
Gene Expression in Barrett’s Esophagus Cell Lines Resemble Esophageal Squamous Cell Carcinoma Instead of Esophageal Adenocarcinoma


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. TCGA Dataset
2.2. RNAseq Data Sets from Barrett’s Epithelial Carcinogenesis (BEC) Model
2.3. Other Gene Expression Datasets
2.4. Data Processing
2.5. Data Analysis
3. Results
3.1. Construction and Validation of Gene Expression Signatures of EAC and ESCC
3.2. ESCC Cell Lines Are Similar to Primary ESCC Tumors, But EAC Cell Lines (Although EAC-Like) Are Considerably Different from Primary EAC Tumors
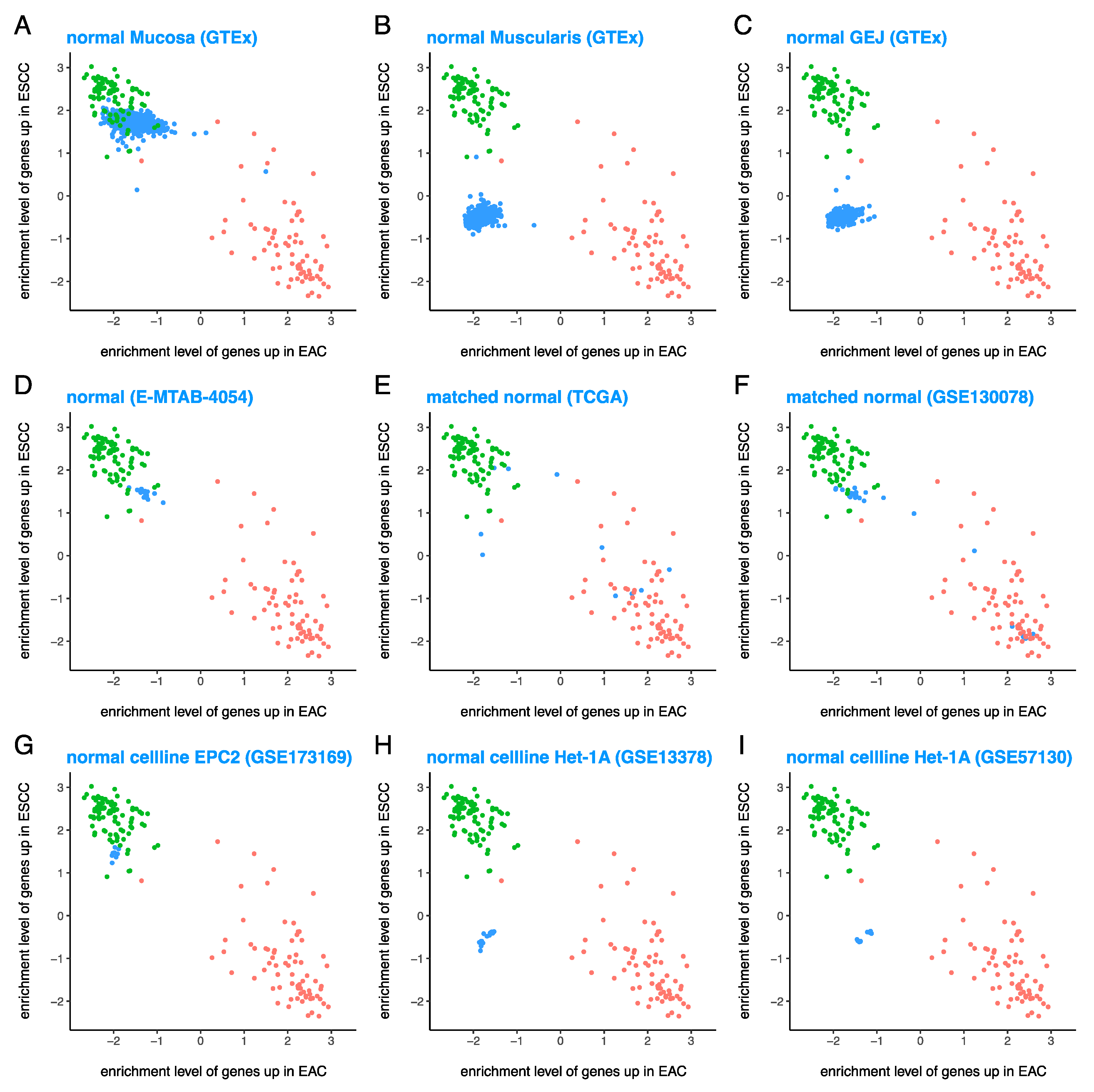
3.3. Normal Esophageal Cell Lines Are Quite Similar to Various Tissues from Normal Esophagus
3.4. BE Cell Lines Are ESCC-Like and Extremely Different from the EAC-Like Primary BE Tissues
3.5. ABS Exposure Was Not Sufficient to Induce EAC-Like GEP in the BE Cell Line BAR-T and It Remained ESCC-Like Even after Malignant Transformation
3.6. Expression Pattern of Individual EAChi and ESCChi Genes in Various Tumor and Normal Tissues and Cell Lines
3.7. The Only EAC Case Comparable to BE Cell Lines Has Features of both EAC and ESCC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siewert, J.R.; Ott, K. Are squamous and adenocarcinomas of the esophagus the same disease? Semin. Radiat. Oncol. 2007, 17, 38–44. [Google Scholar] [CrossRef]
- Hvid-Jensen, F.; Pedersen, L.; Drewes, A.M.; Sorensen, H.T.; Funch-Jensen, P. Incidence of adenocarcinoma among patients with Barrett’s esophagus. N Engl. J. Med. 2011, 365, 1375–1383. [Google Scholar] [CrossRef] [Green Version]
- Bhat, S.; Coleman, H.G.; Yousef, F.; Johnston, B.T.; McManus, D.T.; Gavin, A.T.; Murray, L.J. Risk of malignant progression in Barrett’s esophagus patients: Results from a large population-based study. J. Natl. Cancer Inst. 2011, 103, 1049–1057. [Google Scholar] [CrossRef] [Green Version]
- Desai, T.K.; Krishnan, K.; Samala, N.; Singh, J.; Cluley, J.; Perla, S.; Howden, C.W. The incidence of oesophageal adenocarcinoma in non-dysplastic Barrett’s oesophagus: A meta-analysis. Gut 2012, 61, 970–976. [Google Scholar] [CrossRef] [PubMed]
- Hawe, A.; Payne, W.S.; Weiland, L.H.; Fontana, R.S. Adenocarcinima in the columnar epithelial lined lower (Barret) oesophagus. Thorax 1973, 28, 511–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naef, A.P.; Savary, M.; Ozzello, L. Columnar-lined lower esophagus: An acquired lesion with malignant predisposition. Report on 140 cases of Barrett’s esophagus with 12 adenocarcinomas. J. Thorac. Cardiovasc. Surg. 1975, 70, 826–835. [Google Scholar] [CrossRef]
- Iascone, C.; DeMeester, T.R.; Little, A.G.; Skinner, D.B. Barrett’s esophagus. Functional assessment, proposed pathogenesis, and surgical therapy. Arch. Surg. 1983, 118, 543–549. [Google Scholar] [CrossRef]
- Vaezi, M.F.; Richter, J.E. Role of acid and duodenogastroesophageal reflux in gastroesophageal reflux disease. Gastroenterology 1996, 111, 1192–1199. [Google Scholar] [CrossRef] [PubMed]
- Allison, P.R. Peptic ulcer of the oesophagus. Thorax 1948, 3, 20–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minacapelli, C.D.; Bajpai, M.; Geng, X.; Cheng, C.L.; Chouthai, A.A.; Souza, R.; Spechler, S.J.; Das, K.M. Barrett’s metaplasia develops from cellular reprograming of esophageal squamous epithelium due to gastroesophageal reflux. Am. J. Physiol. Gastrointest Liver Physiol. 2017, 312, G615–G622. [Google Scholar] [CrossRef] [Green Version]
- Lagergren, J.; Bergstrom, R.; Lindgren, A.; Nyren, O. Symptomatic gastroesophageal reflux as a risk factor for esophageal adenocarcinoma. N Engl. J. Med. 1999, 340, 825–831. [Google Scholar] [CrossRef]
- Vaughan, T.L.; Davis, S.; Kristal, A.; Thomas, D.B. Obesity, alcohol, and tobacco as risk factors for cancers of the esophagus and gastric cardia: Adenocarcinoma versus squamous cell carcinoma. Cancer Epidemiol. Biomark. Prev. 1995, 4, 85–92. [Google Scholar]
- Gammon, M.D.; Schoenberg, J.B.; Ahsan, H.; Risch, H.A.; Vaughan, T.L.; Chow, W.H.; Rotterdam, H.; West, A.B.; Dubrow, R.; Stanford, J.L.; et al. Tobacco, alcohol, and socioeconomic status and adenocarcinomas of the esophagus and gastric cardia. J. Natl. Cancer Inst. 1997, 89, 1277–1284. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Johnson, A.; Ali, S.M.; Klempner, S.J.; Bekaii-Saab, T.; Vacirca, J.L.; Khaira, D.; Yelensky, R.; Chmielecki, J.; Elvin, J.A.; et al. Comprehensive Genomic Profiling of Advanced Esophageal Squamous Cell Carcinomas and Esophageal Adenocarcinomas Reveals Similarities and Differences. Oncologist 2015, 20, 1132–1139. [Google Scholar] [CrossRef] [Green Version]
- Alexandrov, L.B.; Nik-Zainal, S.; Wedge, D.C.; Aparicio, S.A.; Behjati, S.; Biankin, A.V.; Bignell, G.R.; Bolli, N.; Borg, A.; Borresen-Dale, A.L.; et al. Signatures of mutational processes in human cancer. Nature 2013, 500, 415–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cancer Genome Atlas Research Network; Analysis Working Group; Kim, J.; Bowlby, R.; Mungall, A.J.; Robertson, A.G.; Odze, R.D.; Cherniack, A.D.; Shih, J.; Pedamallu, C.S.; et al. Integrated genomic characterization of oesophageal carcinoma. Nature 2017, 541, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Hur, C.; Miller, M.; Kong, C.Y.; Dowling, E.C.; Nattinger, K.J.; Dunn, M.; Feuer, E.J. Trends in esophageal adenocarcinoma incidence and mortality. Cancer 2013, 119, 1149–1158. [Google Scholar] [CrossRef] [Green Version]
- Bajpai, M.; Liu, J.; Geng, X.; Souza, R.F.; Amenta, P.S.; Das, K.M. Repeated exposure to acid and bile selectively induces colonic phenotype expression in a heterogeneous Barrett’s epithelial cell line. Lab. Investig. 2008, 88, 643–651. [Google Scholar] [CrossRef] [Green Version]
- Das, K.M.; Kong, Y.; Bajpai, M.; Kulkarni, D.; Geng, X.; Mishra, P.; Banerjee, D.; Hirshfield, K. Transformation of benign Barrett’s epithelium by repeated acid and bile exposure over 65 weeks: A novel in vitro model. Int. J. Cancer 2011, 128, 274–282. [Google Scholar] [CrossRef] [Green Version]
- Bajpai, M.; Aviv, H.; Das, K.M. Prolonged exposure to acid and bile induces chromosome abnormalities that precede malignant transformation of benign Barrett’s epithelium. Mol. Cytogenet. 2012, 5, 43. [Google Scholar] [CrossRef] [Green Version]
- Bajpai, M.; Kessel, R.; Bhagat, T.; Nischal, S.; Yu, Y.; Verma, A.; Das, K.M. High resolution integrative analysis reveals widespread genetic and epigenetic changes after chronic in-vitro acid and bile exposure in Barrett’s epithelium cells. Genes Chromosomes Cancer 2013, 52, 1123–1132. [Google Scholar] [CrossRef] [PubMed]
- Bajpai, M.; Panda, A.; Birudaraju, K.; Van Gurp, J.; Chak, A.; Das, K.M.; Javidian, P.; Aviv, H. Recurring Translocations in Barrett’s Esophageal Adenocarcinoma. Front. Genet. 2021, 12, 674741. [Google Scholar] [CrossRef]
- Panda, A.; Shin, M.R.; Cheng, C.; Bajpai, M. Barrett’s Epithelium to Esophageal Adenocarcinoma: Is There a “Point of No Return”? Front. Genet. 2021, 12, 706706. [Google Scholar] [CrossRef]
- Jaiswal, K.R.; Morales, C.P.; Feagins, L.A.; Gandia, K.G.; Zhang, X.; Zhang, H.Y.; Hormi-Carver, K.; Shen, Y.; Elder, F.; Ramirez, R.D.; et al. Characterization of telomerase-immortalized, non-neoplastic, human Barrett’s cell line (BAR-T). Dis. Esophagus 2007, 20, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakravarty, D.; Gao, J.; Phillips, S.M.; Kundra, R.; Zhang, H.; Wang, J.; Rudolph, J.E.; Yaeger, R.; Soumerai, T.; Nissan, M.H.; et al. OncoKB: A Precision Oncology Knowledge Base. JCO Precis. Oncol. 2017, 2017. [Google Scholar] [CrossRef]
- Ghandi, M.; Huang, F.W.; Jane-Valbuena, J.; Kryukov, G.V.; Lo, C.C.; McDonald, E.R., 3rd; Barretina, J.; Gelfand, E.T.; Bielski, C.M.; Li, H.; et al. Next-generation characterization of the Cancer Cell Line Encyclopedia. Nature 2019, 569, 503–508. [Google Scholar] [CrossRef]
- G. TEx Consortium. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef]
- Duggan, S.P.; Gallagher, W.M.; Fox, E.J.; Abdel-Latif, M.M.; Reynolds, J.V.; Kelleher, D. Low pH induces co-ordinate regulation of gene expression in oesophageal cells. Carcinogenesis 2006, 27, 319–327. [Google Scholar] [CrossRef]
- Duggan, S.P.; Behan, F.M.; Kirca, M.; Smith, S.; Reynolds, J.V.; Long, A.; Kelleher, D. An integrative genomic approach in oesophageal cells identifies TRB3 as a bile acid responsive gene, downregulated in Barrett’s oesophagus, which regulates NF-kappaB activation and cytokine levels. Carcinogenesis 2010, 31, 936–945. [Google Scholar] [CrossRef] [Green Version]
- Silvers, A.L.; Lin, L.; Bass, A.J.; Chen, G.; Wang, Z.; Thomas, D.G.; Lin, J.; Giordano, T.J.; Orringer, M.B.; Beer, D.G.; et al. Decreased selenium-binding protein 1 in esophageal adenocarcinoma results from posttranscriptional and epigenetic regulation and affects chemosensitivity. Clin. Cancer Res. 2010, 16, 2009–2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leicht, D.T.; Kausar, T.; Wang, Z.; Ferrer-Torres, D.; Wang, T.D.; Thomas, D.G.; Lin, J.; Chang, A.C.; Lin, L.; Beer, D.G. TGM2: A cell surface marker in esophageal adenocarcinomas. J. Thorac. Oncol. 2014, 9, 872–881. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Myers, A.L.; Wang, Z.; Nancarrow, D.J.; Ferrer-Torres, D.; Handlogten, A.; Leverenz, K.; Bao, J.; Thomas, D.G.; Wang, T.D.; et al. Osteopontin (OPN/SPP1) isoforms collectively enhance tumor cell invasion and dissemination in esophageal adenocarcinoma. Oncotarget 2015, 6, 22239–22257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, A.L.; Lin, L.; Nancarrow, D.J.; Wang, Z.; Ferrer-Torres, D.; Thomas, D.G.; Orringer, M.B.; Lin, J.; Reddy, R.M.; Beer, D.G.; et al. IGFBP2 modulates the chemoresistant phenotype in esophageal adenocarcinoma. Oncotarget 2015, 6, 25897–25916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahrens, T.D.; Timme, S.; Hoeppner, J.; Ostendorp, J.; Hembach, S.; Follo, M.; Hopt, U.T.; Werner, M.; Busch, H.; Boerries, M.; et al. Selective inhibition of esophageal cancer cells by combination of HDAC inhibitors and Azacytidine. Epigenetics 2015, 10, 431–445. [Google Scholar] [CrossRef] [Green Version]
- Sulahian, R.; Chen, J.; Arany, Z.; Jadhav, U.; Peng, S.; Rustgi, A.K.; Bass, A.J.; Srivastava, A.; Hornick, J.L.; Shivdasani, R.A. SOX15 governs transcription in human stratified epithelia and a subset of esophageal adenocarcinomas. Cell Mol. Gastroenterol. Hepatol. 2015, 1, 598–609.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ykema, B.L.M.; Hoefnagel, S.J.M.; Rigter, L.S.; Kodach, L.L.; Meijer, G.A.; van Leeuwen, F.E.; Khan, H.N.; Snaebjornsson, P.; Aleman, B.M.P.; Broeks, A.; et al. Gene expression profiles of esophageal squamous cell cancers in Hodgkin lymphoma survivors versus sporadic cases. PLoS ONE 2020, 15, e0243178. [Google Scholar] [CrossRef]
- You, B.H.; Yoon, J.H.; Kang, H.; Lee, E.K.; Lee, S.K.; Nam, J.W. HERES, a lncRNA that regulates canonical and noncanonical Wnt signaling pathways via interaction with EZH2. Proc. Natl. Acad. Sci. USA 2019, 116, 24620–24629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Ende, T.; de Clercq, N.C.; van Berge Henegouwen, M.I.; Gisbertz, S.S.; Geijsen, E.D.; Verhoeven, R.H.A.; Meijer, S.L.; Schokker, S.; Dings, M.P.G.; Bergman, J.; et al. Neoadjuvant Chemoradiotherapy Combined with Atezolizumab for Resectable Esophageal Adenocarcinoma: A Single-arm Phase II Feasibility Trial (PERFECT). Clin. Cancer Res. 2021, 27, 3351–3359. [Google Scholar] [CrossRef] [PubMed]
- Janmaat, V.T.; Nesteruk, K.; Spaander, M.C.W.; Verhaar, A.P.; Yu, B.; Silva, R.A.; Phillips, W.A.; Magierowski, M.; van de Winkel, A.; Stadler, H.S.; et al. HOXA13 in etiology and oncogenic potential of Barrett’s esophagus. Nat. Commun. 2021, 12, 3354. [Google Scholar] [CrossRef]
- Klijn, C.; Durinck, S.; Stawiski, E.W.; Haverty, P.M.; Jiang, Z.; Liu, H.; Degenhardt, J.; Mayba, O.; Gnad, F.; Liu, J.; et al. A comprehensive transcriptional portrait of human cancer cell lines. Nat. Biotechnol. 2015, 33, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Maag, J.L.V.; Fisher, O.M.; Levert-Mignon, A.; Kaczorowski, D.C.; Thomas, M.L.; Hussey, D.J.; Watson, D.I.; Wettstein, A.; Bobryshev, Y.V.; Edwards, M.; et al. Novel Aberrations Uncovered in Barrett’s Esophagus and Esophageal Adenocarcinoma Using Whole Transcriptome Sequencing. Mol. Cancer Res. 2017, 15, 1558–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbie, D.A.; Tamayo, P.; Boehm, J.S.; Kim, S.Y.; Moody, S.E.; Dunn, I.F.; Schinzel, A.C.; Sandy, P.; Meylan, E.; Scholl, C.; et al. Systematic RNA interference reveals that oncogenic KRAS-driven cancers require TBK1. Nature 2009, 462, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, K.; Shahmoradgoli, M.; Martinez, E.; Vegesna, R.; Kim, H.; Torres-Garcia, W.; Trevino, V.; Shen, H.; Laird, P.W.; Levine, D.A.; et al. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat. Commun. 2013, 4, 2612. [Google Scholar] [CrossRef] [PubMed]
- Mirabelli, P.; Coppola, L.; Salvatore, M. Cancer Cell Lines Are Useful Model Systems for Medical Research. Cancers 2019, 11, 1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-David, U.; Siranosian, B.; Ha, G.; Tang, H.; Oren, Y.; Hinohara, K.; Strathdee, C.A.; Dempster, J.; Lyons, N.J.; Burns, R.; et al. Genetic and transcriptional evolution alters cancer cell line drug response. Nature 2018, 560, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Salvadores, M.; Fuster-Tormo, F.; Supek, F. Matching cell lines with cancer type and subtype of origin via mutational, epigenomic, and transcriptomic patterns. Sci. Adv. 2020, 6. [Google Scholar] [CrossRef]
- Wang, X.; Ouyang, H.; Yamamoto, Y.; Kumar, P.A.; Wei, T.S.; Dagher, R.; Vincent, M.; Lu, X.; Bellizzi, A.M.; Ho, K.Y.; et al. Residual embryonic cells as precursors of a Barrett’s-like metaplasia. Cell 2011, 145, 1023–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xian, W.; Ho, K.Y.; Crum, C.P.; McKeon, F. Cellular origin of Barrett’s esophagus: Controversy and therapeutic implications. Gastroenterology 2012, 142, 1424–1430. [Google Scholar] [CrossRef]
- Souza, R.F.; Krishnan, K.; Spechler, S.J. Acid, bile, and CDX: The ABCs of making Barrett’s metaplasia. Am. J. Physiol. Gastrointest Liver Physiol. 2008, 295, G211–G218. [Google Scholar] [CrossRef] [Green Version]
- Spechler, S.J.; Souza, R.F. Barrett’s esophagus. N. Engl. J. Med. 2014, 371, 836–845. [Google Scholar] [CrossRef] [PubMed]
- American Gastroenterological, A.; Spechler, S.J.; Sharma, P.; Souza, R.F.; Inadomi, J.M.; Shaheen, N.J. American Gastroenterological Association medical position statement on the management of Barrett’s esophagus. Gastroenterology 2011, 140, 1084–1091. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panda, A.; Bhanot, G.; Ganesan, S.; Bajpai, M. Gene Expression in Barrett’s Esophagus Cell Lines Resemble Esophageal Squamous Cell Carcinoma Instead of Esophageal Adenocarcinoma. Cancers 2021, 13, 5971. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13235971
Panda A, Bhanot G, Ganesan S, Bajpai M. Gene Expression in Barrett’s Esophagus Cell Lines Resemble Esophageal Squamous Cell Carcinoma Instead of Esophageal Adenocarcinoma. Cancers. 2021; 13(23):5971. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13235971
Chicago/Turabian StylePanda, Anshuman, Gyan Bhanot, Shridar Ganesan, and Manisha Bajpai. 2021. "Gene Expression in Barrett’s Esophagus Cell Lines Resemble Esophageal Squamous Cell Carcinoma Instead of Esophageal Adenocarcinoma" Cancers 13, no. 23: 5971. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13235971