
Sex Hormones in Lymphedema

 , , and
, , and 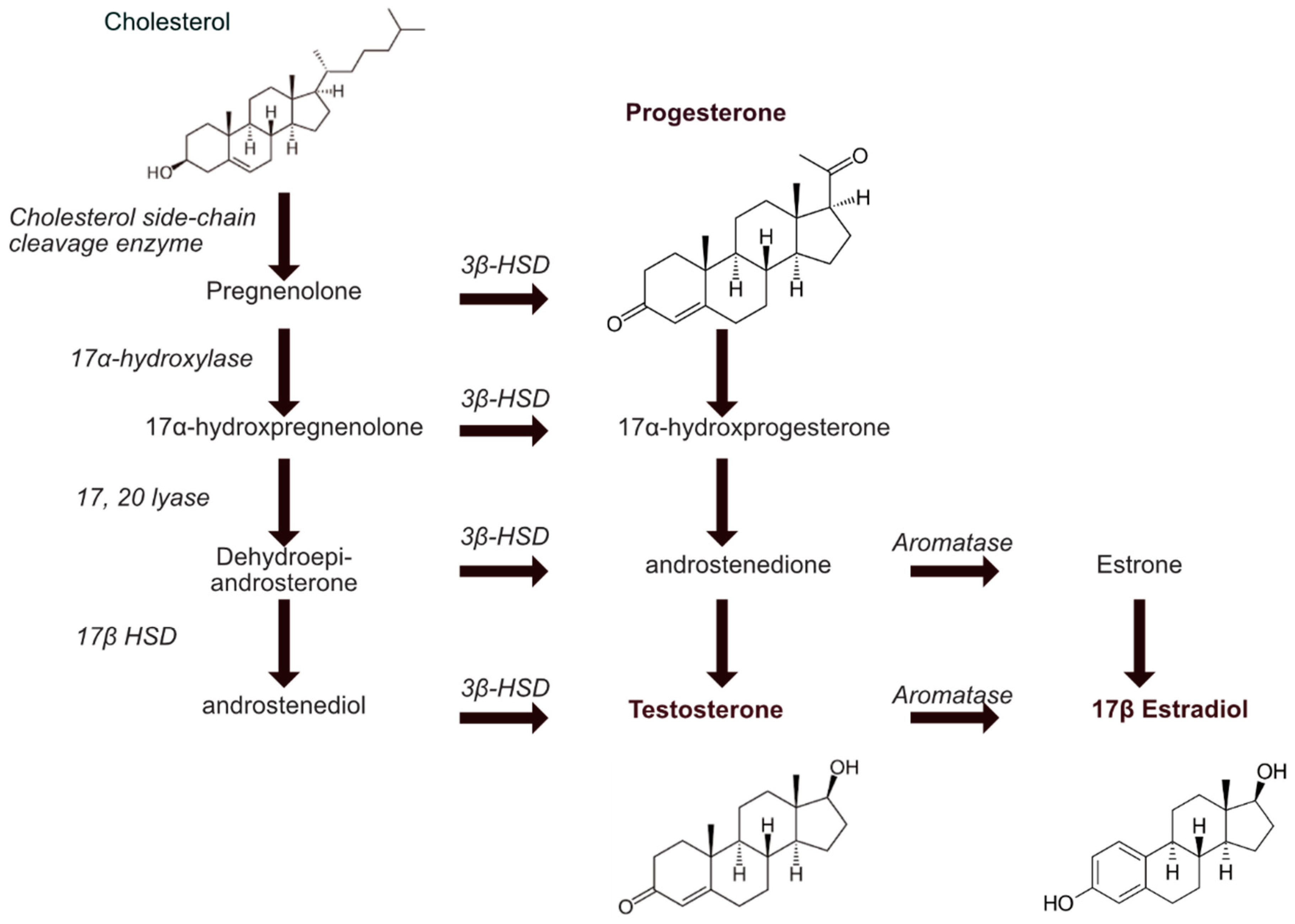{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Sex Hormones and Lymphatic Vasculature
2.1. Steroids Biosynthesis
2.2. Source of Estrogen
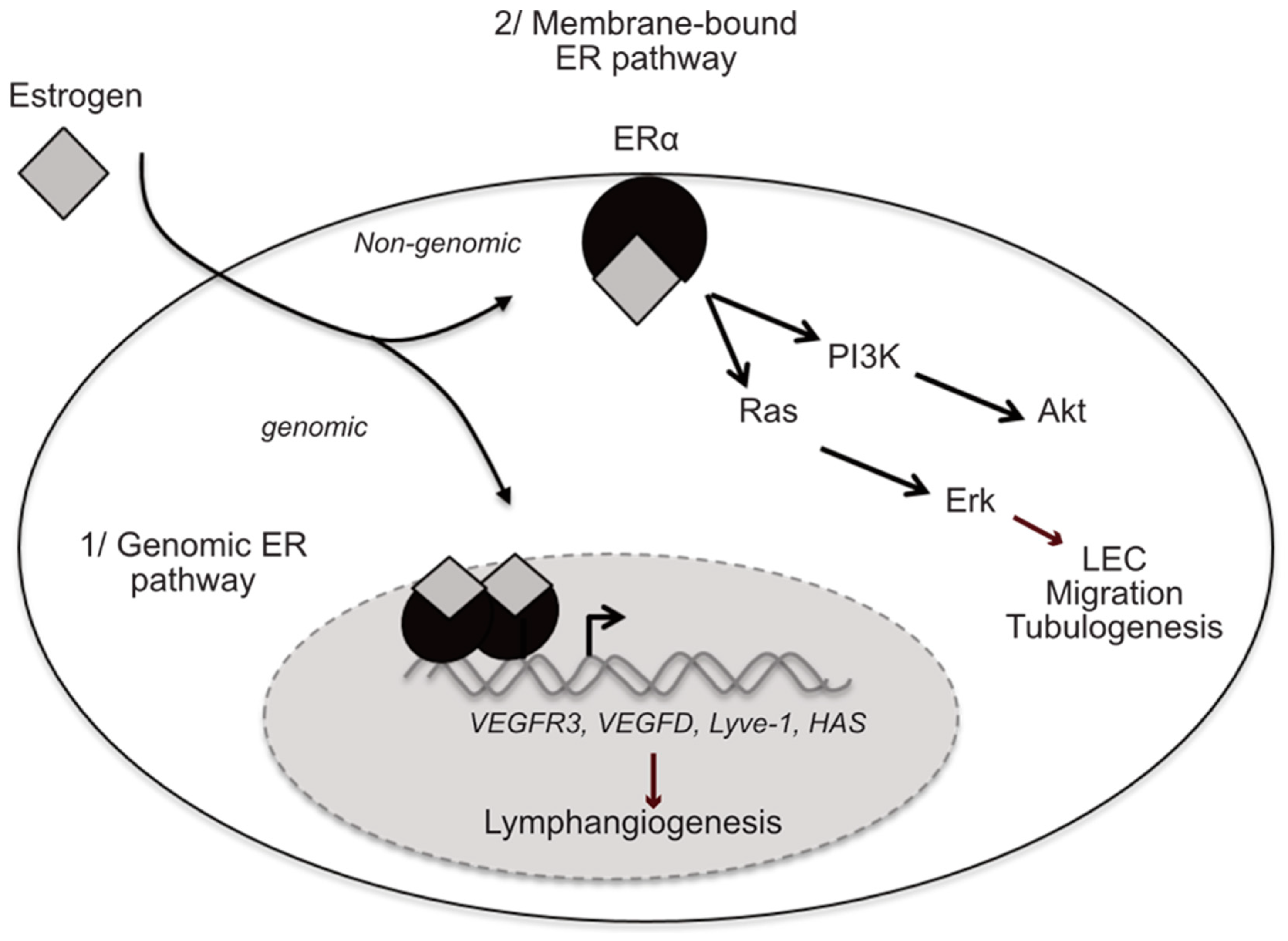
2.3. Estrogen Receptors
2.4. Protective Effect of Estrogen on Vascular and Lymphatic Vessels
3. Sex Hormones and Lymphedema
3.1. Primary Lymphedema
3.2. Secondary Lymphedema
3.2.1. Filariasis Lymphedema
3.2.2. Iatrogenic Lymphedema
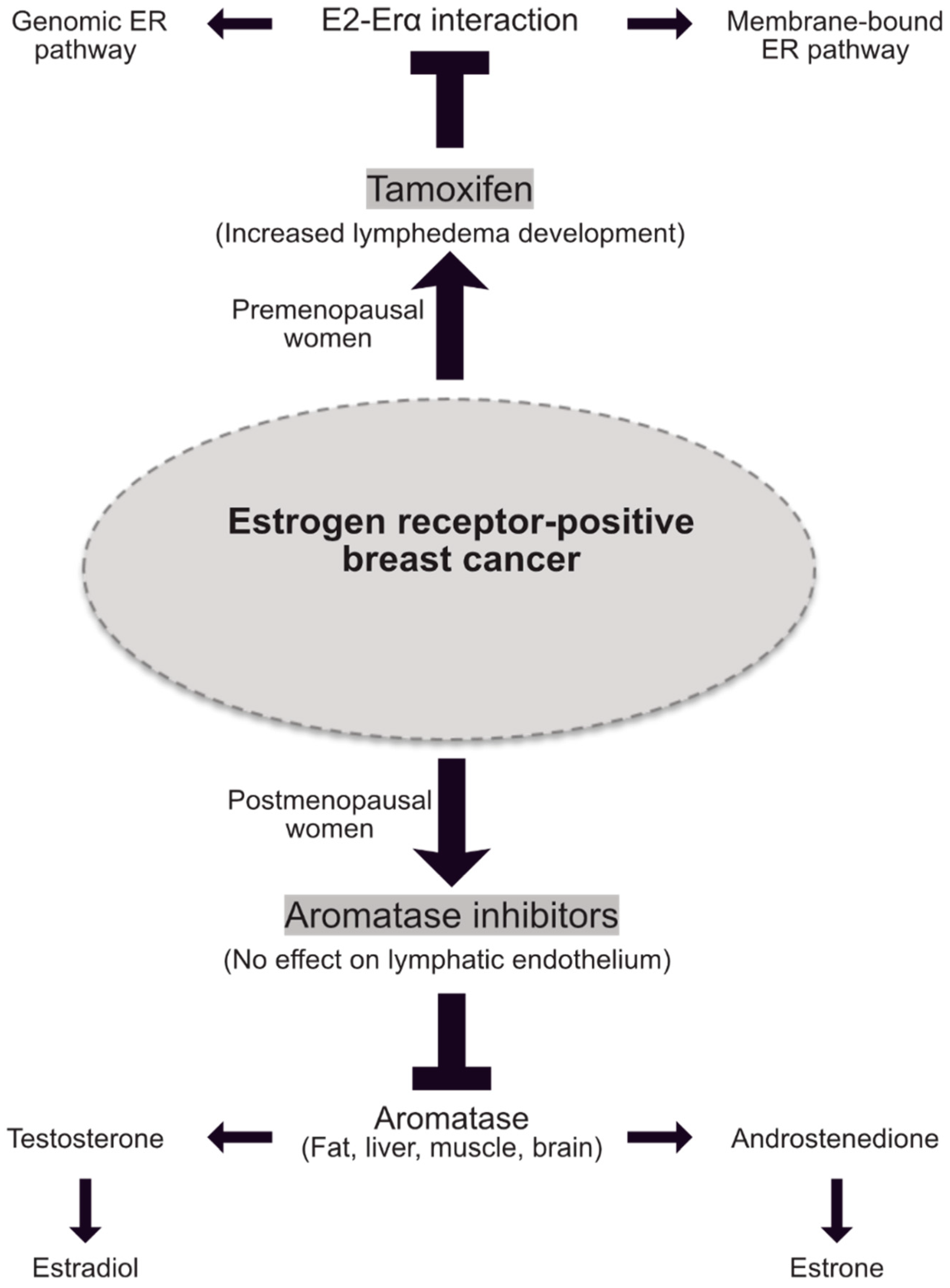
4. Hormone Therapy and Lymphedema
5. Discussion and Future Perspectives
6. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- dos Santos, R.L.; da Silva, F.B.; Ribeiro, R.F.; Stefanon, I. Sex Hormones in the Cardiovascular System. Horm. Mol. Biol. Clin. Investig. 2014, 18, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Arnal, J.-F.; Gourdy, P.; Elhage, R.; Garmy-Susini, B.; Delmas, E.; Brouchet, L.; Castano, C.; Barreira, Y.; Couloumiers, J.C.; Prats, H.; et al. Estrogens and Atherosclerosis. Eur. J. Endocrinol. 2004, 150, 113–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alitalo, K.; Tammela, T.; Petrova, T.V. Lymphangiogenesis in Development and Human Disease. Nature 2005, 438, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Morfoisse, F.; Tatin, F.; Chaput, B.; Therville, N.; Vaysse, C.; Métivier, R.; Malloizel-Delaunay, J.; Pujol, F.; Godet, A.-C.; De Toni, F.; et al. Lymphatic Vasculature Requires Estrogen Receptor-α Signaling to Protect From Lymphedema. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 1346–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unno, N.; Tanaka, H.; Suzuki, M.; Yamamoto, N.; Mano, Y.; Sano, M.; Saito, T.; Konno, H. Influence of Age and Gender on Human Lymphatic Pumping Pressure in the Leg. Lymphology 2011, 44, 113–120. [Google Scholar]
- Szuba, A.; Rockson, S.G. Lymphedema: Classification, Diagnosis and Therapy. Vasc. Med. 1998, 3, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Warren, A.G.; Brorson, H.; Borud, L.J.; Slavin, S.A. Lymphedema: A Comprehensive Review. Ann. Plast. Surg. 2007, 59, 464–472. [Google Scholar] [CrossRef]
- Allam, O.; Park, K.E.; Chandler, L.; Mozaffari, M.A.; Ahmad, M.; Lu, X.; Alperovich, M. The Impact of Radiation on Lymphedema: A Review of the Literature. Gland. Surg. 2020, 9, 596–602. [Google Scholar] [CrossRef]
- Avraham, T.; Daluvoy, S.; Zampell, J.; Yan, A.; Haviv, Y.S.; Rockson, S.G.; Mehrara, B.J. Blockade of Transforming Growth Factor-Β1 Accelerates Lymphatic Regeneration during Wound Repair. Am. J. Pathol. 2010, 177, 3202–3214. [Google Scholar] [CrossRef]
- Clavin, N.W.; Avraham, T.; Fernandez, J.; Daluvoy, S.V.; Soares, M.A.; Chaudhry, A.; Mehrara, B.J. TGF-Beta1 Is a Negative Regulator of Lymphatic Regeneration during Wound Repair. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H2113–H2127. [Google Scholar] [CrossRef] [Green Version]
- Kinashi, H.; Ito, Y.; Sun, T.; Katsuno, T.; Takei, Y. Roles of the TGF-Β–VEGF-C Pathway in Fibrosis-Related Lymphangiogenesis. Int. J. Mol. Sci. 2018, 19, 2487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogino, R.; Hayashida, K.; Yamakawa, S.; Morita, E. Adipose-Derived Stem Cells Promote Intussusceptive Lymphangiogenesis by Restricting Dermal Fibrosis in Irradiated Tissue of Mice. Int. J. Mol. Sci. 2020, 21, 3885. [Google Scholar] [CrossRef] [PubMed]
- Davies, C.; Pan, H.; Peto, R. 10 vs 5 Years of Adjuvant Tamoxifen: Exclusion of 1/402 Centres in ATLAS. Lancet 2017, 389, 1884. [Google Scholar] [CrossRef] [Green Version]
- Dessources, K.; Aviki, E.; Leitao, M.M., Jr. Lower Extremity Lymphedema in Patients with Gynecologic Malignancies. Int. J. Gynecol. Cancer 2020, 30. [Google Scholar] [CrossRef]
- Rasmusson, E.; Gunnlaugsson, A.; Blom, R.; Björk-Eriksson, T.; Nilsson, P.; Ahlgen, G.; Jönsson, C.; Johansson, K.; Kjellén, E. Low Rate of Lymphedema after Extended Pelvic Lymphadenectomy Followed by Pelvic Irradiation of Node-Positive Prostate Cancer. Radiat. Oncol. 2013, 8, 271. [Google Scholar] [CrossRef] [Green Version]
- Bosland, M.C. The Role of Estrogens in Prostate Carcinogenesis: A Rationale for Chemoprevention. Rev. Urol. 2005, 7, S4–S10. [Google Scholar]
- Parker, K.L.; Schimmer, B.P. Transcriptional Regulation of the Genes Encoding the Cytochrome P-450 Steroid Hydroxylases. Vitam. Horm. 1995, 51, 339–370. [Google Scholar] [CrossRef]
- Goddard, L.M.; Murphy, T.J.; Org, T.; Enciso, J.M.; Hashimoto-Partyka, M.K.; Warren, C.M.; Domigan, C.K.; McDonald, A.; He, H.; Sanchez, L.A.; et al. Progesterone Receptor in the Vascular Endothelium Triggers Physiological Uterine Permeability Pre-Implantation. Cell 2014, 156, 549–562. [Google Scholar] [CrossRef] [Green Version]
- Wend, K.; Wend, P.; Krum, S.A. Tissue-Specific Effects of Loss of Estrogen during Menopause and Aging. Front. Endocrinol. (Lausanne) 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Zhou, T.-H.; Gao, Y.; Zhang, N.; Li, J.-C. Ultrastructure and Estrogen Regulation of the Lymphatic Stomata of Ovarian Bursa in Mice. Anat. Rec. (Hoboken) 2007, 290, 1195–1202. [Google Scholar] [CrossRef]
- Hinshelwood, M.M.; Mendelson, C.R. Tissue-Specific Expression of the Human CYP19 (Aromatase) Gene in Ovary and Adipose Tissue of Transgenic Mice. J. Steroid. Biochem. Mol. Biol. 2001, 79, 193–201. [Google Scholar] [CrossRef]
- Manolagas, S.C.; O’Brien, C.A.; Almeida, M. The Role of Estrogen and Androgen Receptors in Bone Health and Disease. Nat. Rev. Endocrinol. 2013, 9, 699–712. [Google Scholar] [CrossRef] [PubMed]
- McTernan, P.G.; Anderson, L.A.; Anwar, A.J.; Eggo, M.C.; Crocker, J.; Barnett, A.H.; Stewart, P.M.; Kumar, S. Glucocorticoid Regulation of P450 Aromatase Activity in Human Adipose Tissue: Gender and Site Differences. J. Clin. Endocrinol. Metab. 2002, 87, 1327–1336. [Google Scholar] [CrossRef] [PubMed]
- Lapid, K.; Lim, A.; Clegg, D.J.; Zeve, D.; Graff, J.M. Oestrogen Signalling in White Adipose Progenitor Cells Inhibits Differentiation into Brown Adipose and Smooth Muscle Cells. Nat. Commun. 2014, 5, 5196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemieux, C.; Phaneuf, D.; Labrie, F.; Giguère, V.; Richard, D.; Deshaies, Y. Estrogen Receptor Alpha-Mediated Adiposity-Lowering and Hypocholesterolemic Actions of the Selective Estrogen Receptor Modulator Acolbifene. Int. J. Obes. (Lond.) 2005, 29, 1236–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieudonne, M.N.; Pecquery, R.; Leneveu, M.C.; Giudicelli, Y. Opposite Effects of Androgens and Estrogens on Adipogenesis in Rat Preadipocytes: Evidence for Sex and Site-Related Specificities and Possible Involvement of Insulin-like Growth Factor 1 Receptor and Peroxisome Proliferator-Activated Receptor Gamma2. Endocrinology 2000, 141, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Thornton, J.W.; Need, E.; Crews, D. Resurrecting the Ancestral Steroid Receptor: Ancient Origin of Estrogen Signaling. Science 2003, 301, 1714–1717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krust, A.; Green, S.; Argos, P.; Kumar, V.; Walter, P.; Bornert, J.M.; Chambon, P. The Chicken Oestrogen Receptor Sequence: Homology with v-ErbA and the Human Oestrogen and Glucocorticoid Receptors. EMBO J. 1986, 5, 891–897. [Google Scholar] [CrossRef]
- Liu, S.; Ding, T.; Liu, H.; Jian, L. GPER Was Associated with Hypertension in Post-Menopausal Women. Open Med. (Wars) 2018, 13, 338–343. [Google Scholar] [CrossRef]
- Arnal, J.-F.; Fontaine, C.; Abot, A.; Valera, M.-C.; Laurell, H.; Gourdy, P.; Lenfant, F. Lessons from the Dissection of the Activation Functions (AF-1 and AF-2) of the Estrogen Receptor Alpha In Vivo. Steroids 2013, 78, 576–582. [Google Scholar] [CrossRef]
- Safe, S.; Kim, K. Non-Classical Genomic Estrogen Receptor (ER)/Specificity Protein and ER/Activating Protein-1 Signaling Pathways. J. Mol. Endocrinol. 2008. [Google Scholar] [CrossRef] [PubMed]
- Chambliss, K.L.; Shaul, P.W. Estrogen Modulation of Endothelial Nitric Oxide Synthase. Endocr. Rev. 2002, 23, 665–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, V.M.; Clarkson, T.B.; Harman, S.M.; Brinton, E.A.; Cedars, M.; Lobo, R.; Manson, J.E.; Merriam, G.R.; Naftolin, F.; Santoro, N. Women, Hormones, and Clinical Trials: A Beginning, Not an End. J. Appl. Physiol. 2005, 99, 381–383. [Google Scholar] [CrossRef] [PubMed]
- Mendelsohn, M.E.; Karas, R.H. Molecular and Cellular Basis of Cardiovascular Gender Differences. Science 2005, 308, 1583–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, E.R. Integration of the Extranuclear and Nuclear Actions of Estrogen. Mol. Endocrinol. 2005, 19, 1951–1959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garmy-Susini, B.; Delmas, E.; Gourdy, P.; Zhou, M.; Bossard, C.; Bugler, B.; Bayard, F.; Krust, A.; Prats, A.C.; Doetschman, T.; et al. Role of Fibroblast Growth Factor-2 Isoforms in the Effect of Estradiol on Endothelial Cell Migration and Proliferation. Circ. Res. 2004, 94, 1301–1309. [Google Scholar] [CrossRef]
- Guo, S.; Colbert, L.S.; Fuller, M.; Zhang, Y.; Gonzalez-Perez, R.R. Vascular Endothelial Growth Factor Receptor-2 in Breast Cancer. Biochim. Biophys. Acta 2010, 1806, 108–121. [Google Scholar] [CrossRef] [Green Version]
- Gormsen, L.C.; Høst, C.; Hjerrild, B.E.; Pedersen, S.B.; Nielsen, S.; Christiansen, J.S.; Gravholt, C.H. Estradiol Acutely Inhibits Whole Body Lipid Oxidation and Attenuates Lipolysis in Subcutaneous Adipose Tissue: A Randomized, Placebo-Controlled Study in Postmenopausal Women. Eur. J. Endocrinol. 2012, 167, 543–551. [Google Scholar] [CrossRef] [Green Version]
- Iorga, A.; Cunningham, C.M.; Moazeni, S.; Ruffenach, G.; Umar, S.; Eghbali, M. The Protective Role of Estrogen and Estrogen Receptors in Cardiovascular Disease and the Controversial Use of Estrogen Therapy. Biol. Sex Differ. 2017, 8, 33. [Google Scholar] [CrossRef]
- Schrott, H.G.; Bittner, V.; Vittinghoff, E.; Herrington, D.M.; Hulley, S. Adherence to National Cholesterol Education Program Treatment Goals in Postmenopausal Women with Heart Disease. The Heart and Estrogen/Progestin Replacement Study (HERS). The HERS Research Group. Jama 1997, 277, 1281–1286. [Google Scholar] [CrossRef]
- Mikkola, T.S.; Anthony, M.S.; Clarkson, T.B.; St Clair, R.W. Serum Cholesterol Efflux Potential in Postmenopausal Monkeys Treated with Tibolone or Conjugated Estrogens. Metabolism 2002, 51, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, M.E.; Kauser, K.; Martin-McNulty, B.; Polinsky, P.; Schwartz, S.M.; Rubanyi, G.M. Estrogen Inhibits the Initiation of Fatty Streaks throughout the Vasculature but Does Not Inhibit Intra-Plaque Hemorrhage and the Progression of Established Lesions in Apolipoprotein E Deficient Mice. Atherosclerosis 2002, 164, 251–259. [Google Scholar] [CrossRef]
- Billon-Galés, A.; Fontaine, C.; Douin-Echinard, V.; Delpy, L.; Berges, H.; Calippe, B.; Lenfant, F.; Laurell, H.; Guéry, J.-C.; Gourdy, P.; et al. Endothelial Estrogen Receptor-α Plays a Crucial Role in the Atheroprotective Action of 17β-Estradiol in Low-Density Lipoprotein Receptor–Deficient Mice. Circulation 2009, 120, 2567–2576. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, C.; Morfoisse, F.; Tatin, F.; Zamora, A.; Zahreddine, R.; Henrion, D.; Arnal, J.-F.; Lenfant, F.; Garmy-Susini, B. The Impact of Estrogen Receptor in Arterial and Lymphatic Vascular Diseases. Int. J. Mol. Sci. 2020, 21, 3244. [Google Scholar] [CrossRef]
- Dufour, C.R.; Levasseur, M.-P.; Pham, N.H.H.; Eichner, L.J.; Wilson, B.J.; Charest-Marcotte, A.; Duguay, D.; Poirier-Héon, J.-F.; Cermakian, N.; Giguère, V. Genomic Convergence among ERRα, PROX1, and BMAL1 in the Control of Metabolic Clock Outputs. PLoS Genet. 2011, 7, e1002143. [Google Scholar] [CrossRef]
- Charest-Marcotte, A.; Dufour, C.R.; Wilson, B.J.; Tremblay, A.M.; Eichner, L.J.; Arlow, D.H.; Mootha, V.K.; Giguère, V. The Homeobox Protein Prox1 Is a Negative Modulator of ERRα/PGC-1α Bioenergetic Functions. Genes Dev. 2010, 24, 537–542. [Google Scholar] [CrossRef] [Green Version]
- Wong, B.W.; Wang, X.; Zecchin, A.; Thienpont, B.; Cornelissen, I.; Kalucka, J.; García-Caballero, M.; Missiaen, R.; Huang, H.; Brüning, U.; et al. The Role of Fatty Acid β-Oxidation in Lymphangiogenesis. Nature 2017, 542, 49–54. [Google Scholar] [CrossRef]
- Deblois, G.; Hall, J.A.; Perry, M.-C.; Laganière, J.; Ghahremani, M.; Park, M.; Hallett, M.; Giguère, V. Genome-Wide Identification of Direct Target Genes Implicates Estrogen-Related Receptor Alpha as a Determinant of Breast Cancer Heterogeneity. Cancer Res. 2009, 69, 6149–6157. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, H.; Takahashi, E.; Kobayashi, M.; Goto, M.; Krust, A.; Chambon, P.; Iguchi, T. The Estrogen-Responsive Adrenomedullin and Receptor-Modifying Protein 3 Gene Identified by DNA Microarray Analysis Are Directly Regulated by Estrogen Receptor. J. Mol. Endocrinol. 2006, 36, 81–89. [Google Scholar] [CrossRef]
- Pearson, L.J.; Rait, C.; Nicholls, M.G.; Yandle, T.G.; Evans, J.J. Regulation of Adrenomedullin Release from Human Endothelial Cells by Sex Steroids and Angiotensin-II. J. Endocrinol. 2006, 191, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Shur, I.; Zemer-Tov, E.; Socher, R.; Benayahu, D. SVEP1 Expression Is Regulated in Estrogen-Dependent Manner. J. Cell. Physiol. 2007, 210, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Stangl, H.; Graf, G.; Yu, L.; Cao, G.; Wyne, K. Effect of Estrogen on Scavenger Receptor BI Expression in the Rat. J. Endocrinol. 2003, 175, 663–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, H.Y.; Thiam, C.H.; Yeo, K.P.; Bisoendial, R.; Hii, C.S.; McGrath, K.C.Y.; Tan, K.W.; Heather, A.; Alexander, J.S.J.; Angeli, V. Lymphatic Vessels Are Essential for the Removal of Cholesterol from Peripheral Tissues by SR-BI-Mediated Transport of HDL. Cell Metab. 2013, 17, 671–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaverien, M.V.; Coroneos, C.J. Surgical Treatment of Lymphedema. Plast. Reconstr. Surg. 2019, 144, 738–758. [Google Scholar] [CrossRef] [PubMed]
- Smeltzer, D.M.; Stickler, G.B.; Schirger, A. Primary Lymphedema in Children and Adolescents: A Follow-up Study and Review. Pediatrics 1985, 76, 206–218. [Google Scholar]
- Kinmonth, J.B.; Taylor, G.W.; Tracy, G.D.; Marsh, J.D. Primary Lymphoedema; Clinical and Lymphangiographic Studies of a Series of 107 Patients in Which the Lower Limbs Were Affected. Br. J. Surg. 1957, 45, 1–9. [Google Scholar] [CrossRef]
- Brunner, U. Natural History of Primary Lymphedema of the Legs. Pathol. Microbiol. 1975, 43, 230–234. [Google Scholar] [CrossRef]
- Gordon, K.; Varney, R.; Keeley, V.; Riches, K.; Jeffery, S.; Van Zanten, M.; Mortimer, P.; Ostergaard, P.; Mansour, S. Update and Audit of the St George’s Classification Algorithm of Primary Lymphatic Anomalies: A Clinical and Molecular Approach to Diagnosis. J. Med. Genet. 2020, 57, 653–659. [Google Scholar] [CrossRef]
- Greene, R.; Fowler, R. Physical Therapy Management of Primary Lymphedema in the Lower Extremities: A Case Report. Physiother. Theory Pract. 2010, 26, 62–68. [Google Scholar] [CrossRef]
- McCarthy, K.; Bondy, C.A. Turner Syndrome in Childhood and Adolescence. Expert Rev. Endocrinol. Metab. 2008, 3, 771–775. [Google Scholar] [CrossRef] [Green Version]
- Shankar, R.K.; Backeljauw, P.F. Current Best Practice in the Management of Turner Syndrome. Ther. Adv. Endocrinol. Metab. 2018, 9, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gravholt, C.H.; Andersen, N.H.; Conway, G.S.; Dekkers, O.M.; Geffner, M.E.; Klein, K.O.; Lin, A.E.; Mauras, N.; Quigley, C.A.; Rubin, K.; et al. Clinical Practice Guidelines for the Care of Girls and Women with Turner Syndrome: Proceedings from the 2016 Cincinnati International Turner Syndrome Meeting. Eur. J. Endocrinol. 2017, 177, G1–G70. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, P.S.; Rockson, S.G. New Developments in Clinical Aspects of Lymphatic Disease. J. Clin. Investig. 2014, 124, 915–921. [Google Scholar] [CrossRef] [PubMed]
- Dale, R.F. The Inheritance of Primary Lymphoedema. J. Med. Genet. 1985, 22, 274–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Zhang, L.; Ding, M.; Luo, Z.; Yuan, S.; Bansal, M.B.; Gilkeson, G.; Lang, R.; Jiang, W. Estrogen Decreases Tight Junction Protein ZO-1 Expression in Human Primary Gut Tissues. Clin. Immunol. 2017, 183, 174–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baluk, P.; Fuxe, J.; Hashizume, H.; Romano, T.; Lashnits, E.; Butz, S.; Vestweber, D.; Corada, M.; Molendini, C.; Dejana, E.; et al. Functionally Specialized Junctions between Endothelial Cells of Lymphatic Vessels. J. Exp. Med. 2007, 204, 2349–2362. [Google Scholar] [CrossRef]
- Silha, J.V.; Krsek, M.; Sucharda, P.; Murphy, L.J. Angiogenic Factors Are Elevated in Overweight and Obese Individuals. Int. J. Obes. (Lond.) 2005, 29, 1308–1314. [Google Scholar] [CrossRef] [Green Version]
- Stachenfeld, N.S. Sex Hormone Effects on Body Fluid Regulation. Exerc. Sport. Sci. Rev. 2008, 36, 152–159. [Google Scholar] [CrossRef] [Green Version]
- Schook, C.C.; Mulliken, J.B.; Fishman, S.J.; Grant, F.D.; Zurakowski, D.; Greene, A.K. Primary Lymphedema: Clinical Features and Management in 138 Pediatric Patients. Plast. Reconstr. Surg. 2011, 127, 2419–2431. [Google Scholar] [CrossRef]
- Heisch, R.B.; Nelson, G.S.; Furlong, M. Studies in Filariasis in East Africa. 1. Filariasis on the Island of Pate, Kenya. Trans. R. Soc. Trop. Med. Hyg. 1959, 53, 41–53. [Google Scholar] [CrossRef]
- Babu, S.; Nutman, T.B. Immunology of Lymphatic Filariasis. Parasite Immunol. 2014, 36, 338–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobson, C. The Age and Sex of the Host as Factors Affecting the Host--Parasite Relationship of the Third-Stage Larva of Amplicaecum Robertsi Sprent & Mines, 1960, in the Laboratory Mouse. Parasitology 1966, 56, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, H.; Horii, Y.; Fujita, K. Effect of Testosterone on the Eosinophil Response of C57BL/6 Mice to Infection with Brugia Pahangi. Immunopharmacology 1992, 23, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, Y.; Berkovich, Z. Testosterone Effect on Bone Marrow, Thymus, and Suppressor T Cells in the (NZB X NZW)F1 Mice: Its Relevance to Autoimmunity. J. Immunol. 1981, 126, 998–1002. [Google Scholar] [PubMed]
- Rajan, T.V.; Nelson, F.K.; Shultz, L.D.; Shultz, K.L.; Beamer, W.G.; Yates, J.; Greiner, D.L. Influence of Gonadal Steroids on Susceptibility to Brugia Malayi in Scid Mice. Acta Trop. 1994, 56, 307–314. [Google Scholar] [CrossRef]
- Ganley, L.; Rajan, T.V. Endogenous Testosterone Levels Do Not Affect Filarial Worm Burdens in Mice. Exp. Parasitol. 2001, 98, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Brabin, L. Sex Differentials in Susceptibility to Lymphatic Filariasis and Implications for Maternal Child Immunity. Epidemiol. Infect. 1990, 105, 335–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernas, M.; Thiadens, S.R.J.; Smoot, B.; Armer, J.M.; Stewart, P.; Granzow, J. Lymphedema Following Cancer Therapy: Overview and Options. Clin. Exp. Metastasis 2018, 35, 547–551. [Google Scholar] [CrossRef]
- Rockson, S.G. Lymphedema after Breast Cancer Treatment. N. Engl. J. Med. 2018, 379, 1937–1944. [Google Scholar] [CrossRef]
- DiSipio, T.; Rye, S.; Newman, B.; Hayes, S. Incidence of Unilateral Arm Lymphoedema after Breast Cancer: A Systematic Review and Meta-Analysis. Lancet Oncol. 2013, 14, 500–515. [Google Scholar] [CrossRef]
- Tiwari, P.; Coriddi, M.; Salani, R.; Povoski, S.P. Breast and Gynecologic Cancer-Related Extremity Lymphedema: A Review of Diagnostic Modalities and Management Options. World J. Surg. Oncol. 2013, 11, 237. [Google Scholar] [CrossRef] [Green Version]
- Meneses, K.D.; McNees, M.P. Upper Extremity Lymphedema after Treatment for Breast Cancer: A Review of the Literature. Ostomy Wound Manag. 2007, 53, 16–29. [Google Scholar]
- Jeselsohn, R.; Buchwalter, G.; De Angelis, C.; Brown, M.; Schiff, R. ESR1 Mutations—a Mechanism for Acquired Endocrine Resistance in Breast Cancer. Nat. Rev. Clin. Oncol. 2015, 12, 573–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komm, B.S. A New Approach to Menopausal Therapy: The Tissue Selective Estrogen Complex. Reprod. Sci. 2008, 15, 984–992. [Google Scholar] [CrossRef] [PubMed]
- Berrodin, T.J.; Chang, K.C.N.; Komm, B.S.; Freedman, L.P.; Nagpal, S. Differential Biochemical and Cellular Actions of Premarin Estrogens: Distinct Pharmacology of Bazedoxifene-Conjugated Estrogens Combination. Mol. Endocrinol. 2009, 23, 74–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kharode, Y.; Bodine, P.V.N.; Miller, C.P.; Lyttle, C.R.; Komm, B.S. The Pairing of a Selective Estrogen Receptor Modulator, Bazedoxifene, with Conjugated Estrogens as a New Paradigm for the Treatment of Menopausal Symptoms and Osteoporosis Prevention. Endocrinology 2008, 149, 6084–6091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazarali, S.A.; Narod, S.A. Tamoxifen for Women at High Risk of Breast Cancer. Breast Cancer (Dove. Med. Press.) 2014, 6, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.-L.; Cheng, M.-H.; Chao, H.-T.; Wang, P.-H. The Role of Selective Estrogen Receptor Modulators on Breast Cancer: From Tamoxifen to Raloxifene. Taiwan J. Obstet. Gynecol. 2008, 47, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Leignadier, J.; Dalenc, F.; Poirot, M.; Silvente-Poirot, S. Improving the Efficacy of Hormone Therapy in Breast Cancer: The Role of Cholesterol Metabolism in SERM-Mediated Autophagy, Cell Differentiation and Death. Biochem. Pharmacol. 2017, 144, 18–28. [Google Scholar] [CrossRef]
- Jordan, V.C. Tamoxifen: A Most Unlikely Pioneering Medicine. Nat. Rev. Drug. Discov. 2003, 2, 205–213. [Google Scholar] [CrossRef]
- Das, N.; Baumgartner, R.N.; Riley, E.C.; Pinkston, C.M.; Yang, D.; Baumgartner, K.B. Treatment-Related Risk Factors for Arm Lymphedema among Long-Term Breast Cancer Survivors. J. Cancer Surviv. 2015, 9, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Shrubb, D.; Mason, W. The Management of Deep Vein Thrombosis in Lymphoedema: A Review. Br. J. Community Nurs. 2006, 11, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Garmy-Susini, B.; UMR 1297-I2MC, Inserm, Université de Toulouse, UT3, Toulouse, France; Morfoisse, F.; UMR 1297-I2MC, Inserm, Université de Toulouse, UT3, Toulouse, France. Personal communication, 2018.
- Henri, O.; Pouehe, C.; Houssari, M.; Galas, L.; Nicol, L.; Edwards-Lévy, F.; Henry, J.-P.; Dumesnil, A.; Boukhalfa, I.; Banquet, S.; et al. Selective Stimulation of Cardiac Lymphangiogenesis Reduces Myocardial Edema and Fibrosis Leading to Improved Cardiac Function Following Myocardial Infarction. Circulation 2016, 133, 1484–1497, discussion 1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldberg, J.I.; Riedel, E.R.; Morrow, M.; Van Zee, K.J. Morbidity of Sentinel Node Biopsy: Relationship between Number of Excised Lymph Nodes and Patient Perceptions of Lymphedema. Ann. Surg. Oncol. 2011, 18, 2866–2872. [Google Scholar] [CrossRef] [PubMed]
- Garmy-Susini, B. Hormone Therapy Outcome in Lymphedema. Aging (Albany NY) 2019, 11, 291–292. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morfoisse, F.; Zamora, A.; Marchaud, E.; Nougue, M.; Diallo, L.H.; David, F.; Roussel, E.; Lacazette, E.; Prats, A.-C.; Tatin, F.; et al. Sex Hormones in Lymphedema. Cancers 2021, 13, 530. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13030530
Morfoisse F, Zamora A, Marchaud E, Nougue M, Diallo LH, David F, Roussel E, Lacazette E, Prats A-C, Tatin F, et al. Sex Hormones in Lymphedema. Cancers. 2021; 13(3):530. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13030530
Chicago/Turabian StyleMorfoisse, Florent, Audrey Zamora, Emmanuelle Marchaud, Manon Nougue, Leila H. Diallo, Florian David, Emilie Roussel, Eric Lacazette, Anne-Catherine Prats, Florence Tatin, and et al. 2021. "Sex Hormones in Lymphedema" Cancers 13, no. 3: 530. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13030530