
KYP-2047, an Inhibitor of Prolyl-Oligopeptidase, Reduces GlioBlastoma Proliferation through Angiogenesis and Apoptosis Modulation

 , , , , , and
, , , , , and 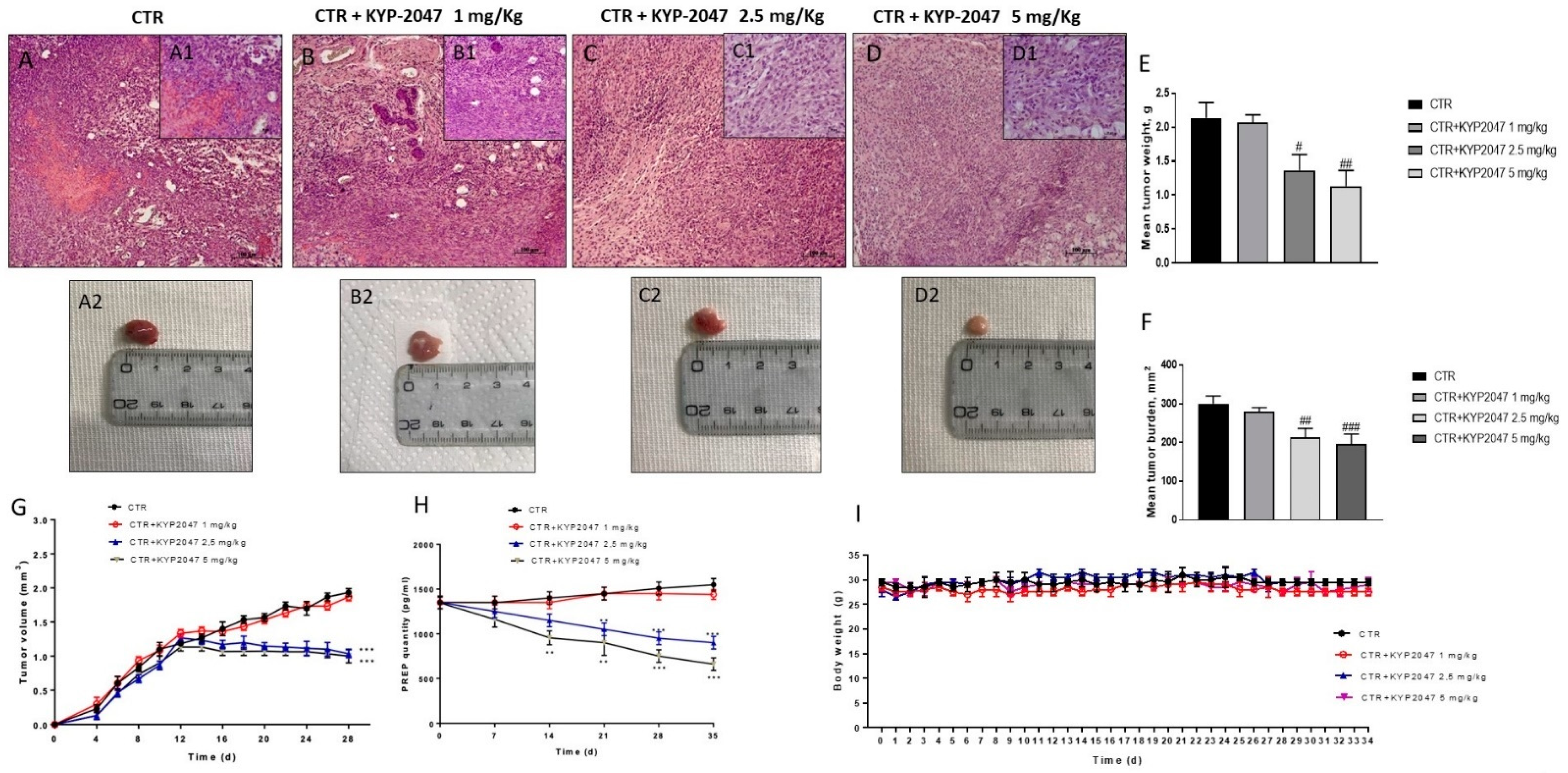{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. In Vivo Studies
2.1.1. Cell Line
2.1.2. Animals
2.1.3. Experimental Design
- Control group (vehicle): weekly intravenous (IV) administration of saline.
- Control group + KYP-2047 1 mg/kg: intraperitoneal (ip) administration of KYP-2047 1 mg/kg dissolved in PBS every three days from day 7.
- Control group + KYP-2047 2.5 mg/kg: intraperitoneal (ip) administration of KYP-2047 2.5 mg/kg dissolved in PBS every three days from day 7.
- Control group + KYP-2047 5 mg/kg: intraperitoneal (ip) administration of KYP-2047 5 mg/kg dissolved in PBS every three days from day 7.
2.1.4. Histological Evaluation
2.1.5. Western Blot Analysis
2.1.6. Immunohistochemical Localization of Vascular Endothelial-Growth-Factor (VEGF), Endothelial Nitric Oxide Synthase (eNOS), CD34, Ki-67, Bcl2 and Caspase-3
2.1.7. Caspase-3 Activity Measurement
2.1.8. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
2.1.9. Enzyme-Linked Immunosorbent Assay (ELISA) Kit
2.1.10. Immunofluorescence Assay
2.2. In Vitro Studies
2.2.1. Cell Lines
2.2.2. Cell Treatment
- Control group (Ctr): human GB cell lines U-87, U-138 and A-172;
- KYP-2047 0.01 μM group: GB cells treated with KYP-2047 0.01 μM;
- KYP-2047 0.1 μM group: GB cells treated with KYP-2047 0.1 μM;
- KYP-2047 0.5 μM group: GB cells treated with KYP-2047 0.5 μM;
- KYP-2047 1 μM group: GB cells treated with KYP-2047 1 μM;
- KYP-2047 10 μM group: GB cells treated with KYP-2047 10 μM;
- KYP-2047 30 μM group: GB cells treated with KYP-2047 30 μM;
- KYP-2047 50 μM group: GB cells treated with KYP-2047 50 μM;
- KYP-2047 100 μM group: GB cells treated with KYP-2047 100 μM;
2.2.3. Cell Viability Assay
2.2.4. Western Blot Analysis
2.2.5. Immunofluorescence Assay for Transforming Growth Factor-β (TGF-β) and Caspase-3
2.3. Materials
2.4. Statistical Analysis
3. Results
3.1. In Vivo Studies
3.1.1. Effect of KYP-2047 on Tumor Growth
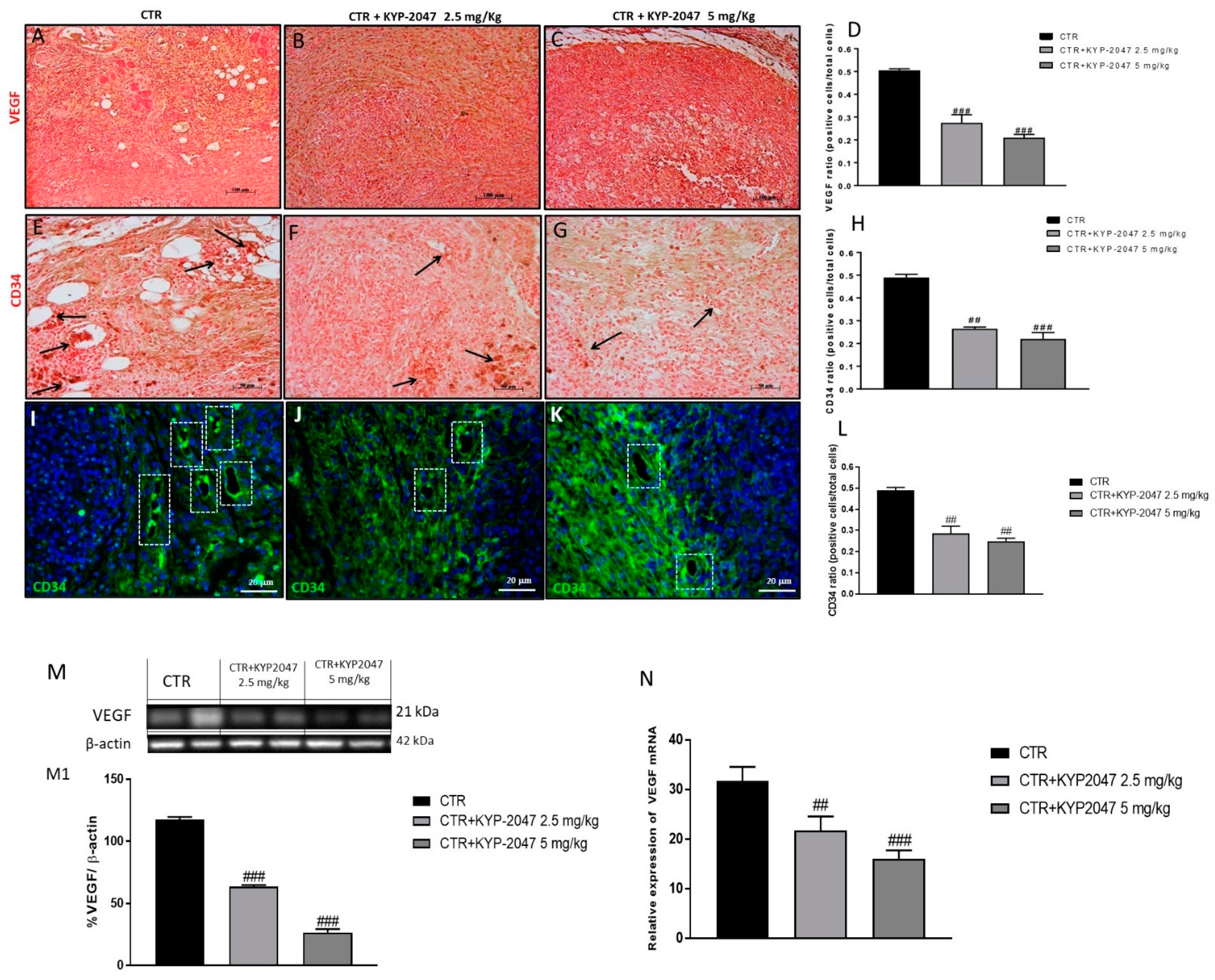
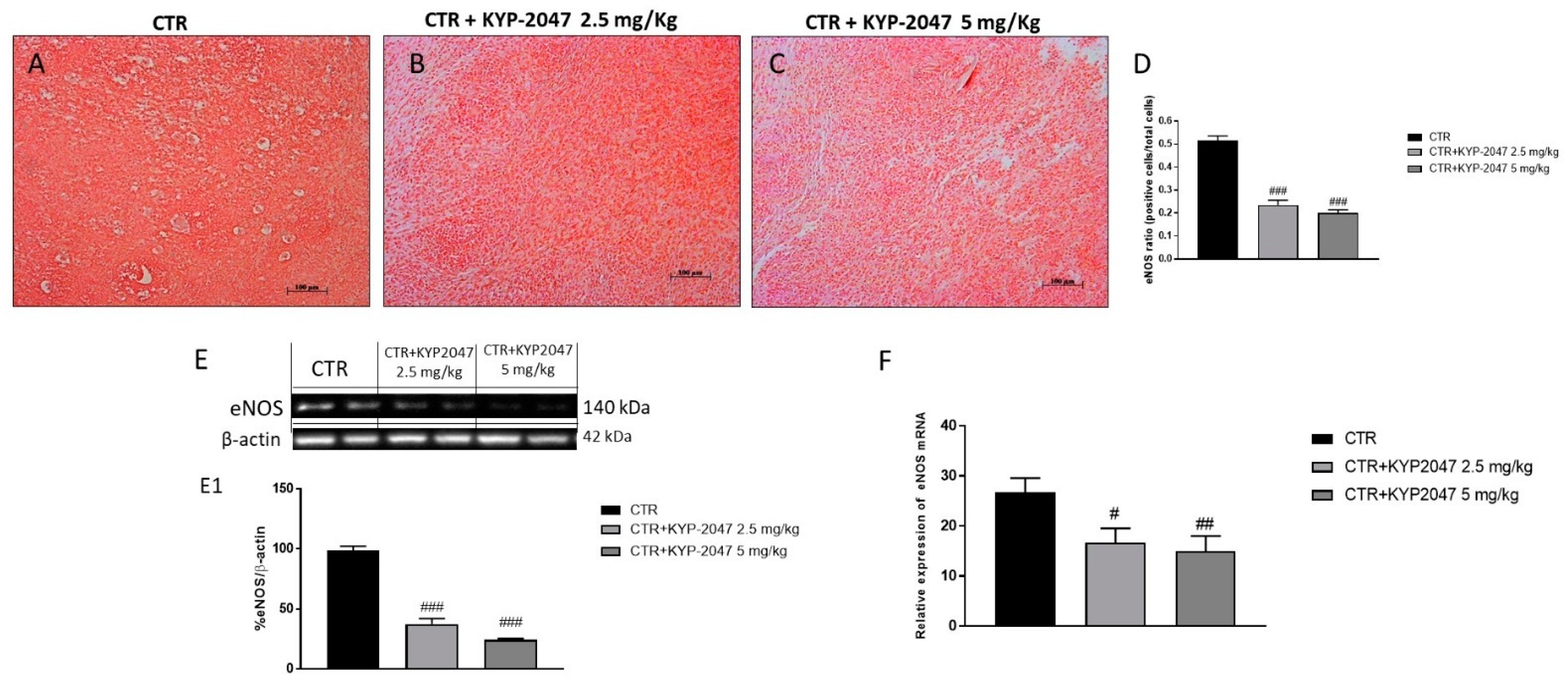
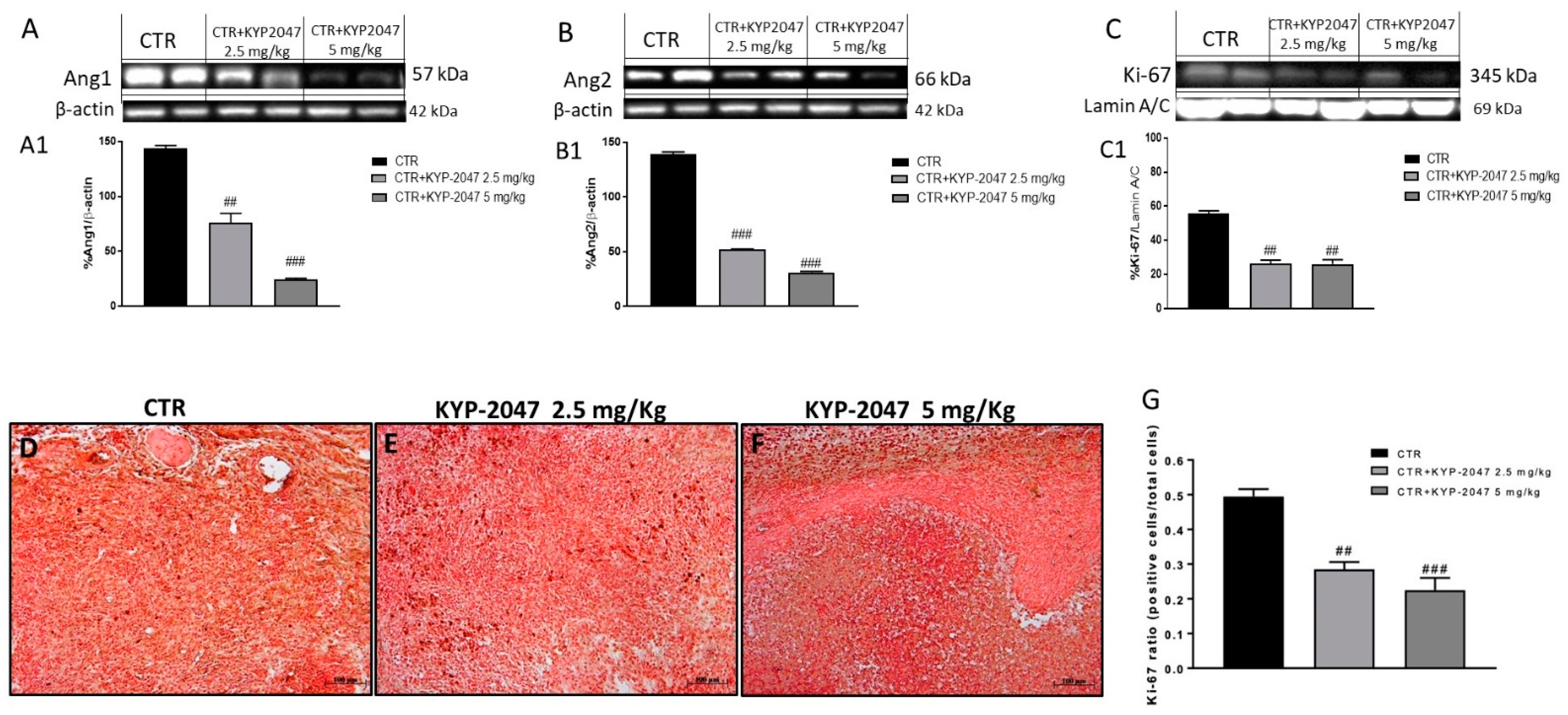
3.1.2. Effect of KYP-2047 on Angiogenesis
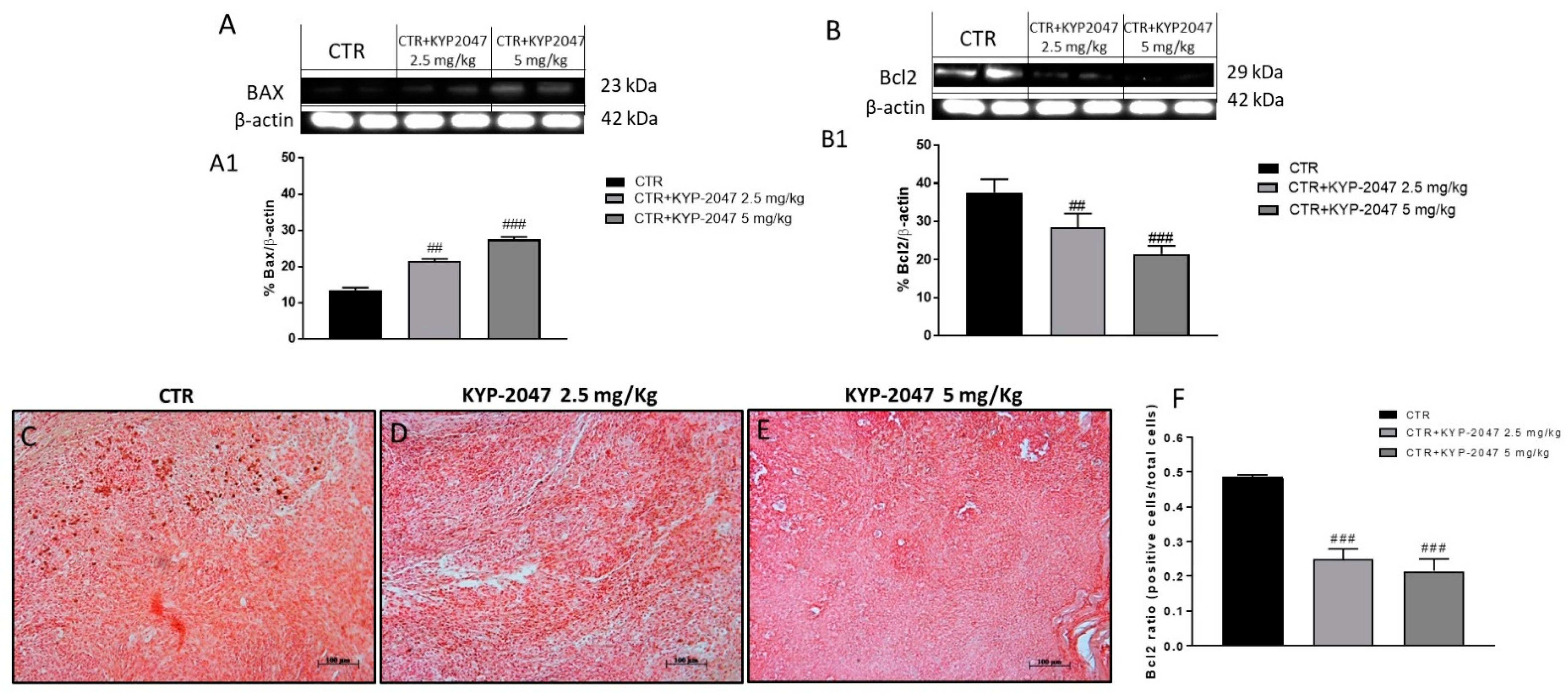
3.1.3. Effect of KYP-2047 on Apoptosis Pathway
3.2. In Vitro Studies
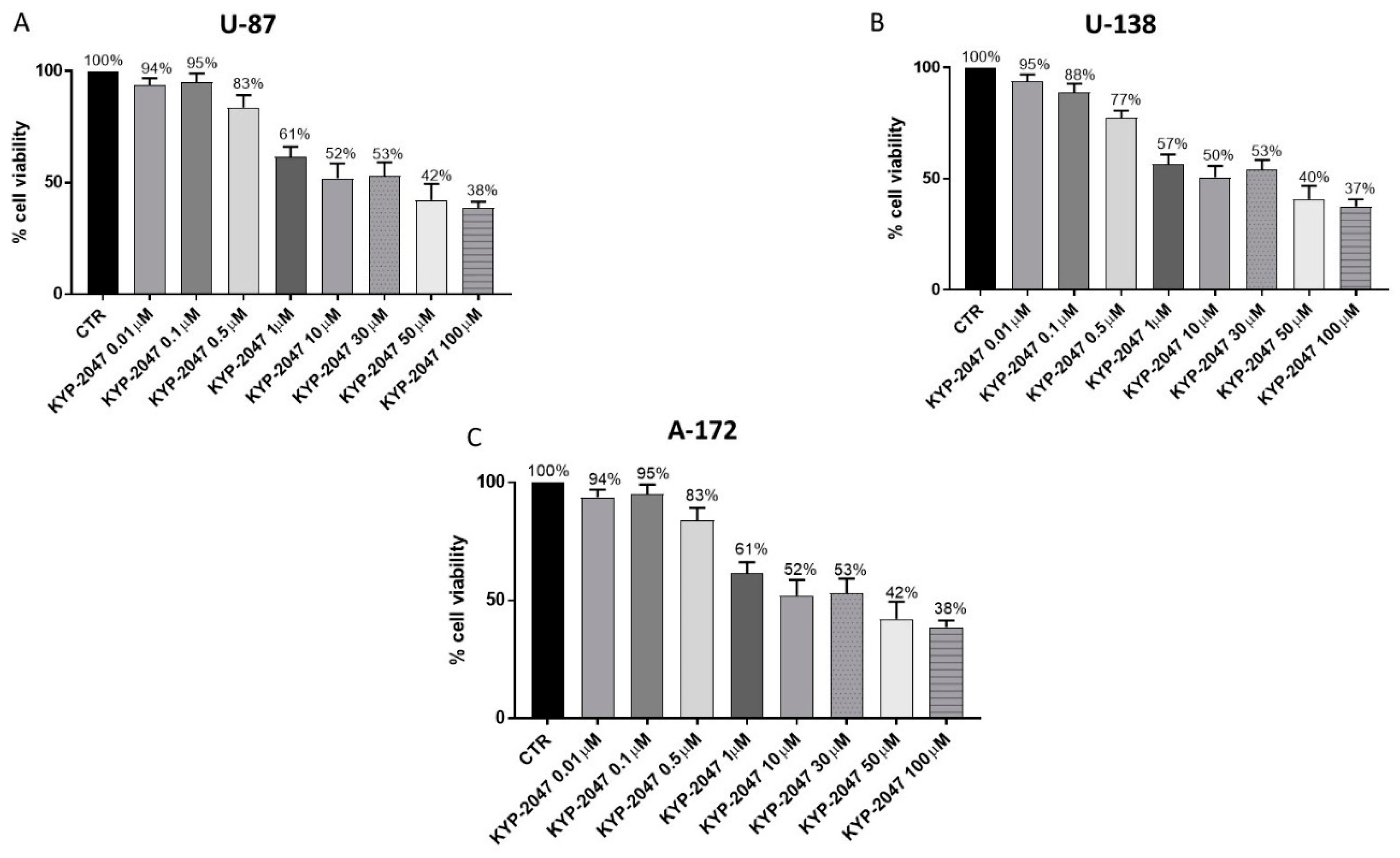
3.2.1. Effect of KYP-2047 on Cell Viability
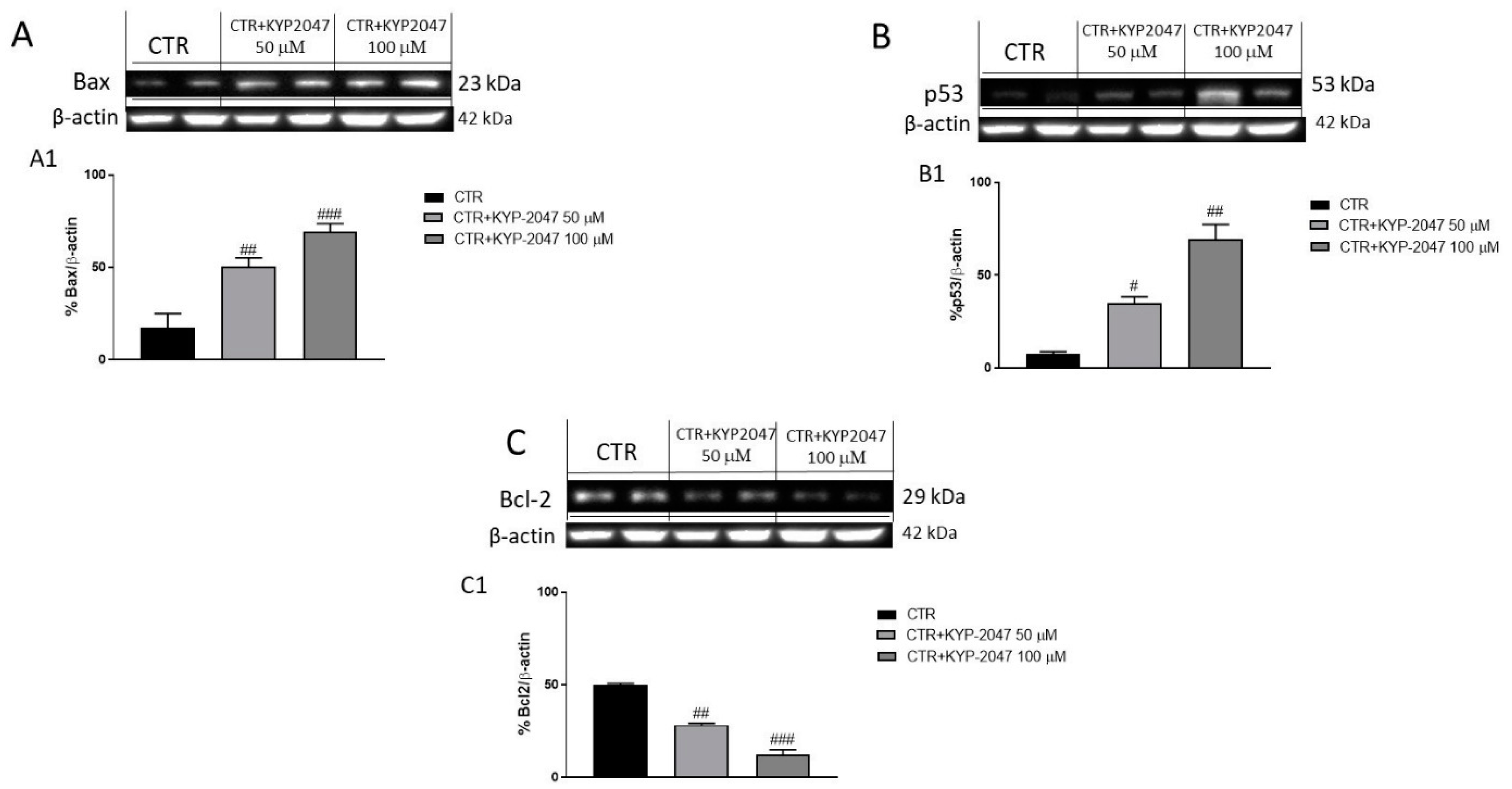
3.2.2. Effect of KYP-2047 on Apoptosis Pathway
3.2.3. Effect of KYP-2047 on TGF-β and Caspase-3 Expression by Immunofluorescence Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- DeAngelis, L.M. Brain tumors. N. Engl. J. Med. 2001, 344, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Hanif, F.; Muzaffar, K.; Perveen, K.; Malhi, S.M.; Simjee, S.U. Glioblastoma Multiforme: A Review of its Epidemiology and Pathogenesis through Clinical Presentation and Treatment. Asian Pac. J. Cancer Prev. 2017, 18, 3–9. [Google Scholar] [CrossRef]
- Lieberman, F. Glioblastoma update: Molecular biology, diagnosis, treatment, response assessment, and translational clinical trials. F1000Research 2017, 6, 1892. [Google Scholar] [CrossRef] [Green Version]
- Sattiraju, A.; Sai, K.K.S.; Mintz, A. Glioblastoma Stem Cells and Their Microenvironment. Adv. Exp. Med. Biol. 2017, 1041, 119–140. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, M.Y.; Assem, M. Glioblastoma Genomics: A Very Complicated Story. In Glioblastoma; De Vleeschouwer, S., Ed.; Codon Publications: Brisbane, Australia, 2017. [Google Scholar] [CrossRef]
- Okada, M.; Miyake, K.; Tamiya, T. Glioblastoma Treatment in the Elderly. Neurol. Med. Chir. 2017, 57, 667–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, X.; Sughrue, M.E. Glioblastoma: New therapeutic strategies to address cellular and genomic complexity. Oncotarget 2018, 9, 9540–9554. [Google Scholar] [CrossRef] [Green Version]
- Broekman, M.L.; Maas, S.L.N.; Abels, E.R.; Mempel, T.R.; Krichevsky, A.M.; Breakefield, X.O. Multidimensional communication in the microenvirons of glioblastoma. Nat. Rev. Neurol. 2018, 14, 482–495. [Google Scholar] [CrossRef] [PubMed]
- Trejo-Solis, C.; Serrano-Garcia, N.; Escamilla-Ramirez, A.; Castillo-Rodriguez, R.A.; Jimenez-Farfan, D.; Palencia, G.; Calvillo, M.; Alvarez-Lemus, M.A.; Flores-Najera, A.; Cruz-Salgado, A.; et al. Autophagic and Apoptotic Pathways as Targets for Chemotherapy in Glioblastoma. Int. J. Mol. Sci. 2018, 19, 3773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hundsberger, T.; Reardon, D.A.; Wen, P.Y. Angiogenesis inhibitors in tackling recurrent glioblastoma. Expert Rev. Anticancer Ther. 2017, 17, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Casili, G.; Lanza, M.; Scuderi, S.A.; Messina, S.; Paterniti, I.; Campolo, M.; Esposito, E. The Inhibition of Prolyl Oligopeptidase as New Target to Counteract Chronic Venous Insufficiency: Findings in a Mouse Model. Biomedicines 2020, 8, 604. [Google Scholar] [CrossRef]
- Jalkanen, A.J.; Leikas, J.V.; Forsberg, M.M. KYP-2047 penetrates mouse brain and effectively inhibits mouse prolyl oligopeptidase. Basic Clin. Pharmacol. Toxicol. 2014, 114, 460–463. [Google Scholar] [CrossRef] [Green Version]
- Myohanen, T.T.; Tenorio-Laranga, J.; Jokinen, B.; Vazquez-Sanchez, R.; Moreno-Baylach, M.J.; Garcia-Horsman, J.A.; Mannisto, P.T. Prolyl oligopeptidase induces angiogenesis both in vitro and in vivo in a novel regulatory manner. Br. J. Pharmacol. 2011, 163, 1666–1678. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.; Zhu, S.; Qiao, Y.; Liu, Y.J.; Chen, W.; Zhao, G.; Chen, J. Recent advances in the role of toll-like receptors and TLR agonists in immunotherapy for human glioma. Protein Cell 2014, 5, 899–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ovcharenko, D.; Chitjian, C.; Kashkin, A.; Fanelli, A.; Ovcharenko, V. Two dichloric compounds inhibit in vivo U87 xenograft tumor growth. Cancer Biol. Ther. 2019, 20, 1281–1289. [Google Scholar] [CrossRef]
- Esposito, E.; Campolo, M.; Casili, G.; Lanza, M.; Franco, D.; Filippone, A.; Peritore, A.F.; Cuzzocrea, S. Protective Effects of Xyloglucan in Association with the Polysaccharide Gelose in an Experimental Model of Gastroenteritis and Urinary Tract Infections. Int. J. Mol. Sci. 2018, 19, 1844. [Google Scholar] [CrossRef] [Green Version]
- Impellizzeri, D.; Bruschetta, G.; Di Paola, R.; Ahmad, A.; Campolo, M.; Cuzzocrea, S.; Esposito, E.; Navarra, M. The anti-inflammatory and antioxidant effects of bergamot juice extract (BJe) in an experimental model of inflammatory bowel disease. Clin. Nutr. 2015, 34, 1146–1154. [Google Scholar] [CrossRef]
- Weinert, B.T.; Krishnadath, K.K.; Milano, F.; Pedersen, A.W.; Claesson, M.H.; Zocca, M.B. Real-time PCR analysis of genes encoding tumor antigens in esophageal tumors and a cancer vaccine. Cancer Immun. 2009, 9, 9. [Google Scholar] [PubMed]
- Irrera, N.; D’Ascola, A.; Pallio, G.; Bitto, A.; Mannino, F.; Arcoraci, V.; Rottura, M.; Ieni, A.; Minutoli, L.; Metro, D.; et al. Beta-Caryophyllene Inhibits Cell Proliferation through a Direct Modulation of CB2 Receptors in Glioblastoma Cells. Cancers 2020, 12, 1038. [Google Scholar] [CrossRef] [Green Version]
- Campolo, M.; Casili, G.; Lanza, M.; Filippone, A.; Paterniti, I.; Cuzzocrea, S.; Esposito, E. Multiple mechanisms of dimethyl fumarate in amyloid beta-induced neurotoxicity in human neuronal cells. J. Cell. Mol. Med. 2018, 22, 1081–1094. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, J.G. Immunofluorescence Staining. Curr. Protoc. Cell Biol. 2015, 69, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Ahir, B.K.; Engelhard, H.H.; Lakka, S.S. Tumor Development and Angiogenesis in Adult Brain Tumor: Glioblastoma. Mol. Neurobiol. 2020, 57, 2461–2478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Li, X.; Luo, M.; Guo, Y.; Li, C.; Lv, D.; Cheng, Z.; Huang, L.; Shang, F.F.; Huang, B.; et al. Tubeimoside I promotes angiogenesis via activation of eNOS-VEGF signaling pathway. J. Ethnopharmacol. 2021, 267, 113642. [Google Scholar] [CrossRef]
- Wood, K.C.; Cortese-Krott, M.M.; Kovacic, J.C.; Noguchi, A.; Liu, V.B.; Wang, X.; Raghavachari, N.; Boehm, M.; Kato, G.J.; Kelm, M.; et al. Circulating blood endothelial nitric oxide synthase contributes to the regulation of systemic blood pressure and nitrite homeostasis. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1861–1871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chabowski, M.; Nowak, A.; Grzegrzolka, J.; Piotrowska, A.; Janczak, D.; Dziegiel, P. Comparison of Microvessel Density Using Nestin and CD34 in Colorectal Cancer. Anticancer Res. 2018, 38, 3889–3895. [Google Scholar] [CrossRef] [PubMed]
- Sie, M.; Wagemakers, M.; Molema, G.; Mooij, J.J.; de Bont, E.S.; den Dunnen, W.F. The angiopoietin 1/angiopoietin 2 balance as a prognostic marker in primary glioblastoma multiforme. J. Neurosurg. 2009, 110, 147–155. [Google Scholar] [CrossRef]
- Wang, P.F.; Li, H.L.; Qi, X.; Yao, K.; Han, S.; Liu, N.; Yang, Y.K.; Li, S.W.; Yan, C.X. Clinical significance of angiopoietin-like protein 3 expression in patients with glioblastoma. Neoplasma 2016, 63, 93–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rautiola, J.; Lampinen, A.; Mirtti, T.; Ristimaki, A.; Joensuu, H.; Bono, P.; Saharinen, P. Association of Angiopoietin-2 and Ki-67 Expression with Vascular Density and Sunitinib Response in Metastatic Renal Cell Carcinoma. PLoS ONE 2016, 11, e0153745. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Cai, H.; Sun, L.; Zhan, P.; Chen, M.; Zhang, F.; Ran, Y.; Wan, J. LGR5, a novel functional glioma stem cell marker, promotes EMT by activating the Wnt/beta-catenin pathway and predicts poor survival of glioma patients. J. Exp. Clin. Cancer Res. 2018, 37, 225. [Google Scholar] [CrossRef]
- Valdes-Rives, S.A.; Casique-Aguirre, D.; German-Castelan, L.; Velasco-Velazquez, M.A.; Gonzalez-Arenas, A. Apoptotic Signaling Pathways in Glioblastoma and Therapeutic Implications. Biomed. Res. Int. 2017, 2017, 7403747. [Google Scholar] [CrossRef] [Green Version]
- Pistritto, G.; Trisciuoglio, D.; Ceci, C.; Garufi, A.; D’Orazi, G. Apoptosis as anticancer mechanism: Function and dysfunction of its modulators and targeted therapeutic strategies. Aging 2016, 8, 603–619. [Google Scholar] [CrossRef] [Green Version]
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, F.; Ghosh, S.; Sinha, S.; Joshi, S.D.; Mehta, V.S.; Sen, E. TGF-beta-induced hCG-beta regulates redox homeostasis in glioma cells. Mol. Cell. Biochem. 2015, 399, 105–112. [Google Scholar] [CrossRef]
- Margadant, C.; Sonnenberg, A. Integrin-TGF-beta crosstalk in fibrosis, cancer and wound healing. EMBO Rep. 2010, 11, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Siegel, P.M.; Massague, J. Cytostatic and apoptotic actions of TGF-beta in homeostasis and cancer. Nat. Rev. Cancer 2003, 3, 807–821. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Ekman, M.; Thakur, N.; Bu, S.; Davoodpour, P.; Grimsby, S.; Tagami, S.; Heldin, C.H.; Landstrom, M. TGFbeta1-induced activation of ATM and p53 mediates apoptosis in a Smad7-dependent manner. Cell Cycle 2006, 5, 2787–2795. [Google Scholar] [CrossRef] [Green Version]
- Wirsching, H.G.; Galanis, E.; Weller, M. Glioblastoma. Handb. Clin. Neurol. 2016, 134, 381–397. [Google Scholar] [CrossRef] [PubMed]
- Soomro, S.H.; Ting, L.R.; Qing, Y.Y.; Ren, M. Molecular biology of glioblastoma: Classification and mutational locations. J. Pak. Med. Assoc. 2017, 67, 1410–1414. [Google Scholar]
- SongTao, Q.; Lei, Y.; Si, G.; YanQing, D.; HuiXia, H.; XueLin, Z.; LanXiao, W.; Fei, Y. IDH mutations predict longer survival and response to temozolomide in secondary glioblastoma. Cancer Sci. 2012, 103, 269–273. [Google Scholar] [CrossRef]
- Venalainen, J.I.; Garcia-Horsman, J.A.; Forsberg, M.M.; Jalkanen, A.; Wallen, E.A.; Jarho, E.M.; Christiaans, J.A.; Gynther, J.; Mannisto, P.T. Binding kinetics and duration of in vivo action of novel prolyl oligopeptidase inhibitors. Biochem. Pharmacol. 2006, 71, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Weathers, S.P.; de Groot, J. VEGF Manipulation in Glioblastoma. Oncology 2015, 29, 720–727. [Google Scholar] [PubMed]
- Carmeliet, P. VEGF as a key mediator of angiogenesis in cancer. Oncology 2005, 69 (Suppl. S3), 4–10. [Google Scholar] [CrossRef]
- Tanaka, S.; Mori, M.; Sakamoto, Y.; Makuuchi, M.; Sugimachi, K.; Wands, J.R. Biologic significance of angiopoietin-2 expression in human hepatocellular carcinoma. J. Clin. Investig. 1999, 103, 341–345. [Google Scholar] [CrossRef] [Green Version]
- Stoeltzing, O.; Ahmad, S.A.; Liu, W.; McCarty, M.F.; Parikh, A.A.; Fan, F.; Reinmuth, N.; Bucana, C.D.; Ellis, L.M. Angiopoietin-1 inhibits tumour growth and ascites formation in a murine model of peritoneal carcinomatosis. Br. J. Cancer 2002, 87, 1182–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastronardi, L.; Guiducci, A.; Puzzilli, F.; Ruggeri, A. Relationship between Ki-67 labeling index and survival in high-grade glioma patients treated after surgery with tamoxifen. J. Neurosurg. Sci. 1999, 43, 263–270. [Google Scholar] [PubMed]
- Alkhaibary, A.; Alassiri, A.H.; AlSufiani, F.; Alharbi, M.A. Ki-67 labeling index in glioblastoma; does it really matter? Hematol. Oncol. Stem. Cell Ther. 2019, 12, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.S. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [Green Version]
- Katsuno, Y.; Lamouille, S.; Derynck, R. TGF-beta signaling and epithelial-mesenchymal transition in cancer progression. Curr. Opin. Oncol. 2013, 25, 76–84. [Google Scholar] [CrossRef]
- Chio, C.C.; Chen, K.Y.; Chang, C.K.; Chuang, J.Y.; Liu, C.C.; Liu, S.H.; Chen, R.M. Improved effects of honokiol on temozolomide-induced autophagy and apoptosis of drug-sensitive and -tolerant glioma cells. BMC Cancer 2018, 18, 379. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scuderi, S.A.; Casili, G.; Ardizzone, A.; Forte, S.; Colarossi, L.; Sava, S.; Paterniti, I.; Esposito, E.; Cuzzocrea, S.; Campolo, M. KYP-2047, an Inhibitor of Prolyl-Oligopeptidase, Reduces GlioBlastoma Proliferation through Angiogenesis and Apoptosis Modulation. Cancers 2021, 13, 3444. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13143444
Scuderi SA, Casili G, Ardizzone A, Forte S, Colarossi L, Sava S, Paterniti I, Esposito E, Cuzzocrea S, Campolo M. KYP-2047, an Inhibitor of Prolyl-Oligopeptidase, Reduces GlioBlastoma Proliferation through Angiogenesis and Apoptosis Modulation. Cancers. 2021; 13(14):3444. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13143444
Chicago/Turabian StyleScuderi, Sarah Adriana, Giovanna Casili, Alessio Ardizzone, Stefano Forte, Lorenzo Colarossi, Serena Sava, Irene Paterniti, Emanuela Esposito, Salvatore Cuzzocrea, and Michela Campolo. 2021. "KYP-2047, an Inhibitor of Prolyl-Oligopeptidase, Reduces GlioBlastoma Proliferation through Angiogenesis and Apoptosis Modulation" Cancers 13, no. 14: 3444. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13143444