Serum-Derived Exosomal MicroRNA Profiles Can Predict Poor Survival Outcomes in Patients with Extranodal Natural Killer/T-Cell Lymphoma


Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Study Design and Patients
2.2. Isolation of Exosomes from Serum Samples and Nanostring nCounter Analyses
2.3. Cell Lines and Isolation of Exosomes from Cell Culture Medium
2.4. Transmission Electron Microscopy and Nanoparticle Tracking Analysis
2.5. RNA Extraction
2.6. Quantitative Real-Time Polymerase Chain Reaction
2.7. Western Blot
2.8. Establishment of Etoposide-Resistant Cell Lines and Cell Viability Assay
2.9. miRNA Transfection and Cytokine Array
2.10. Transwell Co-Culture Assay
2.11. Data Preparation and Statistical Analyses
3. Results
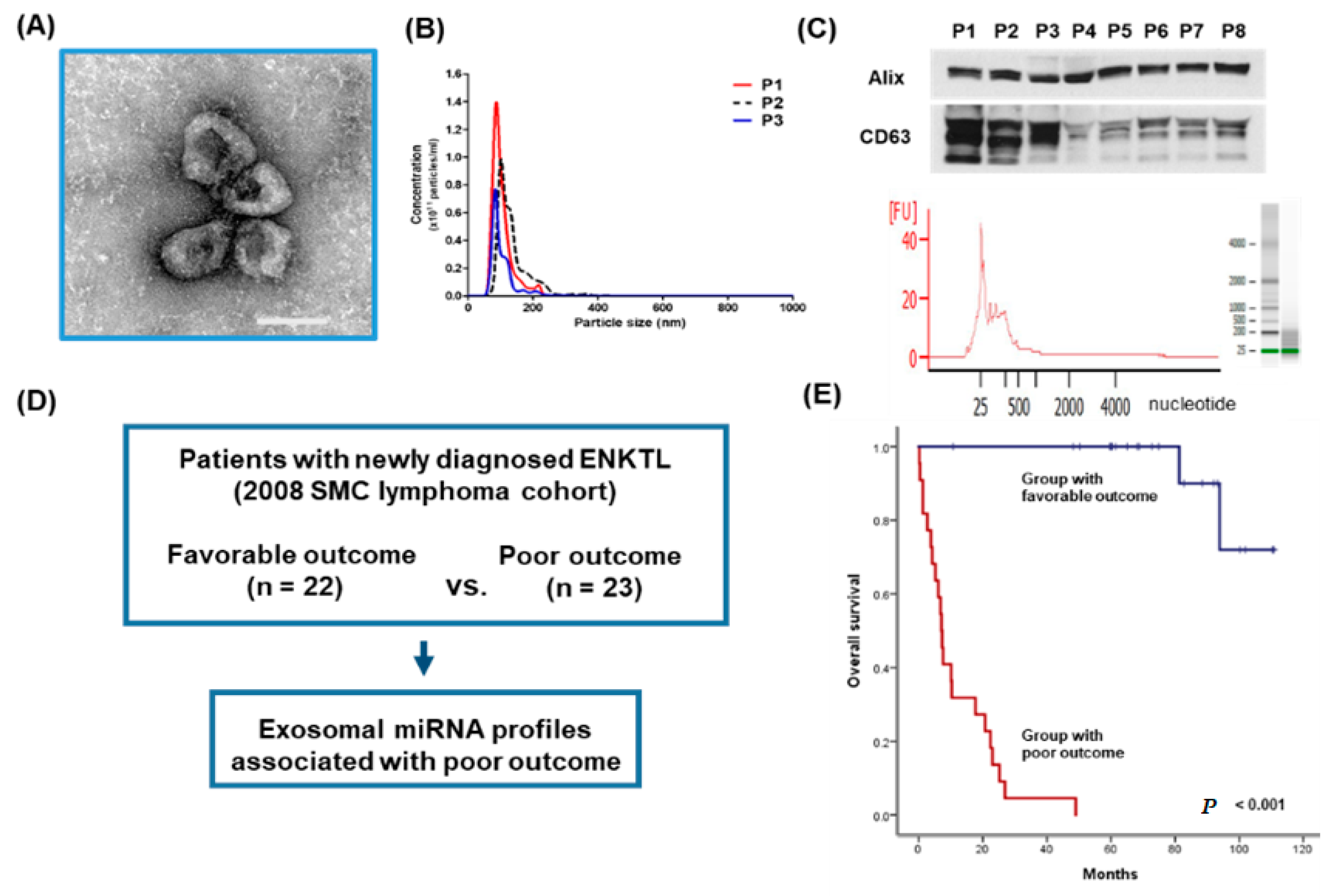
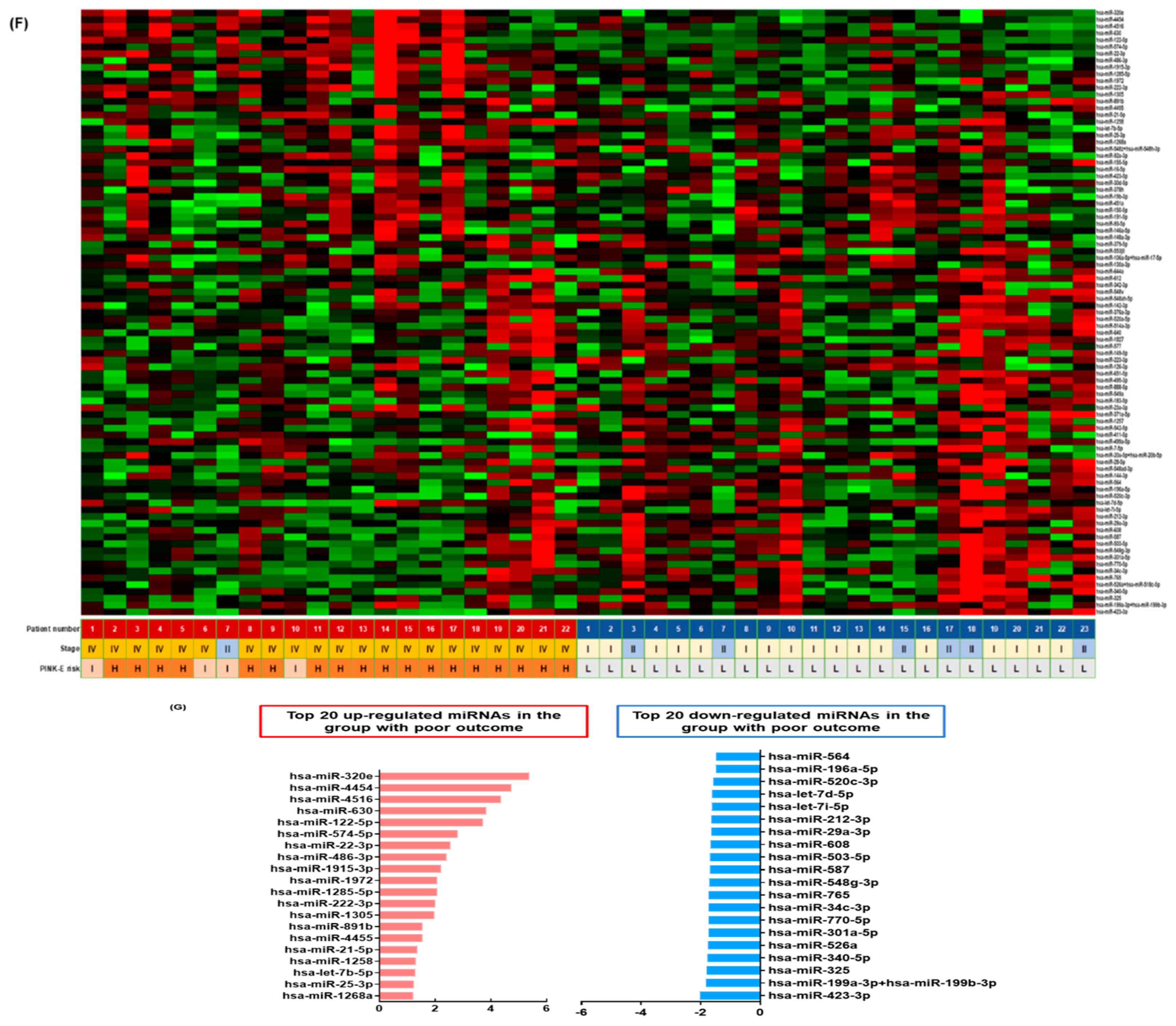
3.1. Isolation of Exosomes from Patients’ Serum in the Training Cohort
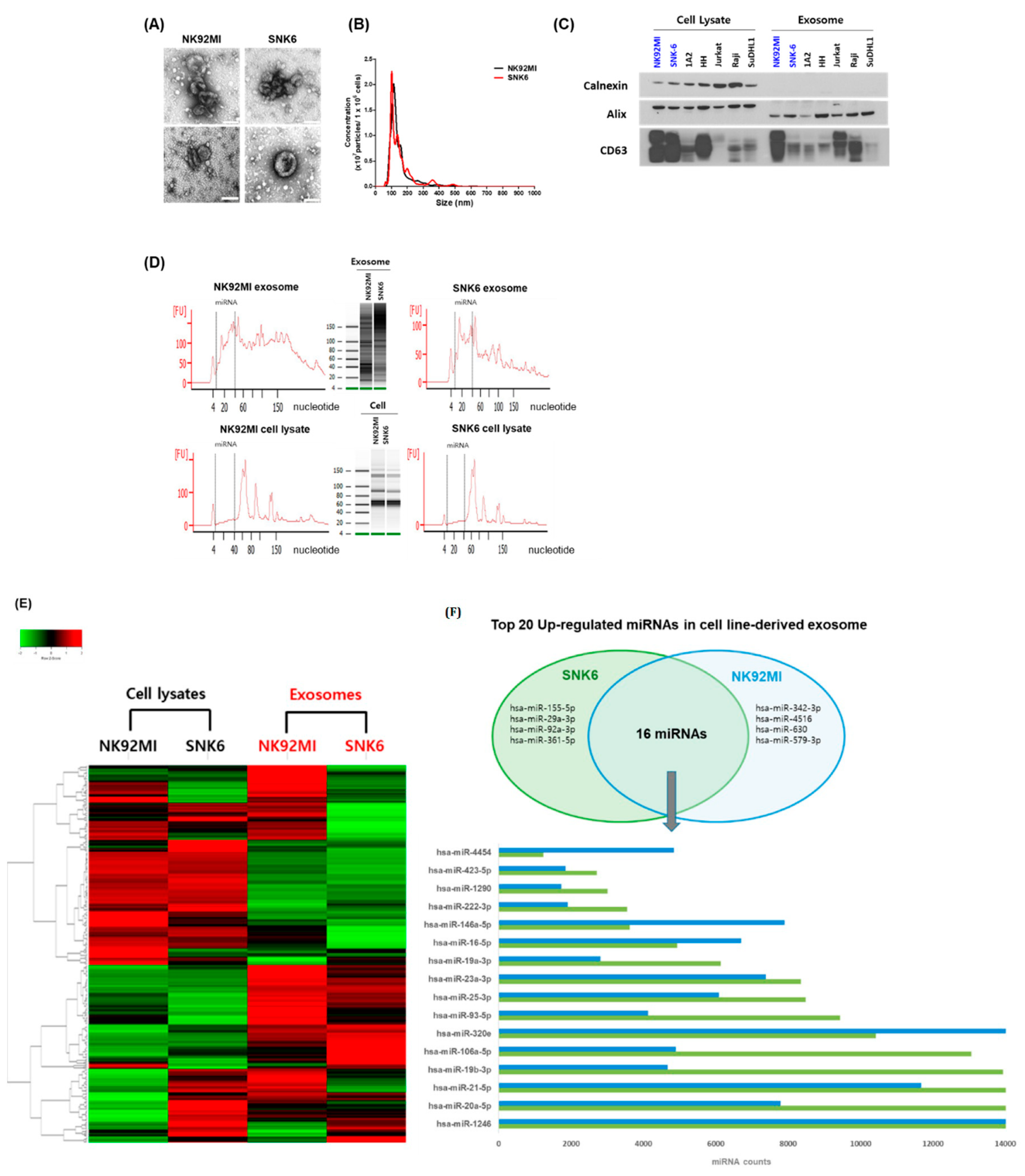
3.2. NKTL Cell Line-Derived Exosomal miRNA Profiles
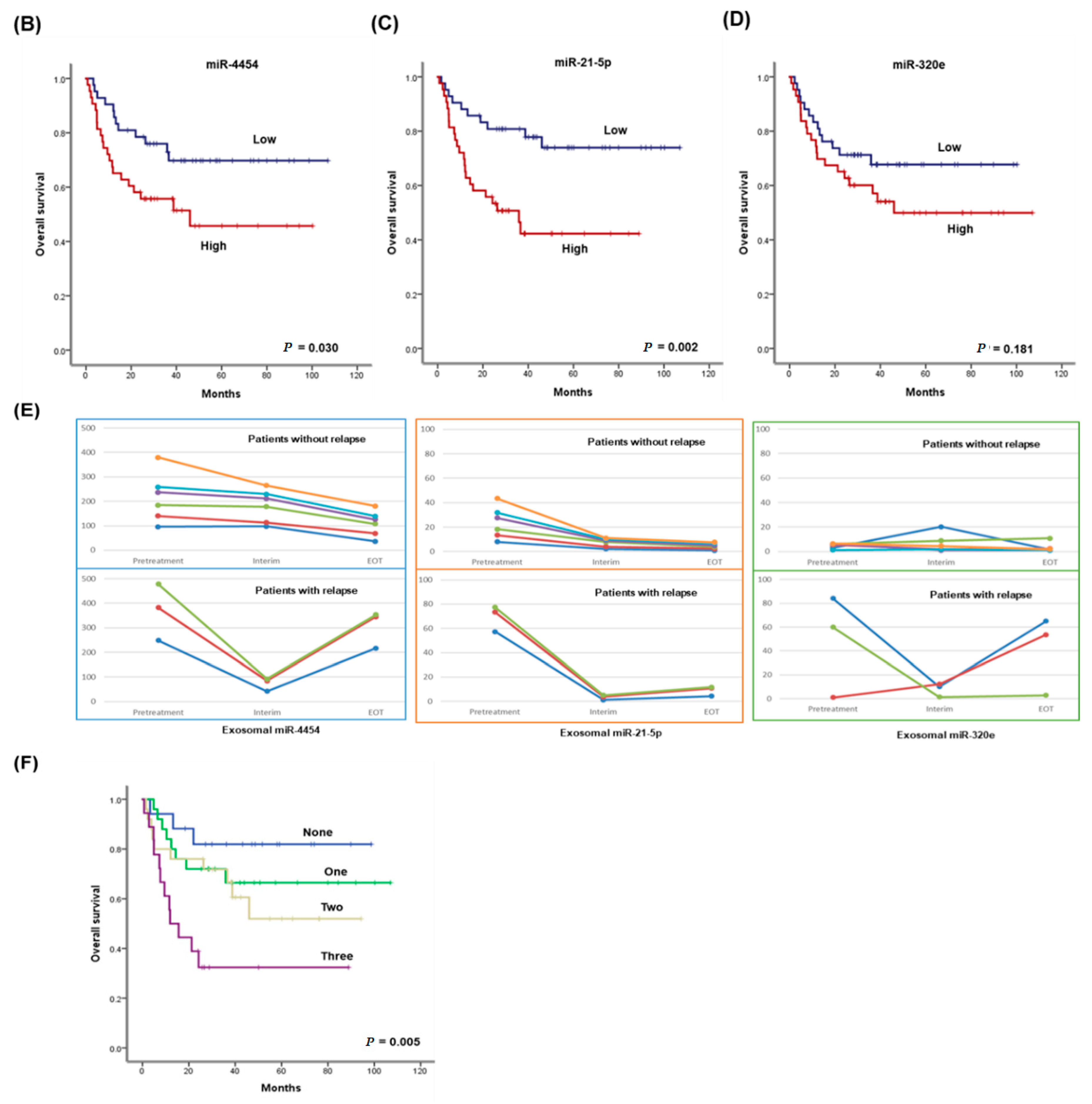
3.3. Validation of the Prognostic Relevance of Exosomal miRNAs in ENKTL
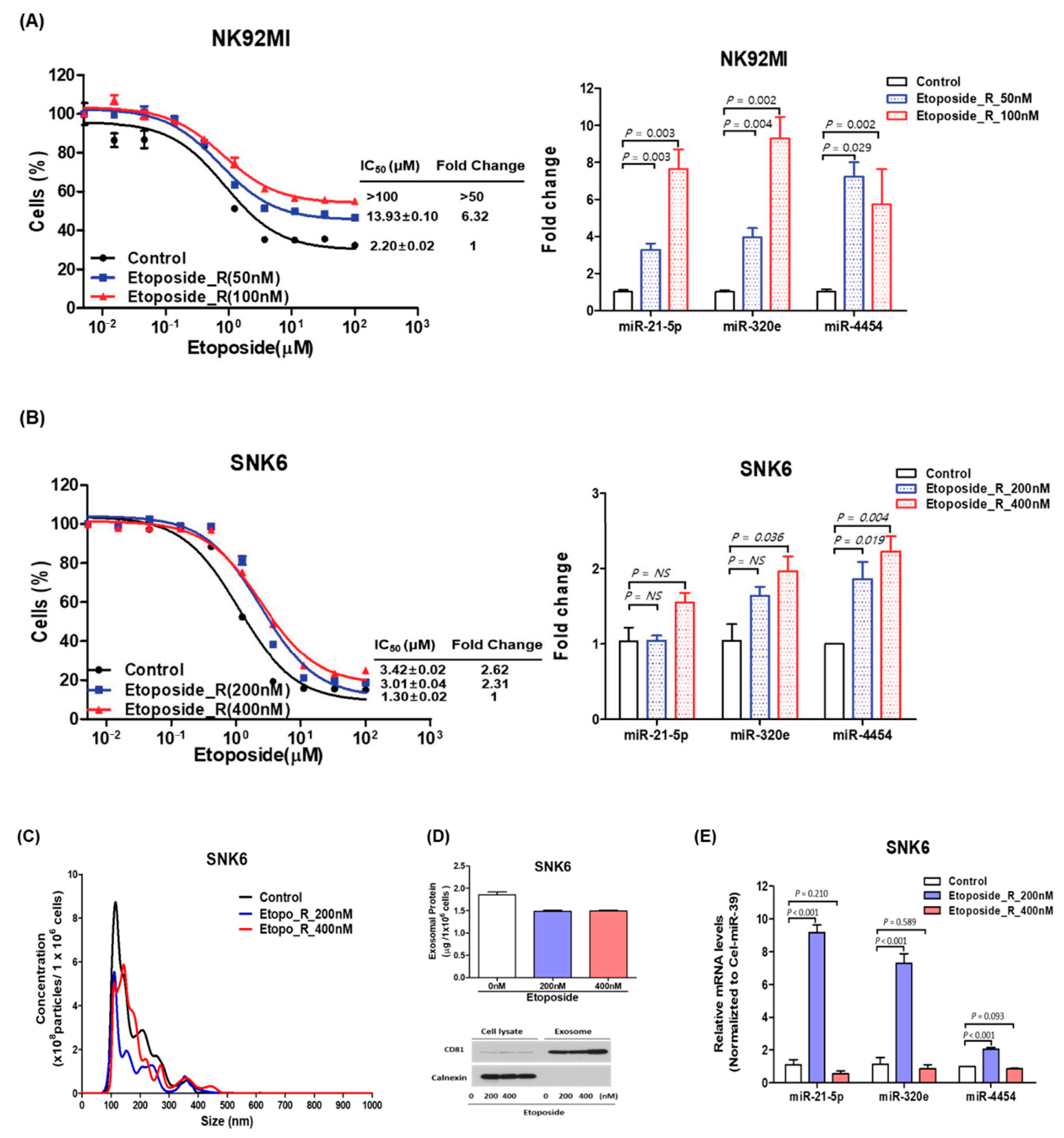
3.4. Expression of miR-21-5p, miR-4454, and miR-320e in Treatment-Resistant NKTL Cell Lines
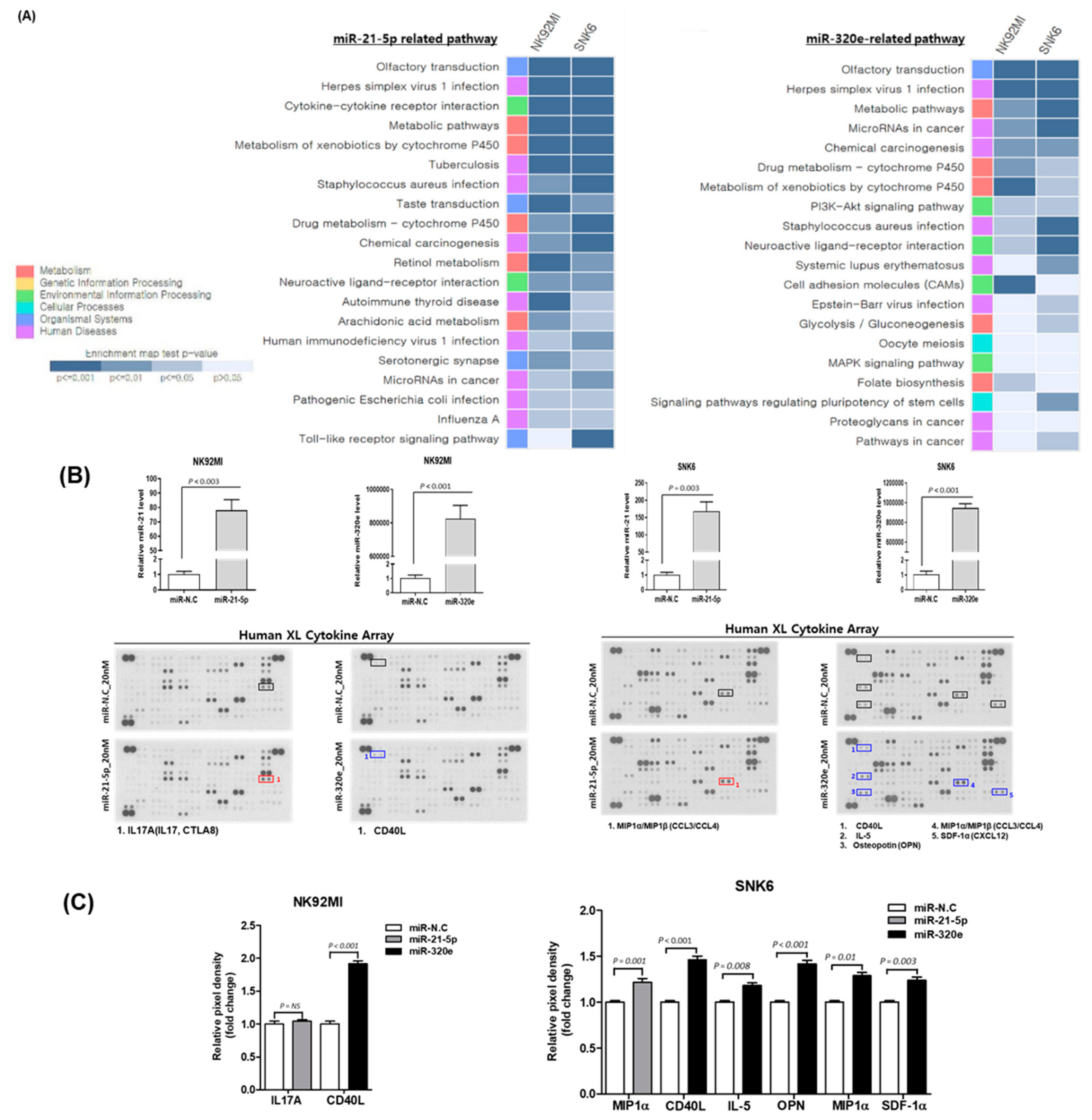
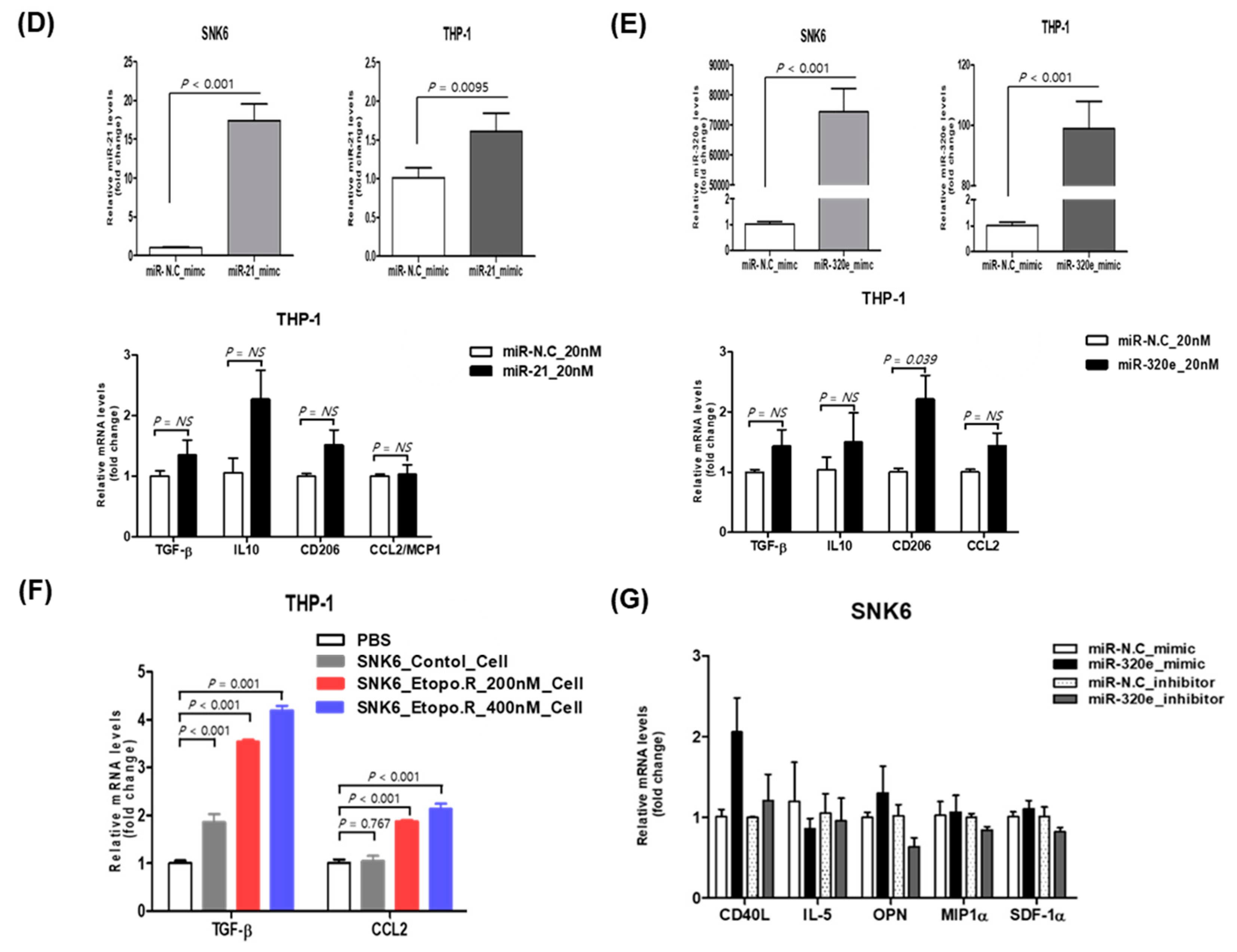
3.5. Effect of Upregulated miR-21-5p and miR-320e on Cytokine Production
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- El Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J.A. Extracellular vesicles: Biology and emerging therapeutic opportunities. Nat. Rev. Drug Discov. 2013, 12, 347–357. [Google Scholar] [CrossRef]
- Yeh, Y.-Y.; Ozer, H.G.; Lehman, A.M.; Maddocks, K.; Yu, L.; Johnson, A.J.; Byrd, J.C. Characterization of CLL exosomes reveals a distinct microRNA signature and enhanced secretion by activation of BCR signaling. Blood 2015, 125, 3297–3305. [Google Scholar] [CrossRef]
- Azmi, A.S.; Bao, B.; Sarkar, F.H. Exosomes in cancer development, metastasis, and drug resistance: A comprehensive review. Cancer Metastasis Rev. 2013, 32, 623–642. [Google Scholar] [CrossRef] [Green Version]
- Thind, A.; Wilson, C. Exosomal miRNAs as cancer biomarkers and therapeutic targets. J. Extracell. Vesicles 2016, 5, 31292. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, T.; Yip, K.W.; Spence, T.; Liu, F.-F. MicroRNAs in extracellular vesicles: Potential cancer biomarkers. J. Hum. Genet. 2016, 62, 67–74. [Google Scholar] [CrossRef]
- An, T.; Qin, S.; Xu, Y.; Tang, Y.; Huang, Y.; Situ, B.; Inal, J.M.; Zheng, L. Exosomes serve as tumour markers for personalized diagnostics owing to their important role in cancer metastasis. J. Extracell. Vesicles 2015, 4, 27522. [Google Scholar] [CrossRef]
- Kwong, Y.-L. Natural killer-cell malignancies: Diagnosis and treatment. Leukemia 2005, 19, 2186–2194. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Yoon, S.E.; Kim, W.S. Treatment of localized extranodal NK/T cell lymphoma, nasal type: A systematic review. J. Hematol. Oncol. 2018, 11, 140. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, M.; Suzuki, R.; Oguchi, M. Advances in the treatment of extranodal NK/T-cell lymphoma, nasal type. Blood 2018, 131, 2528–2540. [Google Scholar] [CrossRef]
- Yamanaka, Y.; Tagawa, H.; Takahashi, N.; Watanabe, A.; Guo, Y.-M.; Iwamoto, K.; Yamashita, J.; Saitoh, H.; Kameoka, Y.; Shimizu, N.; et al. Aberrant overexpression of microRNAs activate AKT signaling via down-regulation of tumor suppressors in natural killer–cell lymphoma/leukemia. Blood 2009, 114, 3265–3275. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.-Q.; Huang, G.L.; Guo, C.-C.; Pu, X.X.; Lin, T.Y. Diagnostic and Prognostic Value of Circulating miR-221 for Extranodal Natural Killer/T-Cell Lymphoma. Dis. Markers 2010, 29, 251–258. [Google Scholar] [CrossRef]
- Zhang, X.; Ji, W.; Huang, R.; Li, L.; Wang, X.; Li, L.; Fu, X.; Sun, Z.; Li, Z.; Chen, Q.; et al. MicroRNA-155 is a potential molecular marker of natural killer/T-cell lymphoma. Oncotarget 2016, 7, 53808–53819. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Yoon, D.H.; Jaccard, A.; Chng, W.J.; Lim, S.T.; Hong, H.; Park, Y.; Chang, K.M.; Maeda, Y.; Ishida, F.; et al. A prognostic index for natural killer cell lymphoma after non-anthracycline-based treatment: A multicentre, retrospective analysis. Lancet Oncol. 2016, 17, 389–400. [Google Scholar] [CrossRef]
- Swerdlow, S.H.; Campo, E.; Harris, N.L.; Jaffe, E.S.; Pileri, S.A.; Stein, H.; Thiele, J.; Vardiman, J.W. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues; IARC Press: Lyon, France, 2008. [Google Scholar]
- Gene Ontology. Available online: http://geneontology.org (accessed on 23 March 2020).
- KEGG. Available online: http://www.genome.jp/kegg (accessed on 23 March 2020).
- R 3.3.3. Available online: https://www.r-project.org/ (accessed on 23 March 2020).
- Zomer, A.; Vendrig, T.; Hopmans, E.S.; van Eijndhoven, M.; Middeldorp, J.M.; Pegtel, D.M. Exosomes: Fit to deliver small RNA. Commun. Integr. Biol. 2010, 3, 447–450. [Google Scholar] [CrossRef]
- Liu, J.; Wang, X. Focus on exosomes—From pathogenic mechanisms to the potential clinical application value in lymphoma. J. Cell. Biochem. 2019, 120, 19220–19228. [Google Scholar] [CrossRef]
- Zare, N.; Javanmard, S.H.; Mehrzad, V.; Eskandari, N.; Kefayat, A. Evaluation of exosomal miR-155, let-7g and let-7i levels as a potential noninvasive biomarker among refractory/relapsed patients, responsive patients and patients receiving R-CHOP. Leuk. Lymphoma 2019, 60, 1877–1889. [Google Scholar] [CrossRef]
- Feng, Y.; Zhong, M.; Zeng, S.; Wang, L.; Liu, P.; Xiao, X.; Liu, Y. Exosome-derived miRNAs as predictive biomarkers for diffuse large B-cell lymphoma chemotherapy resistance. Epigenomics 2019, 11, 35–51. [Google Scholar] [CrossRef]
- Higuchi, H.; Yamakawa, N.; Imadome, K.-I.; Yahata, T.; Kotaki, R.; Ogata, J.; Kakizaki, M.; Fujita, K.; Lu, J.; Yokoyama, K.; et al. Role of exosomes as a proinflammatory mediator in the development of EBV-associated lymphoma. Blood 2018, 131, 2552–2567. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Li, J.; Zhu, W.; Gao, C.; Jiang, R.; Li, W.; Hu, Q.; Zhang, B. MicroRNA-21 and the clinical outcomes of various carcinomas: A systematic review and meta-analysis. BMC Cancer 2014, 14, 819. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Kamohara, H.; Kinoshita, K.; Kurashige, J.; Ishimoto, T.; Iwatsuki, M.; Watanabe, M.; Baba, H. Clinical impact of serum exosomal microRNA-21 as a clinical biomarker in human esophageal squamous cell carcinoma. Cancer 2013, 119, 1159–1167. [Google Scholar] [CrossRef]
- Wang, H.; Hou, L.; Li, A.; Duan, Y.; Gao, H.; Song, X. Expression of Serum Exosomal MicroRNA-21 in Human Hepatocellular Carcinoma. BioMed Res. Int. 2014, 2014, 864894. [Google Scholar] [CrossRef]
- Alles, J.; Menegatti, J.; Motsch, N.; Hart, M.; Eichner, N.; Reinhardt, R.; Meister, G.; Grässer, F.A. miRNA expression profiling of Epstein-Barr virus-associated NKTL cell lines by Illumina deep sequencing. FEBS Open Bio 2016, 6, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Yan, J.; Phyu, T.; Fan, S.; Chung, T.-H.; Mustafa, N.B.; Lin, B.; Wang, L.; Eichhorn, P.J.A.; Goh, B.-C.; et al. MELK mediates the stability of EZH2 through site-specific phosphorylation in extranodal natural killer/T-cell lymphoma. Blood 2019, 134, 2046–2058. [Google Scholar] [CrossRef]
- Yan, J.; Li, B.; Lin, B.; Lee, P.T.; Chung, T.-H.; Tan, J.; Bi, C.; Lee, X.T.; Selvarajan, V.; Ng, S.-B.; et al. EZH2 phosphorylation by JAK3 mediates a switch to noncanonical function in natural killer/T-cell lymphoma. Blood 2016, 128, 948–958. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.; Zhang, W.; Zhang, B.; Zhao, Y.; Zhao, Y.; Li, S.; Liu, Y. miR-21 modulates the effect of EZH2 on the biological behavior of human lung cancer stem cells in vitro. Oncotarget 2017, 8, 85442–85451. [Google Scholar] [CrossRef] [Green Version]
- Perezcarbonell, L.; Sinicrope, F.A.; Alberts, S.R.; Oberg, A.L.; Balaguer, F.; Castells, A.; Boland, C.R.; Goel, A. MiR-320e is a novel prognostic biomarker in colorectal cancer. Br. J. Cancer 2015, 113, 83–90. [Google Scholar] [CrossRef]
- Lim, S.H.; Park, S.; Jang, J.H.; Kim, K.; Kim, H.-J.; Kim, S.-H.; Kang, C.-I.; Chung, D.R.; Peck, K.R.; Lee, J.; et al. Clinical significance of bone marrow hemophagocytosis in adult patients with malignancy and non-malignancy-induced hemophagocytic lymphohistiocytosis. Ann. Hematol. 2016, 95, 325–335. [Google Scholar] [CrossRef]
- Lim, S.W.; Ryu, K.J.; Lee, H.; Ko, Y.H.; Kim, W.S.; Kim, S.J. Serum IL18 is associated with hemophagocytosis and poor survival in extranodal natural killer/T-cell lymphoma. Leuk. Lymphoma 2019, 60, 317–325. [Google Scholar] [CrossRef]
- Menten, P.; Saccani, A.; Dillen, C.; Wuyts, A.; Struyf, S.; Proost, P.; Mantovani, A.; Wang, J.M.; Van Damme, J. Role of the autocrine chemokines MIP-1alpha and MIP-1beta in the metastatic behavior of murine T cell lymphoma. J. Leukoc. Biol. 2002, 72, 780–789. [Google Scholar] [PubMed]
- Cahir-McFarland, E.D.; Carter, K.; Rosenwald, A.; Giltnane, J.M.; Henrickson, S.E.; Staudt, L.M.; Kieff, E. Role of NF-κB in Cell Survival and Transcription of Latent Membrane Protein 1-Expressing or Epstein-Barr Virus Latency III-Infected Cells. J. Virol. 2004, 78, 4108–4119. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Ryu, K.J.; Ko, Y.H.; Kim, H.-J.; Kim, S.-H.; Kim, W.S.; Kim, S.J. Macrophage inflammatory protein 1 alpha (MIP-1alpha) may be associated with poor outcome in patients with extranodal NK/T-cell lymphoma. Hematol. Oncol. 2017, 35, 310–316. [Google Scholar] [CrossRef]
- Mortezaee, K. CXCL12/CXCR4 axis in the microenvironment of solid tumors: A critical mediator of metastasis. Life Sci. 2020, 249, 117534. [Google Scholar] [CrossRef]
- Nakamura, A.; Rampersaud, Y.R.; Sharma, A.; Lewis, S.J.; Wu, B.; Datta, P.; Sundararajan, K.; Endisha, H.; Rossomacha, E.; Rockel, J.S.; et al. Identification of microRNA-181a-5p and microRNA-4454 as mediators of facet cartilage degeneration. JCI Insight 2016, 1, e86820. [Google Scholar] [CrossRef]
- An, Y.; Yang, Q. MiR-21 modulates the polarization of macrophages and increases the effects of M2 macrophages on promoting the chemoresistance of ovarian cancer. Life Sci. 2020, 242, 117162. [Google Scholar] [CrossRef]
- Sahraei, M.; Chaube, B.; Liu, Y.; Sun, J.; Kaplan, A.; Price, N.L.; Ding, W.; Oyaghire, S.; García-Milian, R.; Mehta, S.; et al. Suppressing miR-21 activity in tumor-associated macrophages promotes an antitumor immune response. J. Clin. Investig. 2019, 129, 5518–5536. [Google Scholar] [CrossRef] [Green Version]
- Zheng, P.; Chen, L.; Yuan, X.; Luo, Q.; Liu, Y.; Xie, G.; Ma, Y.; Shen, L. Exosomal transfer of tumor-associated macrophage-derived miR-21 confers cisplatin resistance in gastric cancer cells. J. Exp. Clin. Cancer Res. 2017, 36, 1–13. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Groups | Poor Outcomes n (%) | Favorable Outcomes n (%) | p |
|---|---|---|---|---|
| Age | ≤60 years | 13 (59) | 20 (87) | 0.047 |
| >60 years | 9 (41) | 3 (13) | ||
| Sex | Male | 13 (59) | 15 (65) | 0.763 |
| Female | 9 (41) | 8 (35) | ||
| Performance status | ECOG 0/1 | 15 (68) | 23 (100) | 0.004 |
| ECOG ≥ 2 | 7 (32) | 0 | ||
| Serum LDH | Normal | 2 (9) | 17 (74) | <0.001 |
| Increased | 20 (91) | 6 (26) | ||
| Stage | I/II | 1 (5) | 23 (100) | <0.001 |
| III/IV | 21 (95) | 0 | ||
| Extranodal involvement | Number 0/1 | 4 (18) | 20 (87) | <0.001 |
| Number ≥ 2 | 18 (82) | 3 (13) | ||
| Bone marrow | Not involved | 8 (36) | 23 (100) | <0.001 |
| Involved | 14 (64) | 0 | ||
| Blood EBV DNA | Not detected | 0 | 23 (100) | <0.001 |
| Detected | 22 (100) | 0 | ||
| PINK-E risk | Low | 0 | 23 (100) | <0.001 |
| Intermediate | 4 (18) | 0 | ||
| High | 18 (82) | 0 | ||
| Primary treatment | SMILE | 11 (50) | 2 (9) | <0.001 |
| VIDL | 5 (23) | 0 | ||
| VIPD | 3 (14) | 0 | ||
| MIDLE | 1 (5) | 0 | ||
| CCRT followed by VIDL | 10 (44) | |||
| CCRT followed by VIPD | 5 (22) | |||
| CCRT followed by MIDLE | 3 (13) | |||
| Other | 1 (5) | 3 (13) | ||
| Relapse or progression | Did not occur | 0 | 17 (74) | <0.001 |
| Occurred | 22 (100) | 6 (26) | ||
| Survival outcome | Alive | 0 | 21 (91) | <0.001 |
| Dead | 22 (100) | 2 (9) |
| Characteristics | miR-4454 | miR-21-5p | miR-320e | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Low n (%) | High n (%) | p | Low n (%) | High n (%) | p | Low n (%) | High n (%) | p | |
| Age | |||||||||
| ≤60 years | 27 (64) | 35 (81) | 0.091 | 30 (71) | 32 (74) | 0.810 | 29 (69) | 33 (77) | 0.471 |
| >60 years | 15 (36) | 8 (19) | 12 (29) | 11 (26) | 13 (31) | 10 (23) | |||
| Sex | |||||||||
| Male | 32 (76) | 27 (63) | 0.240 | 30 (71) | 29 (67) | 0.815 | 27 (64) | 32 (74) | 0.353 |
| Female | 10 (24) | 16 (37) | 12 (29) | 14 (33) | 15 (36) | 11 (26) | |||
| Serum LDH | |||||||||
| Normal | 28 (67) | 19 (44) | 0.050 | 30 (71) | 17 (40) | 0.004 | 25 (60) | 22 (51) | 0.515 |
| Increased | 14 (33) | 24 (56) | 12 (29) | 26 (60) | 17 (40) | 21 (49) | |||
| Stage | |||||||||
| I/II | 29 (69) | 21 (49) | 0.078 | 33 (79) | 17 (40) | <0.001 | 28 (67) | 22 (51) | 0.188 |
| III/IV | 13 (31) | 22 (51) | 9 (21) | 26 (60) | 14 (33) | 21 (49) | |||
| Extranodal involvement | |||||||||
| Number 0/1 | 26 (62) | 22 (51) | 0.384 | 29 (69) | 19 (44) | 0.029 | 27 (64) | 21 (49) | 0.191 |
| Number ≥ 2 | 16 (38) | 21 (49) | 13 (31) | 24 (56) | 15 (36) | 22 (51) | |||
| Bone marrow | |||||||||
| Not involved | 39 (93) | 35 (81) | 0.195 | 39 (93) | 35 (81) | 0.195 | 39 (93) | 35 (81) | 0.195 |
| Involved | 3 (7) | 8 (19) | 3 (7) | 8 (19) | 3 (7) | 8 (19) | |||
| Blood EBV DNA | |||||||||
| Not detected | 20 (48) | 13 (30) | 0.122 | 24 (57) | 9 (21) | 0.001 | 16 (38) | 17 (40) | >0.999 |
| Detected | 22 (52) | 30 (70) | 18 (43) | 34 (79) | 26 (62) | 26 (60) | |||
| PINK-E risk | |||||||||
| Low | 22 (52) | 18 (41) | 0.420 | 26 (62) | 14 (33) | 0.021 | 21 (50) | 19 (44) | 0.146 |
| Intermediate | 9 (21) | 8 (19) | 7 (17) | 10 (23) | 11 (26) | 6 (14) | |||
| High | 11 (26) | 17 (40) | 9 (21) | 19 (44) | 10 (24) | 18 (42) | |||
| Relapse or progression | |||||||||
| Did not occur | 25 (60) | 16 (37) | 0.052 | 27 (64) | 14 (33) | 0.005 | 21 (50) | 20 (47) | 0.829 |
| Occurred | 17 (40) | 27 (63) | 15 (36) | 29 (67) | 21 (50) | 23 (53) | |||
| Survival | |||||||||
| Alive | 30 (71) | 22 (51) | 0.075 | 32 (76) | 20 (47) | 0.007 | 29 (69) | 23 (54) | 0.183 |
| Dead | 12 (29) | 21 (49) | 10 (24) | 23 (53) | 13 (31) | 20 (46) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryu, K.J.; Lee, J.Y.; Choi, M.E.; Yoon, S.E.; Cho, J.; Ko, Y.H.; Shim, J.H.; Kim, W.S.; Park, C.; Kim, S.J. Serum-Derived Exosomal MicroRNA Profiles Can Predict Poor Survival Outcomes in Patients with Extranodal Natural Killer/T-Cell Lymphoma. Cancers 2020, 12, 3548. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12123548
Ryu KJ, Lee JY, Choi ME, Yoon SE, Cho J, Ko YH, Shim JH, Kim WS, Park C, Kim SJ. Serum-Derived Exosomal MicroRNA Profiles Can Predict Poor Survival Outcomes in Patients with Extranodal Natural Killer/T-Cell Lymphoma. Cancers. 2020; 12(12):3548. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12123548
Chicago/Turabian StyleRyu, Kyung Ju, Ji Young Lee, Myung Eun Choi, Sang Eun Yoon, Junhun Cho, Young Hyeh Ko, Joon Ho Shim, Won Seog Kim, Chaehwa Park, and Seok Jin Kim. 2020. "Serum-Derived Exosomal MicroRNA Profiles Can Predict Poor Survival Outcomes in Patients with Extranodal Natural Killer/T-Cell Lymphoma" Cancers 12, no. 12: 3548. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12123548