
Key Enzymes in Pyrimidine Synthesis, CAD and CPS1, Predict Prognosis in Hepatocellular Carcinoma

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
2.1. CAD Is Induced in Recurrent Hepatocellular Carcinoma and Predicts Prognosis
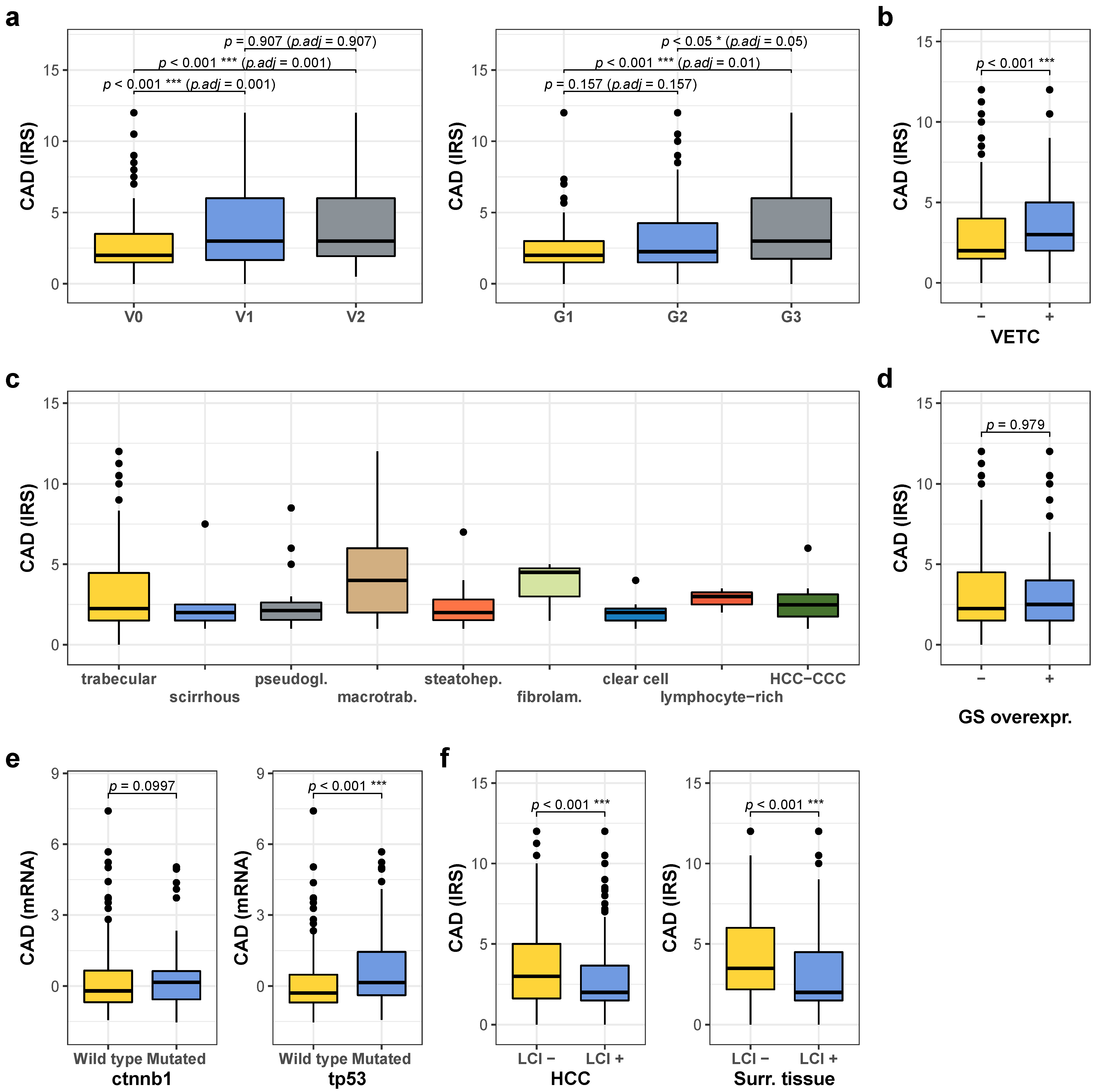
2.2. High CAD Expression Is Associated with Unfavorable Prognostic Factors
2.3. CPS1 Is Downregulated in Hepatocellular Carcinoma, Further Reduced in Recurrent Tumors and Metastases, and Predicts Prognosis
2.4. Low CPS1 Expression Is Associated with Unfavorable Prognostic Factors
2.5. Combined Immunoreactive Score of CAD and CPS1 Improves Prognostic Prediction and Is Particularly Predictive in Early Stage HCC without Vascular Invasion
3. Discussion
4. Materials and Methods
4.1. Patients and Samples
4.2. Cell Culture
4.3. Immunohistochemistry
4.4. Immunofluorescence
4.5. Immunoblotting
4.6. Analysis of Publicly Available Data Sets
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
- Anti-AFP (Dako, Hamburg, Germany, #IR500, rabbit polyclonal, ready to use, Tris/EDTA buffer, pH 9 (Dako #8024))
- Anti-CPS1 (HepPar1, Dako, #IR624, mouse monoclonal, ready to use, Tris/EDTA buffer, pH 9)
- Anti-CAD (Santa Cruz, Heidelberg, Germany, #sc-376072, mouse monoclonal, 1:75, Tris/EDTA buffer, pH 9)
- Anti-cleaved Caspase 3 (Cell Signaling, Frankfurt, Germany, #9664, rabbit monoclonal, 1:200, citrate buffer, pH = 6.1 (Dako #GV805))
- Anti-CD34 (Dako, #IR632, mouse monoclonal, ready to use, Tris/EDTA buffer, pH 9)
- Anti-CK19 (Dako #IR615, mouse monoclonal, ready to use, Tris/EDTA buffer, pH 9)
- Anti-Glutaminsynthetase (Roche, Mannheim, Germany, #760-4898, mouse monoclonal, ready to use, cell conditioning solution 1 (Roche, #950-124))
- Anti-Glypican 3 (Roche, #790-4564, mouse monoclonal, ready to use, cell conditioning solution 1 (Roche, #950-124))
- Anti-γH2AX (Cell Signaling, #9718, rabbit monoclonal, 1:200, citrate buffer, pH = 6.1)
- Anti-HSP70 (Santa Cruz, #sc-24, 1:250, cell conditioning solution 1 (Roche, #950-124))
- Anti-Ki67 (Dako, #IR626, mouse monoclonal, ready to use, citrate buffer, pH = 6.1)
- Anti-ZEB1 (Novusbio, Centennial CO, USA, NBP1-05987, rabbit polyclonal, 1:500, Tris/EDTA buffer, pH 9)
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- European Association for the Study of the Liver. Clinical Practice Guidelines: Management of Hepatocellular Carcinoma. J. Hepatol. 2018, 69, 182–236. [Google Scholar] [CrossRef] [PubMed]
- Pavlova, N.N.; Thompson, C.B. The Emerging Hallmarks of Cancer Metabolism. Cell Metab. 2016, 23, 27–47. [Google Scholar] [CrossRef]
- Ah Mew, N.; Simpson, K.L.; Gropman, A.L.; Lanpher, B.C.; Chapman, K.A.; Summar, M.L. Urea Cycle Disorders Overview. In GeneReviews®; Adam, M.P., Ardinger, H.H., Pagon, R.A., Wallace, S.E., Bean, L.J., Stephens, K., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Keshet, R.; Szlosarek, P.; Carracedo, A.; Erez, A. Rewiring Urea Cycle Metabolism in Cancer to Support Anabolism. Nat. Rev. Cancer 2018, 18, 634–645. [Google Scholar] [CrossRef]
- Lee, J.S.; Adler, L.; Karathia, H.; Carmel, N.; Rabinovich, S.; Auslander, N.; Keshet, R.; Stettner, N.; Silberman, A.; Agemy, L.; et al. Urea Cycle Dysregulation Generates Clinically Relevant Genomic and Biochemical Signatures. Cell 2018, 174, 1559–1570.e22. [Google Scholar] [CrossRef]
- Wang, X.; Yang, K.; Wu, Q.; Kim, L.J.Y.; Morton, A.R.; Gimple, R.C.; Prager, B.C.; Shi, Y.; Zhou, W.; Bhargava, S.; et al. Targeting Pyrimidine Synthesis Accentuates Molecular Therapy Response in Glioblastoma Stem Cells. Sci. Transl. Med. 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.R.; Guy, H.I. Mammalian Pyrimidine Biosynthesis: Fresh Insights into an Ancient Pathway. J. Biol. Chem. 2004, 279, 33035–33038. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; Weber, G. Carbamoyl Phosphate Synthetase (Glutamine-Hydrolyzing): Increased Activity in Cancer Cells. Science 1981, 212, 463–465. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Wang, X.; Li, X.; Xu, G.; Bai, Y.; Wu, J.; Piao, Y.; Shi, Y.; Xiang, R.; Wang, L. Nucleotide de Novo Synthesis Increases Breast Cancer Stemness and Metastasis via CGMP-PKG-MAPK Signaling Pathway. PLoS Biol. 2020, 18, e3000872. [Google Scholar] [CrossRef]
- Morin, A.; Fritsch, L.; Mathieu, J.R.R.; Gilbert, C.; Guarmit, B.; Firlej, V.; Gallou-Kabani, C.; Vieillefond, A.; Delongchamps, N.B.; Cabon, F. Identification of CAD as an Androgen Receptor Interactant and an Early Marker of Prostate Tumor Recurrence. FASEB J. 2012, 26, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Butler, S.L.; Dong, H.; Cardona, D.; Jia, M.; Zheng, R.; Zhu, H.; Crawford, J.M.; Liu, C. The Antigen for Hep Par 1 Antibody Is the Urea Cycle Enzyme Carbamoyl Phosphate Synthetase 1. Lab. Invest. 2008, 88, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Wennerberg, A.E.; Nalesnik, M.A.; Coleman, W.B. Hepatocyte Paraffin 1: A Monoclonal Antibody That Reacts with Hepatocytes and Can Be Used for Differential Diagnosis of Hepatic Tumors. Am. J. Pathol. 1993, 143, 1050–1054. [Google Scholar] [PubMed]
- Liu, H.; Dong, H.; Robertson, K.; Liu, C. DNA Methylation Suppresses Expression of the Urea Cycle Enzyme Carbamoyl Phosphate Synthetase 1 (CPS1) in Human Hepatocellular Carcinoma. Am. J. Pathol. 2011, 178, 652–661. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Research Network. Comprehensive and Integrative Genomic Characterization of Hepatocellular Carcinoma. Cell 2017, 169, 1327–1341.e23. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Ding, W.; Zhang, J.; Gao, Q.; Yang, H.; Cao, W.; Wang, Z.; Fang, L.; Du, R. Significant Down-Regulation of Urea Cycle Generates Clinically Relevant Proteomic Signature in Hepatocellular Carcinoma Patients with Macrovascular Invasion. J. Proteome Res. 2019, 18, 2032–2044. [Google Scholar] [CrossRef]
- Dumenci, O.E.; MR U, A.; Khan, S.A.; Holmes, E.; Taylor-Robinson, S.D. Exploring Metabolic Consequences of CPS1 and CAD Dysregulation in Hepatocellular Carcinoma by Network Reconstruction. J. Hepatocell. Carcinoma 2020, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Remmele, W.; Stegner, H.E. Recommendation for uniform definition of an immunoreactive score (IRS) for immunohistochemical estrogen receptor detection (ER-ICA) in breast cancer tissue. Pathologe 1987, 8, 138–140. [Google Scholar] [PubMed]
- Trevisani, F.; Garuti, F.; Cucchetti, A.; Lenzi, B.; Bernardi, M. De Novo Hepatocellular Carcinoma of Liver Allograft: A Neglected Issue. Cancer Lett 2015, 357, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Imamura, H.; Matsuyama, Y.; Tanaka, E.; Ohkubo, T.; Hasegawa, K.; Miyagawa, S.; Sugawara, Y.; Minagawa, M.; Takayama, T.; Kawasaki, S.; et al. Risk Factors Contributing to Early and Late Phase Intrahepatic Recurrence of Hepatocellular Carcinoma after Hepatectomy. J. Hepatol. 2003, 38, 200–207. [Google Scholar] [CrossRef]
- Zheng, J.; Chou, J.F.; Gönen, M.; Vachharajani, N.; Chapman, W.C.; Majella Doyle, M.B.; Turcotte, S.; Vandenbroucke-Menu, F.; Lapointe, R.; Buettner, S.; et al. Prediction of Hepatocellular Carcinoma Recurrence Beyond Milan Criteria After Resection: Validation of a Clinical Risk Score in an International Cohort. Ann. Surg. 2017, 266, 693–701. [Google Scholar] [CrossRef]
- Poon, R.T.P. Differentiating Early and Late Recurrences after Resection of HCC in Cirrhotic Patients: Implications on Surveillance, Prevention, and Treatment Strategies. Ann. Surg. Oncol. 2009, 16, 792–794. [Google Scholar] [CrossRef]
- Du, Z.-G.; Wei, Y.-G.; Chen, K.-F.; Li, B. Risk Factors Associated with Early and Late Recurrence after Curative Resection of Hepatocellular Carcinoma: A Single Institution’s Experience with 398 Consecutive Patients. Hepatobiliary Pancreat. Dis. Int. 2014, 13, 153–161. [Google Scholar] [CrossRef]
- Boyd, K.E.; Farnham, P.J. Myc versus USF: Discrimination at the Cad Gene Is Determined by Core Promoter Elements. Mol. Cell Biol. 1997, 17, 2529–2537. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Abdelrahim, M.; Samudio, I.; Safe, S. Estrogen Receptor/Sp1 Complexes Are Required for Induction of Cad Gene Expression by 17beta-Estradiol in Breast Cancer Cells. Endocrinology 2003, 144, 2325–2335. [Google Scholar] [CrossRef] [PubMed]
- Renne, S.L.; Woo, H.Y.; Allegra, S.; Rudini, N.; Yano, H.; Donadon, M.; Viganò, L.; Akiba, J.; Lee, H.S.; Rhee, H.; et al. Vessels Encapsulating Tumor Clusters (VETC) Is a Powerful Predictor of Aggressive Hepatocellular Carcinoma. Hepatology 2020, 71, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Lokuhetty, D. WHO Classification of Tumours; WHO: Geneva, Switzerland, 2019; ISBN 978-92-832-4499-8. [Google Scholar]
- Ziol, M.; Poté, N.; Amaddeo, G.; Laurent, A.; Nault, J.-C.; Oberti, F.; Costentin, C.; Michalak, S.; Bouattour, M.; Francoz, C.; et al. Macrotrabecular-Massive Hepatocellular Carcinoma: A Distinctive Histological Subtype with Clinical Relevance. Hepatology 2018, 68, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Zucman-Rossi, J.; Benhamouche, S.; Godard, C.; Boyault, S.; Grimber, G.; Balabaud, C.; Cunha, A.S.; Bioulac-Sage, P.; Perret, C. Differential Effects of Inactivated Axin1 and Activated Beta-Catenin Mutations in Human Hepatocellular Carcinomas. Oncogene 2007, 26, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.-M.; Cao, L.; Li, B.; Zhang, R.-X.; Sui, C.-J.; Yin, Z.-F.; Yang, J.-M. Clinicopathological Significance of ZEB1 Protein in Patients with Hepatocellular Carcinoma. Ann. Surg. Oncol. 2012, 19, 1700–1706. [Google Scholar] [CrossRef]
- Drápela, S.; Bouchal, J.; Jolly, M.K.; Culig, Z.; Souček, K. ZEB1: A Critical Regulator of Cell Plasticity, DNA Damage Response, and Therapy Resistance. Front. Mol. Biosci. 2020, 7, 36. [Google Scholar] [CrossRef]
- Kim, J.; Hu, Z.; Cai, L.; Li, K.; Choi, E.; Faubert, B.; Bezwada, D.; Rodriguez-Canales, J.; Villalobos, P.; Lin, Y.-F.; et al. CPS1 Maintains Pyrimidine Pools and DNA Synthesis in KRAS/LKB1-Mutant Lung Cancer Cells. Nature 2017, 546, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-R.; Sekine, K.; Nakamura, K.; Yanai, H.; Tanaka, M.; Miyajima, A. Y-Box Binding Protein-1 down-Regulates Expression of Carbamoyl Phosphate Synthetase-I by Suppressing CCAAT Enhancer-Binding Protein-Alpha Function in Mice. Gastroenterology 2009, 137, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Tang, N.; Wang, X.; Chen, Y. The Activity of the Carbamoyl Phosphate Synthase 1 Promoter in Human Liver-Derived Cells Is Dependent on Hepatocyte Nuclear Factor 3-Beta. J. Cell. Mol. Med. 2017, 21, 2036–2045. [Google Scholar] [CrossRef] [PubMed]
- Mondada, D.; Bosman, F.T.; Fontolliet, C.; Seelentag, W.K.F. Elevated Hepatocyte Paraffin 1 and Neprilysin Expression in Hepatocellular Carcinoma Are Correlated with Longer Survival. Virchows Arch. 2006, 448, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.C.; Gong, C.; Song, J.; Krausz, T.; Tretiakova, M.; Hyjek, E.; Al-Ahmadie, H.; Alves, V.; Xiao, S.-Y.; Anders, R.A.; et al. Arginase-1. Am. J. Surg. Pathol. 2010, 34, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Torzilli, G.; Belghiti, J.; Kokudo, N.; Takayama, T.; Capussotti, L.; Nuzzo, G.; Vauthey, J.-N.; Choti, M.A.; De Santibanes, E.; Donadon, M.; et al. A Snapshot of the Effective Indications and Results of Surgery for Hepatocellular Carcinoma in Tertiary Referral Centers: Is It Adherent to the EASL/AASLD Recommendations?: An Observational Study of the HCC East-West Study Group. Ann. Surg. 2013, 257, 929–937. [Google Scholar] [CrossRef]
- Di Sandro, S.; Centonze, L.; Pinotti, E.; Lauterio, A.; De Carlis, R.; Romano, F.; Gianotti, L.; De Carlis, L. NTF Research Group Surgical and Oncological Outcomes of Hepatic Resection for BCLC-B Hepatocellular Carcinoma: A Retrospective Multicenter Analysis among 474 Consecutive Cases. Updates Surg. 2019, 71, 285–293. [Google Scholar] [CrossRef]
- Furukawa, K.; Shiba, H.; Horiuchi, T.; Shirai, Y.; Haruki, K.; Fujiwara, Y.; Sakamoto, T.; Gocho, T.; Yanaga, K. Survival Benefit of Hepatic Resection for Hepatocellular Carcinoma beyond the Barcelona Clinic Liver Cancer Classification. J. Hepato-Biliary Pancreat. Sci. 2017, 24, 199–205. [Google Scholar] [CrossRef]
- Koh, Y.X.; Tan, H.L.; Lye, W.K.; Kam, J.H.; Chiow, A.K.H.; Tan, S.S.; Choo, S.P.; Chung, A.Y.F.; Goh, B.K.P. Systematic Review of the Outcomes of Surgical Resection for Intermediate and Advanced Barcelona Clinic Liver Cancer Stage Hepatocellular Carcinoma: A Critical Appraisal of the Evidence. World J. Hepatol. 2018, 10, 433–447. [Google Scholar] [CrossRef]
- Heinrich, S.; Sprinzl, M.; Schmidtmann, I.; Heil, E.; Koch, S.; Czauderna, C.; Heinrich, B.; Philippe, P.; Diggs, L.; Wörns, M.-A.; et al. Validation of Prognostic Accuracy of MESH, HKLC, and BCLC Classifications in a Large German Cohort of Hepatocellular Carcinoma Patients. United Eur. Gastroenterol. J. 2020, 8, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Sohn, J.H.; Duran, R.; Zhao, Y.; Fleckenstein, F.; Chapiro, J.; Sahu, S.P.; Schernthaner, R.E.; Qian, T.; Lee, H.; Zhao, L.; et al. Validation of the Hong Kong Liver Cancer Staging System in Determining Prognosis of the North American Patients Following Intra-Arterial Therapy. Clin. Gastroenterol. Hepatol. 2017, 15, 746–755.e4. [Google Scholar] [CrossRef] [PubMed]
- Adhoute, X.; Pénaranda, G.; Raoul, J.L.; Edeline, J.; Blanc, J.-F.; Pol, B.; Campanile, M.; Perrier, H.; Bayle, O.; Monnet, O.; et al. Barcelona Clinic Liver Cancer Nomogram and Others Staging/Scoring Systems in a French Hepatocellular Carcinoma Cohort. World J. Gastroenterol. 2017, 23, 2545–2555. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.-H.; Hsu, C.-Y.; Hsia, C.-Y.; Lee, Y.-H.; Huang, Y.-H.; Su, C.-W.; Lee, F.-Y.; Lin, H.-C.; Huo, T.-I. Proposal and Validation of a New Model to Estimate Survival for Hepatocellular Carcinoma Patients. Eur. J. Cancer 2016, 63, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Lai, Q.; Avolio, A.W.; Manzia, T.M.; Agnes, S.; Tisone, G.; Berloco, P.B.; Rossi, M. Role of Alpha-Fetoprotein in Selection of Patients with Hepatocellular Carcinoma Waiting for Liver Transplantation: Must We Reconsider It? Int. J. Biol. Markers 2011, 26, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Lomb, D.J.; Haigis, M.C.; Guarente, L. SIRT5 Deacetylates Carbamoyl Phosphate Synthetase 1 and Regulates the Urea Cycle. Cell 2009, 137, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Kozlowski, P.; Collins, M.; Wang, Y.; Haystead, T.A.; Graves, L.M. Caspase-Dependent Cleavage of Carbamoyl Phosphate Synthetase II during Apoptosis. Mol. Pharmacol. 2002, 61, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Ben-Sahra, I.; Howell, J.J.; Asara, J.M.; Manning, B.D. Stimulation of de Novo Pyrimidine Synthesis by Growth Signaling Through MTOR and S6K1. Science 2013, 339, 1323–1328. [Google Scholar] [CrossRef] [PubMed]
- Graves, L.M.; Guy, H.I.; Kozlowski, P.; Huang, M.; Lazarowski, E.; Pope, R.M.; Collins, M.A.; Dahlstrand, E.N.; Earp, H.S.; Evans, D.R. Regulation of Carbamoyl Phosphate Synthetase by MAP Kinase. Nature 2000, 403, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.S.; Sung, J.L.; Sheu, J.C.; Lai, M.Y.; How, S.W.; Hsu, H.C.; Lee, C.S.; Wei, T.C. Serum Alpha-Fetoprotein in the Early Stage of Human Hepatocellular Carcinoma. Gastroenterology 1984, 86, 1404–1409. [Google Scholar] [CrossRef]
- Saffroy, R.; Pham, P.; Reffas, M.; Takka, M.; Lemoine, A.; Debuire, B. New Perspectives and Strategy Research Biomarkers for Hepatocellular Carcinoma. Clin. Chem. Lab. Med. 2007, 45, 1169–1179. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, J.; Shen, F.; Lau, W.Y. Significance of Presence of Microvascular Invasion in Specimens Obtained after Surgical Treatment of Hepatocellular Carcinoma. J. Gastroenterol. Hepatol. 2018, 33, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, S.; Yamashita, S.; Matsumura, S.; Takeda, K.; Minagawa, T.; Kobayashi, S.; Hibi, T.; Shinoda, M.; Endo, I.; Tanabe, M.; et al. Significance of a Preoperative Tumor Marker Gradient for Predicting Microvascular Invasion in Cases of Hepatocellular Carcinoma. Mol. Clin. Oncol. 2020, 12, 290–294. [Google Scholar] [CrossRef] [PubMed]
- DeVito, S.R.; Ortiz-Riaño, E.; Martínez-Sobrido, L.; Munger, J. Cytomegalovirus-Mediated Activation of Pyrimidine Biosynthesis Drives UDP–Sugar Synthesis to Support Viral Protein Glycosylation. Proc. Natl. Acad. Sci. USA 2014, 111, 18019–18024. [Google Scholar] [CrossRef] [PubMed]
- Verrier, E.R.; Weiss, A.; Bach, C.; Heydmann, L.; Turon-Lagot, V.; Kopp, A.; El Saghire, H.; Crouchet, E.; Pessaux, P.; Garcia, T.; et al. Combined Small Molecule and Loss-of-Function Screen Uncovers Estrogen Receptor Alpha and CAD as Host Factors for HDV Infection and Antiviral Targets. Gut 2020, 69, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.; Li, J.; Wu, D.; Xia, Y.; Wang, Q.; Si, A.; Wang, K.; Wan, X.; Lau, W.Y.; Wu, M.; et al. Nomogram for Preoperative Estimation of Microvascular Invasion Risk in Hepatitis B Virus-Related Hepatocellular Carcinoma Within the Milan Criteria. JAMA Surg. 2016, 151, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, Y.-I.; Imai, K.; Yusa, T.; Nakao, Y.; Kitano, Y.; Nakagawa, S.; Okabe, H.; Chikamoto, A.; Ishiko, T.; Yoshizumi, T.; et al. Microvascular Invasion of Single Small Hepatocellular Carcinoma ≤3 Cm: Predictors and Optimal Treatments. Ann. Gastroenterol. Surg. 2018, 2, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, L.; Zhang, X.; Zhang, Y.; Liu, H.; Sun, B.; Zhao, L.; Ge, N.; Qian, H.; Yang, Y.; et al. Detection of Circulating Tumor Cells in Hepatocellular Carcinoma Using Antibodies against Asialoglycoprotein Receptor, Carbamoyl Phosphate Synthetase 1 and Pan-Cytokeratin. PLoS ONE 2014, 9, e96185. [Google Scholar] [CrossRef] [PubMed]
- Park, M.-J.; D’Alecy, L.G.; Anderson, M.A.; Basrur, V.; Feng, Y.; Brady, G.F.; Kim, D.; Wu, J.; Nesvizhskii, A.I.; Lahann, J.; et al. Constitutive Release of CPS1 in Bile and Its Role as a Protective Cytokine during Acute Liver Injury. Proc. Natl. Acad. Sci. USA 2019, 116, 9125–9134. [Google Scholar] [CrossRef]
- El-Sheikh, R.M.; Mansy, S.S.; Nessim, I.G.; Hosni, H.N.; El Hindawi, A.; Hassanein, M.H.; AbdelFattah, A.S. Carbamoyl Phosphate Synthetase 1 (CPS1) as a Prognostic Marker in Chronic Hepatitis C Infection. APMIS 2019, 127, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Uhlén, M.; Björling, E.; Agaton, C.; Szigyarto, C.A.-K.; Amini, B.; Andersen, E.; Andersson, A.-C.; Angelidou, P.; Asplund, A.; Asplund, C.; et al. A Human Protein Atlas for Normal and Cancer Tissues Based on Antibody Proteomics. Mol. Cell Proteomics 2005, 4, 1920–1932. [Google Scholar] [CrossRef] [PubMed]
- Weinmann, A.; Koch, S.; Niederle, I.M.; Schulze-Bergkamen, H.; König, J.; Hoppe-Lotichius, M.; Hansen, T.; Pitton, M.B.; Düber, C.; Otto, G.; et al. Trends in Epidemiology, Treatment, and Survival of Hepatocellular Carcinoma Patients between 1998 and 2009: An Analysis of 1066 Cases of a German HCC Registry. J. Clin. Gastroenterol. 2014, 48, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The CBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Zhang, C.; Lee, S.; Sjöstedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A Pathology Atlas of the Human Cancer Transcriptome. Science 2017, 357. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://Www.R-Project.Org/ (accessed on 11 February 2021).
- Budczies, J.; Klauschen, F.; Sinn, B.V.; Győrffy, B.; Schmitt, W.D.; Darb-Esfahani, S.; Denkert, C. Cutoff Finder: A Comprehensive and Straightforward Web Application Enabling Rapid Biomarker Cutoff Optimization. PLoS ONE 2012, 7, e51862. [Google Scholar] [CrossRef] [PubMed]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S. In Modern Applied Statistics with S.; Venables, W.N., Ripley, B.D., Eds.; Springer: New York, NY, USA, 2002; pp. 301–330. ISBN 978-0-387-21706-2. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | CAD Low | CAD High | p-Value 2 | |
|---|---|---|---|---|
| N = 459 (86%) 1 | N = 74 (14%) 1 | |||
| Median age in years (range) | 64.7 (56.2, 70.7) | 62.6 (56.8, 72.3) | 0.9 | |
| Median tumor size in mm | 40.0 (25.0, 78.0) | 45.0 (27.5, 96.5) | 0.2 | |
| Number of tumors | 0.6 | |||
| 1 | 244 (63%) | 44 (67%) | ||
| ≥2 | 146 (37%) | 22 (33%) | ||
| Gender | 0.042 | |||
| Male | 368 (80%) | 51 (69%) | ||
| Female | 91 (20%) | 23 (31%) | ||
| Etiology of liver disease | ||||
| Alcohol abuse | 144 (31%) | 14 (19%) | 0.041 | |
| HCV | 93 (20%) | 14 (19%) | >0.9 | |
| HBV | 88 (19%) | 11 (15%) | 0.5 | |
| NASH | 34 (7.4%) | 8 (11%) | 0.4 | |
| Hemochromatosis | 20 (4.4%) | 4 (5.4%) | 0.8 | |
| Unknown/Other | 111 (24%) | 15 (20%) | 0.6 | |
| BCLC | 0.006 | |||
| A | 88 (19%) | 10 (14%) | ||
| B | 290 (63%) | 50 (68%) | ||
| C | 47 (10%) | 14 (19%) | ||
| D | 34 (7.4%) | 0 (0%) | ||
| ECOG PST | 0.7 | |||
| 0–1 | 438 (96%) | 72 (100%) | ||
| 2 | 9 (2.0%) | 0 (0%) | ||
| 3 | 5 (1.1%) | 0 (0%) | ||
| 4 | 6 (1.3%) | 0 (0%) | ||
| Liver cirrhosis | 0.3 | |||
| Absent | 161 (35%) | 31 (42%) | ||
| Present | 298 (65%) | 43 (58%) | ||
| Child–Pugh score | 0.024 | |||
| A | 164 (52%) | 30 (61%) | ||
| B | 119 (38%) | 19 (39%) | ||
| C | 34 (11%) | 0 (0%) | ||
| Portal vein thrombosis | 48 (10%) | 16 (22%) | 0.011 | |
| Vascular invasion | <0.001 | |||
| Absent | 314 (68%) | 30 (41%) | ||
| Micro | 83 (18%) | 26 (35%) | ||
| Macro | 62 (14%) | 18 (24%) | ||
| Tumor grading | <0.001 | |||
| G1 | 95 (23%) | 4 (6.0%) | ||
| G2 | 229 (56%) | 36 (54%) | ||
| G3 | 88 (21%) | 27 (40%) | ||
| Macrotrabecular subtype | 0.082 | |||
| no | 420 (93%) | 64 (86%) | ||
| yes | 31 (6.9%) | 10 (14%) | ||
| VETC | 0.2 | |||
| negative | 382 (83%) | 56 (76%) | ||
| positive | 77 (17%) | 18 (24%) | ||
| GS overexpression | 0.3 | |||
| no | 360 (79%) | 63 (85%) | ||
| yes | 98 (21%) | 11 (15%) | ||
| Characteristic | CPS1 Low | CPS1 High | p-Value 2 | |
|---|---|---|---|---|
| N = 143 (27%) 1 | N = 393 (73%) 1 | |||
| Median age in years (range) | 64.0 (56.2, 69.3) | 64.8 (56.5, 71.3) | 0.2 | |
| Median tumor size in mm | 48.0 (29.0, 90.8) | 38.0 (24.0, 78.0) | 0.039 | |
| Number of tumors | >0.9 | |||
| 1 | 76 (63%) | 214 (64%) | ||
| ≥2 | 44 (37%) | 123 (36%) | ||
| Gender | 0.054 | |||
| Male | 104 (73%) | 318 (81%) | ||
| Female | 39 (27%) | 75 (19%) | ||
| Etiology of liver disease | ||||
| Alcohol abuse | 40 (28%) | 121 (31%) | 0.6 | |
| HCV | 24 (17%) | 83 (21%) | 0.3 | |
| HBV | 30 (21%) | 70 (18%) | 0.5 | |
| NASH | 10 (7.0%) | 31 (7.9%) | 0.9 | |
| Hemochromatosis | 8 (5.6%) | 17 (4.3%) | 0.7 | |
| Unknown/Other | 40 (28%) | 88 (22%) | 0.2 | |
| BCLC | 0.014 | |||
| A | 19 (13%) | 79 (20%) | ||
| B | 104 (73%) | 239 (61%) | ||
| C | 17 (12%) | 44 (11%) | ||
| D | 3 (2.1%) | 31 (7.9%) | ||
| ECOG PST | 0.7 | |||
| 0–1 | 137 (97%) | 376 (96%) | ||
| 2 | 2 (1.4%) | 7 (1.8%) | ||
| 3 | 0 (0%) | 5 (1.3%) | ||
| 4 | 2 (1.4%) | 4 (1.0%) | ||
| Liver cirrhosis | 0.008 | |||
| Absent | 65 (45%) | 128 (33%) | ||
| Present | 78 (55%) | 265 (67%) | ||
| Child–Pugh score | 0.11 | |||
| A | 47 (55%) | 146 (52%) | ||
| B | 35 (41%) | 105 (37%) | ||
| C | 3 (3.5%) | 31 (11%) | ||
| Portal vein thrombosis | 16 (11%) | 48 (12%) | 0.9 | |
| Vascular invasion | <0.001 | |||
| Absent | 76 (53%) | 271 (69%) | ||
| Micro | 47 (33%) | 62 (16%) | ||
| Macro | 20 (14%) | 60 (15%) | ||
| Tumor grading | <0.001 | |||
| G1 | 17 (13%) | 83 (24%) | ||
| G2 | 64 (49%) | 202 (58%) | ||
| G3 | 50 (38%) | 65 (19%) | ||
| Macrotrabecular subtype | <0.001 | |||
| no | 113 (79%) | 374 (97%) | ||
| yes | 30 (21%) | 10 (2.6%) | ||
| VETC | >0.9 | |||
| negative | 117 (82%) | 323 (82%) | ||
| positive | 26 (18%) | 69 (18%) | ||
| GS overexpression | 0.004 | |||
| no | 125 (88%) | 298 (76%) | ||
| yes | 17 (12%) | 93 (24%) | ||
| Characteristic | Univariate | Multivariate | ||||||
|---|---|---|---|---|---|---|---|---|
| HR | Conf.Low | Conf.High | p.Value | HR | Conf.Low | Conf.High | p.Value | |
| Clinical Features | ||||||||
| Age (≥60 vs. <60) | 1.43 | 1.13 | 1.80 | <0.01 | 1.44 | 1.13 | 1.84 | 0.003 |
| Sex (male vs. female) | 1.07 | 0.82 | 1.39 | 0.64 | ||||
| Alcohol abuse (tr. vs. f.) | 0.87 | 0.69 | 1.10 | 0.25 | ||||
| NASH (true vs. false) | 1.08 | 0.70 | 1.65 | 0.74 | ||||
| Haemochrom. (tr. vs. f.) | 0.70 | 0.41 | 1.19 | 0.19 | ||||
| HBV (pos. vs. neg.) | 0.89 | 0.67 | 1.17 | 0.40 | ||||
| HCV (pos. vs. neg.) | 1.07 | 0.82 | 1.40 | 0.61 | ||||
| Child–Pugh (B/C vs. A) | 0.97 | 0.74 | 1.26 | 0.81 | ||||
| ECOG (PST1-4 vs. PST0) | 1.43 | 1.13 | 1.79 | <0.01 | 1.47 | 1.15 | 1.87 | 0.002 |
| BCLC new (B-D vs. A) | 1.83 | 1.35 | 2.50 | <0.001 | 1.43 | 1.03 | 1.98 | 0.035 |
| AFP (≥ vs. < 20 ng/mL) | 1.58 | 1.13 | 2.21 | <0.01 | ||||
| Albumin (≥ vs. < 25.75 g/l) | 0.56 | 0.42 | 0.74 | <0.0001 | ||||
| TNM M (M1 vs. M0) | 2.65 | 1.41 | 4.99 | <0.01 | ||||
| Pathologic features | ||||||||
| Liver cirrhosis (yes vs. no) | 0.85 | 0.68 | 1.07 | 0.18 | ||||
| Grading (G3 vs. G1/G2) | 1.50 | 1.15 | 1.95 | <0.01 | ||||
| TNM N (N1 vs. N0) | 1.68 | 0.79 | 3.57 | 0.17 | ||||
| VI (micro vs. none) | 1.47 | 1.12 | 1.93 | <0.01 | 1.17 | 0.87 | 1.57 | 0.309 |
| VI (macro vs. none) | 3.18 | 2.39 | 4.25 | <0.0001 | 2.57 | 1.89 | 3.51 | <0.0001 |
| Macrotrabecular Subtype | 1.96 | 1.34 | 2.85 | <0.001 | 1.43 | 0.94 | 2.20 | 0.098 |
| VETC (pos. vs. neg.) | 1.50 | 1.14 | 1.99 | <0.01 | 1.32 | 0.99 | 1.77 | 0.055 |
| GS (≥9 vs. <9) | 0.77 | 0.58 | 1.02 | 0.07 | ||||
| CAD–CPS1 (high vs. low) | 1.56 | 1.24 | 1.96 | <0.001 | 1.37 | 1.06 | 1.75 | 0.014 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ridder, D.A.; Schindeldecker, M.; Weinmann, A.; Berndt, K.; Urbansky, L.; Witzel, H.R.; Heinrich, S.; Roth, W.; Straub, B.K. Key Enzymes in Pyrimidine Synthesis, CAD and CPS1, Predict Prognosis in Hepatocellular Carcinoma. Cancers 2021, 13, 744. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13040744
Ridder DA, Schindeldecker M, Weinmann A, Berndt K, Urbansky L, Witzel HR, Heinrich S, Roth W, Straub BK. Key Enzymes in Pyrimidine Synthesis, CAD and CPS1, Predict Prognosis in Hepatocellular Carcinoma. Cancers. 2021; 13(4):744. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13040744
Chicago/Turabian StyleRidder, Dirk Andreas, Mario Schindeldecker, Arndt Weinmann, Kristina Berndt, Lana Urbansky, Hagen Roland Witzel, Stefan Heinrich, Wilfried Roth, and Beate Katharina Straub. 2021. "Key Enzymes in Pyrimidine Synthesis, CAD and CPS1, Predict Prognosis in Hepatocellular Carcinoma" Cancers 13, no. 4: 744. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13040744