
Inhibition of Liver Metastasis in Colorectal Cancer by Targeting IL-13/IL13Rα2 Binding Site with Specific Monoclonal Antibodies

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Lines, Reagents and Antibodies
2.2. Peptide Design, Immunization, Preparation and Selection of Anti-IL13Rα2 Mouse Monoclonal Antibodies
2.3. Chicken Antibodies
2.4. Capture ELISA
2.5. Competition ELISAs
2.6. Antibody Confocal Microscopy
2.7. Flow Cytometry
2.8. Western Blot Analysis and Immunoprecipitation
2.9. IL13Rα2 Internalization and Degradation
2.10. Cell Signaling
2.11. Cell Adhesion and Proliferation
2.12. Wound Healing
2.13. Cell Invasion Assays
2.14. Experimental Metastasis in Nude Mice
2.15. Statistical Analyses
3. Results
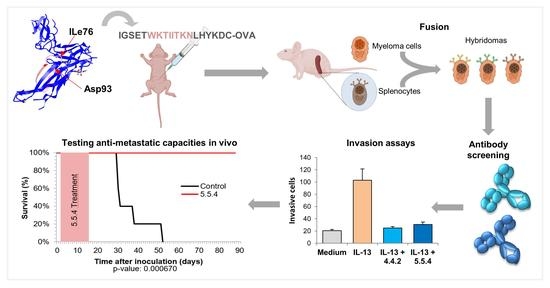
3.1. Antibody Selection with the Capacity for Blocking IL13Rα2-Mediated Invasion
3.2. Characterization of IL13Rα2 D1-Specific mAbs
3.3. IL13Rα2 D1-Specific mAbs Inhibit Metastatic Properties in Colorectal Cancer Cells
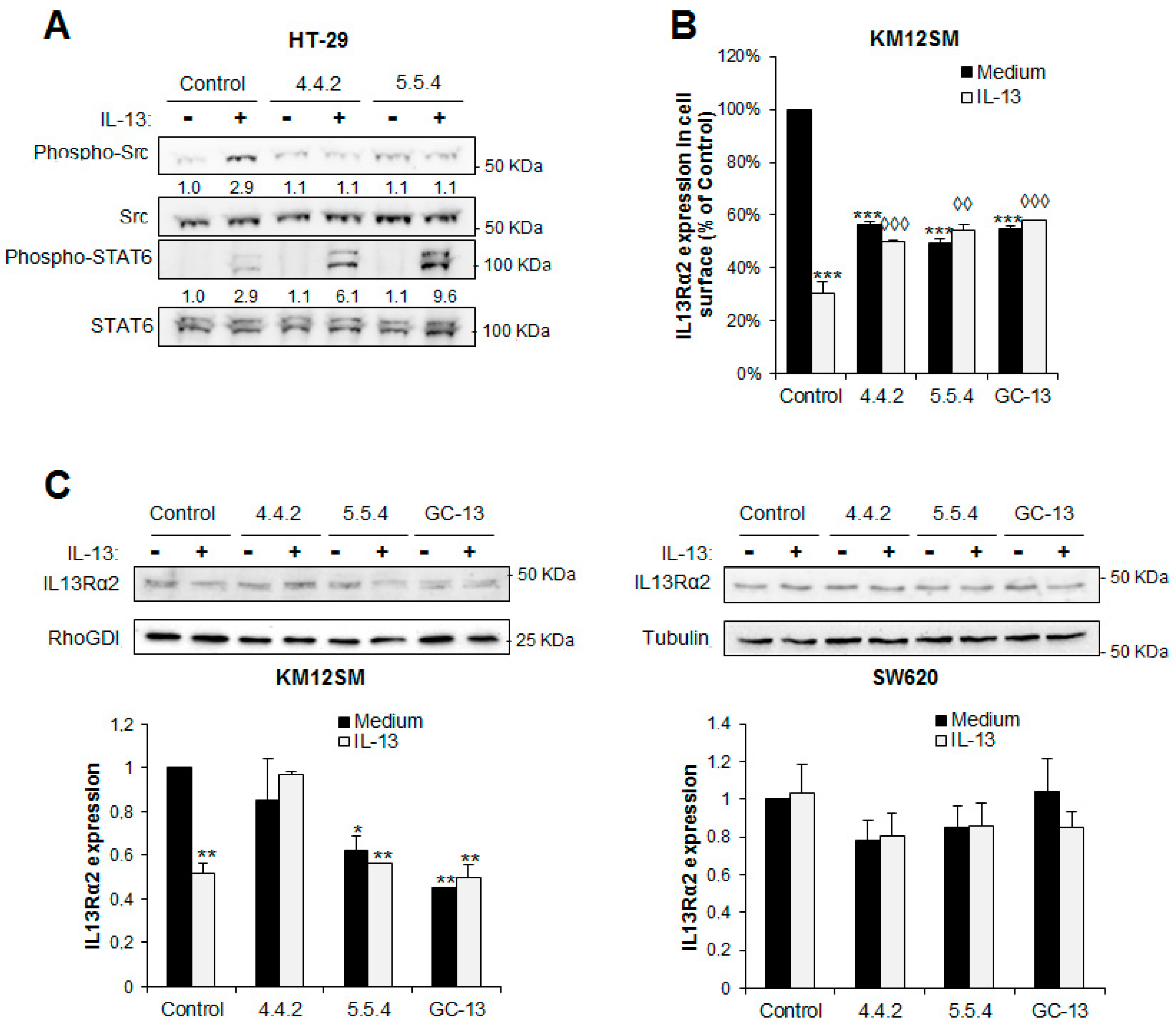
3.4. IL13Rα2 D1-Specific mAbs Inhibit Ligand-Induced Phosphorylation of Downstream Signaling Molecules
3.5. IL13Rα2 D1 mAbs Inhibit IL13Rα2 Activation But Not IL13Rα1 Signaling
3.6. IL13Rα2 D1 mAbs Prevent the Internalization Capacity of IL-13 Binding to IL13Rα2
3.7. Monoclonal Antibody 5.5.4 Is a Potent Inhibitor of Colorectal Cancer Metastasis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holmes, D. A disease of growth. Nat. Cell Biol. 2015, 521, S2–S3. [Google Scholar] [CrossRef] [PubMed]
- Piawah, S.; Venook, A.P. Targeted therapy for colorectal cancer metastases: A review of current methods of molecularly targeted therapy and the use of tumor biomarkers in the treatment of metastatic colorectal cancer. Cancer 2019, 125, 4139–4147. [Google Scholar] [CrossRef] [PubMed]
- Van Cutsem, E.; Köhne, C.-H.; Láng, I.; Folprecht, G.; Nowacki, M.P.; Cascinu, S.; Shchepotin, I.; Maurel, J.; Cunningham, D.; Tejpar, S.; et al. Cetuximab Plus Irinotecan, Fluorouracil, and Leucovorin As First-Line Treatment for Metastatic Colorectal Cancer: Updated Analysis of Overall Survival According to Tumor KRAS and BRAF Mutation Status. J. Clin. Oncol. 2011, 29, 2011–2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciardiello, F.; Tortora, G. EGFR Antagonists in Cancer Treatment. N. Engl. J. Med. 2008, 358, 1160–1174. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Wang, L.; Qiu, H.; Zhang, M.; Sun, L.; Peng, P.; Yu, Q.; Yuan, X. Mechanisms of resistance to anti-EGFR therapy in colorectal cancer. Oncotarget 2017, 8, 3980–4000. [Google Scholar] [CrossRef] [Green Version]
- Martini, G.; Troiani, T.; Cardone, C.; Vitiello, P.; Sforza, V.; Ciardiello, D.; Napolitano, S.; Della Corte, C.M.; Morgillo, F.; Raucci, A.; et al. Present and future of metastatic colorectal cancer treatment: A review of new candidate targets. World J. Gastroenterol. 2017, 23, 4675–4688. [Google Scholar] [CrossRef]
- Debinski, W.; Gibo, D.M. Molecular Expression Analysis of Restrictive Receptor for Interleukin 13, a Brain Tumor-associated Cancer/Testis Antigen. Mol. Med. 2000, 6, 440–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debinski, W.; Gibo, D.M.; Hulet, S.W.; Connor, J.R.; Gillespie, G.Y. Receptor for interleukin 13 is a marker and therapeutic target for human high-grade gliomas. Clin. Cancer Res. 1999, 5, 985–990. [Google Scholar]
- Fujisawa, T.; Joshi, B.H.; Nakajima, A.; Puri, R.K. A Novel Role of Interleukin-13 Receptor 2 in Pancreatic Cancer Invasion and Metastasis. Cancer Res. 2009, 69, 8678–8685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, J.; Treton, D.; Vermot-Desroches, C.; Boden, C.; Horellou, P.; Angevin, E.; Galanaud, P.; Wijdenes, J.; Richard, Y. Expression of interleukin 13 receptor in glioma and renal cell carcinoma: IL13Ralpha2 as a decoy receptor for IL13. Lab. Investig. 2001, 81, 1223–1231. [Google Scholar] [CrossRef] [Green Version]
- Beard, R.E.; Abate-Daga, D.; Rosati, S.F.; Zheng, Z.; Wunderlich, J.R.; Rosenberg, S.A.; Morgan, R.A. Gene Expression Profiling using Nanostring Digital RNA Counting to Identify Potential Target Antigens for Melanoma Immunotherapy. Clin. Cancer Res. 2013, 19, 4941–4950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takenouchi, M.; Hirai, S.; Sakuragi, N.; Yagita, H.; Hamada, H.; Kato, K. Epigenetic modulation enhances the therapeutic effect of anti-IL-13R(alpha)2 antibody in human mesothelioma xenografts. Clin. Cancer Res. 2011, 17, 2819–2829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawakami, M.; Kawakami, K.; Kasperbauer, J.L.; Hinkley, L.L.; Tsukuda, M.; E Strome, S.; Puri, R.K. Interleukin-13 receptor alpha2 chain in human head and neck cancer serves as a unique diagnostic marker. Clin. Cancer Res. 2003, 9, 6381–6388. [Google Scholar]
- Kioi, M.; Kawakami, M.; Shimamura, T.; Husain, S.R.; Puri, R.K. Interleukin-13 receptor alpha2 chain: A potential biomarker and molecular target for ovarian cancer therapy. Cancer 2006, 107, 1407–1418. [Google Scholar] [CrossRef]
- Hallett, M.A.; Venmar, K.T.; Fingleton, B. Cytokine Stimulation of Epithelial Cancer Cells: The Similar and Divergent Functions of IL-4 and IL-13. Cancer Res. 2012, 72, 6338–6343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barderas, R.; Bartolomé, R.Á.; Fernandez-Aceñero, M.J.; Torres, S.; Casal, J.I. High Expression of IL-13 Receptor α2 in Colorectal Cancer Is Associated with Invasion, Liver Metastasis, and Poor Prognosis. Cancer Res. 2012, 72, 2780–2790. [Google Scholar] [CrossRef] [Green Version]
- Fichtner-Feigl, S.; Strober, W.; Kawakami, K.; Puri, R.K.; Kitani, A. IL-13 signaling through the IL-13alpha2 receptor is involved in induction of TGF-beta1 production and fibrosis. Nat. Med. 2006, 12, 99–106. [Google Scholar] [CrossRef]
- Fujisawa, T.; Joshi, B.H.; Puri, R.K. IL-13 regulates cancer invasion and metastasis through IL-13Ralpha2 via ERK/AP-1 pathway in mouse model of human ovarian cancer. Int. J. Cancer 2012, 131, 344–356. [Google Scholar] [CrossRef]
- Bartolome, R.A.; Martin-Regalado, A.; Jaen, M.; Zannikou, M.; Zhang, P.; de Los Rios, V.; Balyasnikova, I.V.; Casal, J.I. Protein Tyrosine Phosphatase-1B Inhibition Disrupts IL13Ralpha2-Promoted Invasion and Metastasis in Cancer Cells. Cancers 2020, 12, 500. [Google Scholar] [CrossRef] [Green Version]
- Bartolomé, R.Á.; García-Palmero, I.; Torres, S.; López-Lucendo, M.F.; Balyasnikova, I.V.; Casal, J.I. IL13 Receptor α2 Signaling Requires a Scaffold Protein, FAM120A, to Activate the FAK and PI3K Pathways in Colon Cancer Metastasis. Cancer Res. 2015, 75, 2434–2444. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, R.; Suzuki, A.; Leland, P.; Joshi, B.H.; Puri, R.K. Identification of a novel role of IL-13Ralpha2 in human Glioblastoma multiforme: Interleukin-13 mediates signal transduction through AP-1 pathway. J Transl. Med. 2018, 16, 369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaci, B.; Brown, C.E.; Binello, E.; Werbaneth, K.; Sampath, P.; Sengupta, S. Significance of interleukin-13 receptor alpha 2-targeted glioblastoma therapy. Neuro-Oncology 2014, 16, 1304–1312. [Google Scholar] [CrossRef] [Green Version]
- Jarboe, J.S.; Johnson, K.R.; Choi, Y.; Lonser, R.R.; Park, J.K. Expression of Interleukin-13 Receptor α2 in Glioblastoma Multiforme: Implications for Targeted Therapies. Cancer Res. 2007, 67, 7983–7986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaeger, R.; Chatila, W.K.; Lipsyc, M.D.; Hechtman, J.F.; Cercek, A.; Sanchez-Vega, F.; Jayakumaran, G.; Middha, S.; Zehir, A.; Donoghue, M.T.A.; et al. Clinical Sequencing Defines the Genomic Landscape of Metastatic Colorectal Cancer. Cancer Cell 2018, 33, 125–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupardus, P.J.; Birnbaum, M.E.; Garcia, K.C. Molecular basis for shared cytokine recognition revealed in the structure of an unusually high affinity complex between IL-13 and IL-13Ralpha2. Structure 2010, 18, 332–342. [Google Scholar] [CrossRef] [Green Version]
- Bartolome, R.A.; Jaen, M.; Casal, J.I. An IL13Ralpha2 peptide exhibits therapeutic activity against metastatic colorectal cancer. Br. J. Cancer 2018, 119, 940–949. [Google Scholar] [CrossRef] [Green Version]
- Kioi, M.; Seetharam, S.; Puri, R.K. Targeting IL-13Ralpha2-positive cancer with a novel recombinant immunotoxin composed of a single-chain antibody and mutated Pseudomonas exotoxin. Mol. Cancer Ther. 2008, 7, 1579–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balyasnikova, I.V.; Wainwright, D.A.; Solomaha, E.; Lee, G.; Han, Y.; Thaci, B.; Lesniak, M.S. Characterization and Immunotherapeutic Implications for a Novel Antibody Targeting Interleukin (IL)-13 Receptor α2. J. Biol. Chem. 2012, 287, 30215–30227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartolomé, R.A.; Aizpurua, C.; Jaén, M.; Torres, S.; Calviño, E.; Imbaud, J.I.; Casal, J.I. Monoclonal Antibodies Directed against Cadherin RGD Exhibit Therapeutic Activity against Melanoma and Colorectal Cancer Metastasis. Clin. Cancer Res. 2017, 24, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Castle, J.C.; Loewer, M.; Boegel, S.; De Graaf, J.; Bender, C.; Tadmor, A.D.; Boisguerin, V.; Bukur, T.; Sorn, P.; Paret, C.; et al. Immunomic, genomic and transcriptomic characterization of CT26 colorectal carcinoma. BMC Genom. 2014, 15, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Berg, K.C.G.; Eide, P.W.; Eilertsen, I.A.; Johannessen, B.; Bruun, J.; Danielsen, S.A.; Bjørnslett, M.; Meza-Zepeda, L.A.; Eknæs, M.; Lind, G.E.; et al. Multi-omics of 34 colorectal cancer cell lines—A resource for biomedical studies. Mol. Cancer 2017, 16, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Adams, G.P.; Schier, R.; McCall, A.M.; Simmons, H.H.; Horak, E.M.; Alpaugh, R.K.; Marks, J.D.; Weiner, L.M. High affinity restricts the localization and tumor penetration of single-chain fv antibody molecules. Cancer Res. 2001, 61, 4750–4755. [Google Scholar]
- Rudnick, S.I.; Lou, J.; Shaller, C.C.; Tang, Y.; Klein-Szanto, A.J.; Weiner, L.M.; Marks, J.D.; Adams, G.P. Influence of Affinity and Antigen Internalization on the Uptake and Penetration of Anti-HER2 Antibodies in Solid Tumors. Cancer Res. 2011, 71, 2250–2259. [Google Scholar] [CrossRef] [Green Version]
- Barderas, R.; Desmet, J.; Timmerman, P.; Meloen, R.; Casal, J.I. Affinity maturation of antibodies assisted by in silico modeling. Proc. Natl. Acad. Sci. USA 2008, 105, 9029–9034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caballero, O.L.; Chen, Y.-T. Cancer/Testis Antigens: Potential Targets for Immunotherapy. In Innate Immune Regulation and Cancer Immunotherapy; Metzler, J.B., Ed.; Springer: New York, NY, USA, 2011; pp. 347–369. [Google Scholar]
- Vieira, A.V.; Lamaze, C.; Schmid, S.L. Control of EGF Receptor Signaling by Clathrin-Mediated Endocytosis. Science 1996, 274, 2086–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaelsen, T.E.; Aase, A.; Norderhaug, L.; Sandlie, I. Antibody dependent cell-mediated cytotoxicity induced by chimeric mouse-human IgG subclasses and IgG3 antibodies with altered hinge region. Mol. Immunol. 1992, 29, 319–326. [Google Scholar] [CrossRef]
- Scott, A.M.; Wolchok, J.D.; Old, L.J. Antibody therapy of cancer. Nat. Rev. Cancer 2012, 12, 278–287. [Google Scholar] [CrossRef]
- O’Toole, M.; Legault, H.; Ramsey, R.; Wynn, T.A.; Kasaian, M.T. A novel and sensitive ELISA reveals that the soluble form of IL-13R-α2 is not expressed in plasma of healthy or asthmatic subjects. Clin. Exp. Allergy 2008, 38, 594–601. [Google Scholar] [CrossRef] [Green Version]
- Lal, A.; Glazer, C.A.; Martinson, H.M.; Friedman, H.S.; Archer, G.E.; Sampson, J.H.; Riggins, G.J. Mutant epidermal growth factor receptor up-regulates molecular effectors of tumor invasion. Cancer Res. 2002, 62, 3335–3339. [Google Scholar]
- Banerjee, S.; Flores-Rozas, H. Monoclonal antibodies for targeted therapy in colorectal cancer. Cancer Biol. Ther. 2010, 9, 563–571. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaén, M.; Bartolomé, R.A.; Aizpurua, C.; Martin-Regalado, Á.; Imbaud, J.I.; Casal, J.I. Inhibition of Liver Metastasis in Colorectal Cancer by Targeting IL-13/IL13Rα2 Binding Site with Specific Monoclonal Antibodies. Cancers 2021, 13, 1731. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13071731
Jaén M, Bartolomé RA, Aizpurua C, Martin-Regalado Á, Imbaud JI, Casal JI. Inhibition of Liver Metastasis in Colorectal Cancer by Targeting IL-13/IL13Rα2 Binding Site with Specific Monoclonal Antibodies. Cancers. 2021; 13(7):1731. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13071731
Chicago/Turabian StyleJaén, Marta, Rubén A. Bartolomé, Carmen Aizpurua, Ángela Martin-Regalado, J. Ignacio Imbaud, and J. Ignacio Casal. 2021. "Inhibition of Liver Metastasis in Colorectal Cancer by Targeting IL-13/IL13Rα2 Binding Site with Specific Monoclonal Antibodies" Cancers 13, no. 7: 1731. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13071731