Serum Type XIX Collagen is Significantly Elevated in Non-Small Cell Lung Cancer: A Preliminary Study on Biomarker Potential

 , ,
, ,
Abstract
:1. Introduction
2. Results
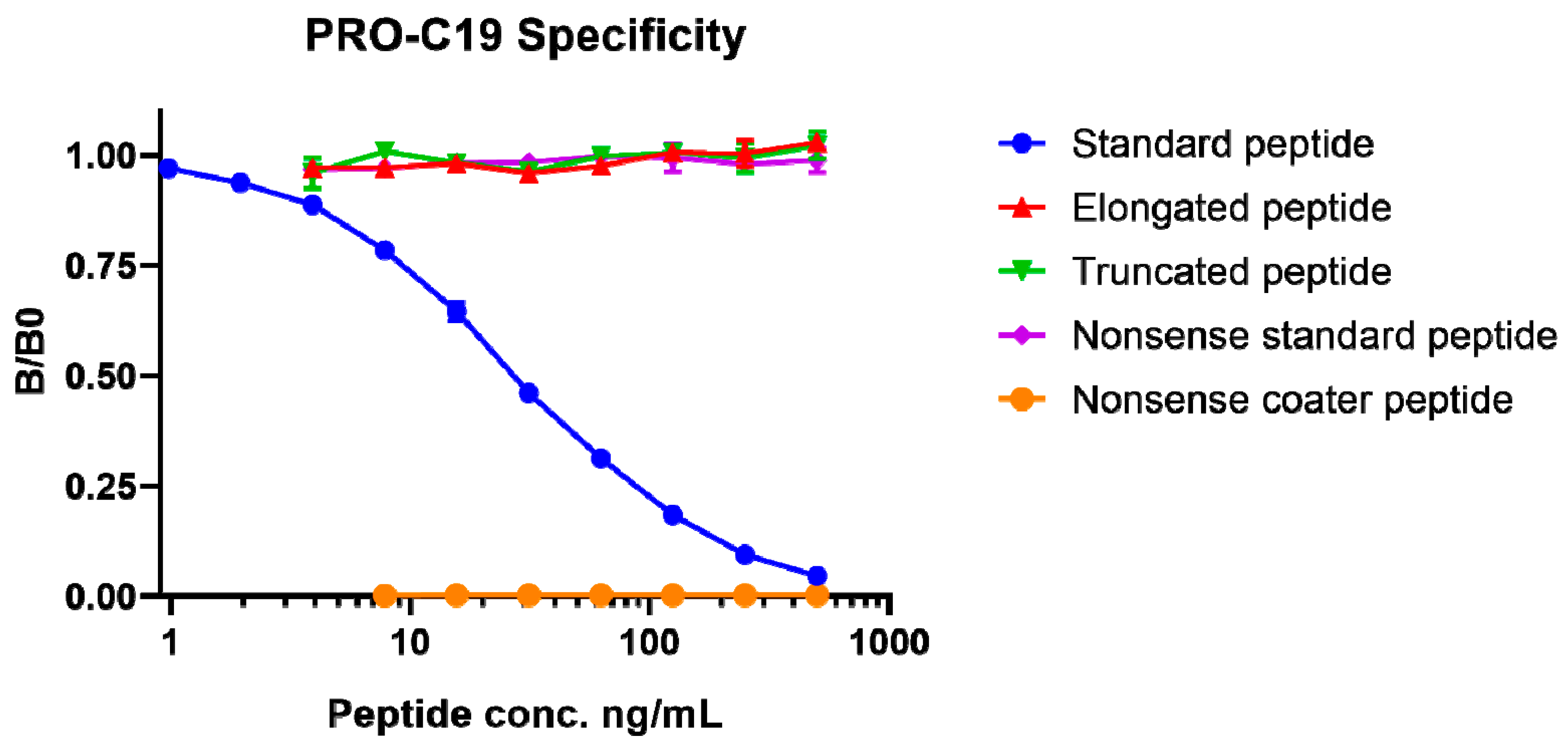
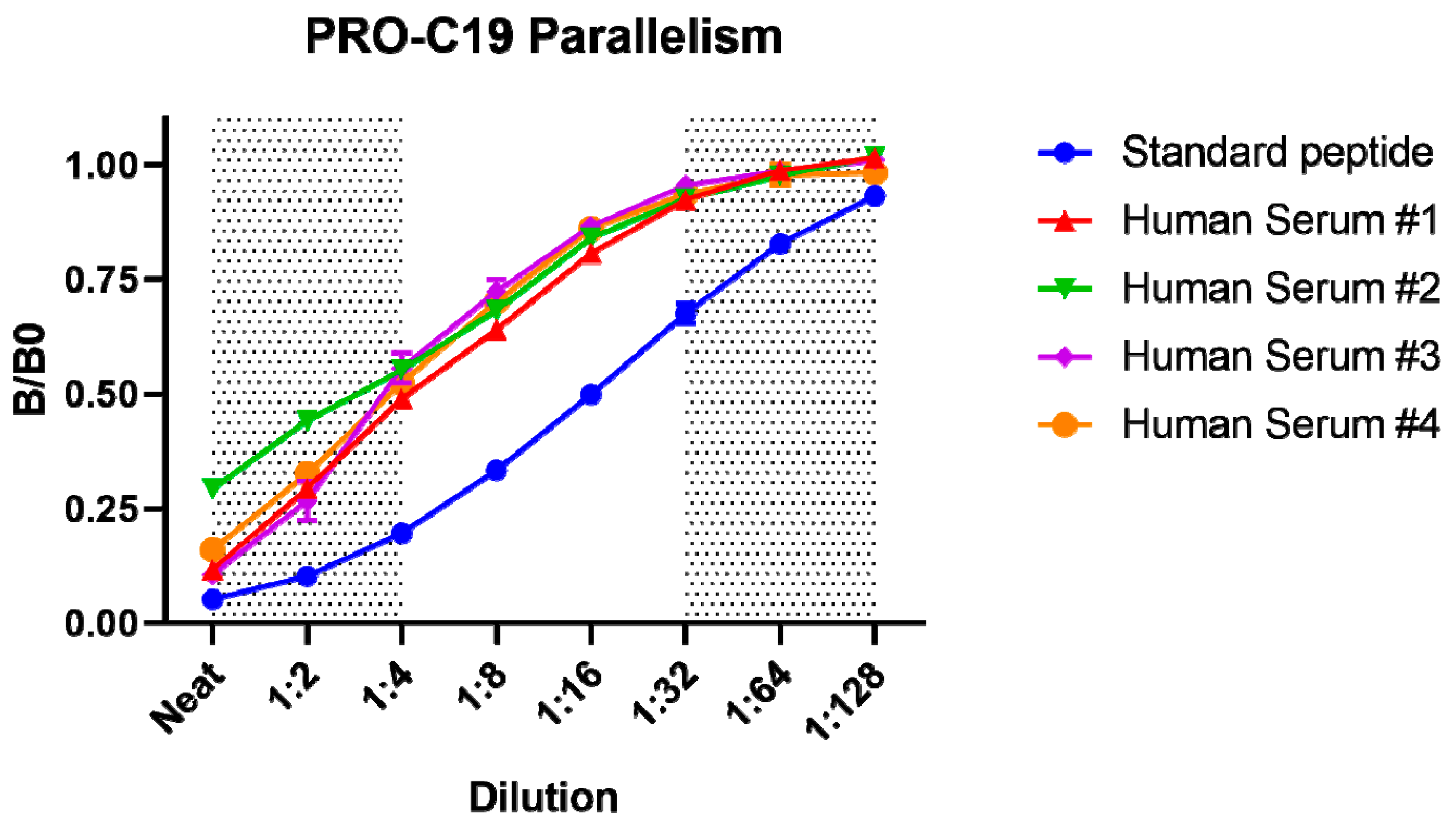
2.1. PRO-C19 ELISA Development
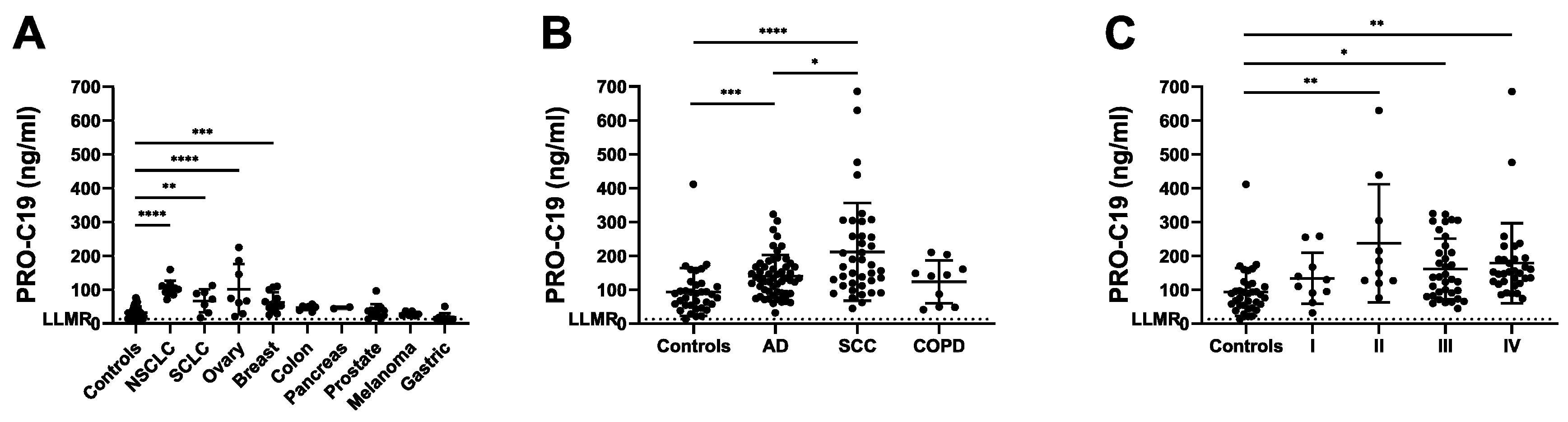
2.2. PRO-C19 in Serum of Cancer Patients (Cohort 1)
2.3. PRO-C19 in Serum of NSCLC Patients (Cohort 2)
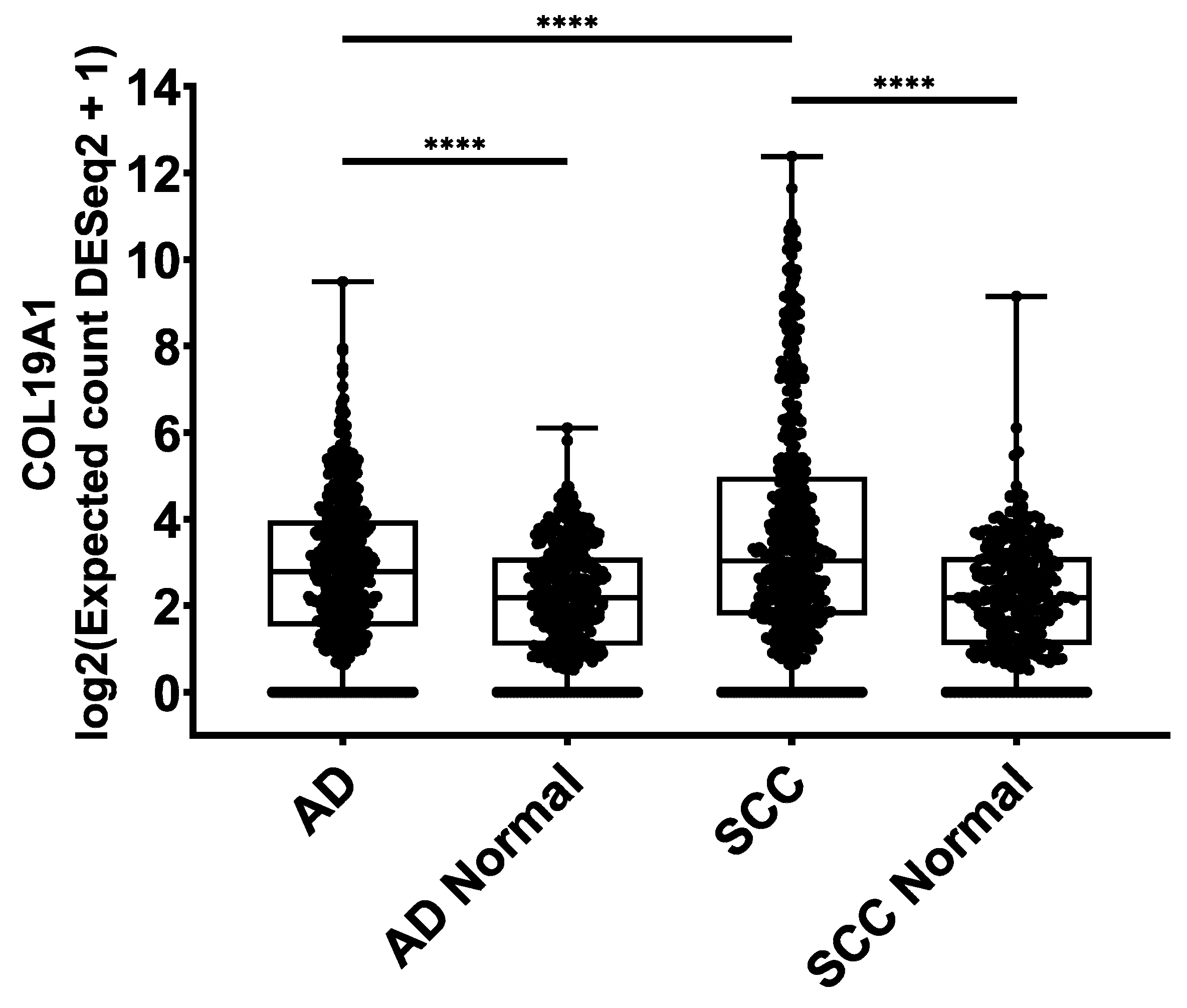
2.4. COL19A1 Gene Expression in Publicly Available Lung Cancer Databases
3. Discussion
4. Materials and Methods
4.1. PRO-C19 ELISA Protocol
4.2. Technical Validation of the PRO-C19 ELISA
4.3. Patient Samples
4.4. Analysis of Publicly Available Genomics Databases
4.5. Statistics:
5. Conclusions
6. Patents
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Molina, J.R.; Yang, P.; Cassivi, S.D.; Schild, S.E.; Adjei, A.A. Non-Small Cell Lung Cancer: Epidemiology, Risk Factors, Treatment, and Survivorship. Mayo Clin. Proc. 2008, 83, 584–594. [Google Scholar] [CrossRef]
- Herbst, R.S.; Morgensztern, D.; Boshoff, C. The biology and management of non-small cell lung cancer. Nature 2018, 553, 446–454. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA A Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.A.; Weinberg, R. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Pickup, M.W.; Mouw, J.K.; Weaver, V.M. The extracellular matrix modulates the hallmarks of cancer. EMBO Rep. 2014, 15, 1243–1253. [Google Scholar] [CrossRef] [Green Version]
- Ricard-Blum, S. The collagen family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef] [Green Version]
- Willumsen, N.; Bager, C.L.; Leeming, D.J.; Smith, V.; Karsdal, M.A.; Dornan, D.; Bay-Jensen, A.-C. Extracellular matrix specific protein fingerprints measured in serum can separate pancreatic cancer patients from healthy controls. BMC Cancer 2013, 13, 554. [Google Scholar] [CrossRef] [Green Version]
- Willumsen, N.; Bager, C.L.; Leeming, D.J.; Smith, V.; Christiansen, C.; Karsdal, M.A.; Dornan, D.; Bay-Jensen, A.-C. Serum biomarkers reflecting specific tumor tissue remodeling processes are valuable diagnostic tools for lung cancer. Cancer Med. 2014, 3, 1136–1145. [Google Scholar] [CrossRef] [PubMed]
- Lipton, A.; Leitzel, K.; Ali, S.M.; Polimera, H.V.; Nagabhairu, V.; Marks, E.; Richardson, A.E.; Krecko, L.; Ali, A.; Koestler, W.; et al. High turnover of extracellular matrix reflected by specific protein fragments measured in serum is associated with poor outcomes in two metastatic breast cancer cohorts. Int. J. Cancer 2018, 143, 3027–3034. [Google Scholar] [CrossRef] [PubMed]
- Thorlacius-Ussing, J.; Kehlet, S.N.; Rønnow, S.R.; Karsdal, M.A.; Willumsen, N. Non-invasive profiling of protease-specific elastin turnover in lung cancer: Biomarker potential. J. Cancer Res. Clin. Oncol. 2018, 145, 383–392. [Google Scholar] [CrossRef]
- Jensen, C.; Madsen, D.H.; Hansen, M.; Schmidt, H.; Svane, I.M.; Karsdal, M.A.; Willumsen, N. Non-invasive biomarkers derived from the extracellular matrix associate with response to immune checkpoint blockade (anti-CTLA-4) in metastatic melanoma patients. J. Immunother. Cancer 2018, 6, 152. [Google Scholar] [CrossRef] [Green Version]
- Willumsen, N.; Jørgensen, L.N.; Karsdal, M.A. Vastatin (the NC1 domain of human type VIII collagen a1 chain) is linked to stromal reactivity and elevated in serum from patients with colorectal cancer. Cancer Boil. Ther. 2019, 20, 692–699. [Google Scholar] [CrossRef]
- Willumsen, N.; Bager, C.A.; Karsdal, M. Matrix Metalloprotease Generated Fragments of Type VI Collagen Have Serum Biomarker Potential in Cancer – A Proof of Concept Study. Transl. Oncol. 2019, 12, 693–698. [Google Scholar] [CrossRef]
- Banys-Paluchowski, M.; Loibl, S.; Witzel, I.; Mundhenke, C.; Lederer, B.; Solbach, C.; Karn, T.; Marmé, F.; Nekljudova, V.; Schem, C.; et al. Clinical Relevance of Collagen Protein Degradation Markers C3M and C4M in the Serum of Breast Cancer Patients Treated with Neoadjuvant Therapy in the GeparQuinto Trial. Cancers 2019, 11, 1186. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.M.; Willumsen, N.; Dehlendorff, C.; Johansen, A.Z.; Jensen, B.V.; Hansen, C.P.; Hasselby, J.P.; Bojesen, S.E.; Pfeiffer, P.; Nielsen, S.E.; et al. Clinical value of serum hyaluronan and propeptide of type III collagen in patients with pancreatic cancer. Int. J. Cancer 2019, 146, 2913–2922. [Google Scholar] [CrossRef] [PubMed]
- Willumsen, N.; Ali, S.M.; Leitzel, K.; Drabick, J.J.; Yee, N.; Polimera, H.V.; Nagabhairu, V.; Krecko, L.; Ali, A.; Maddukuri, A.; et al. Collagen fragments quantified in serum as measures of desmoplasia associate with survival outcome in patients with advanced pancreatic cancer. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, H.; Zhang, H.; Ramirez, F.; Mattei, M.-G.; Moradi-Améli, M.; Van Der Rest, M.; Gordon, M.K. Synteny between the loci for a novel FACIT-like collagen locus (D6S228E) and α1(IX) collagen (COL9A1) on 6q12–q14 in humans. Genomics 1992, 13, 884–886. [Google Scholar] [CrossRef]
- Inoguchi, K.; Yoshioka, H.; Khaleduzzaman, M.; Ninomiya, Y. The mRNA for α1(XIX) Collagen Chain, a New Member of FACITs, Contains a Long Unusual 3′ Untranslated Region and Displays Many Unique Splicing Variants1. J. Biochem. 1995, 117, 137–146. [Google Scholar] [CrossRef]
- Myers, J.C.; Yang, H.A.; D’Ippolito, J.; Presente, A.; Miller, M.K.; Dion, A.S. The triple-helical region of human type XIX collagen consists of multiple collagenous subdomains and exhibits limited sequence homology to alpha 1(XVI). J. Boil. Chem. 1994, 269, 18549–18557. [Google Scholar]
- Myers, J.C.; Li, D.; Bageris, A.; Abraham, V.; Dion, A.S.; Amenta, P.S. Biochemical and immunohistochemical characterization of human type XIX defines a novel class of basement membrane zone collagens. Am. J. Pathol. 1997, 151, 1729–1740. [Google Scholar]
- Amenta, P.S.; Hadad, S.; Lee, M.T.; Barnard, N.; Li, D.; Myers, J.C. Loss of types XV and XIX collagen precedes basement membrane invasion in ductal carcinoma of the female breast. J. Pathol. 2003, 199, 298–308. [Google Scholar] [CrossRef]
- Ramont, L.; Brassart-Pasco, S.; Thevenard, J.; Deshorgue, A.; Venteo, L.; Laronze, J.Y.; Pluot, M.; Monboisse, J.C.; Maquart, F.-X. The NC1 domain of type XIX collagen inhibits in vivo melanoma growth. Mol. Cancer Ther. 2007, 6, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Oudart, J.-B.; Brassart-Pasco, S.; Vautrin, A.; Sellier, C.; Machado, C.; Dupont-Deshorgue, A.; Brassart, B.; Baud, S.; Dauchez, M.; Monboisse, J.-C.; et al. Plasmin releases the anti-tumor peptide from the NC1 domain of collagen XIX. Oncotarget 2015, 6, 3656–3668. [Google Scholar] [CrossRef]
- Oudart, J.-B.; Doué, M.; Vautrin, A.; Brassart, B.; Sellier, C.; Dupont-Deshorgue, A.; Monboisse, J.-C.; Maquart, F.-X.; Brassart-Pasco, S.; Ramont, L. The anti-tumor NC1 domain of collagen XIX inhibits the FAK/ PI3K/Akt/mTOR signaling pathway through αvβ3 integrin interaction. Oncotarget 2015, 7, 1516–1528. [Google Scholar] [CrossRef] [Green Version]
- The Cancer Genome Atlas Program. Available online: https://www.cancer.gov/tcga (accessed on 29 May 2020).
- The Genotype-Tissue Expression (GTEx) Project. Available online: https://www.gtexportal.org (accessed on 29 May 2020).
- Sumiyoshi, H.; Inoguchi, K.; Khaleduzzaman, M.; Ninomiya, Y.; Yoshioka, H. Ubiquitous Expression of the α1(XIX) Collagen Gene (Col19a1) during Mouse Embryogenesis Becomes Restricted to a Few Tissues in the Adult Organism. J. Boil. Chem. 1997, 272, 17104–17111. [Google Scholar] [CrossRef] [Green Version]
- Sedgwick, A.E.; D’Souza-Schorey, C. Wnt Signaling in Cell Motility and Invasion: Drawing Parallels between Development and Cancer. Cancers 2016, 8, 80. [Google Scholar] [CrossRef] [Green Version]
- Giroux-Leprieur, E.; Costantini, A.; Ding, V.W.; He, B. Hedgehog Signaling in Lung Cancer: From Oncogenesis to Cancer Treatment Resistance. Int. J. Mol. Sci. 2018, 19, 2835. [Google Scholar] [CrossRef] [Green Version]
- Chaffer, C.L.; Juan, B.P.S.; Lim, E.A.; Weinberg, R. EMT, cell plasticity and metastasis. Cancer Metastasis Rev. 2016, 35, 645–654. [Google Scholar] [CrossRef]
- Kehlet, S.; Manon-Jensen, T.; Sun, S.; Brix, S.; Leeming, D.; Karsdal, M.A.; Willumsen, N. A fragment of SPARC reflecting increased collagen affinity shows pathological relevance in lung cancer – implications of a new collagen chaperone function of SPARC. Cancer Boil. Ther. 2018, 19, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Nissen, N.I.; Karsdal, M.; Willumsen, N. Post-translational modifications of vimentin reflect different pathological processes associated with non-small cell lung cancer and chronic obstructive pulmonary disease. Oncotarget 2019, 10, 6829–6841. [Google Scholar] [CrossRef]
- Leeming, D.J.; Willumsen, N.; Sand, J.; Nielsen, S.H.; Dasgupta, B.; Brodmerkel, C.; Curran, M.; Bager, C.; Karsdal, M.A. A serological marker of the N-terminal neoepitope generated during LOXL2 maturation is elevated in patients with cancer or idiopathic pulmonary fibrosis. Biochem. Biophys. Rep. 2018, 17, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.H.; Willumsen, N.; Leeming, D.J.; Daniels, S.J.; Brix, S.; Karsdal, M.A.; Genovese, F.; Nielsen, M.J. Serological Assessment of Activated Fibroblasts by alpha-Smooth Muscle Actin (α-SMA): A Noninvasive Biomarker of Activated Fibroblasts in Lung Disorders. Transl. Oncol. 2018, 12, 368–374. [Google Scholar] [CrossRef]
- Nielsen, S.H.; Willumsen, N.; Brix, S.; Sun, S.; Manon-Jensen, T.; Karsdal, M.; Genovese, F. Tumstatin, a Matrikine Derived from Collagen Type IVα3, is Elevated in Serum from Patients with Non–Small Cell Lung Cancer. Transl. Oncol. 2018, 11, 528–534. [Google Scholar] [CrossRef]
- Kehlet, S.; Bager, C.; Willumsen, N.; Dasgupta, B.; Brodmerkel, C.; Curran, M.; Brix, S.; Leeming, D.J.; Karsdal, M.A. Cathepsin-S degraded decorin are elevated in fibrotic lung disorders – development and biological validation of a new serum biomarker. BMC Pulm. Med. 2017, 17, 110. [Google Scholar] [CrossRef] [Green Version]
- Willumsen, N.; Bager, C.L.; Leeming, D.J.; Bay-Jensen, A.-C.; Karsdal, M.A. Nidogen-1 Degraded by Cathepsin S can be Quantified in Serum and is Associated with Non-Small Cell Lung Cancer. Neoplasia 2017, 19, 271–278. [Google Scholar] [CrossRef]
- Myers, J.C.; Li, D.; Amenta, P.S.; Clark, C.C.; Nagaswami, C.; Weisel, J.W. Type XIX Collagen Purified from Human Umbilical Cord Is Characterized by Multiple Sharp Kinks Delineating Collagenous Subdomains and by Intermolecular Aggregates via Globular, Disulfide-linked, and Heparin-binding Amino Termini. J. Boil. Chem. 2003, 278, 32047–32057. [Google Scholar] [CrossRef] [Green Version]
- Oudart, J.; Brassart-Pasco, S.; Luczka, E.; Dupont-Deshorgue, A.; Bellon, G.; Boudko, S.; Bachinger, H.; Monboisse, J.C.; Maquart, F.-X.; Ramont, L. Analytical methods for measuring collagen XIX in human cell cultures, tissue extracts, and biological fluids. Anal. Biochem. 2013, 437, 111–117. [Google Scholar] [CrossRef]
- Määttä, M.; Virtanen, I.; Burgeson, R.; Autio–Harmainen, H. Comparative Analysis of the Distribution of Laminin Chains in the Basement Membranes in Some Malignant Epithelial Tumors: The ?1 Chain of Laminin Shows a Selected Expression Pattern in Human Carcinomas. J. Histochem. Cytochem. 2001, 49, 711–725. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Chen, J.; Lippold, K.; Monavarfeshani, A.; Carrillo, G.L.; Jenkins, R.A.; Fox, M. Collagen-derived matricryptins promote inhibitory nerve terminal formation in the developing neocortex. J. Cell Boil. 2016, 212, 721–736. [Google Scholar] [CrossRef]
- Caccavari, F.; Valdembri, D.; Sandri, C.; Bussolino, F.; Serini, G.G. Integrin signaling and lung cancer. Cell Adhes. Migr. 2010, 4, 124–129. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.; Fu, C.; Bhattacharya, J. Vascular expression of the αvβ3-integrin in lung and other organs. Am. J. Physiol. Cell. Mol. Physiol. 2000, 278, L217–L226. [Google Scholar] [CrossRef] [Green Version]
- Schaffner, F.; Ray, A.-M.; Dontenwill, M. Integrin α5β1, the Fibronectin Receptor, as a Pertinent Therapeutic Target in Solid Tumors. Cancers 2013, 5, 27–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Infante, J.; Prieto, C.; Sierra, M.; Sánchez-Juan, P.; González-Aramburu, I.; Sánchez-Quintana, C.; Berciano, J.; Combarros, O.; Sainz, J. Identification of candidate genes for Parkinson’s disease through blood transcriptome analysis in LRRK2-G2019S carriers, idiopathic cases, and controls. Neurobiol. Aging 2015, 36, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Shtilbans, A.; Choi, S.G.; Fowkes, M.E.; Khitrov, G.; Shahbazi, M.; Ting, J.; Zhang, W.; Sun, Y.; Sealfon, S.C.; Lange, D.J. Differential gene expression in patients with amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. 2011, 12, 250–256. [Google Scholar] [CrossRef]
- Calvo, A.; Manzano, R.; Atencia-Cibreiro, G.; Oliván, S.; Muñoz, M.J.; Zaragoza, P.; Cordero-Vázquez, P.; Esteban-Pérez, J.; García-Redondo, A.; Osta, R. Genetic Biomarkers for ALS Disease in Transgenic SOD1G93A Mice. PLoS ONE 2012, 7, e32632. [Google Scholar] [CrossRef] [Green Version]
- Calvo, A.; Cibreiro, G.A.; Merino, P.T.; Roy, J.F.; Galiana, A.; Rufián, A.J.; Cano, J.M.; Martínez, L.M.; Moreno, L.; Larrodé, P.; et al. Collagen XIX Alpha 1 Improves Prognosis in Amyotrophic Lateral Sclerosis. Aging Dis. 2019, 10, 278–292. [Google Scholar] [CrossRef] [Green Version]
- Neergaard, J.; Dragsbæk, K.; Kehlet, S.; Hansen, H.; Hansen, G.; Byrjalsen, I.; Alexandersen, P.; Lindgren, L.; Bihlet, A.; Riis, B.; et al. Cohort Profile: The Prospective Epidemiological Risk Factor (PERF) study. Int. J. Epidemiol. 2016, 46, 1104. [Google Scholar] [CrossRef] [Green Version]
- Goldman, M.; Craft, B.; Hastie, M.; Repečka, K.; Kamath, A.; McDade, F.; Rogers, D.; Brooks, A.N.; Zhu, J.; Haussler, D.; et al. The UCSC Xena platform for public and private cancer genomics data visualization and interpretation 2018, 326470. BioRxiv 2018, 326470. [Google Scholar]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Vivian, J.; Rao, A.; Nothaft, F.A.; Ketchum, C.; Armstrong, J.; Novak, A.M.; Pfeil, J.; Narkizian, J.; DeRan, A.D.; Musselman-Brown, A.; et al. Toil enables reproducible, open source, big biomedical data analyses. Nat. Biotechnol. 2017, 35, 314–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Boil. 2014, 15, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test | Result |
|---|---|
| IC50 | 29.5 ng/mL |
| Measurement range | 3.31–214 ng/Ll |
| Detection range | 1.23–443 ng/L |
| Minimum required dilution in human serum | 1:4 |
| Dilution recovery of human serum below 1:4 | 102% |
| Spiking recovery of peptide in serum | 119% |
| Interference hemoglobin, low/high conc. | 95.1%/93.4% |
| Interference lipids, low/high conc. | 104%/104% |
| Interference biotin, low/high conc. | 101%/101% |
| Inter-assay variation | 10.9% |
| Intra-assay variation | 6.63% |
| Analyte stability (24 hrs 4 °C/4 hrs 20 °C) | 85.3%/82.3% |
| Freeze-thaw stability up to four cycles | 97.7% |
| Cohort 1 | Healthy Controls (n = 38) | Cancers (n = 75) | p-Value | ||
| Age, median (min-max) | 71.5 (60–82) | 60 (30–84) | <0.0001 | ||
| Male, n (%) | 0 (0.00%) | 39 (52.0%) | <0.0001 | ||
| Tumour type and stage, n | - | Breast cancer (n = 12), II:10, III:2 | - | ||
| Colon cancer (n = 7), II:3, III:4 | |||||
| Gastric cancer (n = 9), I:4, II:1, III:3, IV:1 | |||||
| Melanoma (n = 6), II:5, III:1 | |||||
| NSCLC (n = 11), I:5, II:3, III:3 | |||||
| Ovarian cancer (n = 8), I:1, II:2, III:5 | |||||
| Pancreatic cancer (n = 2), Not available | |||||
| Prostate cancer (n = 13), I:1, II:12 | |||||
| SCLC (n = 7), I:2, II:1, III:3, IV:1 | |||||
| Cohort 2 | Healthy controls (n = 35) | AD (n = 55) | SCC (n = 39) | COPD (n = 10) | p-Value |
| Age, median (min–max) | 60 (51–72) | 62 (41–78) | 62 (50–71) | 54 (50–60) | <0.0001 |
| Male, n (%) | 21 (60%) | 21 (42%) | 27 (73%) | 5 (50%) | 0.0339 |
| Tumour stage, n | - | I:5, II:4, III:22, IV:19 | I:5, II:6, III:13, IV:13 | - | - |
| Test | AUROC (95%CI) | p-Value | Sensitivity, % (95%CI) | Specificity, % (95%CI) | Cut-Off (ng/mL) |
|---|---|---|---|---|---|
| Cohort 1 | |||||
| Controls v NSCLC | 0.995 (0.918 – 1.00) | <0.0001 | 100 (71.5 – 100.0) | 94.74 (82.3 – 99.4) | 63.3 |
| Controls v SCLC | 0.808 (0.663 – 0.910) | 0.0048 | 71.4 (29.0 – 96.3) | 84.2 (68.7 – 94.0) | 54.3 |
| Controls v breast cancer | 0.814 (0.678 – 0.910) | <0.0001 | 75.0 (42.8 – 94.5) | 78.95 (62.7 – 90.4) | 41.9 |
| Controls v ovarian cancer | 0.839 (0.701 – 0.931) | 0.0003 | 75.0 (34.9 – 96.8) | 92.1 (78.6 – 98.3) | 60.3 |
| Cohort 2 | |||||
| Controls v AD+SCC | 0.781 (0.700 – 0.849) | <0.0001 | 73.4 (63.3 – 82.0) | 74.29 (56.7 – 87.5) | 108.5 |
| Controls v AD | 0.749 (0.647 – 0.835) | <0.0001 | 69.09 (55.2 – 80.9) | 74.29 (56.7 – 87.5) | 108.5 |
| Controls v SCC | 0.826 (0.721 – 0.905) | <0.0001 | 71.79 (55.1 – 85.0) | 82.86 (66.4 – 93.4) | 124.1 |
| AD vs SCC | 0.654 (0.548 – 0.749) | 0.0096 | 41.03 (25.6 – 57.9) | 87.27 (75.5 – 94.7) | 194.0 |
| Controls v COPD | 0.646 (0.489 – 0.782) | 0.1926 | 60.00 (26.2 – 87.8) | 82.86 (66.4 – 93.4) | 124.1 |
| AD vs COPD | 0.556 (0.428 – 0.680) | 0.6251 | 30.00 (6.7 – 65.2) | 98.18 (90.3 – 100.0) | 49.1 |
| SCC vs COPD | 0.690 (0.542 – 0.814) | 0.0377 | 100.00 (69.2 – 100.0) | 35.90 (21.2 – 52.8) | 210.2 |
| Controls v AD I+II | 0.733 (0.579 – 0.855) | 0.0197 | 88.89 (51.8 – 99.7) | 57.14 (39.4 – 73.7) | 89.1 |
| Controls v AD III+IV | 0.756 (0.644 – 0.847) | <0.0001 | 68.29 (51.9 – 81.9) | 77.14 (59.9 – 89.6) | 113.9 |
| Controls v SCC I+II | 0.818 (0.677 – 0.916) | <0.0001 | 81.82 (48.2 – 97.7) | 80.00 (63.1 – 91.6) | 118.9 |
| Controls v SCC III+IV | 0.833 (0.716 – 0.916) | <0.0001 | 73.08 (52.2 – 88.4) | 82.86 (66.4 – 93.4) | 124.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thorlacius-Ussing, J.; Manon-Jensen, T.; Sun, S.; Leeming, D.J.; Sand, J.M.; Karsdal, M.; Willumsen, N. Serum Type XIX Collagen is Significantly Elevated in Non-Small Cell Lung Cancer: A Preliminary Study on Biomarker Potential. Cancers 2020, 12, 1510. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12061510
Thorlacius-Ussing J, Manon-Jensen T, Sun S, Leeming DJ, Sand JM, Karsdal M, Willumsen N. Serum Type XIX Collagen is Significantly Elevated in Non-Small Cell Lung Cancer: A Preliminary Study on Biomarker Potential. Cancers. 2020; 12(6):1510. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12061510
Chicago/Turabian StyleThorlacius-Ussing, Jeppe, Tina Manon-Jensen, Shu Sun, Diana J. Leeming, Jannie M. Sand, Morten Karsdal, and Nicholas Willumsen. 2020. "Serum Type XIX Collagen is Significantly Elevated in Non-Small Cell Lung Cancer: A Preliminary Study on Biomarker Potential" Cancers 12, no. 6: 1510. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12061510