
Pyroptosis in Cancer: Friend or Foe?



Melanoma and Sarcoma Medical Oncology Unit, State Key Laboratory of Oncology in South China, Collaborative Innovation Center for Cancer Medicine, Sun Yat-sen University Cancer Center, Guangzhou 510060, China
*
Author to whom correspondence should be addressed.
Cancers 2021, 13(14), 3620; https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13143620
Submission received: 11 June 2021
/
Revised: 9 July 2021
/
Accepted: 14 July 2021
/
Published: 20 July 2021
Abstract
:Simple Summary
Pyroptosis is a new form of programmed cell death that differs from apoptosis in terms of its release of inflammatory factors and its characteristic bubble-like morphology. Pyroptosis was first discovered in the process of immune defense against bacterial infection, but the field of research soon spread to other inflammatory diseases and cancer. As cancer constitutes a serious risk for public health, numerous studies investigating pyroptosis in cancer have been carried out during these years. Tumorigenesis and new therapeutic treatments have been the focus of much recent research. This review discusses the role of pyroptosis in tumorigenesis and its influence on tumor immunity.
Abstract
Pyroptosis is an inflammatory form of programmed cell death that is mediated by pore-forming proteins such as the gasdermin family (GSDMs), including GSDMA-E. Upon cleavage by activated caspases or granzyme proteases, the N-terminal of GSDMs oligomerizes in membranes to form pores, resulting in pyroptosis. Though all the gasdermin proteins have been studied in cancer, the role of pyroptosis in cancer remains mysterious, with conflicting findings. Numerous studies have shown that various stimuli, such as pathogen-associated molecular patterns (PAMPs), damage-associated molecular patterns (DAMPs), and chemotherapeutic drugs, could trigger pyroptosis when the cells express GSDMs. However, it is not clear whether pyroptosis in cancer induced by chemotherapeutic drugs or CAR T cell therapy is beneficial or harmful for anti-tumor immunity. This review discusses the discovery of pyroptosis as well as its role in inflammatory diseases and cancer, with an emphasis on tumor immunity.
1. Introduction
Programmed cell death mediated by specific signaling pathways plays a vital role in morphogenesis, the maintenance of homeostasis, and various diseases [1]. According to the morphology changes involved, programmed cell death can be divided into lytic and non-lytic cell death. Apoptosis is a well-studied form of non-lytic cell death without the release of pro-inflammatory factors. However, pyroptosis and necroptosis are lytic and highly inflammatory in nature [2,3]. Cross talk between the various forms of cell death is mediated by numerous pathways. Among these, caspase 8 is considered as a molecular switch that controls apoptosis, necroptosis, and pyroptosis [4,5].
Pyroptosis was first studied in the field of immune defense against pathogens. This field of research gradually extended to other diseases, especially cancer, as cancer has become a leading heath menace in recent decades. Many studies have found that tumor cells can be killed through pyroptosis, but the role of pyroptosis in tumorigenesis or anti-tumor immunity has not yet been clearly summarized. In this review, we aim to discuss how pyroptosis was discovered and the roles of pyroptosis in human diseases, including tumors, with an emphasis on the relationship between pyroptosis and tumor immunity.
2. The Discovery of Pyroptosis
Pyroptosis was first observed in 1992 when A. Zychlinsky treated macrophages with Shigella flexneri and saw lytic cell death [6]. However, this was considered to be a case of apoptosis at that time, and not until 2000 did Boise and Collins name it pyroptosis [7]. Cookson [8] demonstrated that, unlike apoptosis, pyroptosis was associated with the release of some proinflammatory factors and was dependent on caspase-1 instead of caspase-3, as in apoptosis. With the breakthrough in research on the inflammasome [9], pyroptosis was found to be induced upon the activation of caspase-1/4/5/11 and to occur in both macrophage and non-macrophage cells [10,11,12,13,14]. The underlying mechanism of how inflammatory caspases trigger pyroptosis remained unknown until Feng Shao [15] discovered that one of the gasdermin proteins, GSDMD, is required for caspase-1-mediated pyroptosis. When cells respond to an exogenous pathogen or endogenous damage, a canonical inflammasome is formed by the adaptor protein ASC and upstream sensors NLRP1b/NLRP3/NLRC4/AIM2/Pyrin, which in turn activates the effector pro-caspase 1, leading to its self-cleavage. The activated caspase 1 then cleaves GSDMD and the inflammatory factors pro-IL-18 and IL-1β. In the non-canonical inflammasome pathway, lipopolysaccharide (LPS) directly binds to and activates pro-caspase 4/5 or murine pro-caspase 11. Then, the activated caspases cleave GSDMD, contributing to the oligomerization of the N-terminal in membranes to form pores [16,17,18,19,20], leading to lytic cell death and the release of inflammatory cytokines such as IL-18, IL1β, and HMGB1 [21]. Later, it was discovered that the gasdermin family GSDMA/B/C/D/E could all be cleaved by activated caspases and granzyme proteases, and the N-terminal oligomerizes in membranes to form pores [22], leading to pyroptosis (Figure 1).
3. The Role of Pyroptosis in Inflammatory Diseases
Although pyroptosis was first discovered in Shigella flexneri-infected macrophages [23], Salmonella, various other bacteria [24], and viruses soon demonstrated pyroptosis inducing capability in target cells besides macrophages. In addition to its role in inflammatory diseases, pyroptosis also plays a vital part in other ailments. Diabetes is a major risk factor underlying cardiovascular diseases. Studies have shown that NLRP3 is a mediator of pyroptosis in cardiomyocytes, leading to diabetic cardiomyopathy [25,26,27]. Moreover, nicotine [28], cholesterol crystals [29], oxLDL [30], and cadmium [31] can disrupt the coronary arterial endothelial cells via pyroptosis, causing atherosclerosis. Additionally, pyroptosis in vascular smooth muscle cells and monocytes/macrophages also accelerates the pathogenesis of coronary atherosclerosis. Pyroptosis has also been reported in common neurological disorders. Increased intracellular calcium, mitochondrial dysfunction, and other endogenous compounds can activate the NLRP3 inflammasome and trigger pyroptosis in microglia and dopamine neurons, increasing the incidence of neuroinflammation and promoting the progression of Parkinson’s disease [32] and inflammatory demyelination during multiple sclerosis (MS) [33].
Taken together, cell death by pyroptosis has been found to be a common factor associated with the progression of many inflammation-associated diseases. Numerous studies have shown that it is possible to attenuate inflammation by blocking pyroptosis with inhibitors targeting caspase-1/3/4/5/11 or the NLRP3 inflammasome. HIV infection induces the cell death of quiescent lymphoid CD4 T cells by caspase-1-mediated pyroptosis, and those dying CD4 T cells release inflammatory signals that cause more cells to die [34]. This indicates that caspase 1 inhibitors could be a new class of ‘anti-AIDS’ therapeutic targets.
4. The Roles of Pyroptosis in Cancer: Promoting or Inhibiting the Tumor?
4.1. The Role of Inflammasomes in Cancer
Inflammasomes are protein complexes that are assembled in response to pathogens or intercellular danger. The type of inflammasome involved depends on the sensor used. These sensors consist of pyrin, AIM2, and nucleotide-binding domain (NOD)-like receptors (NLRs) such as NLRP1, NLRP3, and NLRC4, among which NLRP3 has been the most widely studied [35]. The NLRP3 inflammasome is involved in a variety of inflammation-associated diseases. In terms of tumors, the NLRP3 inflammasome could enhance the proliferation and migration of A549 lung cancer cells [36] (Table 1), and an increased expression of NLRP3 was positively correlated to tumor growth in oral squamous cell carcinoma (OSCC) [37] (Table 1). Another study showed that NLRP3 could promote epithelial to mesenchymal transition in colon cancer cells in an inflammasome-independent manner [38] (Table 1). Moreover, the NLRP3 inflammasome is involved in chemoresistance in oral squamous cell carcinoma [39] (Table 1) and insensitivity to radiotherapy in glioblastoma [40] (Table 1). Various studies have confirmed that the NLRP3-mediated release of IL-18 [41,42,43] or IL-1β [44,45,46,47] enhances the malignancy of lymphoma, gastric cancer, breast cancer, colon cancer, and glioblastoma via promoting the proliferation of cancer, immunosuppression, angiogenesis, and metastasis [48,49]. Similarly, Zhai et al. studied the tumor-promoting role of NLRP1 and reported that NLRP1 could augment the activation of inflammasomes and suppress apoptosis in metastatic melanoma [50].
On the contrary, a number of studies have also shown the anti-tumor activity of inflammasomes. Lower levels of NLRP3 [51] (Table 1), NLRP1 [54], and AIM2 [55] proteins were found in hepatocellular carcinoma (HCC), colorectal cancer (CRC), and gastric cancer (GC) compared to the adjacent normal tissue. A low level of these proteins was correlated with the advanced stage and poor prognosis of HCC, suggesting that the inflammasomes served as negative regulators in HCC tumorigenesis. The anti-tumor role of inflammasomes has been extensively studied in colitis-associated cancer, showing that the secretion of IL-18 mediated by the NLRP3 inflammasome could protect enterocytes from early-stage colitis induced by dextran sulfate sodium (DSS) or azoxymethane (AOM) [52,56,57,58] (Table 1). Furthermore, a study found that NLRC4 in tumor-associated macrophages (TAMs) inhibited the progression of melanoma independently of inflammasome activation [59]. Additionally, the NLRP3 inflammasome in Kupffer cells suppressed colorectal cancer metastatic growth in the liver by promoting the maturation of NK cells and tumoricidal activity mediated by IL-18 [53] (Table 1).
Overall, the role of the inflammasome in different cancer types is ambiguous (Figure 2).
4.2. The Role of Gasdermin Family Proteins in Cancer
Gasdermin family proteins, which include GSDMA, GSDMB, GSDMC, GSDMD, and GSDME, had been studied in other fields but not in cancers. Only in the last few years have their roles in pyroptosis and cancer been studied.
4.2.1. GSDMA, GSDMB and GSDMC
To date, the roles of GSDMA/B/C in tumors have been described only vaguely, and very limited information is available about them. GSDMA is usually expressed in epithelial cells and its lineage GSDMA3 has been proposed to be involved in epidermal differentiation [60], TNF-α-induced apoptosis [61], skin inflammation, and hair loss [62,63,64]. Polymorphisms of GSDMB as well as GSDMA were found to be linked to the development of childhood asthma [65,66,67]. GSDMA is frequently silenced in gastric cancers [68,69], indicating that GSDMA might function as a regulator of tumor suppression. Interestingly, GSDMB could be considered as an oncogene because of its enhanced expression in gastric cancers, hepatic carcinomas, cervical tumors, and breast cancer [70,71,72]. Additionally, many studies have also proposed that GSDMB possessed protumor functions such as migration, metastasis, and resistance to therapy in HER2 breast cancer [73,74].
Due to its constricted expression, there are no known diseases associated with GSDMC. GSDMC has been reported to be upregulated by transforming growth factor beta receptor 2 (TGFBR2) mutation in colorectal cancer (CRC), promoting tumor cell proliferation [75]. Likewise, the overexpression of GSDMC was confirmed in lung adenocarcinoma (LUAD) patients, acting as a promising predictive factor [76]. A recent study demonstrated that, under hypoxia, nuclear PD-L1 was found to enhance the expression of GSDMC, and breast cancer patients with an increased expression of GSDMC had poorer outcomes, suggesting that the chronic tumor necrosis induced by GSDMC in the center of hypoxia regions may promote tumor progression [77]. However, GSDMC expression is suppressed in some cell lines of both esophageal and gastric cancers [78], supporting its potential role as an anti-tumor agent.
From the above studies, we could not decide the definite roles of GSDMA/B/C in cancer. However, as a substrate of caspases or granzymes, GSDMs could be cleaved and the oligomerization of N-terminals in membranes would trigger pyroptosis, which has been confirmed in many cancers.
4.2.2. GSDMD
Despite its role in innate immunity, GSDMD also plays a crucial part in carcinogenesis. The protein level of GSDMD was positively associated with the aggressiveness of non-small cell lung cancer (NSCLC). GSDMD modulated the EGFR/Akt signaling in NSCLC and promoted the proliferation of tumor by inhibiting apoptosis [79]. However, another study showed that the expression of GSDMD in gastric cancer (GC) was decreased compared to that in matched adjacent non-cancerous tissues and that the downregulation of GSDMD accelerated S/G2 cell transition, suggesting that GSDMD may serve as a tumor suppressor [80].
Many small-molecule inhibitors or drugs displaying tumoricidal activity through apoptosis or autography were also found to function by inducing GSDMD-mediated pyroptosis. Docosahexaenoic acid (DHA), an omega-3 fatty acid with anticancer effects, was verified to induce pyroptotic cell death in triple-negative breast cancer cells [81] (Table 2). Likewise, α-NETA induces the pyroptosis of epithelial ovarian cancer cells through the GSDMD/caspase-4 pathway [82] (Table 2). Polyphyllin VI (PPVI) [83] (Table 2) suppressed the proliferation of non-small cell lung cancer (NSCLC) by inducing pyroptosis via the induction of the ROS/NF-κB/NLRP3/GSDMD signal axis in NSCLC. Metformin could induce GSDMD-mediated pyroptosis in esophageal squamous cell carcinoma (ESCC) by targeting the miR-497-PELP1 axis, indicating that pyroptosis-inducing reagents could serve as alternative treatments for chemo- and radiotherapy refractory ESCC [84] (Table 2). Many new therapeutic targets and tumor markers have also been discovered in the process of studying tumoricidal drugs and pyroptosis. Small-molecule inhibitors of the serine dipeptidases DPP8 and DPP9 (DPP8/9) activated CARD8 and caspase-1 to trigger pyroptosis in the majority of human acute myeloid leukemia (AML) [85] (Table 2), indicating the protein CARD8 could be a novel therapeutic target for AML. The activation of pyroptosis in head and neck squamous cell carcinoma (HNSCC) was found to be a calcium-dependent process, and the reduced expression of calcium ion regulator CD38 could prevent inflammasome-induced pyroptosis in HNSCC, indicating that CD38 may function as a tumor suppressor in HNSCC [86].
Overall, despite its uncertain role in tumorigenesis, GSDMD-mediated pyroptosis in cancer has attracted huge interest from researchers worldwide and has remained a critical study topic, indicating its importance as a new therapeutic target.
4.2.3. GSDME
As one of the members of the gasdermin family, GSDME is also known as deafness, autosomal dominant 5 (DFNA5). Based on several studies, the mutations at the DNA level result in the skipping of exon 8 [93,94,95] and the truncation of the protein, which augments the intrinsic pore-forming activity due to loss of the inhibitory C-terminal domain and thus damages hearing-associated cells and leads to hearing loss [96,97,98]. In contrast, the inactivation of GSDME in the form of decreased expression instead of genetic mutation is associated with cancer. Methylation analysis shows that in breast cancer [99,100,101,102] and colorectal cancer [103,104,105], the hypermethylated promoter of GSDME was relevant to patients’ prognosis, suggesting that GSDME methylation could be a potential detection and prognostic marker [99,105]. In most cancers, GSDME expression is decreased in tumor tissues compared to normal tissues [99,102,104,106,107]. Furthermore more, the induced expression of GSDME in cancer cell lines leads to decreased proliferation [108] and suppressed colony formation [104], while GSDME deficiency contributes to the malignancy of tumors [109] and drug resistance [110]. These studies have suggested that GSDME may function as a tumor suppressor gene. However, some other studies found that there is no clear correlation between GSDME promoter hypermethylation and GSDME expression [107], and there is no significant difference in GSDME expression between tumors and normal tissues [103]. Furthermore, GSDME-WT cancer cells develop tumors of comparable size to that of the GSDME-KO cells in colon cancer [87,111] (Table 2) and lung cancer [112] models, demonstrating that GSDME may not be involved in tumorigenesis. Surprisingly, one study even reported that higher GSDME expression in esophageal squamous cell carcinoma [88] (Table 2) was considered as a promising prognostic marker. Based on the above studies, the roles of GSDME in cancer are still controversial.
After Feng Shao discovered that the activation of caspase-3 by chemotherapeutic agents could trigger pyroptosis by cleaving GSDME [113], numerous studies showed that small-molecule inhibitors or traditional chemotherapeutic drugs could cause cell death via pyroptosis when tumor-expressing GSDME was cleaved by activated caspase-3 in various cancers [87,88,89,92] (Table 2). As we mentioned before that the expression of GSDME was associated with the sensitivity of chemotherapy, studies showed that GSDME deficiency could impair the efficacy of cisplatin [89] (Table 2), sulfasalazine together with iron dextran [114], or ceritinib [112]. However, in some other cases, GSDME did not affect the tumoricidal activity of 5-FU [90] (Table 2) or lobaplatin [87] (Table 2). All these tumoricidal substances kill cancer cells not only by pyroptosis but also through apoptosis or autophagy. However, in the absence of GSDME, pyroptosis could not be triggered.
Taken together, the role of GSDME in cancer could not be determined, with conflicting results in tumorigenesis. Thus, it still necessary to conduct further research, especially concerning the induction of pyroptosis in tumors.
4.2.4. The Significance of Pyroptosis in Tumorigenesis or Tumor Progression?
Thus far, all gasdermin family proteins have been studied in cancers but neither the relationship between their expression and tumor malignancy nor the function of GSDMs in tumorigenesis are not consistent. Although some studies have shown that the ectopic expression of GSDMs promoted the formation of tumors [115], the underlying mechanism has not been revealed. It is well known that tumor development is a multi-stage process and different mechanisms operate at different times in various cancers [116,117]. The factors involved in tumor initiation, such as the loss-of-function mutations in p53 [118] or the formation of oncoprotein BCR-ABL [119], have been proven to play a role in the initial stages. As the tumor grows, the inner part of tumor cells suffers from more hypoxia stress, starvation, and other harsh conditions. One recent study found that pyroptosis in a small fraction of tumor cells in the central hypoxia region of cancer contributed to chronic tumor necrosis, which in turn suppressed the anti-tumor immunity and promoted tumor progression [77]. Without therapeutic regents, the pyroptosis of cancer cells triggered by an adverse tumor microenvironment is more likely to accelerate the progression. Thus, as far as we are concerned, pyroptosis is more consistently related to metastasis and late-stage solid tumors than to preneoplastic lesions or early-stage tumors. Moreover, gasdermin family proteins may have some other cellular functions despite their contribution to pyroptosis, and this also deserves further research.
5. The Correlation between Pyroptosis and Therapeutic Reagents
Apoptosis is the most widely studied process of tumor inhibition and numerous chemotherapeutic agents for inducing apoptosis have been extensively studied. Of late, many researchers have clearly demonstrated that apoptosis could be switched to pyroptosis [120]. Pyroptosis is quite common in cancer cells treated with traditional chemotherapeutic drugs or promising small-molecule targets. Though there is no solid evidence that GSDMs are linked to tumorigenesis, the expression of the gasdermin family members in caners determines whether pyroptosis could be induced or not. Moreover, the expression of GSDMs is always relatively higher in normal tissues as compared to cancers, and under this condition pyroptosis would be more easily induced in normal tissues instead of tumors, potentiating unwanted effects or side effects of tumoricidal drugs. One study showed that doxorubicin induced-cardiotoxicity was mediated by pyroptosis in cardiomyocyte regulated through the Bnip3-caspase-3-GSDME pathway [121]. Similarly, the chemotherapeutic drugs cisplatin and doxorubicin also induced nephrotoxicity through pyroptosis [122]. Based on these results, we could see that pyroptosis not only mediated the antitumor activity of the therapeutic agents but also their side effects. At present, studies in this area mostly focus on the therapeutic effects of the targeting drugs and whether the drugs induce pyroptosis and fuel anti-tumor immunity.
All in all, as a form of cell death, pyroptosis is universal among cancer cells and normal cells treated with tumoricidal reagents. Patients may benefit more from more tumor-specific treatment, such as the selective induction of pyroptosis in tumor cells, which directly kills tumors but also activates anti-tumor immunity.
6. The Relationship between Pyroptosis and Anti-Tumor Immunity
Numerous studies have revealed that pyroptosis could fuel anti-tumor immunity. The application of a biorthogonal system to GSDMA3 in tumor cells showed that pyroptosis may augment the antitumor immune response and increase the efficacy of immune checkpoint blockade [123] (Table 3). BRAF inhibitors together with MEK inhibitors could induce pyroptosis in melanoma with a higher HMGB1, more tumor-associated T cells, and less dendritic cell infiltration, while GSDME deficiency would reverse the anti-tumor immunity [91] (Table 2 and Table 3). Two other recent studies found that the granzymes released from CD8 +T cells and NK cells could cleave GSDMB/E and thus trigger the pyroptosis of tumor cells, indicating that pyroptosis might serve as an important effector in anti-tumor immunity [115,124] (Table 3). However, Gao Tan’s [21] study found that HMGB1, a proinflammatory factor released from GSDME-mediated pyroptotic epithelial cells, induces colorectal cancer proliferation through the ERK1/2 pathway. Furthermore, Liu’s research found that pyroptosis-released factors from tumor cells could again trigger pyroptosis in macrophages, resulting in the release of cytokines and subsequent cytokine release syndrome (CRS) [125] (Table 3). Moreover, as mentioned before, IL-18 has both pro [41,42,43] and antitumorigenic effects [52,53]. The effects of other intercellular substances in the tumor microenvironment, such as ATP, HMGB1, IL1β, and LDH, are still uncertain. This is a topic that deserves further study. From these results, we can see that pyroptosis mediates the tumoricidal effect of some inhibitors or cytolytic immune cells, and pyroptosis-released inflammatory factors may fuel the anti-tumor immunity or be harmful to patients by promoting tumor or causing inflammatory cascades. Thus, the relationship between pyroptosis and anti-tumor immunity is not certain and is worthy of further investigation.
7. Conclusions and Future Prospects
Compared with apoptosis or autophagy, pyroptosis is a form of proinflammatory cell death with released inflammatory factors. Pyroptosis was first discovered in the defense of pathogenic insults and later researchers found that pyroptosis mediates many inflammatory diseases. It has been proposed that the inhibition of pyroptosis through targeting caspase-1/3/4/5/11 or the NLRP3 inflammasome could relieve inflammatory diseases. However, some other studies have found that the induction of pyroptosis in tumor cells would enhance the tumoricidal effect. Various tumoricidal substances such as chemotherapeutic drugs or granzymes released from cytotoxic T cells could kill cancer cells via canonical or non-canonical pyroptosis. However, there is no confirmed conclusion that inflammasomes or GSDMs are involved in tumorigenesis or act as tumor suppressors, and it is not certain that pyroptosis induced by tumoricidal drugs is beneficial for tumor patients in the long run because other normal cells could also die by pyroptosis when stimulated by DAMPs released from pyroptotic tumor cells. Moreover, the underlying mechanism that regulates pyroptosis has not been revealed so far and the roles of pyroptosis in anti-tumor immunity are still ambiguous. Above all, every aspect of pyroptosis needs to be elucidated further by more in-depth studies to confirm its function and mechanism. Additionally, it would be also interesting to explore some drug candidates that could inhibit tumors via pyroptosis and facilitate anti-tumor immunity.
Author Contributions
X.L. wrote the manuscript. T.G. and X.Z. revised and corrected the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (81772863, 82072958).
Acknowledgments
We would like to thank Gaurab Roy, Sun Yat-sen University Cancer Center, for providing the professional medical writing assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hughes, L.D.; Wang, Y.; Meli, A.P.; Rothlin, C.V.; Ghosh, S. Decoding Cell Death: From a Veritable Library of Babel to Vade Mecum? Annu. Rev. Immunol. 2021, 39, 791–817. [Google Scholar] [CrossRef] [PubMed]
- Demarco, B.; Chen, K.W.; Broz, P. Cross talk between intracellular pathogens and cell death. Immunol. Rev. 2020, 297, 174–193. [Google Scholar] [CrossRef] [PubMed]
- Bedoui, S.; Herold, M.J.; Strasser, A. Emerging connectivity of programmed cell death pathways and its physiological implications. Nat. Rev. Mol. Cell Biol. 2020, 21, 678–695. [Google Scholar] [CrossRef] [PubMed]
- Fritsch, M.; Günther, S.D.; Schwarzer, R.; Albert, M.C.; Schorn, F.; Werthenbach, J.P.; Schiffmann, L.M.; Stair, N.; Stocks, H.; Seeger, J.M.; et al. Caspase-8 is the molecular switch for apoptosis, necroptosis and pyroptosis. Nature 2019, 575, 683–687. [Google Scholar] [CrossRef]
- Schwarzer, R.; Laurien, L.; Pasparakis, M. New insights into the regulation of apoptosis, necroptosis, and pyroptosis by receptor interacting protein kinase 1 and caspase-8. Curr. Opin. Cell Biol. 2020, 63, 186–193. [Google Scholar] [CrossRef]
- Zychlinsky, A.; Prevost, M.C.; Sansonetti, P.J. Shigella flexneri induces apoptosis in infected macrophages. Nature 1992, 358, 167–169. [Google Scholar] [CrossRef]
- Boise, L.H.; Collins, C.M. Salmonella-induced cell death: Apoptosis, necrosis or programmed cell death? Trends Microbiol. 2001, 9, 64–67. [Google Scholar] [CrossRef]
- Cookson, B.T.; Brennan, M.A. Pro-inflammatory programmed cell death. Trends Microbiol. 2001, 9, 113–114. [Google Scholar] [CrossRef]
- Li, Y.; Huang, H.; Liu, B.; Zhang, Y.; Pan, X.; Yu, X.Y.; Shen, Z.; Song, Y.H. Inflammasomes as therapeutic targets in human diseases. Signal Transduct. Target. Ther. 2021, 6, 247. [Google Scholar] [CrossRef]
- Miao, E.A.; Leaf, I.A.; Treuting, P.M.; Mao, D.P.; Dors, M.; Sarkar, A.; Warren, S.E.; Wewers, M.D.; Aderem, A. Caspase-1-induced pyroptosis is an innate immune effector mechanism against intracellular bacteria. Nat. Immunol. 2010, 11, 1136–1142. [Google Scholar] [CrossRef]
- Miao, E.A.; Rajan, J.V.; Aderem, A. Caspase-1-induced pyroptotic cell death. Immunol. Rev. 2011, 243, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Broz, P.; Ruby, T.; Belhocine, K.; Bouley, D.M.; Kayagaki, N.; Dixit, V.M.; Monack, D.M. Caspase-11 increases susceptibility to Salmonella infection in the absence of caspase-1. Nature 2012, 490, 288–291. [Google Scholar] [CrossRef] [PubMed]
- Hagar, J.A.; Powell, D.A.; Aachoui, Y.; Ernst, R.K.; Miao, E.A. Cytoplasmic LPS activates caspase-11: Implications in TLR4-independent endotoxic shock. Science 2013, 341, 1250–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wree, A.; Eguchi, A.; McGeough, M.D.; Pena, C.A.; Johnson, C.D.; Canbay, A.; Hoffman, H.M.; Feldstein, A.E. NLRP3 inflammasome activation results in hepatocyte pyroptosis, liver inflammation, and fibrosis in mice. Hepatology 2014, 59, 898–910. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Zhao, Y.; Wang, K.; Shi, X.; Wang, Y.; Huang, H.; Zhuang, Y.; Cai, T.; Wang, F.; Shao, F. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 2015, 526, 660–665. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Z.; Ruan, J.; Pan, Y.; Magupalli, V.G.; Wu, H.; Lieberman, J. Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature 2016, 535, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Wang, K.; Liu, W.; She, Y.; Sun, Q.; Shi, J.; Sun, H.; Wang, D.C.; Shao, F. Pore-forming activity and structural autoinhibition of the gasdermin family. Nature 2016, 535, 111–116. [Google Scholar] [CrossRef]
- Kayagaki, N.; Stowe, I.B.; Lee, B.L.; O’Rourke, K.; Anderson, K.; Warming, S.; Cuellar, T.; Haley, B.; Roose-Girma, M.; Phung, Q.T.; et al. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature 2015, 526, 666–671. [Google Scholar] [CrossRef]
- Tan, Y.; Chen, Q.; Li, X.; Zeng, Z.; Xiong, W.; Li, G.; Li, X.; Yang, J.; Xiang, B.; Yi, M. Pyroptosis: A new paradigm of cell death for fighting against cancer. J. Exp. Clin. Cancer Res. 2021, 40, 153. [Google Scholar] [CrossRef]
- He, W.T.; Wan, H.; Hu, L.; Chen, P.; Wang, X.; Huang, Z.; Yang, Z.H.; Zhong, C.Q.; Han, J. Gasdermin D is an executor of pyroptosis and required for interleukin-1 β secretion. Cell Res. 2015, 25, 1285–1298. [Google Scholar] [CrossRef]
- Tan, G.; Huang, C.; Chen, J.; Zhi, F. HMGB1 released from GSDME-mediated pyroptotic epithelial cells participates in the tumorigenesis of colitis-associated colorectal cancer through the ERK1/2 pathway. J. Hematol. Oncol. 2020, 13, 149. [Google Scholar] [CrossRef]
- Liu, X.; Xia, S.; Zhang, Z.; Wu, H.; Lieberman, J. Channelling inflammation: Gasdermins in physiology and disease. Nat. Rev. Drug Discov. 2021, 20, 384–405. [Google Scholar] [CrossRef]
- Sarhan, J.; Liu, B.C.; Muendlein, H.I.; Li, P.; Nilson, R.; Tang, A.Y.; Rongvaux, A.; Bunnell, S.C.; Shao, F.; Green, D.R.; et al. Caspase-8 induces cleavage of gasdermin D to elicit pyroptosis during Yersinia infection. Proc. Natl. Acad. Sci. USA 2018, 115, E10888–E10897. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.; Wang, X.; Zheng, Y.; Jiang, J.; Hu, J. What role does pyroptosis play in microbial infection? J. Cell Physiol. 2019, 234, 7885–7892. [Google Scholar] [CrossRef]
- Luo, B.; Huang, F.; Liu, Y.; Liang, Y.; Wei, Z.; Ke, H.; Zeng, Z.; Huang, W.; He, Y. NLRP3 Inflammasome as a Molecular Marker in Diabetic Cardiomyopathy. Front. Physiol. 2017, 8, 519. [Google Scholar] [CrossRef] [Green Version]
- Luo, B.; Li, B.; Wang, W.; Liu, X.; Xia, Y.; Zhang, C.; Zhang, M.; Zhang, Y.; An, F. NLRP3 gene silencing ameliorates diabetic cardiomyopathy in a type 2 diabetes rat model. PLoS ONE 2014, 9, e104771. [Google Scholar] [CrossRef]
- Li, X.; Du, N.; Zhang, Q.; Li, J.; Chen, X.; Liu, X.; Hu, Y.; Qin, W.; Shen, N.; Xu, C.; et al. MicroRNA-30d regulates cardiomyocyte pyroptosis by directly targeting foxo3a in diabetic cardiomyopathy. Cell Death Dis. 2014, 5, e1479. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Zhang, H.; Qi, W.; Zhang, Y.; Li, J.; Li, Z.; Lin, Y.; Bai, X.; Liu, X.; Chen, X.; et al. Nicotine promotes atherosclerosis via ROS-NLRP3-mediated endothelial cell pyroptosis. Cell Death Dis. 2018, 9, 171. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X.; Pitzer, A.L.; Chen, Y.; Wang, L.; Li, P.L. Coronary endothelial dysfunction induced by nucleotide oligomerization domain-like receptor protein with pyrin domain containing 3 inflammasome activation during hypercholesterolemia: Beyond inflammation. Antioxid. Redox Signal. 2015, 22, 1084–1096. [Google Scholar] [CrossRef] [Green Version]
- Zhaolin, Z.; Jiaojiao, C.; Peng, W.; Yami, L.; Tingting, Z.; Jun, T.; Shiyuan, W.; Jinyan, X.; Dangheng, W.; Zhisheng, J.; et al. OxLDL induces vascular endothelial cell pyroptosis through miR-125a-5p/TET2 pathway. J. Cell Physiol. 2019, 234, 7475–7491. [Google Scholar] [CrossRef]
- Accarias, S.; Lugo-Villarino, G.; Foucras, G.; Neyrolles, O.; Boullier, S.; Tabouret, G. Pyroptosis of resident macrophages differentially orchestrates inflammatory responses to Staphylococcus aureus in resistant and susceptible mice. Eur. J. Immunol. 2015, 45, 794–806. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yuan, Y.H.; Chen, N.H.; Wang, H.B. The mechanisms of NLRP3 inflammasome/pyroptosis activation and their role in Parkinson’s disease. Int. Immunopharmacol. 2019, 67, 458–464. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, B.A.; Mamik, M.K.; Saito, L.B.; Boghozian, R.; Monaco, M.C.; Major, E.O.; Lu, J.Q.; Branton, W.G.; Power, C. Caspase-1 inhibition prevents glial inflammasome activation and pyroptosis in models of multiple sclerosis. Proc. Natl. Acad. Sci. USA 2018, 115, E6065–E6074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doitsh, G.; Galloway, N.L.; Geng, X.; Yang, Z.; Monroe, K.M.; Zepeda, O.; Hunt, P.W.; Hatano, H.; Sowinski, S.; Muñoz-Arias, I.; et al. Cell death by pyroptosis drives CD4 T-cell depletion in HIV-1 infection. Nature 2014, 505, 509–514. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Sharif, H.; Vora, S.M.; Zheng, Y.; Wu, H. Structures and functions of the inflammasome engine. J. Allergy Clin. Immunol. 2021, 147, 2021–2029. [Google Scholar] [CrossRef]
- Wang, Y.; Kong, H.; Zeng, X.; Liu, W.; Wang, Z.; Yan, X.; Wang, H.; Xie, W. Activation of NLRP3 inflammasome enhances the proliferation and migration of A549 lung cancer cells. Oncol. Rep. 2016, 35, 2053–2064. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Luo, Q.; Feng, X.; Zhang, R.; Li, J.; Chen, F. NLRP3 promotes tumor growth and metastasis in human oral squamous cell carcinoma. BMC Cancer 2018, 18, 500. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wang, Y.; Du, Q.; Lu, P.; Fan, H.; Lu, J.; Hu, R. Inflammasome-independent NLRP3 is required for epithelial-mesenchymal transition in colon cancer cells. Exp. Cell Res. 2016, 342, 184–192. [Google Scholar] [CrossRef]
- Feng, X.; Luo, Q.; Zhang, H.; Wang, H.; Chen, W.; Meng, G.; Chen, F. The role of NLRP3 inflammasome in 5-fluorouracil resistance of oral squamous cell carcinoma. J. Exp. Clin. Cancer Res. 2017, 36, 81. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Liu, Y. Aging-related gene signature regulated by Nlrp3 predicts glioma progression. Am. J. Cancer Res. 2015, 5, 442–449. [Google Scholar]
- Zhao, X.; Zhang, C.; Hua, M.; Wang, R.; Zhong, C.; Yu, J.; Han, F.; He, N.; Zhao, Y.; Liu, G.; et al. NLRP3 inflammasome activation plays a carcinogenic role through effector cytokine IL-18 in lymphoma. Oncotarget 2017, 8, 108571–108583. [Google Scholar] [CrossRef] [Green Version]
- Deswaerte, V.; Nguyen, P.; West, A.; Browning, A.F.; Yu, L.; Ruwanpura, S.M.; Balic, J.; Livis, T.; Girard, C.; Preaudet, A.; et al. Inflammasome Adaptor ASC Suppresses Apoptosis of Gastric Cancer Cells by an IL18-Mediated Inflammation-Independent Mechanism. Cancer Res. 2018, 78, 1293–1307. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Cheon, S.; Jung, M.K.; Song, S.B.; Kim, D.; Kim, H.J.; Park, H.; Bang, S.I.; Cho, D. Interleukin-18 enhances breast cancer cell migration via down-regulation of claudin-12 and induction of the p38 MAPK pathway. Biochem. Biophys. Res. Commun. 2015, 459, 379–386. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhu, M.; Lance, P. IL1β-mediated Stromal COX-2 signaling mediates proliferation and invasiveness of colonic epithelial cancer cells. Exp. Cell Res. 2012, 318, 2520–2530. [Google Scholar] [CrossRef]
- Tsai, C.Y.; Lee, T.S.; Kou, Y.R.; Wu, Y.L. Glucosamine inhibits IL-1beta-mediated IL-8 production in prostate cancer cells by MAPK attenuation. J. Cell Biochem. 2009, 108, 489–498. [Google Scholar] [CrossRef]
- Fathima, H.K.; Ramaswamy, P.; Nandakumar, D.N. IL-1β microenvironment promotes proliferation, migration, and invasion of human glioma cells. Cell Biol. Int. 2014, 38, 1415–1422. [Google Scholar] [CrossRef]
- Li, S.; Liang, X.; Ma, L.; Shen, L.; Li, T.; Zheng, L.; Sun, A.; Shang, W.; Chen, C.; Zhao, W.; et al. MiR-22 sustains NLRP3 expression and attenuates H. pylori-induced gastric carcinogenesis. Oncogene 2018, 37, 884–896. [Google Scholar] [CrossRef]
- Terme, M.; Ullrich, E.; Aymeric, L.; Meinhardt, K.; Desbois, M.; Delahaye, N.; Viaud, S.; Ryffel, B.; Yagita, H.; Kaplanski, G.; et al. IL-18 induces PD-1-dependent immunosuppression in cancer. Cancer Res. 2011, 71, 5393–5399. [Google Scholar] [CrossRef] [Green Version]
- Saijo, Y.; Tanaka, M.; Miki, M.; Usui, K.; Suzuki, T.; Maemondo, M.; Hong, X.; Tazawa, R.; Kikuchi, T.; Matsushima, K.; et al. Proinflammatory cytokine IL-1 beta promotes tumor growth of Lewis lung carcinoma by induction of angiogenic factors: In vivo analysis of tumor-stromal interaction. J. Immunol. 2002, 169, 469–475. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Z.; Liu, W.; Kaur, M.; Luo, Y.; Domenico, J.; Samson, J.M.; Shellman, Y.G.; Norris, D.A.; Dinarello, C.A.; Spritz, R.A.; et al. NLRP1 promotes tumor growth by enhancing inflammasome activation and suppressing apoptosis in metastatic melanoma. Oncogene 2017, 36, 3820–3830. [Google Scholar] [CrossRef] [Green Version]
- Wei, Q.; Mu, K.; Li, T.; Zhang, Y.; Yang, Z.; Jia, X.; Zhao, W.; Huai, W.; Guo, P.; Han, L. Deregulation of the NLRP3 inflammasome in hepatic parenchymal cells during liver cancer progression. Lab. Investig. 2014, 94, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Zaki, M.H.; Vogel, P.; Body-Malapel, M.; Lamkanfi, M.; Kanneganti, T.D. IL-18 production downstream of the Nlrp3 inflammasome confers protection against colorectal tumor formation. J. Immunol. 2010, 185, 4912–4920. [Google Scholar] [CrossRef] [Green Version]
- Dupaul-Chicoine, J.; Arabzadeh, A.; Dagenais, M.; Douglas, T.; Champagne, C.; Morizot, A.; Rodrigue-Gervais, I.G.; Breton, V.; Colpitts, S.L.; Beauchemin, N.; et al. The Nlrp3 Inflammasome Suppresses Colorectal Cancer Metastatic Growth in the Liver by Promoting Natural Killer Cell Tumoricidal Activity. Immunity 2015, 43, 751–763. [Google Scholar] [CrossRef] [Green Version]
- Williams, T.M.; Leeth, R.A.; Rothschild, D.E.; Coutermarsh-Ott, S.L.; McDaniel, D.K.; Simmons, A.E.; Heid, B.; Cecere, T.E.; Allen, I.C. The NLRP1 inflammasome attenuates colitis and colitis-associated tumorigenesis. J. Immunol. 2015, 194, 3369–3380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dihlmann, S.; Tao, S.; Echterdiek, F.; Herpel, E.; Jansen, L.; Chang-Claude, J.; Brenner, H.; Hoffmeister, M.; Kloor, M. Lack of Absent in Melanoma 2 (AIM2) expression in tumor cells is closely associated with poor survival in colorectal cancer patients. Int. J. Cancer 2014, 135, 2387–2396. [Google Scholar] [CrossRef]
- Nowarski, R.; Jackson, R.; Gagliani, N.; de Zoete, M.R.; Palm, N.W.; Bailis, W.; Low, J.S.; Harman, C.C.; Graham, M.; Elinav, E.; et al. Epithelial IL-18 Equilibrium Controls Barrier Function in Colitis. Cell 2015, 163, 1444–1456. [Google Scholar] [CrossRef] [Green Version]
- Bauer, C.; Duewell, P.; Mayer, C.; Lehr, H.A.; Fitzgerald, K.A.; Dauer, M.; Tschopp, J.; Endres, S.; Latz, E.; Schnurr, M. Colitis induced in mice with dextran sulfate sodium (DSS) is mediated by the NLRP3 inflammasome. Gut 2010, 59, 1192–1199. [Google Scholar] [CrossRef] [Green Version]
- Du, Q.; Wang, Q.; Fan, H.; Wang, J.; Liu, X.; Wang, H.; Wang, Y.; Hu, R. Dietary cholesterol promotes AOM-induced colorectal cancer through activating the NLRP3 inflammasome. Biochem. Pharmacol. 2016, 105, 42–54. [Google Scholar] [CrossRef]
- Janowski, A.M.; Colegio, O.R.; Hornick, E.E.; McNiff, J.M.; Martin, M.D.; Badovinac, V.P.; Norian, L.A.; Zhang, W.; Cassel, S.L.; Sutterwala, F.S. NLRC4 suppresses melanoma tumor progression independently of inflammasome activation. J. Clin. Investig. 2016, 126, 3917–3928. [Google Scholar] [CrossRef] [Green Version]
- Lin, P.H.; Lin, H.Y.; Kuo, C.C.; Yang, L.T. N-terminal functional domain of Gasdermin A3 regulates mitochondrial homeostasis via mitochondrial targeting. J. Biomed. Sci. 2015, 22, 44. [Google Scholar] [CrossRef] [Green Version]
- Lei, M.; Bai, X.; Yang, T.; Lai, X.; Qiu, W.; Yang, L.; Lian, X. Gsdma3 is a new factor needed for TNF-α-mediated apoptosis signal pathway in mouse skin keratinocytes. Histochem. Cell Biol. 2012, 138, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Jiang, X.; Gu, P.; Chen, W.; Zeng, X.; Gao, X. Gsdma3 mutation causes bulge stem cell depletion and alopecia mediated by skin inflammation. Am. J. Pathol. 2012, 180, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Runkel, F.; Marquardt, A.; Stoeger, C.; Kochmann, E.; Simon, D.; Kohnke, B.; Korthaus, D.; Wattler, F.; Fuchs, H.; Hrabé, D.A.M.; et al. The dominant alopecia phenotypes Bareskin, Rex-denuded, and Reduced Coat 2 are caused by mutations in gasdermin 3. Genomics 2004, 84, 824–835. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Rathkolb, B.; Budde, B.S.; Nürnberg, P.; de Angelis, M.H.; Aigner, B.; Schneider, M.R. Gsdma3(I359N) is a novel ENU-induced mutant mouse line for studying the function of Gasdermin A3 in the hair follicle and epidermis. J. Derm. Sci. 2012, 67, 190–192. [Google Scholar] [CrossRef]
- Yu, J.; Kang, M.J.; Kim, B.J.; Kwon, J.W.; Song, Y.H.; Choi, W.A.; Shin, Y.J.; Hong, S.J. Polymorphisms in GSDMA and GSDMB are associated with asthma susceptibility, atopy and BHR. Pediatr. Pulmonol. 2011, 46, 701–708. [Google Scholar] [CrossRef]
- Stein, M.M.; Thompson, E.E.; Schoettler, N.; Helling, B.A.; Magnaye, K.M.; Stanhope, C.; Igartua, C.; Morin, A.; Washington, C.R.; Nicolae, D.; et al. A decade of research on the 17q12-21 asthma locus: Piecing together the puzzle. J. Allergy Clin. Immunol. 2018, 142, 749–764.e3. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, M.A.; Matheson, M.C.; Tang, C.S.; Granell, R.; Ang, W.; Hui, J.; Kiefer, A.K.; Duffy, D.L.; Baltic, S.; Danoy, P.; et al. Genome-wide association analysis identifies 11 risk variants associated with the asthma with hay fever phenotype. J. Allergy Clin. Immunol. 2014, 133, 1564–1571. [Google Scholar] [CrossRef] [Green Version]
- Saeki, N.; Kim, D.H.; Usui, T.; Aoyagi, K.; Tatsuta, T.; Aoki, K.; Yanagihara, K.; Tamura, M.; Mizushima, H.; Sakamoto, H.; et al. GASDERMIN, suppressed frequently in gastric cancer, is a target of LMO1 in TGF-beta-dependent apoptotic signalling. Oncogene 2007, 26, 6488–6498. [Google Scholar] [CrossRef] [Green Version]
- Saeki, N.; Kuwahara, Y.; Sasaki, H.; Satoh, H.; Shiroishi, T. Gasdermin (Gsdm) localizing to mouse Chromosome 11 is predominantly expressed in upper gastrointestinal tract but significantly suppressed in human gastric cancer cells. Mamm. Genome. 2000, 11, 718–724. [Google Scholar] [CrossRef]
- Carl-McGrath, S.; Schneider-Stock, R.; Ebert, M.; Röcken, C. Differential expression and localisation of gasdermin-like (GSDML), a novel member of the cancer-associated GSDMDC protein family, in neoplastic and non-neoplastic gastric, hepatic, and colon tissues. Pathology 2008, 40, 13–24. [Google Scholar] [CrossRef]
- Sun, Q.; Yang, J.; Xing, G.; Sun, Q.; Zhang, L.; He, F. Expression of GSDML Associates with Tumor Progression in Uterine Cervix Cancer. Transl. Oncol. 2008, 1, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Hergueta-Redondo, M.; Sarrió, D.; Molina-Crespo, A.; Megias, D.; Mota, A.; Rojo-Sebastian, A.; García-Sanz, P.; Morales, S.; Abril, S.; Cano, A.; et al. Gasdermin-B promotes invasion and metastasis in breast cancer cells. PLoS ONE 2014, 9, e90099. [Google Scholar] [CrossRef] [Green Version]
- Molina-Crespo, A.; Cadete, A.; Sarrio, D.; Gámez-Chiachio, M.; Martinez, L.; Chao, K.; Olivera, A.; Gonella, A.; Díaz, E.; Palacios, J.; et al. Intracellular Delivery of an Antibody Targeting Gasdermin-B Reduces HER2 Breast Cancer Aggressiveness. Clin. Cancer Res. 2019, 25, 4846–4858. [Google Scholar] [CrossRef] [Green Version]
- Hergueta-Redondo, M.; Sarrio, D.; Molina-Crespo, A.; Vicario, R.; Bernadó-Morales, C.; Martínez, L.; Rojo-Sebastián, A.; Serra-Musach, J.; Mota, A.; Martínez-Ramírez, A.; et al. Gasdermin B expression predicts poor clinical outcome in HER2-positive breast cancer. Oncotarget 2016, 7, 56295–56308. [Google Scholar] [CrossRef] [Green Version]
- Miguchi, M.; Hinoi, T.; Shimomura, M.; Adachi, T.; Saito, Y.; Niitsu, H.; Kochi, M.; Sada, H.; Sotomaru, Y.; Ikenoue, T.; et al. Gasdermin C Is Upregulated by Inactivation of Transforming Growth Factor β Receptor Type II in the Presence of Mutated Apc, Promoting Colorectal Cancer Proliferation. PLoS ONE 2016, 11, e0166422. [Google Scholar] [CrossRef]
- Wei, J.; Xu, Z.; Chen, X.; Wang, X.; Zeng, S.; Qian, L.; Yang, X.; Ou, C.; Lin, W.; Gong, Z.; et al. Overexpression of GSDMC is a prognostic factor for predicting a poor outcome in lung adenocarcinoma. Mol. Med. Rep. 2020, 21, 360–370. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Zhao, R.; Xia, W.; Chang, C.-W.; You, Y.; Hsu, J.-M.; Nie, L.; Chen, Y.; Wang, Y.-C.; Liu, C.; et al. PD-L1-mediated gasdermin C expression switches apoptosis to pyroptosis in cancer cells and facilitates tumour necrosis. Nat. Cell Biol. 2020, 22, 1264–1275. [Google Scholar] [CrossRef]
- Saeki, N.; Usui, T.; Aoyagi, K.; Kim, D.H.; Sato, M.; Mabuchi, T.; Yanagihara, K.; Ogawa, K.; Sakamoto, H.; Yoshida, T.; et al. Distinctive expression and function of four GSDM family genes (GSDMA-D) in normal and malignant upper gastrointestinal epithelium. Genes Chromosomes Cancer 2009, 48, 261–271. [Google Scholar] [CrossRef]
- Gao, J.; Qiu, X.; Xi, G.; Liu, H.; Zhang, F.; Lv, T.; Song, Y. Downregulation of GSDMD attenuates tumor proliferation via the intrinsic mitochondrial apoptotic pathway and inhibition of EGFR/Akt signaling and predicts a good prognosis in non-small cell lung cancer. Oncol. Rep. 2018, 40, 1971–1984. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.J.; Chen, D.; Jiang, M.Z.; Xu, B.; Li, X.W.; Chu, Y.; Zhang, Y.J.; Mao, R.; Liang, J.; Fan, D.M. Downregulation of gasdermin D promotes gastric cancer proliferation by regulating cell cycle-related proteins. J. Dig. Dis. 2018, 19, 74–83. [Google Scholar] [CrossRef]
- Pizato, N.; Luzete, B.C.; Kiffer, L.; Corrêa, L.H.; de Oliveira, S.I.; Assumpção, J.; Ito, M.K.; Magalhães, K.G. Omega-3 docosahexaenoic acid induces pyroptosis cell death in triple-negative breast cancer cells. Sci. Rep. 2018, 8, 1952. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Wu, X.; Zhang, J.; Liu, L.; Sui, X.; Zhang, R.; Liu, W.; Shen, F.; Sun, Y.; Xi, X. α-NETA induces pyroptosis of epithelial ovarian cancer cells through the GSDMD/caspase-4 pathway. FASEB J. 2019, 33, 12760–12767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, J.F.; Mei, Q.B.; Zhou, X.G.; Tang, Y.; Xiong, R.; Qiu, W.Q.; Pan, R.; Law, B.Y.; Wong, V.K.; Yu, C.L.; et al. Polyphyllin VI Induces Caspase-1-Mediated Pyroptosis via the Induction of ROS/NF-κB/NLRP3/GSDMD Signal Axis in Non-Small Cell Lung Cancer. Cancers 2020, 12, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Li, K.; Lin, X.; Yao, Z.; Wang, S.; Xiong, X.; Ning, Z.; Wang, J.; Xu, X.; Jiang, Y.; et al. Metformin induces human esophageal carcinoma cell pyroptosis by targeting the miR-497/PELP1 axis. Cancer Lett. 2019, 450, 22–31. [Google Scholar] [CrossRef]
- Johnson, D.C.; Taabazuing, C.Y.; Okondo, M.C.; Chui, A.J.; Rao, S.D.; Brown, F.C.; Reed, C.; Peguero, E.; de Stanchina, E.; Kentsis, A.; et al. DPP8/DPP9 inhibitor-induced pyroptosis for treatment of acute myeloid leukemia. Nat. Med. 2018, 24, 1151–1156. [Google Scholar] [CrossRef]
- Zhang, M.J.; Gao, W.; Liu, S.; Siu, S.P.; Yin, M.; Ng, J.C.; Chow, V.L.; Chan, J.Y.; Wong, T.S. CD38 triggers inflammasome-mediated pyroptotic cell death in head and neck squamous cell carcinoma. Am. J. Cancer Res. 2020, 10, 2895–2908. [Google Scholar]
- Tang, J.; Bei, M.; Zhu, J.; Xu, G.; Chen, D.; Jin, X.; Huang, J.; Dong, J.; Shi, L.; Xu, L.; et al. Acute cadmium exposure induces GSDME-mediated pyroptosis in triple-negative breast cancer cells through ROS generation and NLRP3 inflammasome pathway activation. Env. Toxicol. Pharm. 2021, 87, 103686. [Google Scholar] [CrossRef]
- Wu, M.; Wang, Y.; Yang, D.; Gong, Y.; Rao, F.; Liu, R.; Danna, Y.; Li, J.; Fan, J.; Chen, J.; et al. A PLK1 kinase inhibitor enhances the chemosensitivity of cisplatin by inducing pyroptosis in oesophageal squamous cell carcinoma. EBioMedicine 2019, 41, 244–255. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.C.; Li, C.G.; Wang, Y.F.; Xu, L.H.; He, X.H.; Zeng, Q.Z.; Zeng, C.Y.; Mai, F.Y.; Hu, B.; Ouyang, D.Y. Chemotherapeutic paclitaxel and cisplatin differentially induce pyroptosis in A549 lung cancer cells via caspase-3/GSDME activation. Apoptosis 2019, 24, 312–325. [Google Scholar] [CrossRef]
- Wang, Y.; Yin, B.; Li, D.; Wang, G.; Han, X.; Sun, X. GSDME mediates caspase-3-dependent pyroptosis in gastric cancer. Biochem. Biophys. Res. Commun. 2018, 495, 1418–1425. [Google Scholar] [CrossRef]
- Erkes, D.A.; Cai, W.; Sanchez, I.M.; Purwin, T.J.; Rogers, C.; Field, C.O.; Berger, A.C.; Hartsough, E.J.; Rodeck, U.; Alnemri, E.S.; et al. Mutant BRAF and MEK Inhibitors Regulate the Tumor Immune Microenvironment via Pyroptosis. Cancer Discov. 2020, 10, 254–269. [Google Scholar] [CrossRef]
- Yu, P.; Wang, H.Y.; Tian, M.; Li, A.X.; Chen, X.S.; Wang, X.L.; Zhang, Y.; Cheng, Y. Eukaryotic elongation factor-2 kinase regulates the cross-talk between autophagy and pyroptosis in doxorubicin-treated human melanoma cells in vitro. Acta Pharm. Sin. 2019, 40, 1237–1244. [Google Scholar] [CrossRef]
- Van Laer, L.; Huizing, E.H.; Verstreken, M.; van Zuijlen, D.; Wauters, J.G.; Bossuyt, P.J.; Van de Heyning, P.; McGuirt, W.T.; Smith, R.J.; Willems, P.J.; et al. Nonsyndromic hearing impairment is associated with a mutation in DFNA5. Nat. Genet. 1998, 20, 194–197. [Google Scholar] [CrossRef]
- Cheng, J.; Han, D.Y.; Dai, P.; Sun, H.J.; Tao, R.; Sun, Q.; Yan, D.; Qin, W.; Wang, H.Y.; Ouyang, X.M.; et al. A novel DFNA5 mutation, IVS8+4 A>G, in the splice donor site of intron 8 causes late-onset non-syndromic hearing loss in a Chinese family. Clin. Genet. 2007, 72, 471–477. [Google Scholar] [CrossRef]
- Li-Yang, M.N.; Shen, X.F.; Wei, Q.J.; Yao, J.; Lu, Y.J.; Cao, X.; Xing, G.Q. IVS8+1 DelG, a Novel Splice Site Mutation Causing DFNA5 Deafness in a Chinese Family. Chin. Med. J. 2015, 128, 2510–2515. [Google Scholar] [CrossRef]
- Bischoff, A.M.; Luijendijk, M.W.; Huygen, P.L.; van Duijnhoven, G.; De Leenheer, E.M.; Oudesluijs, G.G.; Van Laer, L.; Cremers, F.P.; Cremers, C.W.; Kremer, H. A novel mutation identified in the DFNA5 gene in a Dutch family: A clinical and genetic evaluation. Audiol. Neurootol. 2004, 9, 34–46. [Google Scholar] [CrossRef]
- Chai, Y.; Chen, D.; Wang, X.; Wu, H.; Yang, T. A novel splice site mutation in DFNA5 causes late-onset progressive non-syndromic hearing loss in a Chinese family. Int. J. Pediatr. Otorhinolaryngol. 2014, 78, 1265–1268. [Google Scholar] [CrossRef]
- Booth, K.T.; Azaiez, H.; Kahrizi, K.; Wang, D.; Zhang, Y.; Frees, K.; Nishimura, C.; Najmabadi, H.; Smith, R.J. Exonic mutations and exon skipping: Lessons learned from DFNA5. Hum. Mutat. 2018, 39, 433–440. [Google Scholar] [CrossRef]
- Croes, L.; Beyens, M.; Fransen, E.; Ibrahim, J.; Vanden, B.W.; Suls, A.; Peeters, M.; Pauwels, P.; Van Camp, G.; Op, D.B.K. Large-scale analysis of DFNA5 methylation reveals its potential as biomarker for breast cancer. Clin. Epigenetics 2018, 10, 51. [Google Scholar] [CrossRef]
- Croes, L.; de Beeck, K.O.; Pauwels, P.; Vanden, B.W.; Peeters, M.; Fransen, E.; Van Camp, G. DFNA5 promoter methylation a marker for breast tumorigenesis. Oncotarget 2017, 8, 31948–31958. [Google Scholar] [CrossRef] [Green Version]
- Fujikane, T.; Nishikawa, N.; Toyota, M.; Suzuki, H.; Nojima, M.; Maruyama, R.; Ashida, M.; Ohe-Toyota, M.; Kai, M.; Nishidate, T.; et al. Genomic screening for genes upregulated by demethylation revealed novel targets of epigenetic silencing in breast cancer. Breast Cancer Res. Treat. 2010, 122, 699–710. [Google Scholar] [CrossRef]
- Kim, M.S.; Lebron, C.; Nagpal, J.K.; Chae, Y.K.; Chang, X.; Huang, Y.; Chuang, T.; Yamashita, K.; Trink, B.; Ratovitski, E.A.; et al. Methylation of the DFNA5 increases risk of lymph node metastasis in human breast cancer. Biochem. Biophys. Res. Commun. 2008, 370, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, J.; Op, D.B.K.; Fransen, E.; Croes, L.; Beyens, M.; Suls, A.; Vanden, B.W.; Peeters, M.; Van Camp, G. Methylation analysis of Gasdermin E shows great promise as a biomarker for colorectal cancer. Cancer Med. 2019, 8, 2133–2145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.S.; Chang, X.; Yamashita, K.; Nagpal, J.K.; Baek, J.H.; Wu, G.; Trink, B.; Ratovitski, E.A.; Mori, M.; Sidransky, D. Aberrant promoter methylation and tumor suppressive activity of the DFNA5 gene in colorectal carcinoma. Oncogene 2008, 27, 3624–3634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokomizo, K.; Harada, Y.; Kijima, K.; Shinmura, K.; Sakata, M.; Sakuraba, K.; Kitamura, Y.; Shirahata, A.; Goto, T.; Mizukami, H.; et al. Methylation of the DFNA5 gene is frequently detected in colorectal cancer. Anticancer Res. 2012, 32, 1319–1322. [Google Scholar]
- Akino, K.; Toyota, M.; Suzuki, H.; Imai, T.; Maruyama, R.; Kusano, M.; Nishikawa, N.; Watanabe, Y.; Sasaki, Y.; Abe, T.; et al. Identification of DFNA5 as a target of epigenetic inactivation in gastric cancer. Cancer Sci. 2007, 98, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Stoll, G.; Ma, Y.; Yang, H.; Kepp, O.; Zitvogel, L.; Kroemer, G. Pro-necrotic molecules impact local immunosurveillance in human breast cancer. Oncoimmunology 2017, 6, e1299302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.J.; Tang, L.; Shen, D.W.; Wang, C.; Yuan, Q.Y.; Gao, W.; Wang, Y.K.; Xu, R.H.; Zhang, H. The expression and regulation of DFNA5 in human hepatocellular carcinoma DFNA5 in hepatocellular carcinoma. Mol. Biol. Rep. 2013, 40, 6525–6531. [Google Scholar] [CrossRef] [PubMed]
- Rogers, C.; Erkes, D.A.; Nardone, A.; Aplin, A.E.; Fernandes-Alnemri, T.; Alnemri, E.S. Gasdermin pores permeabilize mitochondria to augment caspase-3 activation during apoptosis and inflammasome activation. Nat. Commun. 2019, 10, 1689. [Google Scholar] [CrossRef] [PubMed]
- Lage, H.; Helmbach, H.; Grottke, C.; Dietel, M.; Schadendorf, D. DFNA5 (ICERE-1) contributes to acquired etoposide resistance in melanoma cells. FEBS Lett. 2001, 494, 54–59. [Google Scholar] [CrossRef] [Green Version]
- Croes, L.; Fransen, E.; Hylebos, M.; Buys, K.; Hermans, C.; Broeckx, G.; Peeters, M.; Pauwels, P.; Op, D.B.K.; Van Camp, G. Determination of the Potential Tumor-Suppressive Effects of Gsdme in a Chemically Induced and in a Genetically Modified Intestinal Cancer Mouse Model. Cancers 2019, 11, 1214. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Zhang, S.; Wu, J.; Chen, M.; Cai, M.C.; Fu, Y.; Li, W.; Wang, J.; Zhao, X.; Yu, Z.; et al. Molecular Targeted Therapies Elicit Concurrent Apoptotic and GSDME-Dependent Pyroptotic Tumor Cell Death. Clin. Cancer Res. 2018, 24, 6066–6077. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gao, W.; Shi, X.; Ding, J.; Liu, W.; He, H.; Wang, K.; Shao, F. Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature 2017, 547, 99–103. [Google Scholar] [CrossRef]
- Zhou, B.; Zhang, J.Y.; Liu, X.S.; Chen, H.Z.; Ai, Y.L.; Cheng, K.; Sun, R.Y.; Zhou, D.; Han, J.; Wu, Q. Tom20 senses iron-activated ROS signaling to promote melanoma cell pyroptosis. Cell Res. 2018, 28, 1171–1185. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhang, Y.; Xia, S.; Kong, Q.; Li, S.; Liu, X.; Junqueira, C.; Meza-Sosa, K.F.; Mok, T.; Ansara, J.; et al. Gasdermin E suppresses tumour growth by activating anti-tumour immunity. J. Nat. 2020, 579, 415–420. [Google Scholar] [CrossRef]
- Vishwakarma, M.; Piddini, E. Outcompeting cancer. Nat. Rev. Cancer 2020, 20, 187–198. [Google Scholar] [CrossRef]
- Cox, T.R. The matrix in cancer. Nat. Rev. Cancer 2021, 21, 217–238. [Google Scholar] [CrossRef]
- Levine, A.J. p53: 800 million years of evolution and 40 years of discovery. Nat. Rev. Cancer 2020, 20, 471–480. [Google Scholar] [CrossRef]
- Vetrie, D.; Helgason, G.V.; Copland, M. The leukaemia stem cell: Similarities, differences and clinical prospects in CML and AML. Nat. Rev. Cancer 2020, 20, 158–173. [Google Scholar] [CrossRef]
- Jiang, M.; Qi, L.; Li, L.; Li, Y. The caspase-3/GSDME signal pathway as a switch between apoptosis and pyroptosis in cancer. Cell Death Discov. 2020, 6, 112. [Google Scholar] [CrossRef]
- Zheng, X.; Zhong, T.; Ma, Y.; Wan, X.; Qin, A.; Yao, B.; Zou, H.; Song, Y.; Yin, D. Bnip3 mediates doxorubicin-induced cardiomyocyte pyroptosis via caspase-3/GSDME. Life Sci. 2020, 242, 117186. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Wang, H.; Weng, C.; Jiang, H.; Chen, J. Caspase 3/GSDME-dependent pyroptosis contributes to chemotherapy drug-induced nephrotoxicity. Cell Death Dis. 2021, 12, 186. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, Y.; Ding, J.; Wang, C.; Zhou, X.; Gao, W.; Huang, H.; Shao, F.; Liu, Z. A bioorthogonal system reveals antitumour immune function of pyroptosis. Nature 2020, 579, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; He, H.; Wang, K.; Shi, X.; Wang, Y.; Su, Y.; Wang, Y.; Li, D.; Liu, W.; Zhang, Y.; et al. Granzyme A from cytotoxic lymphocytes cleaves GSDMB to trigger pyroptosis in target cells. Science 2020, 368, 6494. [Google Scholar] [CrossRef]
- Liu, Y.; Fang, Y.; Chen, X.; Wang, Z.; Liang, X.; Zhang, T.; Liu, M.; Zhou, N.; Lv, J.; Tang, K.; et al. Gasdermin E-mediated target cell pyroptosis by CAR T cells triggers cytokine release syndrome. Sci. Immunol. 2020, 5, 43. [Google Scholar] [CrossRef]
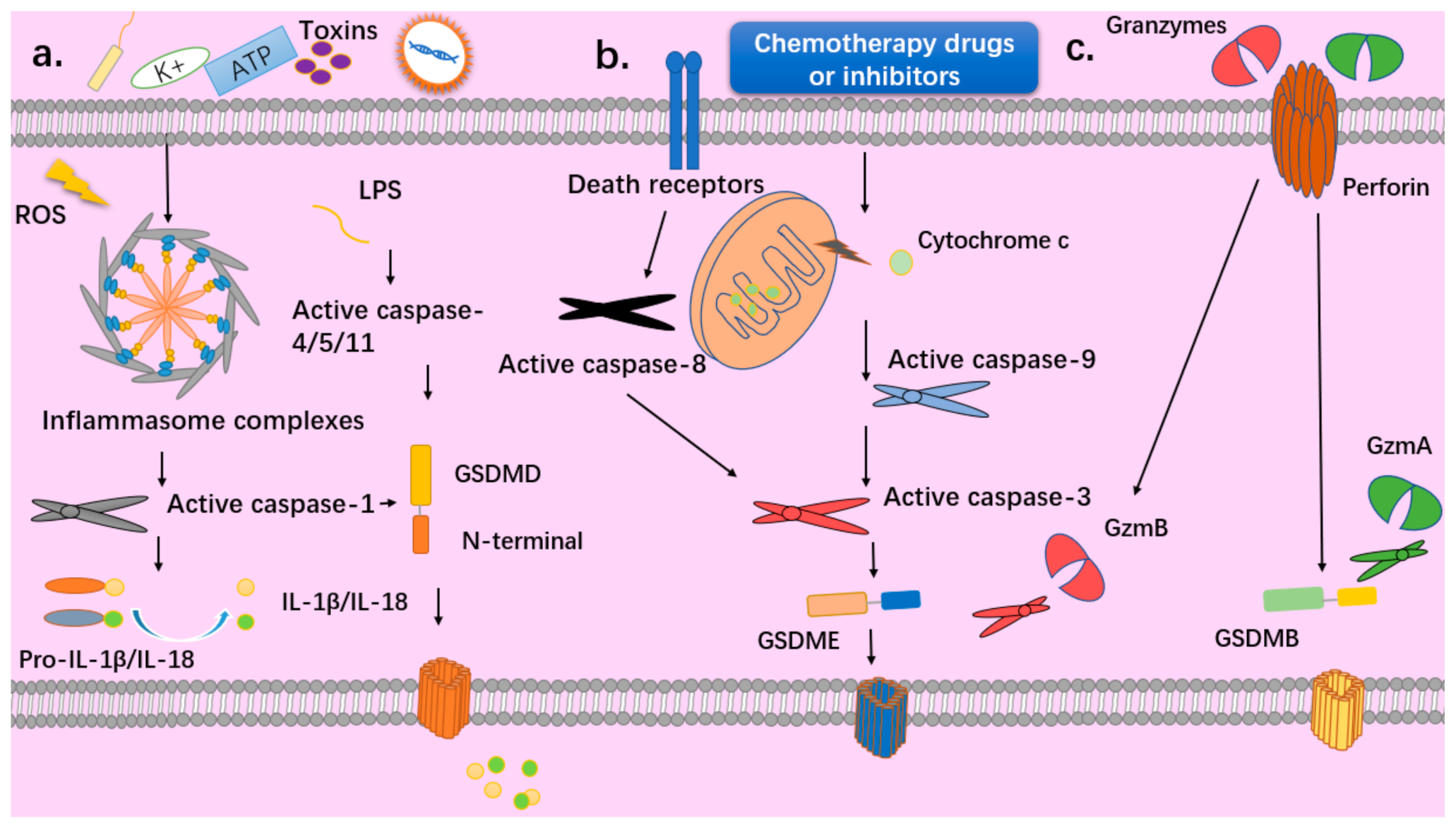
Figure 1.
Different pathways to pyroptosis. (a) Canonical pyroptosis: PAMPs such as Gram-negative bacteria, viruses, toxins and DAMPs such as intracellular ROS, ATP, potassium, cadmium can activate the inflammasomes and activated caspase 1 can cleave GSDMD and pro-IL-1β/IL-18. Non-canonical pyroptosis: Bacterial lipopolysaccharide (LPS) directly binds to and activates pro-caspase 4/5 or murine pro-caspase 11, then the activated caspases cleave GSDMD. (b) Chemotherapeutic drugs or inhibitors can disrupt the mitochondrial membrane and the release of cytochrome c activates caspase 9 and caspase 3 to cleave GSDME and trigger pyroptosis. Caspase 3 could also be activated by caspase 8 when death receptors are stimulated. (c) Cytotoxic T lymphocytes (CTLs) and natural killer (NK) cells release perforin to deliver serine protease granzymes (Gzms) into target cells, and then GzmA and GzmB can cleave GSDMB/E, which triggers pyroptosis.
Figure 1.
Different pathways to pyroptosis. (a) Canonical pyroptosis: PAMPs such as Gram-negative bacteria, viruses, toxins and DAMPs such as intracellular ROS, ATP, potassium, cadmium can activate the inflammasomes and activated caspase 1 can cleave GSDMD and pro-IL-1β/IL-18. Non-canonical pyroptosis: Bacterial lipopolysaccharide (LPS) directly binds to and activates pro-caspase 4/5 or murine pro-caspase 11, then the activated caspases cleave GSDMD. (b) Chemotherapeutic drugs or inhibitors can disrupt the mitochondrial membrane and the release of cytochrome c activates caspase 9 and caspase 3 to cleave GSDME and trigger pyroptosis. Caspase 3 could also be activated by caspase 8 when death receptors are stimulated. (c) Cytotoxic T lymphocytes (CTLs) and natural killer (NK) cells release perforin to deliver serine protease granzymes (Gzms) into target cells, and then GzmA and GzmB can cleave GSDMB/E, which triggers pyroptosis.

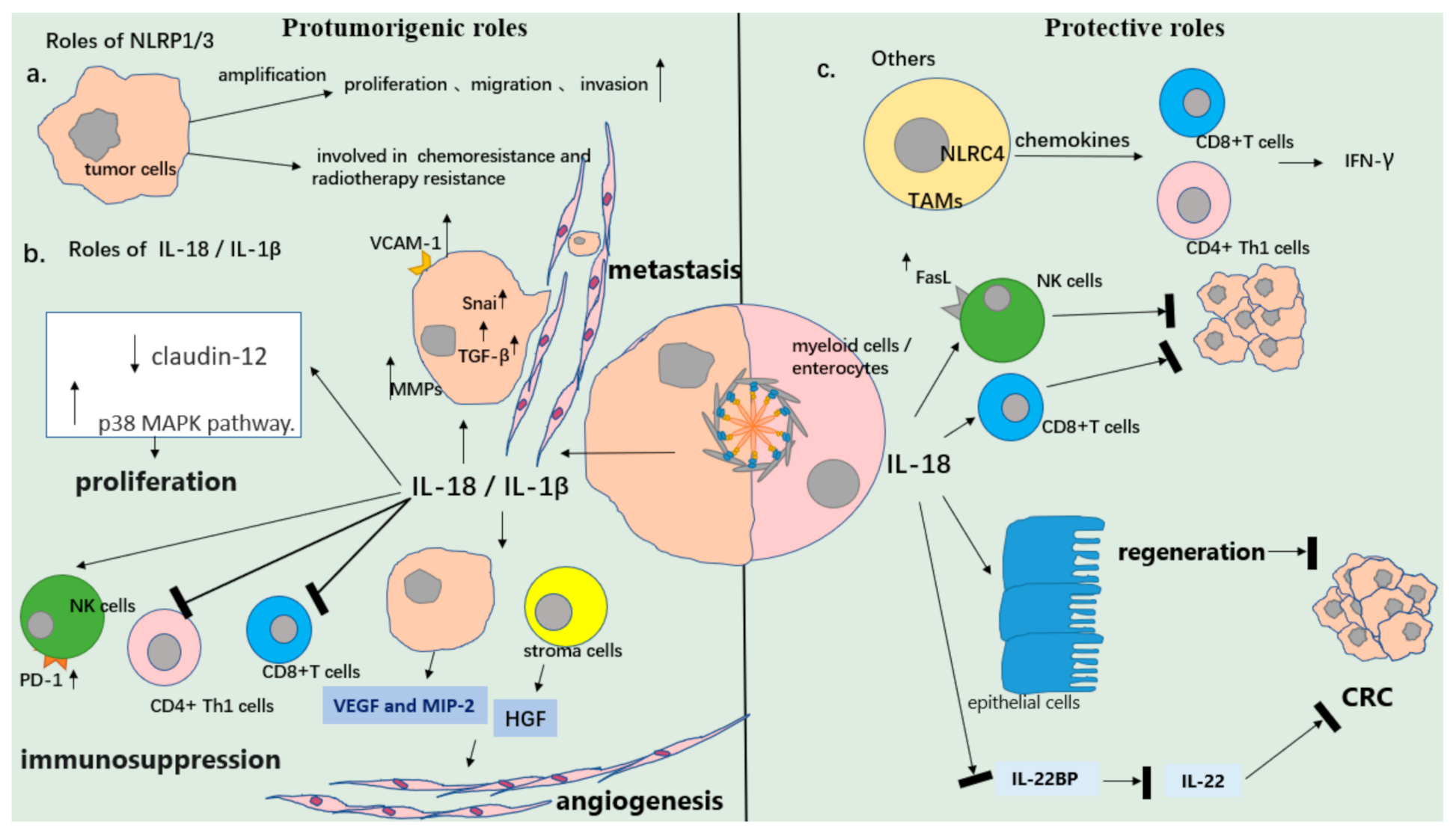
Figure 2.
Diverse roles of the inflammasome in cancer. (a) NLRP3 mediates the proliferation and invasion of tumor cells. A higher expression of NLRP3 in tumor is associated with the clinical outcomes of patients. (b) The release of IL-18 /IL-1βfrom tumor cells induces tumor proliferation, immunosuppression, angiogenesis, and metastasis. However, IL-18 from myeloid cells or enterocytes can activate cytolytic immune cells and inhibit the formation of tumors (colorectal cancer) via initiating tissue repair. (c) Others. Inflammasomes such as NLRC4 in tumor-associated macrophages (TAMs) can inhibit tumor progression.
Figure 2.
Diverse roles of the inflammasome in cancer. (a) NLRP3 mediates the proliferation and invasion of tumor cells. A higher expression of NLRP3 in tumor is associated with the clinical outcomes of patients. (b) The release of IL-18 /IL-1βfrom tumor cells induces tumor proliferation, immunosuppression, angiogenesis, and metastasis. However, IL-18 from myeloid cells or enterocytes can activate cytolytic immune cells and inhibit the formation of tumors (colorectal cancer) via initiating tissue repair. (c) Others. Inflammasomes such as NLRC4 in tumor-associated macrophages (TAMs) can inhibit tumor progression.


{kind=link}
{kind=link}
Table 1.
Roles of the NLRP3 inflammasome in cancer.
| Tumor Types | Significance | Experimental Models | Results | Refs. |
|---|---|---|---|---|
| Lung cancer | tumor promoter | LC cell lines | The activation of NLRP3 increased the tumor proliferation and migration. | [36] |
| OSCC | tumor promoter | OSCC cell lines and tissue | The enhanced expression of NLRP3 was correlated with tumor growth and metastasis. | [37] |
| The upregulation of NLRP3 correlates with the chemoresistance of 5-FU in OSCC. | [39] | |||
| Colon cancer | tumor promoter | CRC cell lines | The NLRP3 level was increased during EMT, which was independent of inflammasome activation. | [38] |
| Glioma | tumor promoter | glioma cell lines | NLRP3 inflammasome contributed to the radiotherapy resistance in a xenograft mouse glioblastoma model. | [40] |
| Liver cancer | tumor suppressor | HCC tissue | The deficiency of the NLRP3 inflammasome was related to higher pathological grades and advanced clinical stages in HCC. | [51] |
| Colorectal cancer | tumor suppressor | Nlrp3−/−, Asc−/− and Caspase1−/− mice | NLRP3 inflammasome protected mice from colitis-associated colorectal tumorigenesis. | [52] |
| macrophages surrounded CRC tissue | The NLRP3 inflammasome inhibited the growth of liver colon cancer metastatic tumor. | [53] |
Abbreviations: LC, lung cancer; OSCC, oral squamous cell carcinoma; CRC, colorectal cancer; EMT, epithelial-mesenchymal transition; HCC, hepatocellular carcinoma.
Table 2.
Induction of pyroptosis in cancer by various therapeutic reagents.
| Therapeutic Reagents | Cancer Type | Pathway | Refs. |
|---|---|---|---|
| Docosahexaenoic acid (DHA) | Breast cancer | NF-κB/Caspase-1/GSDMD | [81] |
| α-NETA | Ovarian cancer | GSDMD/caspase-4 | [82] |
| Polyphyllin VI (PPVI) | NSCLC | ROS/NF-κB/NLRP3/GSDMD | [83] |
| Metformin | ESCC | miR-497/PELP1/GSDMD | [84] |
| DPP8/9 | AML | CARD8/Caspase-1/GSDMD | [85] |
| Cadmium | Breast cancer | [87] | |
| BI2536 and Cisplatin | ESCC | [88] | |
| Paclitaxel and cisplatin | Lung cancer | Caspase-3/GSDME | [89] |
| 5-FU | Gastric cancer | [90] | |
| BRAFi + MEKi | Melanoma | [91] | |
| Doxorubicin | Melanoma | eEF-2K/GSDME | [92] |
Table 3.
Roles of gasdermin family members in tumor immunity.
| Gasdermin Family Members | Roles in Tumor Immunity | Refs. |
|---|---|---|
| GSDMA3 | A small fraction of pyroptotic tumor cells brought about the increased infiltration of cytotoxic T cells and CD4+ T helper cells and other immunological changes within the tumor. | [123] |
| GSDMB | Th expression of GSDMB wild type (WT) promoted cytotoxic T lymphocyte-mediated tumor clearance by antibodies to programmed cell death 1 (PD-1). | [124] |
| GSDME | The ectopic expression of GSDME inhibited tumor growth and enhanced tumor immunity with more and functional tumor-infiltrating natural-killer and CD8+ T lymphocytes. | [115] |
| GSDME | Treatment with BRAF inhibitors and MEK inhibitors in mouse melanoma promoted pyroptosis and the release of HMGB1, which enhanced the levels of tumor-associated T cells and damped dendritic cell infiltrates. | [91] |
| GSDME | Large amounts of damage-associated molecular pattern molecules (DAMPs) from pyroptotic cancer cells activate caspase 1 for GSDMD cleavage in macrophages, resulting in the release of proinflammatory cytokines and subsequent CRS. | [125] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lu, X.; Guo, T.; Zhang, X. Pyroptosis in Cancer: Friend or Foe? Cancers 2021, 13, 3620. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13143620
AMA Style
Lu X, Guo T, Zhang X. Pyroptosis in Cancer: Friend or Foe? Cancers. 2021; 13(14):3620. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13143620
Chicago/Turabian StyleLu, Xiuxia, Tianhui Guo, and Xing Zhang. 2021. "Pyroptosis in Cancer: Friend or Foe?" Cancers 13, no. 14: 3620. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13143620
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.