Curcumin Treatment Identifies Therapeutic Targets within Biomarkers of Liver Colonization by Highly Invasive Mesothelioma Cells—Potential Links with Sarcomas

 ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
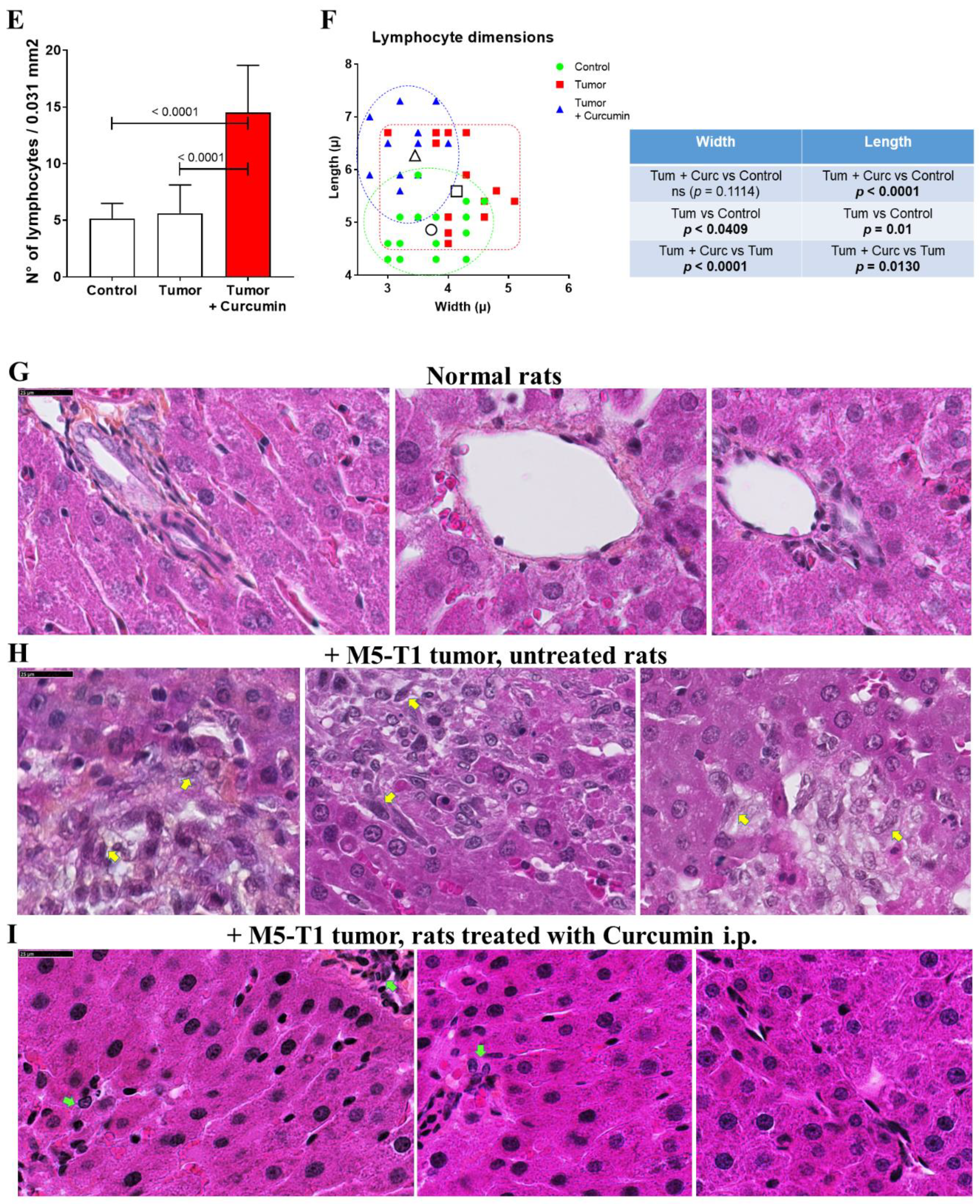
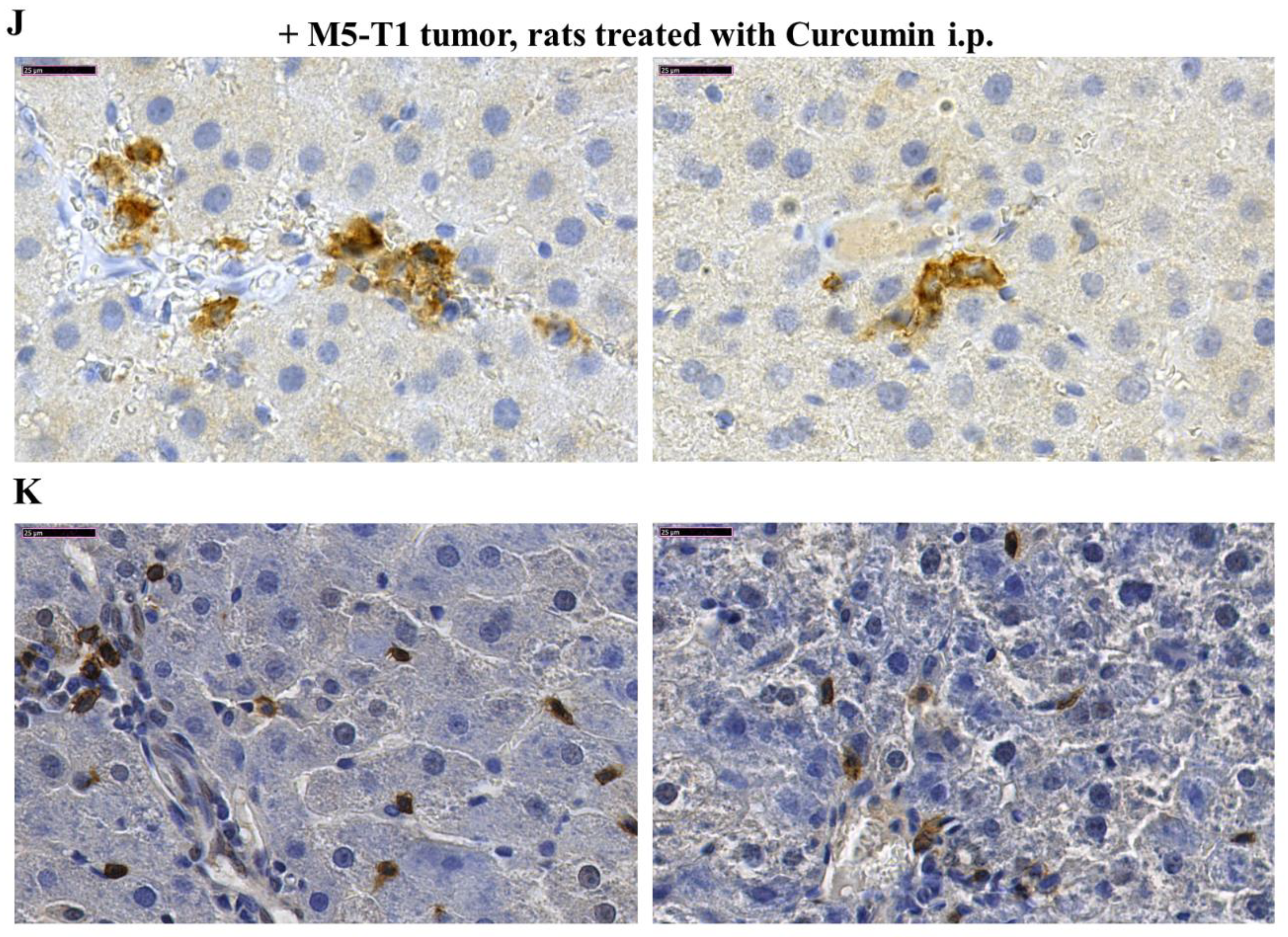
2.1. Histological Characterization of Lymphocyte Changes in the Liver of Curcumin-Treated Rats
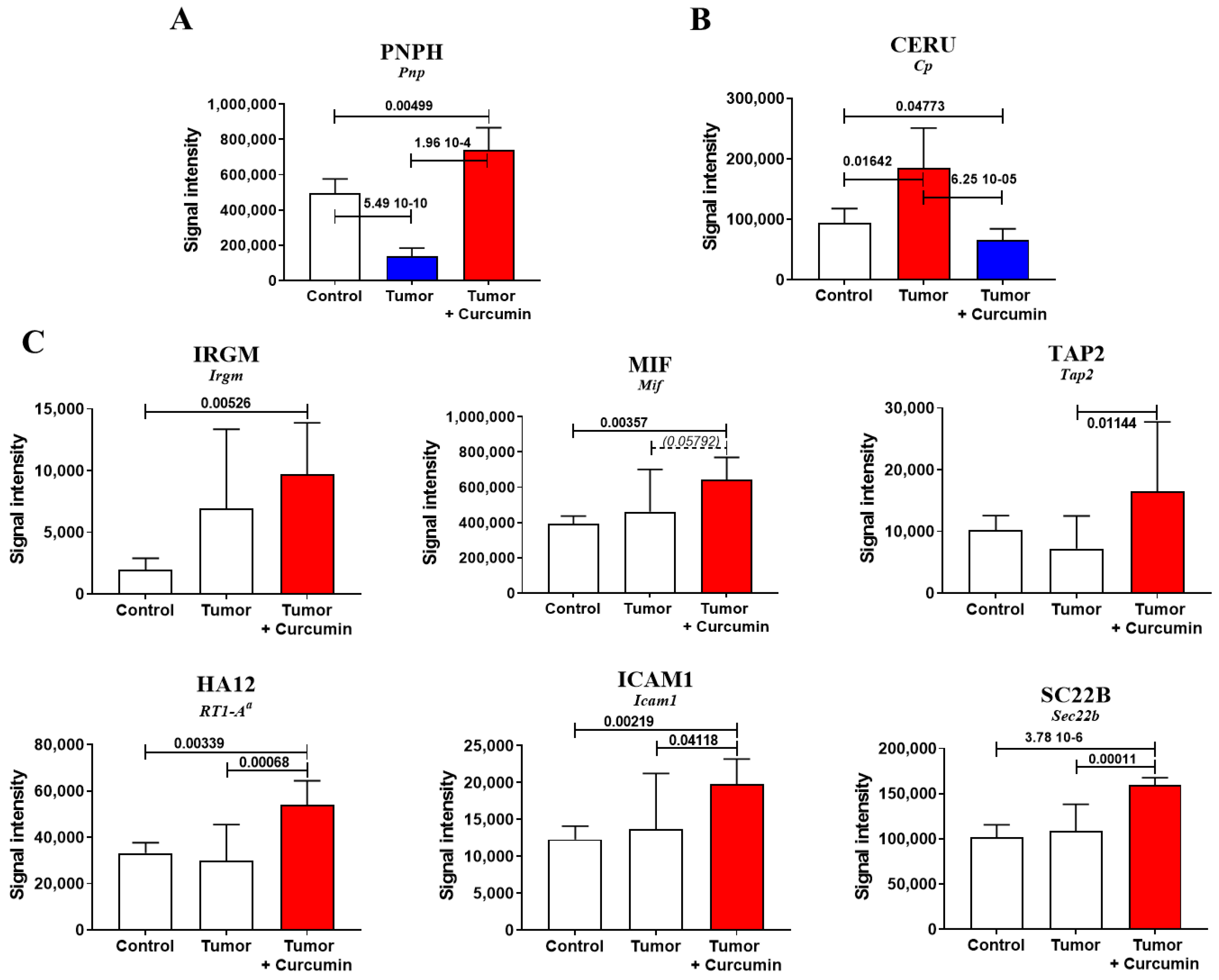
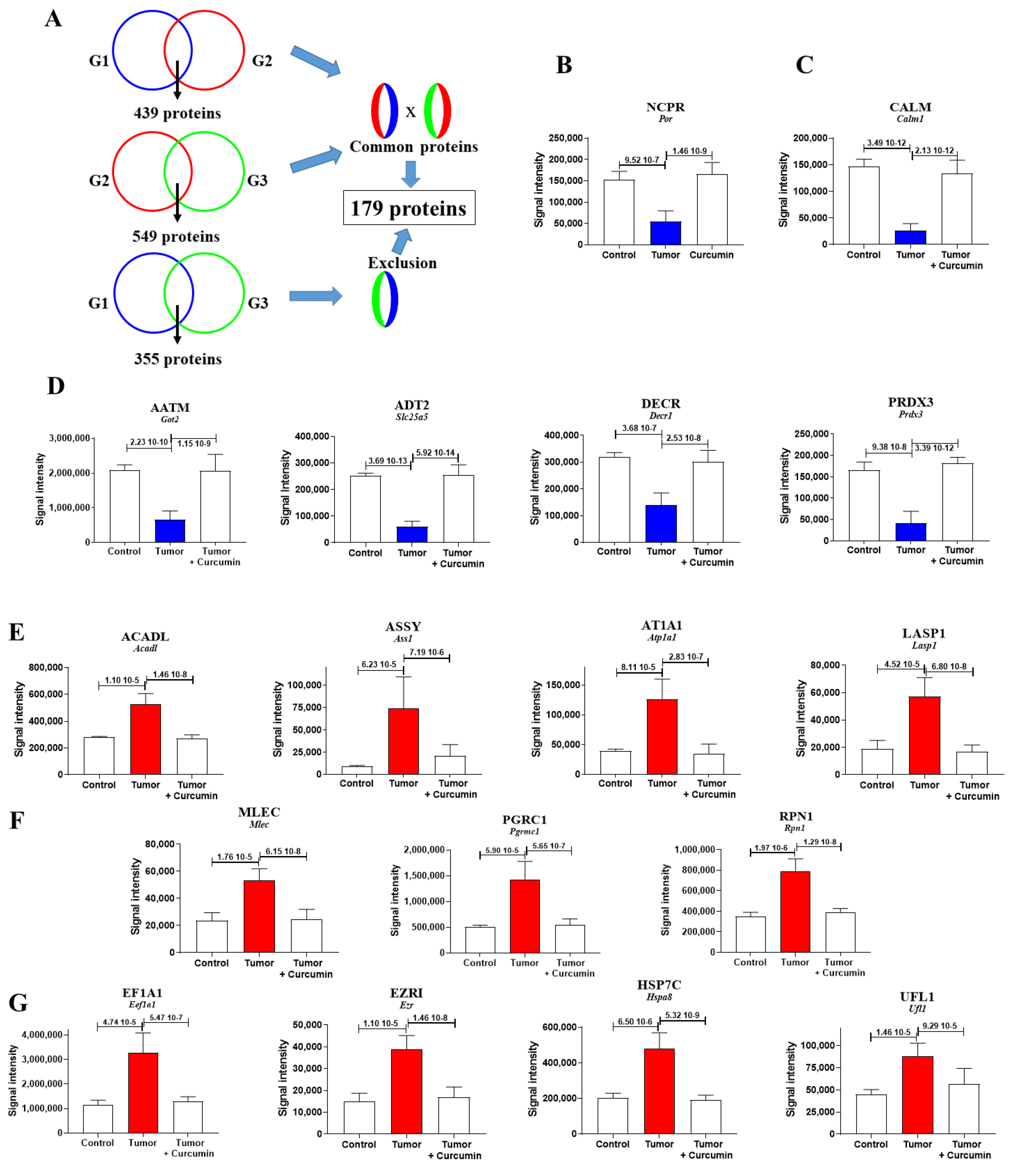
2.2. Biomarkers Involved in the Immune Response Directed against M5-T1 Cells
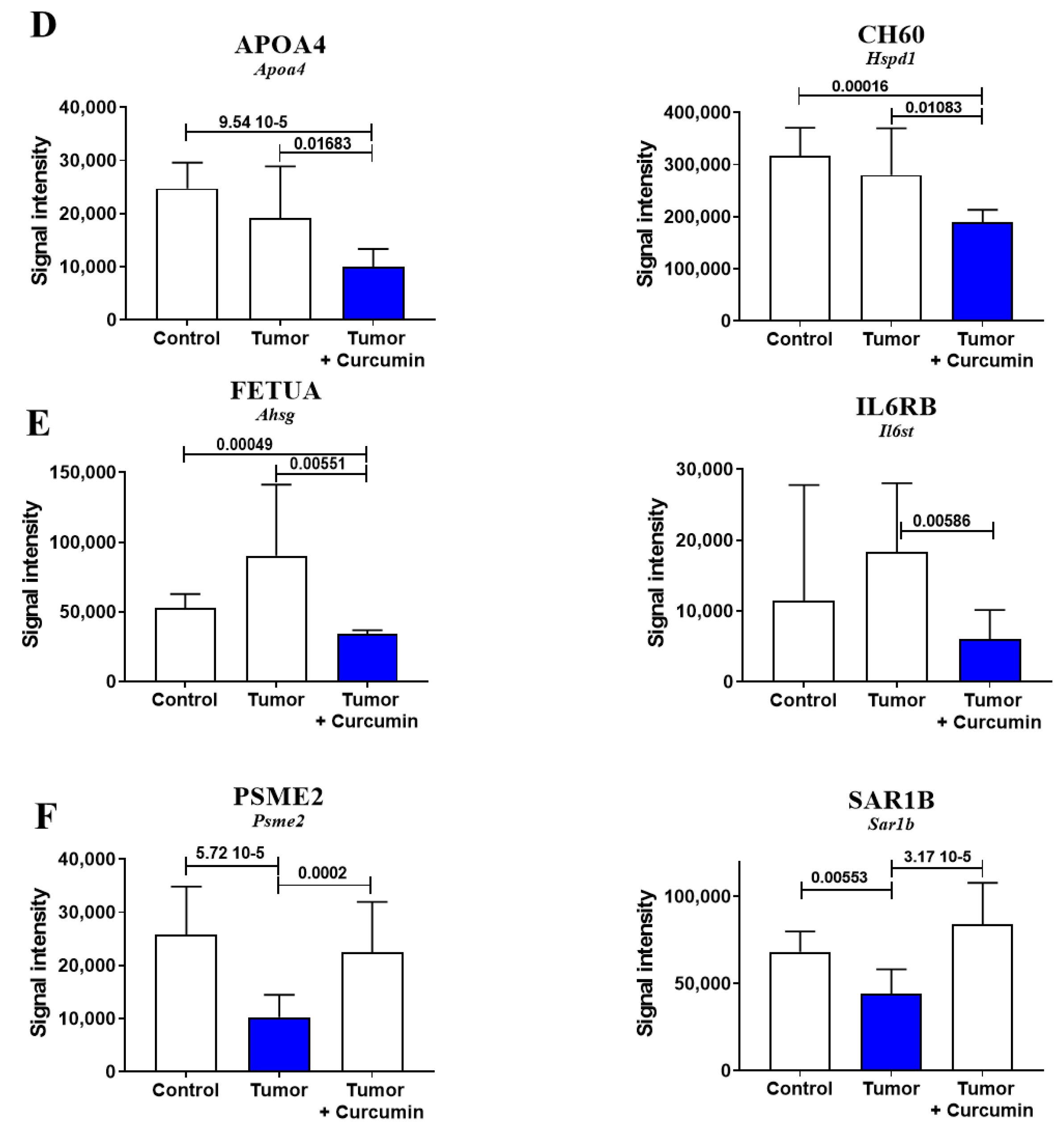
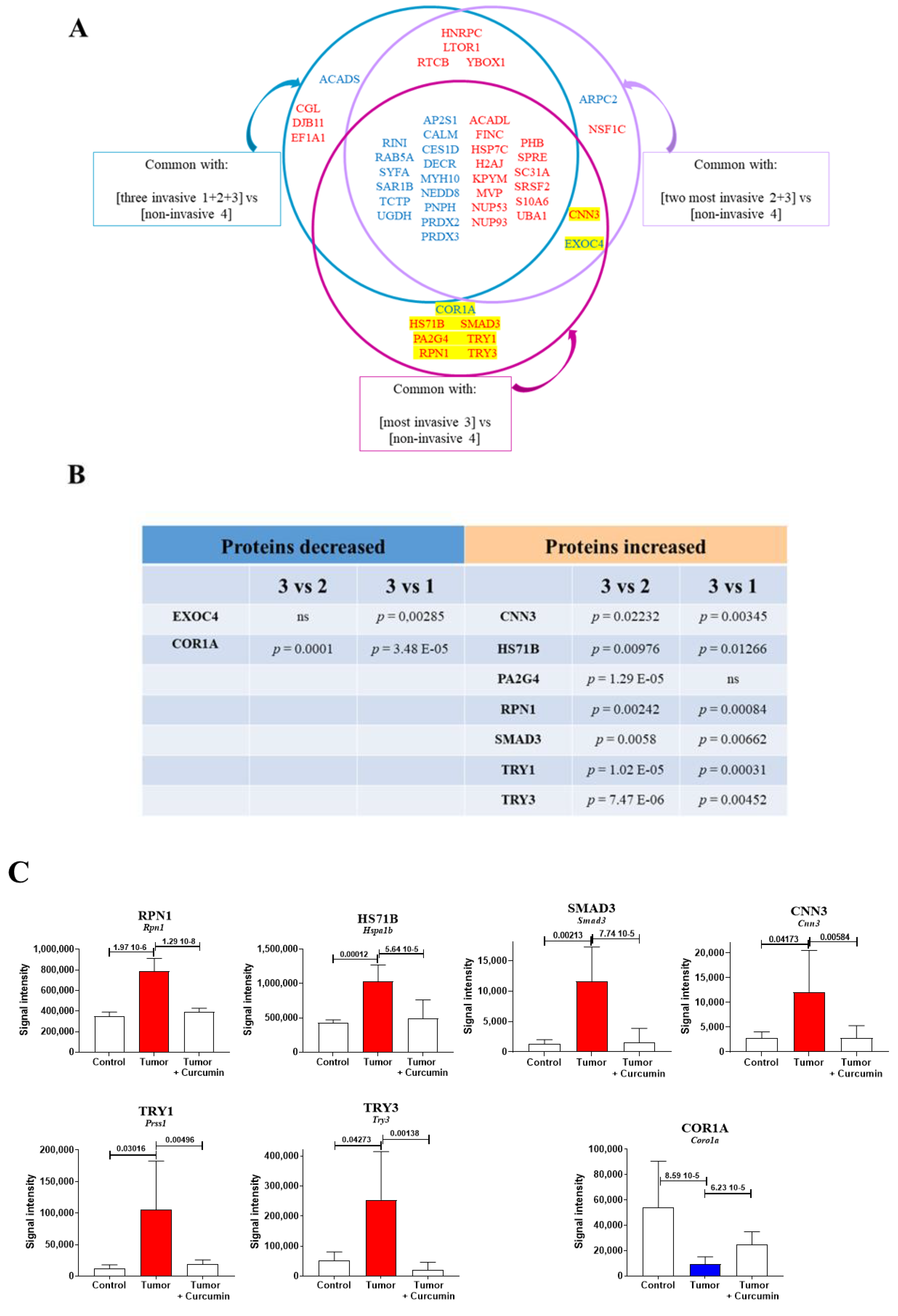
2.3. Biomarkers Specifically Related to Metastatic Colonization of the Liver by M5-T1 Cells
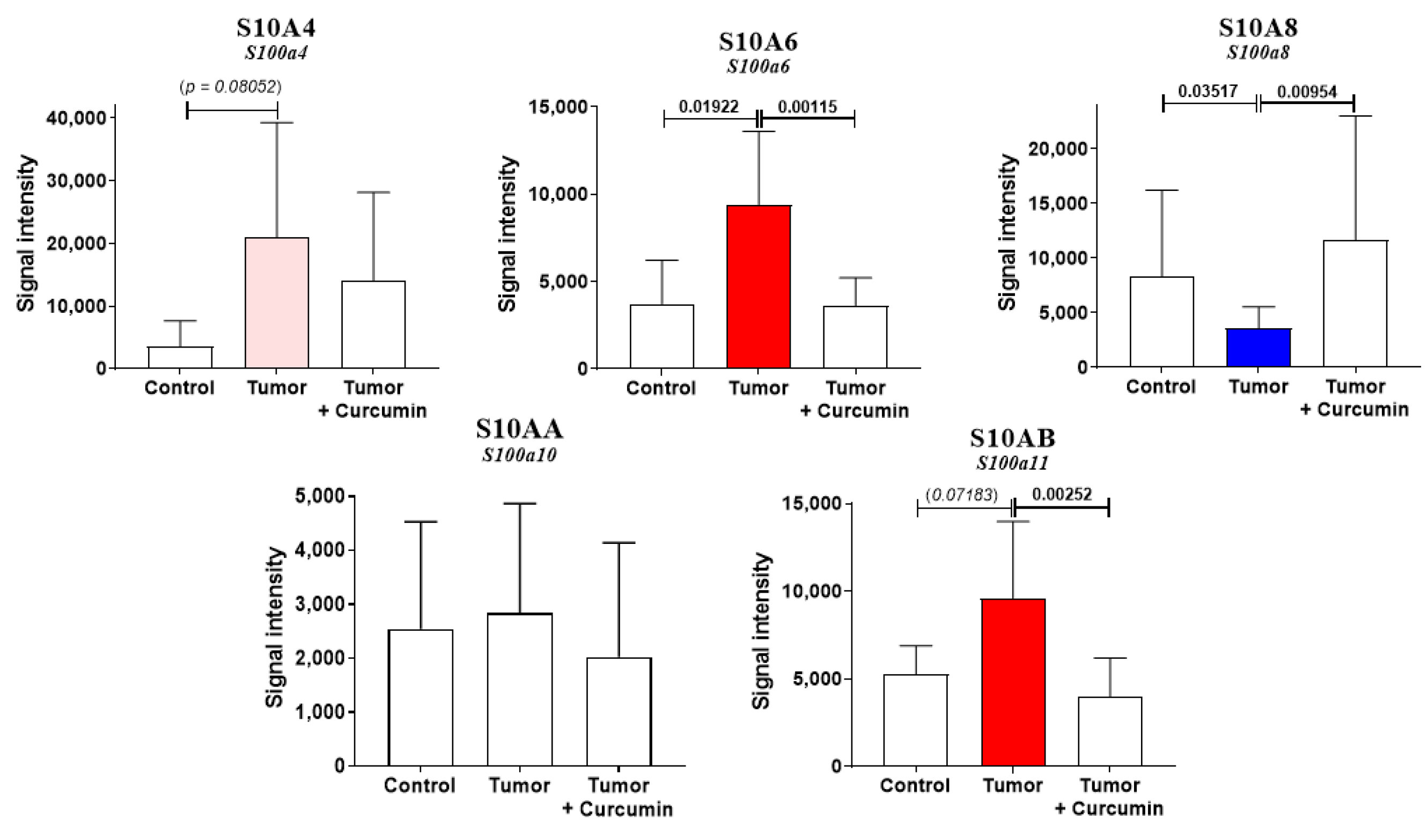
2.4. S100 Differential Abundance Proteins
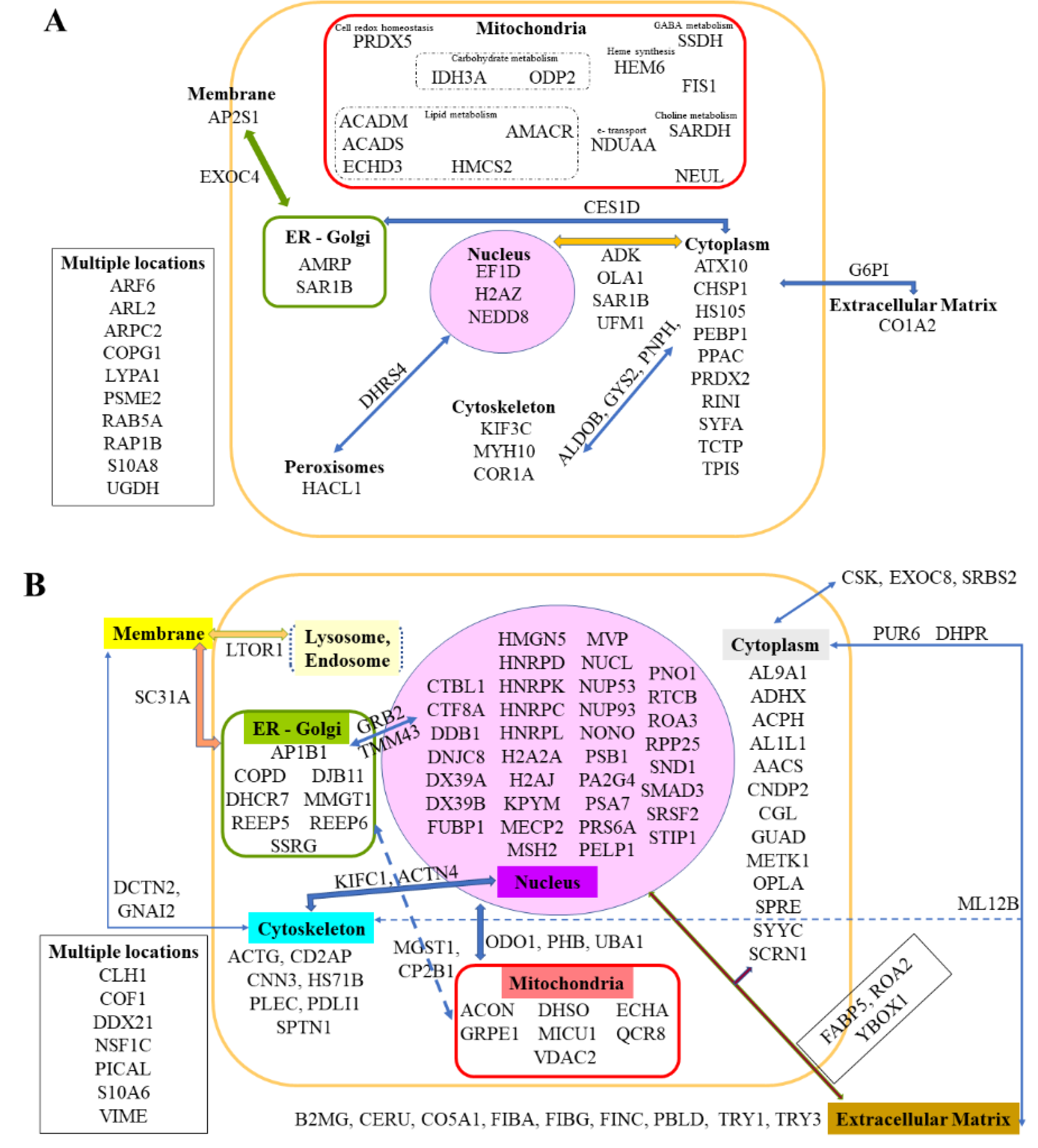
2.5. Common Biomarkers of Liver Colonization by M5-T1 Cells and the M5-T1 Tumor
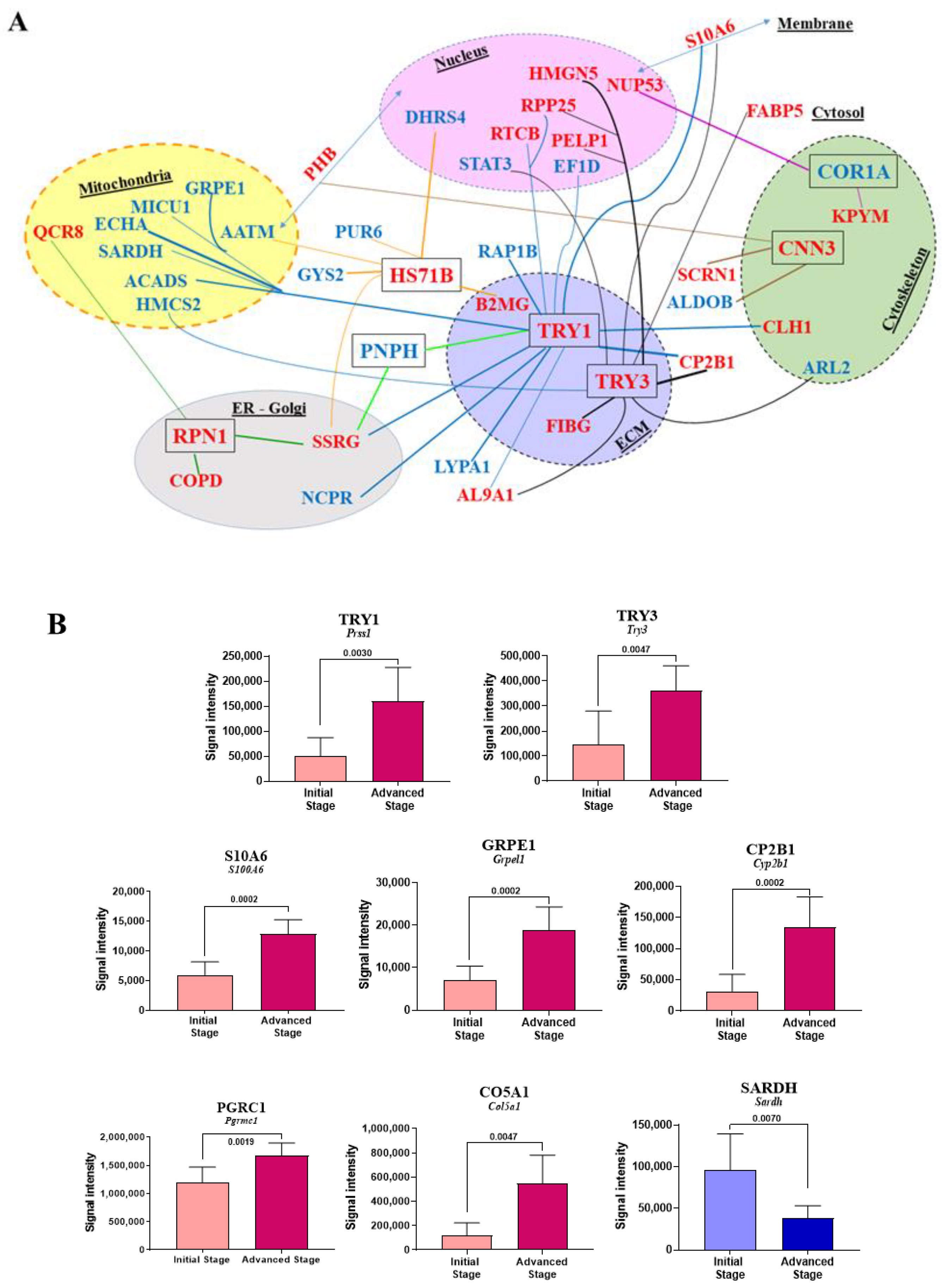
2.6. Correlations between Identified Biomarkers
3. Discussion
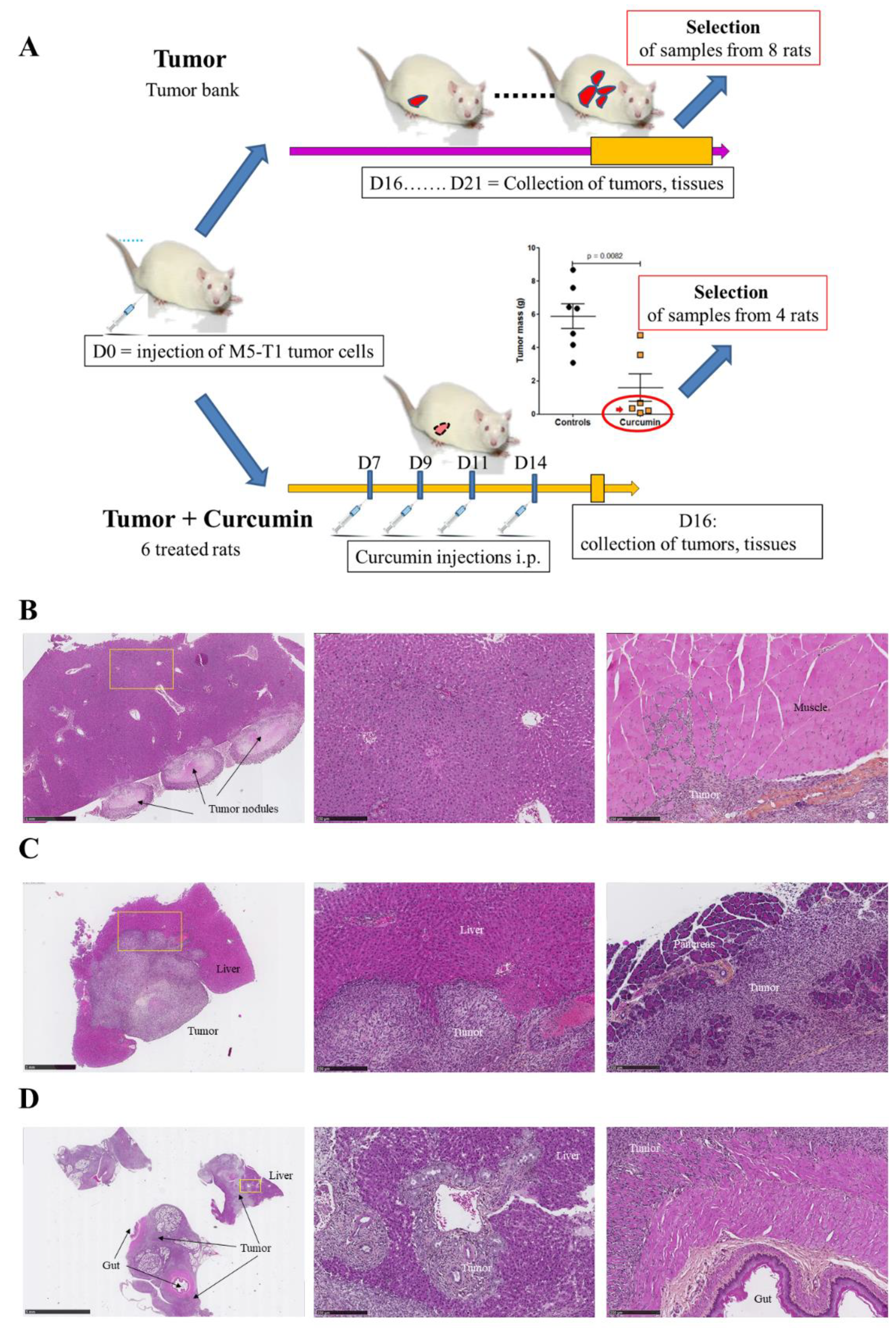
4. Materials and Methods
4.1. NMR Characterization of the Batch of Curcumin Used for Treatment
4.2. Collection of Tissues for Histological and Proteomic Analyses
4.3. Experimental Procedures for In Vivo Manipulations
4.4. Histology
4.5. Sample Preparation for SWATH-MS Analysis
4.6. Relative Quantification by SWATH Acquisition and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Talmadge, J.E.; Fidler, I.J. AACR centennial series: The biology of cancer metastasis: Historical perspective. Cancer Res. 2010, 70, 5649–5669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mielgo, A.; Schmid, M.C. Liver tropism in cancer: The hepatic metastatic niche. Cold Spring Harb. Perspect. Med. 2019. [Google Scholar] [CrossRef]
- Shibue, T.; Weinberg, R.A. Metastatic colonization: Settlement, adaptation and propagation of tumor cells in a foreign tissue environment. Semin. Cancer Biol. 2011, 21, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Pein, M.; Oskarsson, T. Microenvironment in metastasis: Roadblocks and supportive niches. Am. J. Physiol. Cell Physiol. 2015, 309, C627–C638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.W.; Beatty, G.L. Hepatocytes prepare the soil for liver metastasis. Mol. Cell. Oncol. 2019, 6, e1632686. [Google Scholar] [CrossRef]
- Perou, C.M.; Sørlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef]
- Johansson, H.J.; Socciarelli, F.; Vacanti, N.M.; Haugen, M.H.; Zhu, Y.; Siavelis, I.; Fernandez-Woodbridge, A.; Aure, M.R.; Sennblad, B.; Vesterlund, M.; et al. Breast cancer quantitative proteome and proteogenomic landscape. Nat. Commun. 2019, 10, 1600. [Google Scholar] [CrossRef]
- Djomehri, S.I.; Gonzalez, M.E.; da Veiga Leprevost, F.; Tekula, S.R.; Chang, H.-Y.; White, M.J.; Cimino-Mathews, A.; Burman, B.; Basrur, V.; Argani, P.; et al. Quantitative proteomic landscape of metaplastic breast carcinoma pathological subtypes and relationship to triple-negative tumors. Nat. Commun. 2020, 11, 1723. [Google Scholar] [CrossRef] [Green Version]
- Greenblatt, R.B. Sarcomatoid metastases in the lymph nodes draining a primary carcinoma with a sarcomatoid stroma. Am. J. Pathol. 1933, 9, 525–538. [Google Scholar]
- Klebe, S.; Brownlee, N.A.; Mahar, A.; Burchette, J.L.; Sporn, T.A.; Vollmer, R.T.; Roggli, V.L. Sarcomatoid mesothelioma: A clinical-pathological correlation of 326 cases. Mod. Pathol. 2010, 23, 470–479. [Google Scholar] [CrossRef] [Green Version]
- Rice, J.M.; Kovatch, R.M.; Anderson, L.M. Intraperitoneal mesotheliomas induced in mice by a polycyclic aromatic hydrocarbon. J. Toxicol. Environ. Health 1989, 27, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Greim, H.; Utell, M.J.; Maxim, L.D.; Niebo, R. Perspectives on refractory ceramic fiber (RCF) carcinogenicity: Comparisons with other fibers. Inhal. Toxicol. 2014, 26, 789–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, Y.; Hamasaki, M.; Yoshimura, M.; Matsumoto, S.; Sato, A.; Tsujimura, T.; Ueda, H.; Makihata, S.; Kato, F.; Iwasaki, A.; et al. A combination of MTAP and BAP1 immunohistochemistry is effective for distinguishing sarcomatoid mesothelioma from fibrous pleuritis. Lung Cancer 2018, 125, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Abrahao-Machado, L.F.; Antunes, B.; Filippi, R.Z.; Volc, S.; Boldrini, E.; Menezes, W.P.; Reis, R.M.; de Camargo, O.P. Loss of MTAP expression is a negative prognostic marker in Ewing sarcoma family of tumors. Biomark. Med. 2018, 12, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Nader, J.S.; Abadie, J.; Deshayes, S.; Boissard, A.; Blandin, S.; Blanquart, C.; Boisgerault, N.; Coqueret, O.; Guette, C.; Grégoire, M.; et al. Characterization of increasing stages of invasiveness identifies stromal/cancer cell crosstalk in rat models of mesothelioma. Oncotarget 2018, 9, 16311–16329. [Google Scholar] [CrossRef] [Green Version]
- Nader, J.S.; Guillon, J.; Petit, C.; Boissard, A.; Franconi, F.; Blandin, S.; Lambot, S.; Grégoire, M.; Verrièle, V.; Nawrocki-Raby, B.; et al. S100A4 is a biomarker of tumorigenesis, EMT, Invasion, and colonization of host organs in experimental malignant mesothelioma. Cancers 2020, 12, 939. [Google Scholar] [CrossRef] [Green Version]
- Pouliquen, D.L.; Nawrocki-Raby, B.; Nader, J.; Blandin, S.; Robard, M.; Birembaut, P.; Grégoire, M. Evaluation of intracavitary administration of curcumin for the treatment of sarcomatoid mesothelioma. Oncotarget 2017, 8, 57552–57573. [Google Scholar] [CrossRef] [Green Version]
- Rosen, F.S. The acquired immunodeficiency syndrome (AIDS). J. Clin. Investig. 1985, 75, 1–3. [Google Scholar] [CrossRef]
- Bzowska, A.; Kulikowska, E.; Shugar, D. Purine nucleoside phosphorylases: Properties, functions, and clinical aspects. Pharmacol. Ther. 2000, 88, 349–425. [Google Scholar] [CrossRef]
- Arpaia, E.; Benveniste, P.; Di Cristofano, A.; Gu, Y.; Dalal, I.; Kelly, S.; Hershfield, M.; Pandolfi, P.P.; Roifman, C.M.; Cohen, A. Mitochondrial basis for immune deficiency: Evidence from purine nucleoside phosphorylase-deficient mice. J. Exp. Med. 2000, 191, 2197. [Google Scholar] [CrossRef] [Green Version]
- Schramm, V.L. Development of transition state analogues of purine nucleoside phosphorylase as anti-T-cell agents. Biochim. Biophys. Acta 2002, 1587, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Sartorelli, A.C. Conversion of 6-thioguanine to the nucleoside level by purine nucleoside phosphorylase of Sarcoma 180 and Sarcoma 180/TG ascites cells. Cancer Res. 1981, 41, 1086–1090. [Google Scholar] [PubMed]
- Munshi, P.N.; Lubin, M.; Bertino, J.R. 6-thioguanine: A drug with unrealized potential for cancer therapy. Oncologist 2014, 19, 760–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuoka, R.; Shiba-Ishii, A.; Nakano, N.; Togayachi, A.; Sakashita, S.; Sato, Y.; Minami, Y.; Noguchi, M. Heterotopic production of ceruloplasmin by lung adenocarcinoma is significantly correlated with prognosis. Lung Cancer 2018, 118, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, Y.; Sato, Y.; Asahi-Ozaki, Y.; Ito, E.; Honma, R.; Imai, J.; Kanno, T.; Kano, M.; Akiyama, H.; Sata, T.; et al. Effusion and solid lymphomas have distinctive gene and protein expression profiles in an animal model of primary effusion lymphoma. J. Pathol. 2006, 209, 464–473. [Google Scholar] [CrossRef]
- Punwani, D.; Pelz, B.; Yu, J.; Arva, N.C.; Schafernak, K.; Kondratowicz, K.; Makhija, M.; Puck, J.M. Coronin-1A: Immune deficiency in humans and mice. J. Clin. Immunol. 2015, 35, 100–107. [Google Scholar] [CrossRef] [Green Version]
- Ciuba, K.; Hawkes, W.; Tojkander, S.; Kogan, K.; Engel, U.; Iskratsch, T.; Lappalainen, P. Calponin-3 is critical for coordinated contractility of actin stress fibers. Sci. Rep. 2018, 8, 17670. [Google Scholar] [CrossRef]
- Sakamoto, A.; Oda, Y.; Yamamoto, H.; Oshiro, Y.; Miyajima, K.; Itakura, E.; Tamiya, S.; Honda, Y.; Ishihara, A.; Iwamoto, Y.; et al. Calponin and h-caldesmon expression in atypical fibroxanthoma and superficial leiomyosarcoma. Virchows Arch. 2002, 440, 404–409. [Google Scholar] [CrossRef]
- Rekhi, B.; Vogel, U.; Basak, R.; Desai, S.B.; Jambhekar, N.A. Clinicopathological and molecular spectrum of Ewing sarcomas/PNETs, including validation of EWSR1 rearrangement by conventional and array FISH technique in certain cases. Pathol. Oncol. Res. 2014, 20, 503–516. [Google Scholar] [CrossRef]
- Hong, K.-S.; Kim, H.; Kim, S.-H.; Kim, M.; Yoo, J. Calponin 3 regulates cell invasion and doxorubicin resistance in gastric cancer. Gastroenterol. Res. Pract. 2019, 2019, 3024970. [Google Scholar] [CrossRef]
- Nair, V.A.; Al-Khayyal, N.A.; Sivaperumal, S.; Abdel-Rahman, W.M. Calponin 3 promotes invasion and drug resistance of colon cancer cells. World J. Gastrointest. Oncol. 2019, 11, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Gilston, B.A.; Skaar, E.P.; Chazin, W.J. Binding of transition metals to S100 proteins. Sci. China Life Sci. 2016, 59, 792–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, C.-M.; Yang, F.-X.; Wang, P.-L.; Yang, Q.-X.; Sun, X.-R. Clinicopathologic significance of S100A4 expression in osteosarcoma. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 833–839. [Google Scholar] [PubMed]
- Madda, R.; Chen, C.-M.; Wang, J.-Y.; Chen, C.-F.; Chao, K.-Y.; Yang, Y.-M.; Wu, H.-Y.; Chen, W.-M.; Wu, P.-K. Proteomic profiling and identification of significant markers from high-grade osteosarcoma after cryotherapy and irradiation. Sci. Rep. 2020, 10, 2105. [Google Scholar] [CrossRef]
- Martinez-Aguilar, J.; Clifton-Bligh, R.; Molloy, M.P. A multiplexed, targeted mass spectrometry assay of the S100 protein family uncovers the isoform-specific expression in thyroid tumours. BMC Cancer 2015, 15, 199. [Google Scholar] [CrossRef] [Green Version]
- Cabezón, T.; Celis, J.E.; Skibshøj, I.; Klingelhöfer, J.; Grigorian, M.; Gromov, P.; Rank, F.; Myklebust, J.H.; Maelandsmo, G.M.; Lukanidin, E.; et al. Expression of S100A4 by a variety of cell types present in the tumor microenvironment of human breast cancer. Int. J. Cancer 2007, 121, 1433–1444. [Google Scholar] [CrossRef]
- Donato, R.; Sorci, G.; Giambanco, I. S100A6 protein: Functional roles. Cell. Mol. Life Sci. 2017, 74, 2749–2760. [Google Scholar] [CrossRef]
- Zhang, M.-X.; Gan, W.; Jing, C.-Y.; Zheng, S.-S.; Yi, Y.; Zhang, J.; Xu, X.; Lin, J.J.; Zhang, B.-H.; Qiu, S.-J. S100A11 promotes cell proliferation via P38/MAPK signaling pathway in intrahepatic cholangiocarcinoma. Mol. Carcinog. 2018, 58, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Meng, M.; Sang, L.; Wang, X. S100 calcium binding protein A11 (S100A11) promotes the proliferation, migration and invasion of cervical cancer cells, and activates Wnt/b-catenin signaling. Oncotargets Ther. 2019, 12, 8675–8685. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.-F.; Li, T.; Jiang, F.; Ni, W.-K.; Guan, C.-Q.; Liu, Z.-X.; Lu, C.-H.; Ni, R.-Z.; Wu, W.; Xiao, M.-B. Correlation between S100A11 and the TGF-β1/SMAD4 pathway and its effects on the proliferation and apoptosis of pancreatic cancer cell line PANC-1. Mol. Cell. Biochem. 2019, 450, 53–64. [Google Scholar] [CrossRef]
- Wang, C.; Lin, C.; Tao, Q.; Zhao, S.; Liu, H.; Li, L. Evaluation of calcium-binding protein A11 promotes the carcinogenesis of hypopharygeal squamous cell carcinoma via the PI3K/AKT signaling pathway. Am. J. Transl. Res. 2019, 11, 3472–3480. [Google Scholar] [PubMed]
- Reckenbeil, J.; Kraus, D.; Probstmeier, R.; Allam, J.-P.; Novak, N.; Frentzen, M.; Martini, M.; Wenghoefer, M.; Winter, J. Cellular distribution and gene expression pattern of metastasin (S100A4), Calgranulin A (S100A8), and Calgranulin B (S100A9) in oral lesions as markers for molecular biology. Cancer Investig. 2016, 34, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Diederichs, S.; Bulk, E.; Steffen, B.; Ji, P.; Tickenbrock, L.; Lang, K.; Zänker, K.S.; Metzger, R.; Schneider, P.M.; Gerke, V.; et al. S100 family members and trypsinogens are predictors of distant metastasis and survival in early-stage non-small cell lung cancer. Cancer Res. 2004, 64, 5564–5569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilardi, C.; Silini, A.; Figini, S.; Anastasia, A.; Lupi, M.; Fruscio, R.; Giavazzi, R.; Bani, M. Trypsinogen 4 boosts tumor endothelial cells migration through proteolysis of tissue factor pathway inhibitor-2. Oncotarget 2015, 6, 28389–28400. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Hu, Y.-L.; Feng, Y.; Guo, Y.-B.; Liu, Y.-F.; Mao, Q.-S.; Xue, W.-J. High-level expression of PRSS3 correlates with metastasis and poor prognosis in patients with gastric cancer. J. Surg. Oncol. 2019, 119, 1108–1121. [Google Scholar] [CrossRef]
- Fan, Y.; Guo, Y.; Hamblin, M.; Chang, L.; Zhang, J.; Chen, Y.E. Inhibition of gluconeogenic genes by calcium-regulated heat-stable protein 1 via repression of peroxisome proliferator-activated receptor α. J. Biol. Chem. 2011, 286, 40584–40594. [Google Scholar] [CrossRef] [Green Version]
- Petrovchich, I.; Ford, J.M. Genetic predisposition to gastric cancer. Rev. Semin. Oncol. 2016, 43, 554–559. [Google Scholar] [CrossRef] [Green Version]
- Chmielecki, J.; Bailey, M.; He, J.; Elvin, J.; Vergilio, J.-A.; Ramkissoon, S.; Suh, J.; Frampton, G.M.; Sun, J.X.; Morley, S.; et al. Genomic profiling of a large set of diverse pediatric cancers identifies known and novel mutations across tumor spectra. Cancer Res. 2017, 77, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Koivunen, E.; Saksela, O.; Itkonen, O.; Osman, S.; Huhtala, M.-L.; Stenman, U.-K. Human colon carcinoma, fibrosarcoma and leukemia cell lines produce tumor-associated trypsinogen. Int. J. Cancer 1991, 47, 592–596. [Google Scholar] [CrossRef]
- Chen, Q.-Q.; Gao, F.; Zhuang, Z.-H.; Zhang, Z.-B.; Liu, Q.-C.; Chen, J.-T. Trypsin-antitrypsin imbalance in immune escape and clonal proliferation of pancreatic cancer. J. Genet. Syndr. Gene Ther. 2013, 4, 11. [Google Scholar]
- Kirby, G.M.; Batist, G.; Alpert, L.; Lamoureux, E.; Cameron, R.G.; Alaoui-Jamali, M.A. Overexpression of cytochrome P-450 isoforms involved in Aflatoxin B1 bioactivation in human liver with cirrhosis and hepatitis. Toxicol. Pathol. 1996, 24, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Tang, R.; Wu, D.; Sun, X. Research advances in HMGN5 and cancer. Tumor Biol. 2016, 37, 1531–1539. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, X.; Wu, F.; Zhang, X.; Zhang, H.; Gao, D.; Bi, D.; Qu, H.; Ge, J.; Xu, Y.; et al. HADHA overexpression disrupts lipid metabolism and inhibits tumor growth in clear cell renal cell carcinoma. Exp. Cell Res. 2019, 384, 111558. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Barber, G.N. STING an endoplasmic reticulum adaptor that facilitates innate immune signaling. Nature 2008, 455, 674–678. [Google Scholar] [CrossRef]
- Chano, T.; Mori, K.; Scotlandi, K.; Benini, S.; Lapucci, C.; Manara, M.C.; Serra, M.; Picci, P.; Okabe, H.; Baldini, N. Differentially expressed genes in multidrug resistant variants of U-2 OS human osteosarcoma cells. Oncol. Rep. 2004, 11, 1227–1263. [Google Scholar] [CrossRef]
- Dybkaer, K.; Iqbal, J.; Zhou, G.; Geng, H.; Xiao, L.; Schmitz, A.; d’Amore, F.; Chan, W.C. Genome wide transcriptional analysis of resting and IL2 activated human natural killer cells: Gene expression signatures indicative of novel molecular signaling pathways. BMC Genom. 2007, 8, 230. [Google Scholar] [CrossRef] [Green Version]
- Varelas, X.; Bouchie, M.P.; Kukuruzinska, M.A. Protein N-glycosylation in oral cancer: Dysregulated cellular networks among DPAGT1, E-cadherin adhesion and canonical Wnt signaling. Glycobiology 2014, 24, 579–591. [Google Scholar] [CrossRef] [Green Version]
- Harada, Y.; Ohkawa, Y.; Kizuka, Y.; Taniguchi, N. Oligosaccharyltransferase: A gatekeeper of health and tumor progression. Int. J. Mol. Sci. 2019, 20, 6074. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stokes, J., III; Singh, U.P.; Gunn, S.K.; Acharya, A.; Manne, U.; Mishra, M. Targeting Hsp70: A possible therapy for cancer. Cancer Lett. 2016, 374, 156–166. [Google Scholar] [CrossRef] [Green Version]
- Kuramochi, M.; Izawa, T.; Pervin, M.; Bondoc, A.; Kuwamura, M.; LaMarre, J.; Yamate, J. Attenuation of thioacetamide-induced hepatocellular injury by short-term repeated injections associated with down-regulation of metabolic enzymes and relationship with MHC class II-presenting cells. Exp. Toxicol. Pathol. 2017, 69, 589–597. [Google Scholar] [CrossRef]
- Myers, S.M.; Mulligan, L.M. The RET receptor is linked to stress response pathways. Cancer Res. 2004, 64, 4453–4463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.-L.; Zhang, C.Z.; Liu, L.-L.; Lu, S.-X.; Pan, Y.-H.; Wang, C.-H.; He, Y.-F.; Lin, C.-S.; Yang, X.; Xie, D.; et al. A Gys2/p53 negative feedback loop restricts tumor growth in HBV-related hepatocellular carcinoma. Cancer Res. 2019, 79, 534–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldi, A.; De Luca, A.; Maiorano, P.; D’Angelo, C.; Giordano, A. Curcumin as an anticancer agent in malignant mesothelioma: A review. Int. J. Mol. Sci. 2020, 21, 1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hocking, A.; Tommasi, S.; Sordillo, P.; Klebe, S. The safety and exploration of the pharmacokinetics of intrapleural liposomal curcumin. Int. J. Nanomed. 2020, 15, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Benassi, R.; Ferrari, E.; Lazzari, S.; Spagnolo, F.; Saladini, M. Theoretical study on curcumin: A comparison of calculated spectroscopic properties with NMR, UV-vis and IR experimental data. J. Mol. Struct. 2008, 892, 168–176. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Initial St. (G2) /Control (G1) | Advanced St. (G2) /Control (G1) | + Curcumin (G3) /Initial St. (G2) | + Curcumin (G3) /Advanced St. (G2) |
|---|---|---|---|---|
| FETUA | ns (0.3677) | 0.0081 | 0.0104 | 0.0002 |
| IRGM | 0.0283 | 0.0040 | ns (0.1605) | ns (0.2345) |
| TRY1 | 0.0727 | 0.0040 | 0.0830 | 0.0002 |
| TRY3 | ns (0.5697) | 0.0040 | 0.0011 | 0.0002 |
| ACADS | 0.0727 | 0.0081 | ns (0.1049) | 0.0047 |
| CHSP1 | 0.0040 | 0.0040 | 0.0002 | 0.0002 |
| SARDH | ns (0.1535) | 0.0485 | ns (0.1304) | 0.0104 |
| ADHX | 0.0485 | 0.0283 | 0.0830 | 0.0070 |
| AL1L1 | 0.0727 | 0.0040 | ns (0.2345) | 0.0030 |
| CO5A1 | ns (0.1535) | 0.0162 | 0.0281 | 0.0006 |
| CP2B1 | ns (0.5697) | 0.0040 | ns (0.1949) | 0.0002 |
| DNJC8 | ns (0.2141) | 0.0040 | ns (0.3823) | 0.0011 |
| DX39A | ns (0.6828) | ns (0.1091) | 0.0104 | ns (0.8785) |
| FIBG | 0.0162 | 0.0040 | 0.0003 | 0.0002 |
| GRPE1 | ns (0.1091) | 0.0040 | ns (0.2786) | 0.0002 |
| MMGT1 | ns (0.2828) | 0.0162 | ns (0.1049) | 0.0019 |
| PDLI1 | 0.0485 | 0.0040 | 0.0148 | 0.0011 |
| PELP1 | 0.0081 | 0.0040 | ns (0.1049) | 0.0019 |
| PGRC1 | 0.0040 | 0.0040 | 0.0003 | 0.0002 |
| RTCB | 0.0040 | 0.0040 | 0.0019 | 0.0006 |
| S10A6 | ns (0.2141) | 0.0081 | 0.0104 | 0.0002 |
| Protein | Initial St. (G2) /Control (G1) | Advanced St. (G2) /Control (G1) | + Curcumin (G3) /Initial St. (G2) | + Curcumin (G3) /Advanced St. (G2) |
|---|---|---|---|---|
| AP2S1 | 0.0040 | 0.0040 | 0.0003 | 0.0030 |
| ARL2 | 0.0162 | ns (0.1091) | 0.0003 | 0.0148 |
| DJB11 | 0.0040 | 0.0162 | 0.0003 | 0.0030 |
| ECHA | 0.0040 | 0.0081 | 0.0002 | 0.0011 |
| FABP5 | 0.0040 | 0.0283 | 0.0002 | 0.0070 |
| FINC | 0.0040 | 0.0485 | 0.0019 | ns (0.1605) |
| H2AJ | 0.0040 | 0.0081 | 0.0030 | ns (0.1949) |
| HMCS2 | 0.0040 | ns (0.2828) | 0.0002 | ns (0.3823) |
| HNRPK | 0.0040 | ns (0.1091) | 0.0002 | 0.0830 |
| IDH3A | 0.0081 | ns (0.1535) | 0.0006 | 0.0047 |
| LYPA1 | 0.0040 | 0.0040 | 0.0002 | 0.0003 |
| NCPR | 0.0040 | 0.0040 | 0.0002 | 0.0002 |
| PA2G4 | 0.0040 | 0.0162 | 0.0002 | 0.0650 |
| PICAL | 0.0081 | 0.0485 | 0.0011 | 0.0379 |
| PPAC | 0.0040 | 0.0040 | 0.0070 | 0.0104 |
| SRSF2 | 0.0040 | ns (0.1091) | 0.0006 | ns (0.2345) |
| VIME | 0.0040 | 0.0162 | 0.0019 | ns (0.5054) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pouliquen, D.L.; Boissard, A.; Henry, C.; Blandin, S.; Richomme, P.; Coqueret, O.; Guette, C. Curcumin Treatment Identifies Therapeutic Targets within Biomarkers of Liver Colonization by Highly Invasive Mesothelioma Cells—Potential Links with Sarcomas. Cancers 2020, 12, 3384. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12113384
Pouliquen DL, Boissard A, Henry C, Blandin S, Richomme P, Coqueret O, Guette C. Curcumin Treatment Identifies Therapeutic Targets within Biomarkers of Liver Colonization by Highly Invasive Mesothelioma Cells—Potential Links with Sarcomas. Cancers. 2020; 12(11):3384. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12113384
Chicago/Turabian StylePouliquen, Daniel L., Alice Boissard, Cécile Henry, Stéphanie Blandin, Pascal Richomme, Olivier Coqueret, and Catherine Guette. 2020. "Curcumin Treatment Identifies Therapeutic Targets within Biomarkers of Liver Colonization by Highly Invasive Mesothelioma Cells—Potential Links with Sarcomas" Cancers 12, no. 11: 3384. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12113384