
SUMOylation Is Associated with Aggressive Behavior in Chondrosarcoma of Bone

 and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material & Methods
2.1. Compound
2.2. Tissue Microarray (TMA) and Clinicopathological Data
2.3. Immunohistochemistry
2.4. Analysis of Chondrosarcoma TMA Data Set
2.5. Analysis of Chondrosarcoma mRNA Expression Data Set
2.6. Analysis of Soft Tissue Sarcoma Data from the TCGA Repository
2.7. Cell Culture
2.8. Cell Viability Assay
2.9. Colony Formation Assay
2.10. Western Blot
2.11. Cell Cycle Analysis by Flow Cytometry
2.12. Microscopy
2.13. Statistical Analysis
3. Results
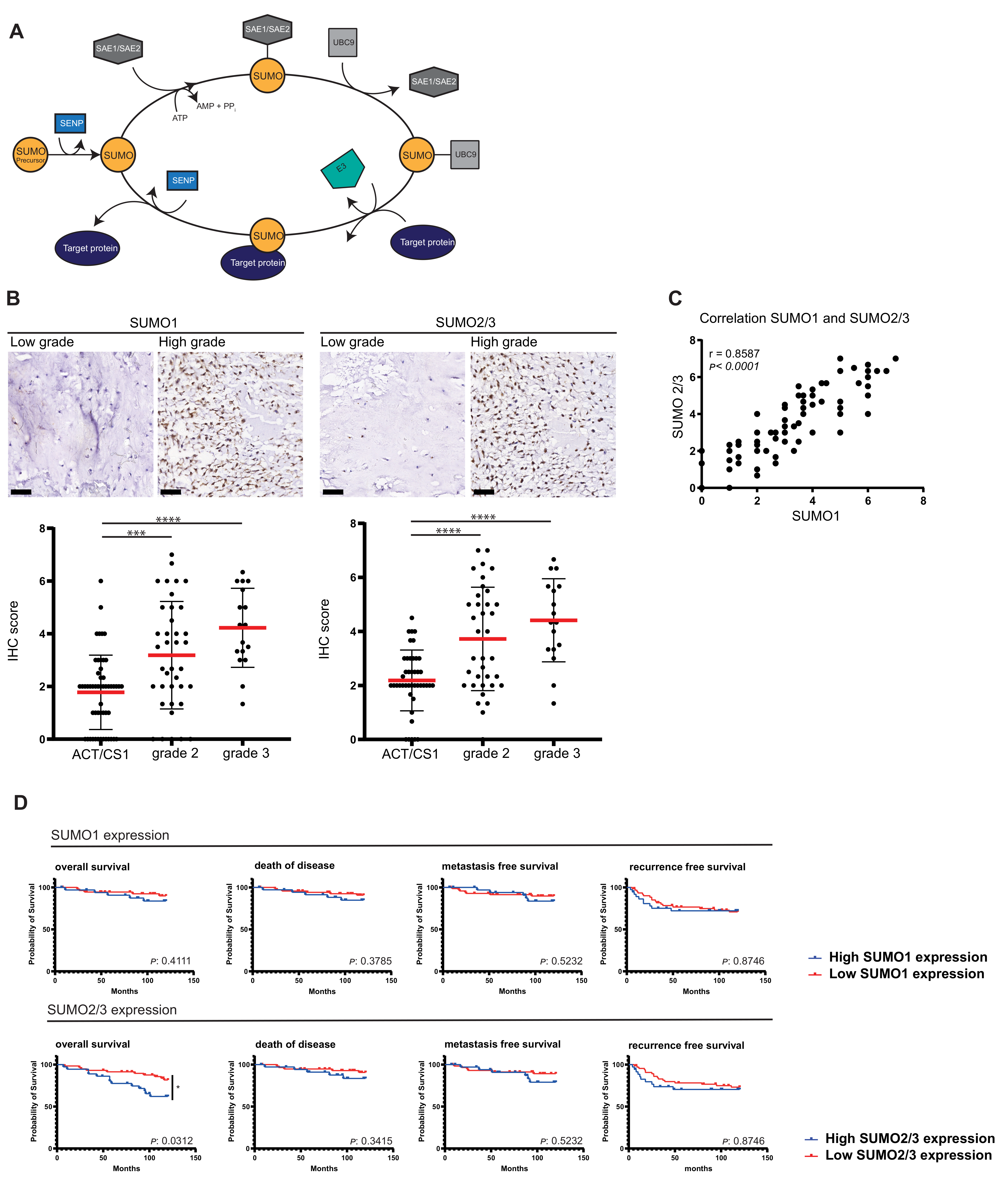
3.1. High Expression Levels of SUMO1–3 Correlate with Aggressiveness in Chondrosarcoma
3.2. High SUMO Expression Correlates with Poorer Survival
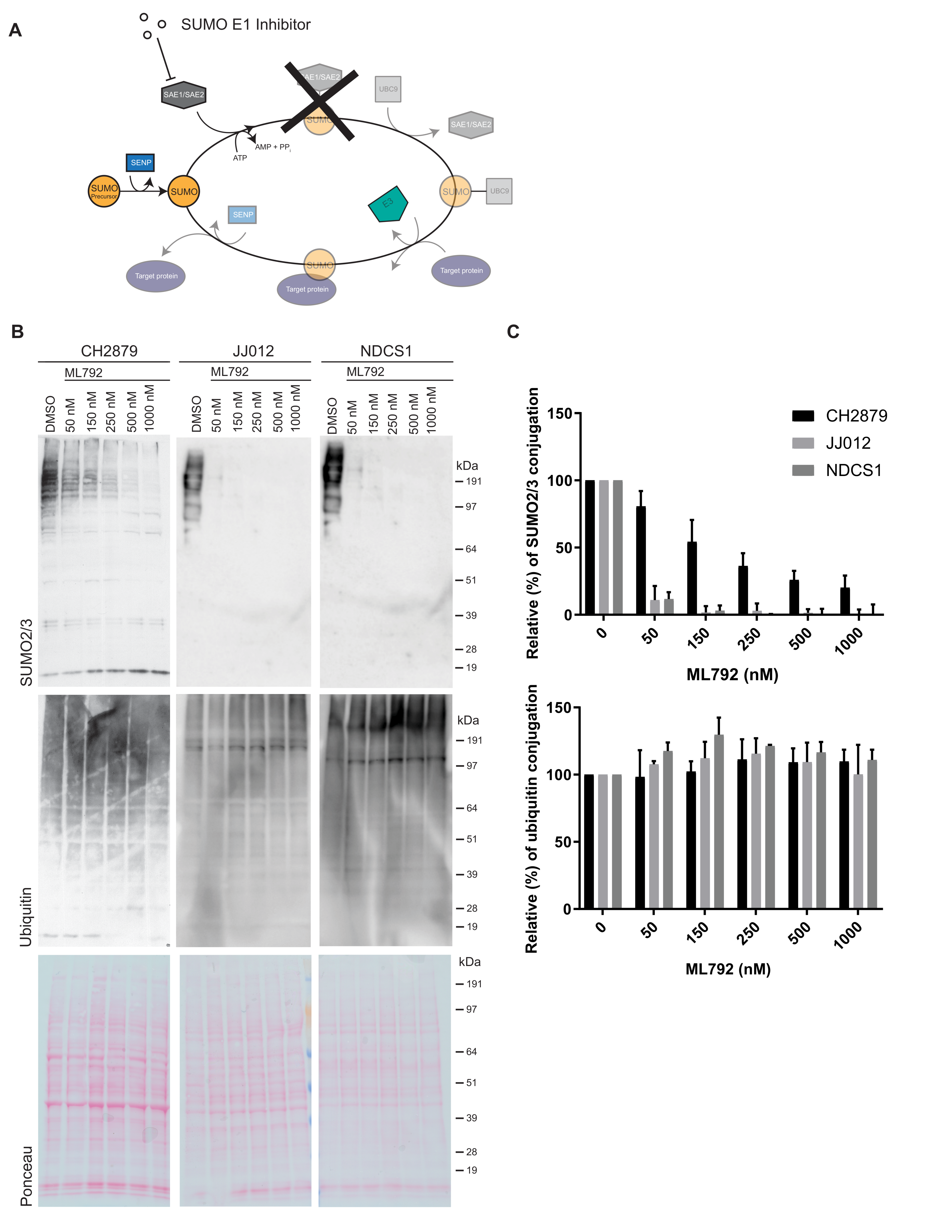
3.3. SUMO E1 Inhibition Decreases SUMO-Conjugation in Chondrosarcoma Cell Lines
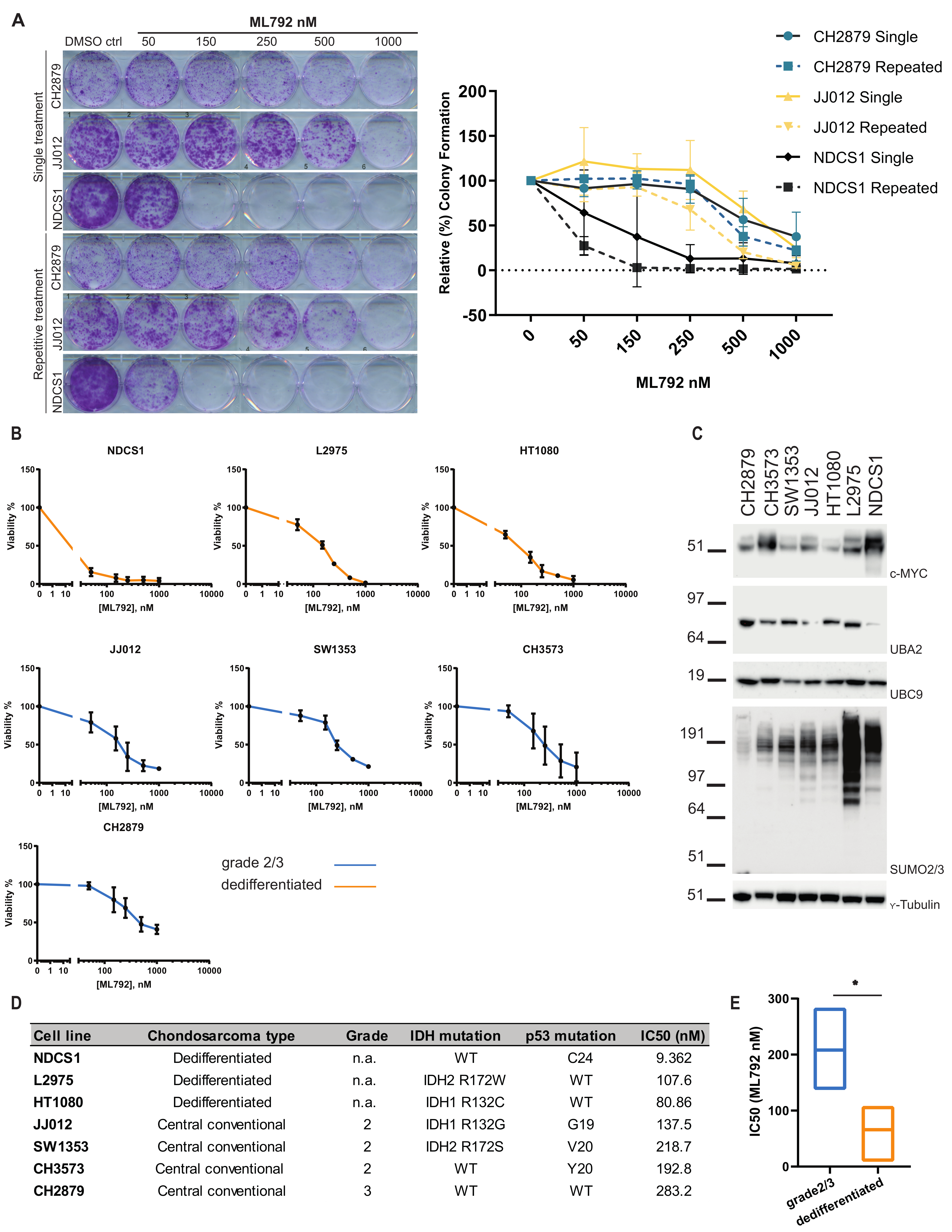
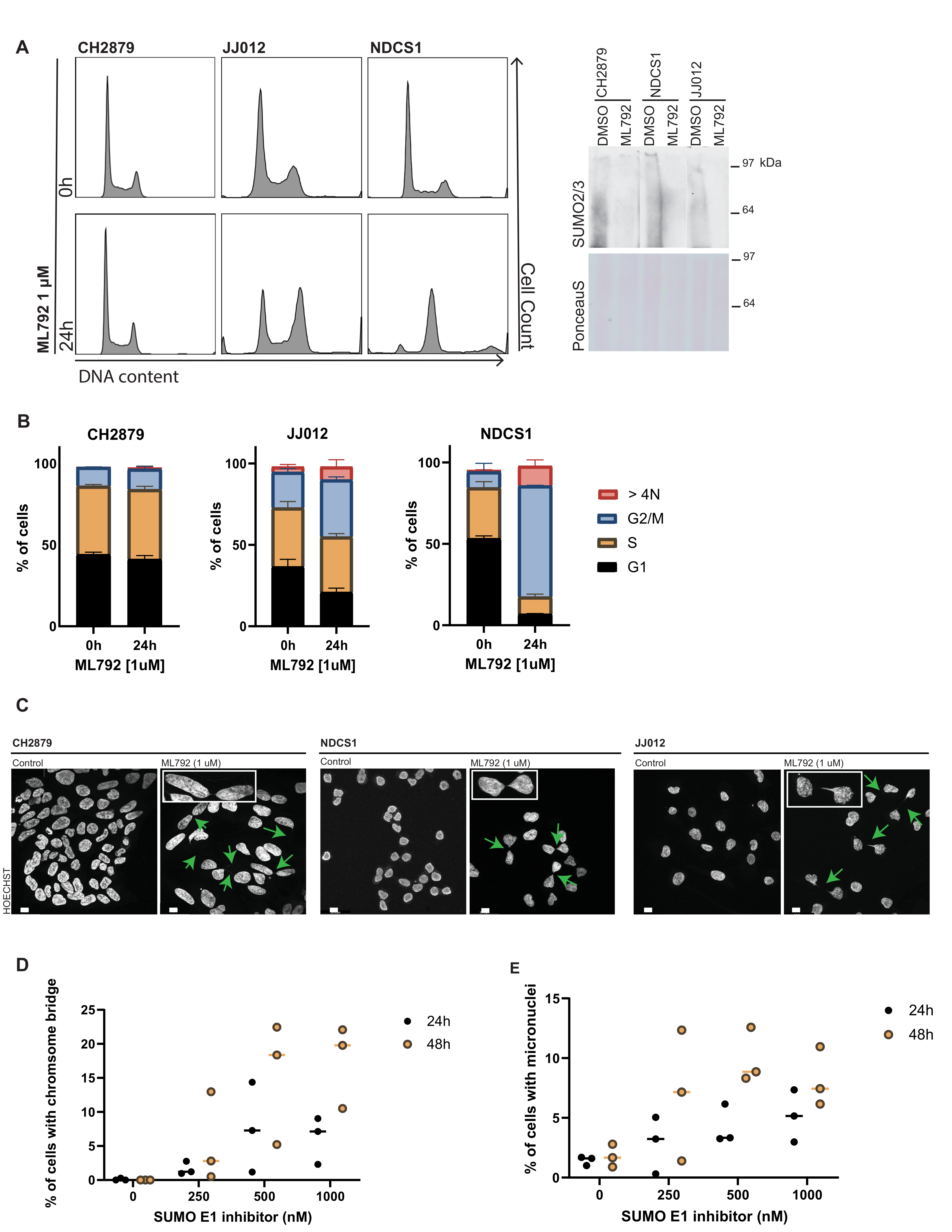
3.4. SUMO E1 Inhibition Reduces Cell Proliferation and Viability In Vitro
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cappadocia, L.; Lima, C.D. Ubiquitin-like Protein Conjugation: Structures, Chemistry, and Mechanism. Chem. Rev. 2018, 118, 889–918. [Google Scholar] [CrossRef]
- Flotho, A.; Melchior, F. Sumoylation: A regulatory protein modification in health and disease. Annu. Rev. Biochem. 2013, 82, 357–385. [Google Scholar] [CrossRef]
- Wang, L.; Wansleeben, C.; Zhao, S.; Miao, P.; Paschen, W.; Yang, W. SUMO2 is essential while SUMO3 is dispensable for mouse embryonic development. EMBO Rep. 2014, 15, 878–885. [Google Scholar] [CrossRef] [Green Version]
- Melchior, F.; Schergaut, M.; Pichler, A. SUMO: Ligases, isopeptidases and nuclear pores. Trends Biochem. Sci. 2003, 28, 612–618. [Google Scholar] [CrossRef]
- Muller, S.; Ledl, A.; Schmidt, D. SUMO: A regulator of gene expression and genome integrity. Oncogene 2004, 23, 1998–2008. [Google Scholar] [CrossRef] [Green Version]
- Jackson, S.P.; Durocher, D. Regulation of DNA damage responses by ubiquitin and SUMO. Mol. Cell 2013, 49, 795–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eifler, K.; Vertegaal, A.C.O. SUMOylation-Mediated Regulation of Cell Cycle Progression and Cancer. Trends Biochem. Sci. 2015, 40, 779–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeler, J.S.; Dejean, A. SUMO and the robustness of cancer. Nat. Rev. Cancer 2017, 17, 184–197. [Google Scholar] [CrossRef] [PubMed]
- Nacerddine, K.; Lehembre, F.; Bhaumik, M.; Artus, J.; Cohen-Tannoudji, M.; Babinet, C.; Pandolfi, P.P.; Dejean, A. The SUMO pathway is essential for nuclear integrity and chromosome segregation in mice. Dev. Cell 2005, 9, 769–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelisch, F.; Sonneville, R.; Pourkarimi, E.; Agostinho, A.; Blow, J.J.; Gartner, A.; Hay, R.T. Dynamic SUMO modification regulates mitotic chromosome assembly and cell cycle progression in Caenorhabditis elegans. Nat. Commun. 2014, 5, 5485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.J.; Hochstrasser, M. A new protease required for cell-cycle progression in yeast. Nature 1999, 398, 246–251. [Google Scholar] [CrossRef]
- Mukhopadhyay, D.; Arnaoutov, A.; Dasso, M. The SUMO protease SENP6 is essential for inner kinetochore assembly. J. Cell Biol. 2010, 188, 681–6692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Bacco, A.; Ouyang, J.; Lee, H.Y.; Catic, A.; Ploegh, H.; Gill, G. The SUMO-specific protease SENP5 is required for cell division. Mol. Cell. Biol. 2006, 26, 4489–4498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liebelt, F.; Jansen, N.S.; Kumar, S.; Gracheva, E.; Claessens, L.A.; Verlaan-de Vries, M.; Willemstein, E.; Vertegaal, A.C.O. The poly-SUMO2/3 protease SENP6 enables assembly of the constitutive centromere-associated network by group deSUMOylation. Nat. Commun. 2019, 10, 3987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendriks, I.A.; Vertegaal, A.C. A comprehensive compilation of SUMO proteomics. Nat. Rev. Mol. Cell Biol. 2016, 17, 581–595. [Google Scholar] [CrossRef]
- Schimmel, J.; Eifler, K.; Sigurethsson, J.O.; Cuijpers, S.A.; Hendriks, I.A.; Verlaan-de Vries, M.; Kelstrup, C.D.; Francavilla, C.; Medema, R.H.; Olsen, J.V.; et al. Uncovering SUMOylation dynamics during cell-cycle progression reveals FoxM1 as a key mitotic SUMO target protein. Mol. Cell 2014, 53, 1053–1066. [Google Scholar] [CrossRef] [Green Version]
- Myatt, S.S.; Kongsema, M.; Man, C.W.Y.; Kelly, D.J.; Gomes, A.R.; Khongkow, P.; Karunarathna, U.; Zona, S.; Langer, J.K.; Dunsby, C.W.; et al. SUMOylation inhibits FOXM1 activity and delays mitotic transition. Oncogene 2014, 33, 4316–4326. [Google Scholar] [CrossRef] [Green Version]
- Agostinho, M.; Santos, V.; Ferreira, F.; Costa, R.; Cardoso, J.; Pinheiro, I.; Rino, J.; Jaffray, E.; Hay, R.T.; Ferreira, J. Conjugation of human topoisomerase 2 alpha with small ubiquitin-like modifiers 2/3 in response to topoisomerase inhibitors: Cell cycle stage and chromosome domain specificity. Cancer Res. 2008, 68, 2409–2418. [Google Scholar] [CrossRef] [Green Version]
- Azuma, Y.; Arnaoutov, A.; Dasso, M. SUMO-2/3 regulates topoisomerase II in mitosis. J. Cell Biol. 2003, 163, 477–4487. [Google Scholar] [CrossRef]
- Yoshida, M.M.; Ting, L.; Gygi, S.P.; Azuma, Y. SUMOylation of DNA topoisomerase IIalpha regulates histone H3 kinase Haspin and H3 phosphorylation in mitosis. J. Cell Biol. 2016, 213, 665–678. [Google Scholar] [CrossRef] [Green Version]
- Cuijpers, S.A.G.; Vertegaal, A.C.O. Guiding Mitotic Progression by Crosstalk between Post-translational Modifications. Trends Biochem. Sci. 2018, 43, 251–268. [Google Scholar] [CrossRef]
- Lee, C.C.; Li, B.; Yu, H.; Matunis, M.J. Sumoylation promotes optimal APC/C Activation and Timely Anaphase. eLife 2018, 7, e29539. [Google Scholar] [CrossRef] [PubMed]
- Eifler, K.; Cuijpers, S.A.G.; Willemstein, E.; Raaijmakers, J.A.; El Atmioui, D.; Ovaa, H.; Medema, R.H.; Vertegaal, A.C.O. SUMO targets the APC/C to regulate transition from metaphase to anaphase. Nat. Commun. 2018, 9, 1119. [Google Scholar] [CrossRef]
- Yatskevich, S.; Kroonen, J.S.; Alfieri, C.; Tischer, T.; Howes, A.C.; Clijsters, L.; Yang, J.; Zhang, Z.; Yan, K.; Vertegaal, A.C.O.; et al. Molecular mechanisms of APC/C release from spindle assembly checkpoint inhibition by APC/C SUMOylation. Cell Rep. 2021, 34. [Google Scholar] [CrossRef]
- WCoTE, B. Soft Tissue and Bone Tumours: WHO Classification of Tumours; IARC: Lyon, France, 2020; Volume 3. [Google Scholar]
- van Praag, V.M.; Rueten-Budde, A.J.; Ho, V.; Dijkstra, P.D.S.; van der Geest, I.C.; Bramer, J.A.; Schaap, G.R.; Jutte, P.C.; Schreuder, H.W.B.; Ploegmakers, J.J.W.; et al. Incidence, outcomes and prognostic factors during 25 years of treatment of chondrosarcomas. Surg. Oncol. 2018, 27, 402–408. [Google Scholar] [CrossRef]
- Bovee, J.V.; Hogendoorn, P.C.; Wunder, J.S.; Alman, B.A. Cartilage tumours and bone development: Molecular pathology and possible therapeutic targets. Nat. Rev. Cancer 2010, 10, 481–488. [Google Scholar] [CrossRef]
- Amary, M.F.; Bacsi, K.; Maggiani, F.; Damato, S.; Halai, D.; Berisha, F.; Pollock, R.; O’Donnell, P.; Grigoriadis, A.; Diss, T.; et al. IDH1 and IDH2 mutations are frequent events in central chondrosarcoma and central and periosteal chondromas but not in other mesenchymal tumours. J. Pathol. 2011, 224, 334–343. [Google Scholar] [CrossRef]
- Pansuriya, T.C.; van Eijk, R.; d’Adamo, P.; van Ruler, M.A.J.H.; Kuijjer, M.L.; Oosting, J.; Cleton-Jansen, A.-M.; van Oosterwijk, J.G.; Verbeke, S.L.J.; Meijer, D.; et al. Somatic mosaic IDH1 and IDH2 mutations are associated with enchondroma and spindle cell hemangioma in Ollier disease and Maffucci syndrome. Nat. Genet. 2011, 43, 1256–1261. [Google Scholar] [CrossRef] [PubMed]
- Suijker, J.; Oosting, J.; Koornneef, A.; Struys, E.A.; Salomons, G.S.; Schaap, F.G.; Waaijer, C.J.; Wijers-Koster, P.M.; Briaire-de Bruijn, I.H.; Haazen, L. Inhibition of mutant IDH1 decreases D-2-HG levels without affecting tumorigenic properties of chondrosarcoma cell lines. Oncotarget 2015, 6, 12505–12519. [Google Scholar] [CrossRef] [Green Version]
- Dang, L.; White, D.W.; Gross, S.; Bennett, B.D.; Bittinger, M.A.; Driggers, E.M.; Fantin, V.R.; Jang, H.G.; Jin, S.; Keenan, M.C.; et al. Cancer-associated IDH1 mutations produce 2-hydroxyglutarate. Nature 2009, 462, 739–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Losman, J.-A.; Kaelin, W.G.J.G. What a difference a hydroxyl makes: Mutant IDH,(R)-2-hydroxyglutarate, and cancer. Genes Dev. 2013, 27, 836–852. [Google Scholar] [CrossRef] [Green Version]
- Morrison, C.; Radmacher, M.; Mohammed, N.; Suster, D.; Auer, H.; Jones, S.; Riggenbach, J.; Kelbick, N.; Bos, G.; Mayerson, J. MYC amplification and polysomy 8 in chondrosarcoma: Array comparative genomic hybridization, fluorescent in situ hybridization, and association with outcome. J. Clin. Oncol. 2005, 23, 9369–9376. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Riceberg, J.; Soucy, T.; Koenig, E.; Minissale, J.; Gallery, M.; Bernard, H.; Yang, X.; Liao, H.; Rabino, C.; et al. Probing the roles of SUMOylation in cancer cell biology by using a selective SAE inhibitor. Nat. Chem. Biol. 2017, 13, 1164–1171. [Google Scholar] [CrossRef]
- van Oosterwijk, J.G.; van Ruler, M.A.; Briaire-de Bruijn, I.H.; Herpers, B.; Gelderblom, H.; van de Water, B.; Bovée, J.V. Src kinases in chondrosarcoma chemoresistance and migration: Dasatinib sensitises to doxorubicin in TP53 mutant cells. Br. J. Cancer 2013, 109, 1214–1222. [Google Scholar] [CrossRef] [Green Version]
- Vertegaal, A.C.; Ogg, S.C.; Jaffray, E.; Rodriguez, M.S.; Hay, R.T.; Andersen, J.S.; Mann, M.; Lamond, A.I. A proteomic study of SUMO-2 target proteins. J. Biol. Chem. 2004, 279, 33791–33798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranski, Z.; Booij, T.H.; Cleton-Jansen, A.-M.; Price, L.S.; van de Water, B.; Bovée, J.V.; Hogendoorn, P.C.; Danen, E.H. Aven-mediated checkpoint kinase control regulates proliferation and resistance to chemotherapy in conventional osteosarcoma. J. Pathol. 2015, 236, 348–359. [Google Scholar] [CrossRef]
- Nicolle, R.; Ayadi, M.; Gomez-Brouchet, A.; Armenoult, L.; Banneau, G.; Elarouci, N.; Tallegas, M.; Decouvelaere, A.-V.; Aubert, S.; Rédini, F.; et al. Integrated molecular characterization of chondrosarcoma reveals critical determinants of disease progression. Nat. Commun. 2019, 10, 4622. [Google Scholar] [CrossRef] [PubMed]
- Nagy, Á.; Munkácsy, G.; Győrffy, B. Pancancer survival analysis of cancer hallmark genes. Sci. Rep. 2021, 11, 6047. [Google Scholar] [CrossRef] [PubMed]
- Gil-Benso, R.; Lopez-Gines, C.; López-Guerrero, J.A.; Carda, C.; Callaghan, R.C.; Navarro, S.; Ferrer, J.; Pellín, A.; Llombart-Bosch, A. Establishment and characterization of a continuous human chondrosarcoma cell line, ch-2879: Comparative histologic and genetic studies with its tumor of origin. Lab. Investig. 2003, 83, 877–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Oosterwijk, J.G.; de Jong, D.; van Ruler, M.A.; Hogendoorn, P.C.; Dijkstra, P.S.; van Rijswijk, C.S.; Machado, I.; Llombart-Bosch, A.; Szuhai, K.; Bovée, J.V. Three new chondrosarcoma cell lines: One grade III conventional central chondrosarcoma and two dedifferentiated chondrosarcomas of bone. BMC Cancer 2012, 12, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scully, S.P.; Berend, K.R.; Toth, A.; Qi, W.N.; Qi, Z.; Block, J.A. Marshall Urist Award. Interstitial collagenase gene expression correlates with in vitro invasion in human chondrosarcoma. Clin. Orthop. Relat. Res. 2000, 291–303. [Google Scholar] [CrossRef]
- Rasheed, S.; Nelson-Rees, W.A.; Toth, E.M.; Arnstein, P.; Gardner, M.B. Characterization of a newly derived human sarcoma cell line (HT-1080). Cancer 1974, 33, 1027–1033. [Google Scholar] [CrossRef]
- Kudo, N.; Ogose, A.; Hotta, T.; Kawashima, H.; Gu, W.; Umezu, H.; Toyama, T.; Endo, N.J.V.A. Establishment of novel human dedifferentiated chondrosarcoma cell line with osteoblastic differentiation. Virchows Arch. 2007, 451, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Calabuig-Fariñas, S.; Benso, R.G.; Szuhai, K.; Machado, I.; López-Guerrero, J.A.; de Jong, D.; Peydró, A.; Miguel, T.S.; Navarro, L.; Pellín, A.; et al. Characterization of a New Human Cell Line (CH-3573) Derived from a Grade II Chondrosarcoma with Matrix Production. Pathol. Oncol. Res. 2012, 18, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Hafner, M.; Niepel, M.; Chung, M.; Sorger, P.K. Growth rate inhibition metrics correct for confounders in measuring sensitivity to cancer drugs. Nat. Methods 2016, 13, 521–527. [Google Scholar] [CrossRef] [Green Version]
- Venneker, S.; Kruisselbrink, A.B.; Briaire-de Bruijn, I.H.; de Jong, Y.; van Wijnen, A.J.; Danen, E.H.J.; Bovée, J. Inhibition of PARP Sensitizes Chondrosarcoma Cell Lines to Chemo- and Radiotherapy Irrespective of the IDH1 or IDH2 Mutation Status. Cancers 2019, 11, 1918. [Google Scholar] [CrossRef] [Green Version]
- Schneekloth, J.S., Jr. Drug discovery: Controlling protein SUMOylation. Nat. Chem. Biol. 2017, 13, 1141–1142. [Google Scholar] [CrossRef]
- Hoellein, A.; Fallahi, M.; Schoeffmann, S.; Steidle, S.; Schaub, F.X.; Rudelius, M.; Laitinen, I.; Nilsson, L.; Goga, A.; Peschel, C.; et al. Myc-induced SUMOylation is a therapeutic vulnerability for B-cell lymphoma. Blood 2014, 124, 2081–2090. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Riceberg, J.; Pulukuri, S.M.; Grossman, S.; Shinde, V.; Shah, P.; Brownell, J.E.; Dick, L.; Newcomb, J.; Bence, N. Characterization of the loss of SUMO pathway function on cancer cells and tumor proliferation. PLoS ONE 2015, 10, e0123882. [Google Scholar] [CrossRef] [Green Version]
- Seeler, J.S.; Bischof, O.; Nacerddine, K.; Dejean, A. SUMO, the three Rs and cancer. In Acute Promyelocytic Leukemia: Molecular Genetics, Mouse Models and Targeted Therapy; Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2007; Volume 313, pp. 49–71. [Google Scholar]
- Stead, K.; Aguilar, C.; Hartman, T.; Drexel, M.; Meluh, P.; Guacci, V.J.T.J.o.c.b. Pds5p regulates the maintenance of sister chromatid cohesion and is sumoylated to promote the dissolution of cohesion. J. Cell Biol. 2003, 163, 729–741. [Google Scholar] [CrossRef] [Green Version]
- Bachant, J.; Alcasabas, A.; Blat, Y.; Kleckner, N.; Elledge, S.J.J.M.C. The SUMO-1 isopeptidase Smt4 is linked to centromeric cohesion through SUMO-1 modification of DNA topoisomerase II. Mol. Cell 2002, 9, 1169–1182. [Google Scholar] [CrossRef]
- Dawlaty, M.M.; Malureanu, L.; Jeganathan, K.B.; Kao, E.; Sustmann, C.; Tahk, S.; Shuai, K.; Grosschedl, R.; van Deursen, J.M.J.C. Resolution of sister centromeres requires RanBP2-mediated SUMOylation of topoisomerase IIα. Cell 2008, 133, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Rai, R.; Varma, S.P.; Shinde, N.; Ghosh, S.; Kumaran, S.P.; Skariah, G.; Laloraya, S.J.J.o.B.C. Small ubiquitin-related modifier ligase activity of Mms21 is required for maintenance of chromosome integrity during the unperturbed mitotic cell division cycle in Saccharomyces cerevisiae. J. Biol. Chem. 2011, 286, 14516–14530. [Google Scholar] [CrossRef] [Green Version]
- Behlke-Steinert, S.; Touat-Todeschini, L.; Skoufias, D.A.; Margolis, R.L. SMC5 and MMS21 are required for chromosome cohesion and mitotic progression. Cell Cycle 2009, 8, 2211–2218. [Google Scholar] [CrossRef]
- Strotman, P.K.; Reif, T.J.; Kliethermes, S.A.; Sandhu, J.K.; Nystrom, L.M. Dedifferentiated chondrosarcoma: A survival analysis of 159 cases from the SEER database (2001–2011). J. Surg. Oncol. 2017, 116, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Bakker, S.H.; Jacobs, W.C.H.; Pondaag, W.; Gelderblom, H.; Nout, R.A.; Dijkstra, P.D.S.; Peul, W.C.; Vleggeert-Lankamp, C.L.A. Chordoma: A systematic review of the epidemiology and clinical prognostic factors predicting progression-free and overall survival. Eur. Spine J. 2018, 27, 3043–3058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimer, R.J.; Gosheger, G.; Taminiau, A.; Biau, D.; Matejovsky, Z.; Kollender, Y.; San-Julian, M.; Gherlinzoni, F.; Ferrari, C. Dedifferentiated chondrosarcoma: Prognostic factors and outcome from a European group. Eur. J. Cancer 2007, 43, 2060–2065. [Google Scholar] [CrossRef] [PubMed]
- Meijer, D.; de Jong, D.; Pansuriya, T.C.; van den Akker, B.E.; Picci, P.; Szuhai, K.; Bovée, J.V. Genetic characterization of mesenchymal, clear cell, and dedifferentiated chondrosarcoma. Genes Chromosomes Cancer 2012, 51, 899–909. [Google Scholar] [CrossRef] [PubMed]
- Peterse, E.F.; Niessen, B.; Addie, R.D.; de Jong, Y.; Cleven, A.H.; Kruisselbrink, A.B.; van den Akker, B.E.; Molenaar, R.J.; Cleton-Jansen, A.-M.; Bovée, J.V. Targeting glutaminolysis in chondrosarcoma in context of the IDH1/2 mutation. Br. J. Cancer 2018, 118, 1074–1083. [Google Scholar] [CrossRef] [Green Version]
- Addie, R.D.; de Jong, Y.; Alberti, G.; Kruisselbrink, A.B.; Que, I.; Baelde, H.; Bovee, J. Exploration of the chondrosarcoma metabolome; the mTOR pathway as an important pro-survival pathway. J. Bone Oncol. 2019, 15, 100222. [Google Scholar] [CrossRef]
- Song, J.; Wang, X.; Zhu, J.; Liu, J. Rapamycin causes growth arrest and inhibition of invasion in human chondrosarcoma cells. J. BUON 2016, 21, 244–251. [Google Scholar]
- van Oosterwijk, J.G.; Meijer, D.; van Ruler, M.A.J.H.; van den Akker, B.E.W.M.; Oosting, J.; Krenács, T.; Picci, P.; Flanagan, A.M.; Liegl-Atzwanger, B.; Leithner, A.; et al. Screening for Potential Targets for Therapy in Mesenchymal, Clear Cell, and Dedifferentiated Chondrosarcoma Reveals Bcl-2 Family Members and TGFβ as Potential Targets. Am. J. Pathol. 2013, 182, 1347–1356. [Google Scholar] [CrossRef] [PubMed]
- Scotlandi, K.; Hattinger, C.M.; Pellegrini, E.; Gambarotti, M.; Serra, M. Genomics and Therapeutic Vulnerabilities of Primary Bone Tumors. Cells 2020, 9, 968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, L.; Li, Y.J.; Fakih, M.; Wiatrek, R.L.; Duldulao, M.; Chen, Z.B.; Chu, P.G.; Garcia-Aguilar, J.; Chen, Y. Role of SUMO activating enzyme in cancer stem cell maintenance and self-renewal. Nat. Commun. 2016, 7, 12326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroonen, J.S.; Vertegaal, A.C.O. Targeting SUMO Signaling to Wrestle Cancer. Trends Cancer 2020, 7, 496–510. [Google Scholar] [CrossRef]
- Biederstädt, A.; Hassan, Z.; Schneeweis, C.; Schick, M.; Schneider, L.; Muckenhuber, A.; Hong, Y.; Siegers, G.; Nilsson, L.; Wirth, M.; et al. SUMO pathway inhibition targets an aggressive pancreatic cancer subtype. Gut 2020, 69, 1472–1482. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.J.; Du, L.; Wang, J.; Vega, R.; Lee, T.D.; Miao, Y.; Aldana-Masangkay, G.; Samuels, E.R.; Li, B.; Ouyang, S.X.; et al. Allosteric Inhibition of Ubiquitin-like Modifications by a Class of Inhibitor of SUMO-Activating Enzyme. Cell Chem. Biol. 2019, 26, 278–288. [Google Scholar] [CrossRef]
- Langston, S.P.; Grossman, S.; England, D.; Afroze, R.; Bence, N.; Bowman, D.; Bump, N.; Chau, R.; Chuang, B.C.; Claiborne, C.; et al. Discovery of TAK-981, a First-in-Class Inhibitor of SUMO-Activating Enzyme for the Treatment of Cancer. J. Med. Chem. 2021, 64, 2501–2520. [Google Scholar] [CrossRef]
- Shah, J.J.; Jakubowiak, A.J.; O’Connor, O.A.; Orlowski, R.Z.; Harvey, R.D.; Smith, M.R.; Lebovic, D.; Diefenbach, C.; Kelly, K.; Hua, Z.; et al. Phase I Study of the Novel Investigational NEDD8-Activating Enzyme Inhibitor Pevonedistat (MLN4924) in Patients with Relapsed/Refractory Multiple Myeloma or Lymphoma. Clin. Cancer Res. 2016, 22, 34–43. [Google Scholar] [CrossRef] [Green Version]
- Malhab, L.J.B.; Descamps, S.; Delaval, B.; Xirodimas, D.P. The use of the NEDD8 inhibitor MLN4924 (Pevonedistat) in a cyclotherapy approach to protect wild-type p53 cells from MLN4924 induced toxicity. Sci. Rep. 2016, 6, 37775. [Google Scholar] [CrossRef] [Green Version]
- Hyer, M.L.; Milhollen, M.A.; Ciavarri, J.; Fleming, P.; Traore, T.; Sappal, D.; Huck, J.; Shi, J.; Gavin, J.; Brownell, J.; et al. A small-molecule inhibitor of the ubiquitin activating enzyme for cancer treatment. Nat. Med. 2018, 24, 186–193. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Association | Variable | Hazard Ratio (B) | CI (95%) (exp(B)) | p Value |
|---|---|---|---|---|
| 10-year survival | ||||
| SUMO1 expression * | 0.312 | 0.614–0.3042 | 0.444 | |
| ACT/CS1 (compared to grade 3) # | −2.488 | 0.026–0.267 | 0.000 | |
| Grade 2 (compared to grade 3) # | −1.198 | 0.122–0.744 | 0.009 | |
| SUMO2/3exp (grade dependent) # | −0.538 | 0.245–0.1392 | 0.225 | |
| SUMO2/3 expression * | 0.95 | 1.117–5.979 | 0.026 | |
| ACT/CS1 (compared to grade 3) # | −2.692 | 0.014–0.338 | 0.001 | |
| Grade 2 (compared to grade 3) # | −1.155 | 0.109–0.914 | 0.034 | |
| SUMO1exp (grade dependent) # | −0.262 | 0.254–2.331 | 0.644 | |
| 10-year survival (death of disease) | ||||
| SUMO1 expression * | 0.527 | 0.517–5.552 | 0.384 | |
| ACT/CS1 (compared to grade 3) # | −3.773 | 0.003–0.205 | 0.001 | |
| Grade 2 (compared to grade 3) # | −2.047 | 0.032–0.517 | 0.004 | |
| SUMO2/3exp (grade dependent) # | −0.685 | 0.144–1.766 | 0.284 | |
| SUMO2/3 expression * | 0.892 | 0.687–8.654 | 0.168 | |
| ACT/CS1 (compared to grade 3) # | −15.087 | 0.00–8.075E136 | 0.929 | |
| Grade 2 (compared to grade 3) # | −3.075 | 0.007–0.319 | 0.002 | |
| SUMO1exp (grade dependent) # | −1.933 | 0.23–0.896 | 0.038 | |
| Metastasis-free survival (10-year) | ||||
| SUMO1 expression * | 0.523 | 0.515–5.528 | 0.388 | |
| ACT/CS1 (compared to grade 3) # | 10.145 | 0.00–1.112E+120 | 0.940 | |
| Grade 2 (compared to grade 3) # | 10.991 | 0.00–2.586E+120 | 0.936 | |
| SUMO2/3exp (grade dependent) # | 0.407 | 0.398–5.676 | 0.548 | |
| SUMO2/3 expression * | 0.606 | 0.590–5.694 | 0.295 | |
| ACT/CS1 (compared to grade 3) # | −3.941 | 0.002–0.186 | 0.001 | |
| Grade 2 (compared to grade 3) # | −2.064 | 0.033–0.492 | 0.003 | |
| SUMO1exp (grade dependent) # | −0.894 | 0.112–1.491 | 0.175 | |
| Recurrence-free survival (10 year) | ||||
| SUMO1 expression * | 0.171 | 0.552–2.553 | 0.661 | |
| ACT/CS1 (compared to grade 3) # | −3.063 | 0.16–0.140 | 0.000 | |
| Grade 2 (compared to grade 3) # | −1.623 | 0.083–0.468 | 0.000 | |
| SUMO2/3exp (grade dependent) # | −0.897 | 0.177–0.950 | 0.038 | |
| SUMO2/3 expression * | −0.026 | 0.434–2.037 | 0.875 | |
| ACT/CS1 (compared to grade 3) # | −3.076 | 0.015–0.146 | 0.000 | |
| Grade 2 (compared to grade 3) # | −1.53 | 0.092–0.511 | 0.000 | |
| SUMO1exp (grade dependent) # | −1.015 | 0.157–0.939 | 0.018 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kroonen, J.S.; Kruisselbrink, A.B.; Briaire-de Bruijn, I.H.; Olaofe, O.O.; Bovée, J.V.M.G.; Vertegaal, A.C.O. SUMOylation Is Associated with Aggressive Behavior in Chondrosarcoma of Bone. Cancers 2021, 13, 3823. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13153823
Kroonen JS, Kruisselbrink AB, Briaire-de Bruijn IH, Olaofe OO, Bovée JVMG, Vertegaal ACO. SUMOylation Is Associated with Aggressive Behavior in Chondrosarcoma of Bone. Cancers. 2021; 13(15):3823. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13153823
Chicago/Turabian StyleKroonen, Jessie S., Alwine B. Kruisselbrink, Inge H. Briaire-de Bruijn, Olaejirinde O. Olaofe, Judith V. M. G. Bovée, and Alfred C. O. Vertegaal. 2021. "SUMOylation Is Associated with Aggressive Behavior in Chondrosarcoma of Bone" Cancers 13, no. 15: 3823. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13153823