAF | ||||
| WT | 225 | 48 (21.3%) | 177 (78.7%) | 1.0000 |
| Mutation | 5 | 1 (20.0%) | 4 (80.0%) | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shinmura, K.; Kato, H.; Kawanishi, Y.; Yoshimura, K.; Tsuchiya, K.; Takahara, Y.; Hosokawa, S.; Kawase, A.; Funai, K.; Sugimura, H. POLQ Overexpression Is Associated with an Increased Somatic Mutation Load and PLK4 Overexpression in Lung Adenocarcinoma. Cancers 2019, 11, 722. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11050722
Shinmura K, Kato H, Kawanishi Y, Yoshimura K, Tsuchiya K, Takahara Y, Hosokawa S, Kawase A, Funai K, Sugimura H. POLQ Overexpression Is Associated with an Increased Somatic Mutation Load and PLK4 Overexpression in Lung Adenocarcinoma. Cancers. 2019; 11(5):722. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11050722
Chicago/Turabian StyleShinmura, Kazuya, Hisami Kato, Yuichi Kawanishi, Katsuhiro Yoshimura, Kazuo Tsuchiya, Yoshiyuki Takahara, Seiji Hosokawa, Akikazu Kawase, Kazuhito Funai, and Haruhiko Sugimura. 2019. "POLQ Overexpression Is Associated with an Increased Somatic Mutation Load and PLK4 Overexpression in Lung Adenocarcinoma" Cancers 11, no. 5: 722. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11050722