
Angiopoietin-1 Upregulates Cancer Cell Motility in Colorectal Cancer Liver Metastases through Actin-Related Protein 2/3

 , , and
, , and 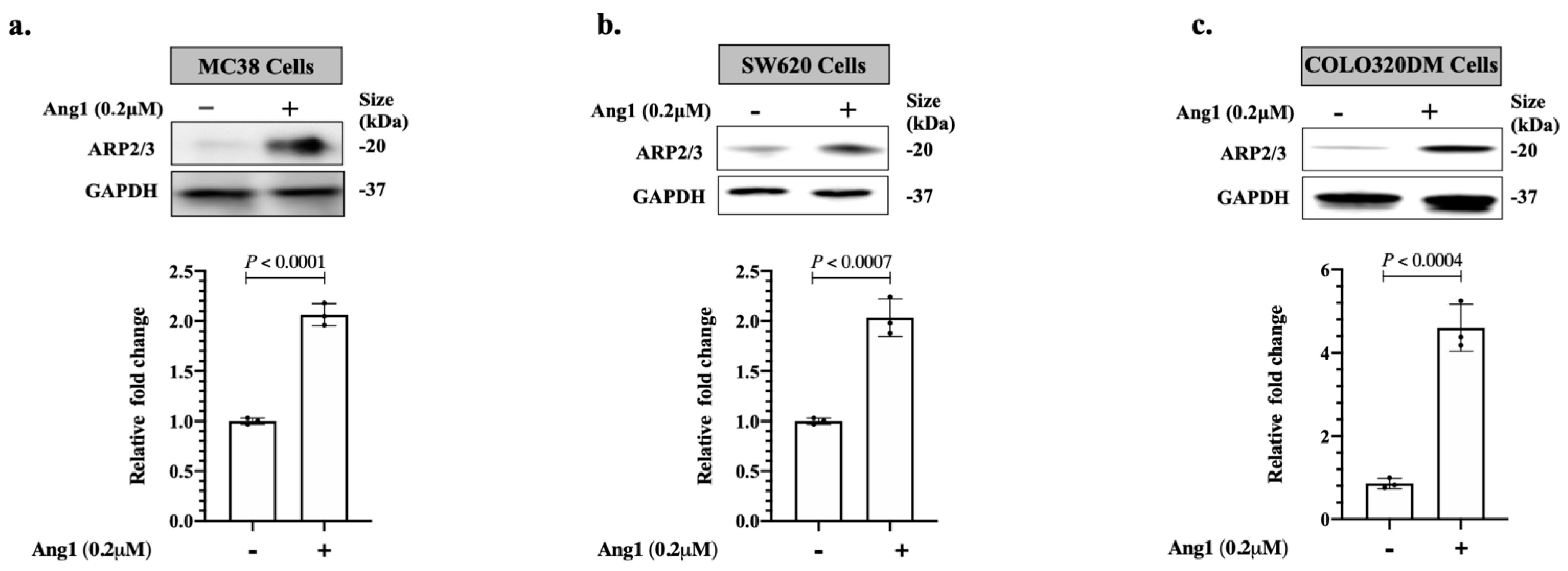{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Samples
2.2. Cell Cultures
2.3. Lentiviral ShRNA Knockdown
2.4. Immunoblotting
2.5. Immunohistochemical Staining
2.6. Immunofluorescence Staining
2.7. Scratch Assay
2.8. Statistical Analysis
3. Results
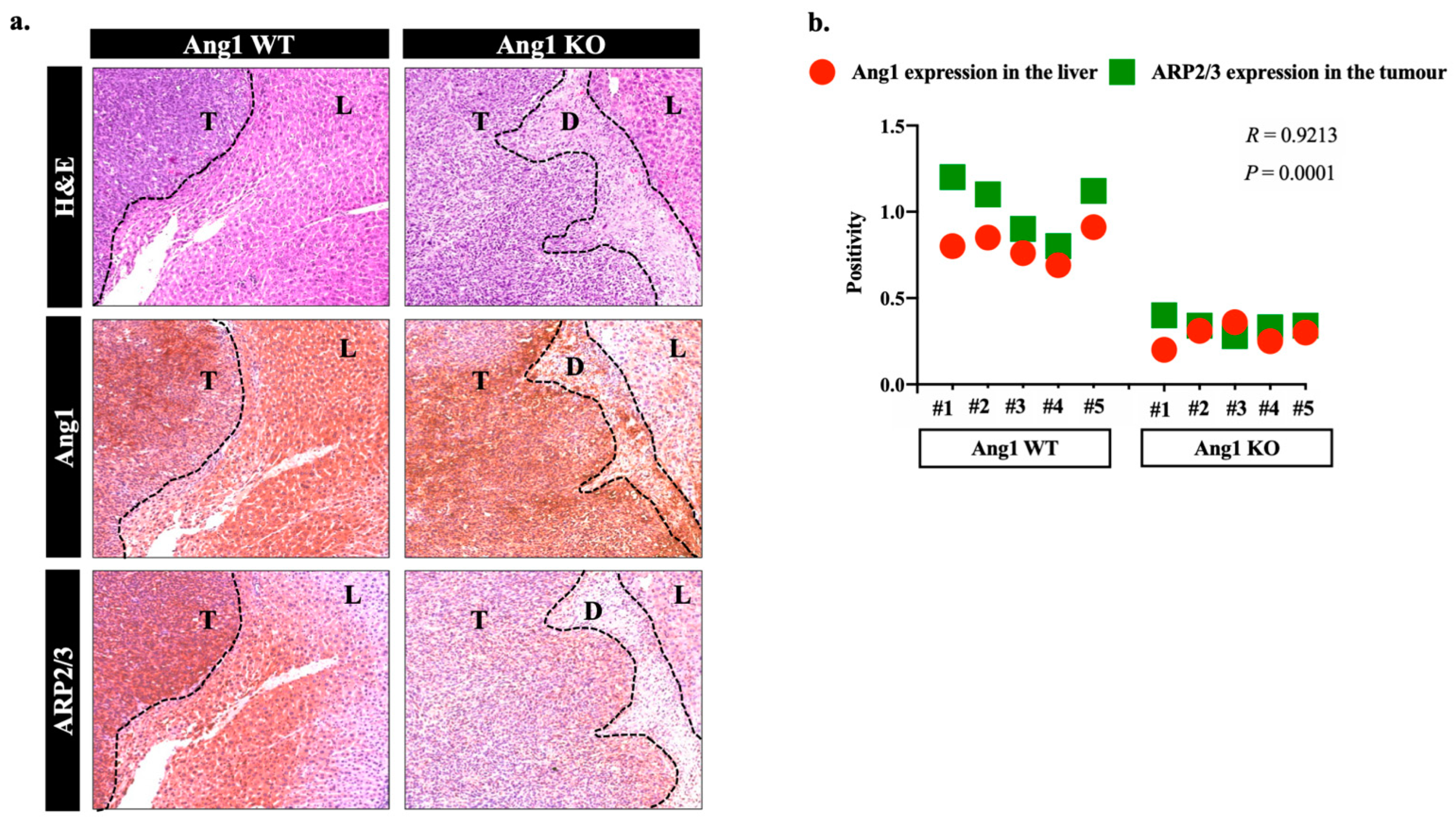
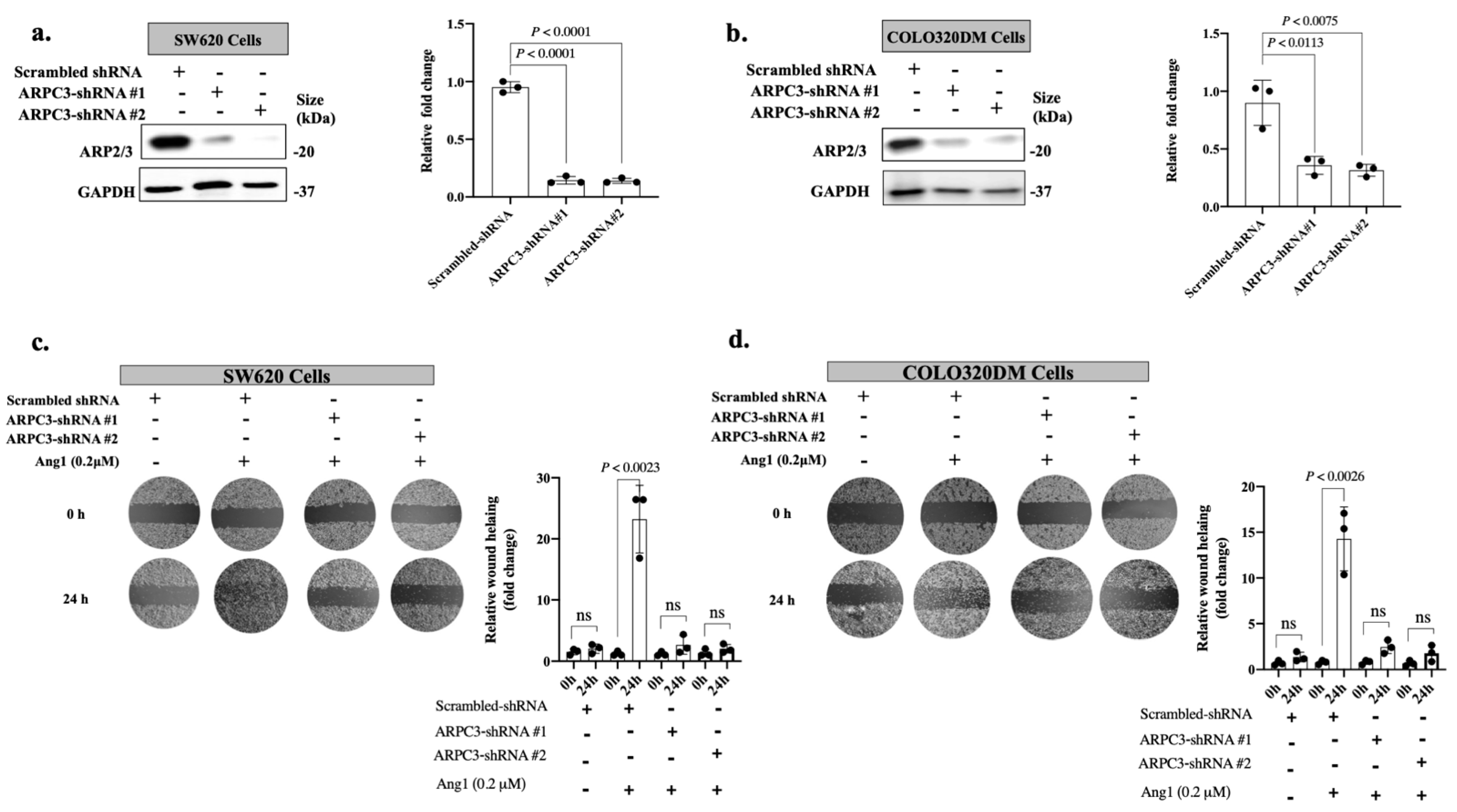
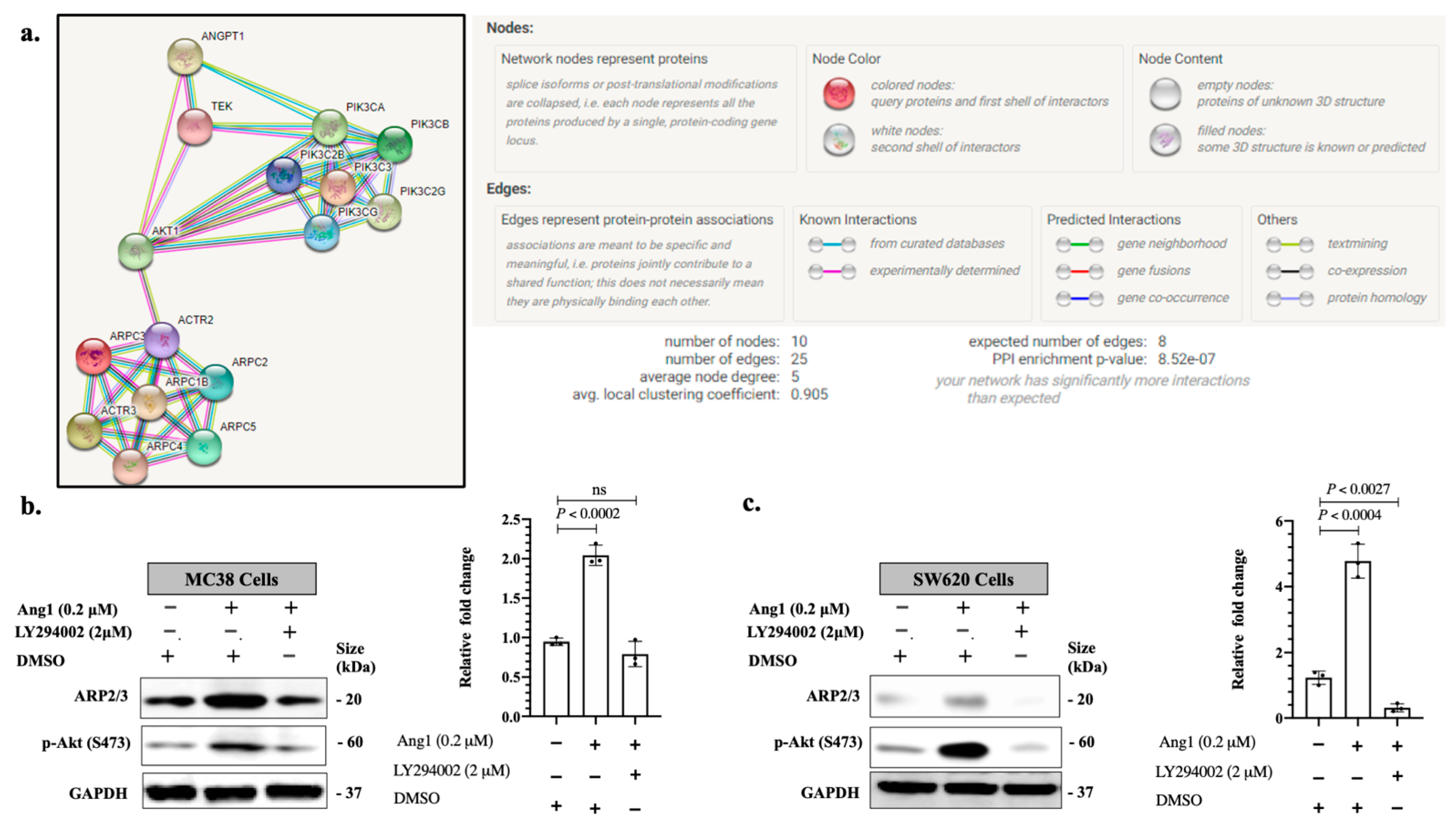
3.1. Ang1 Induces Cancer Cell Motility through ARP2/3
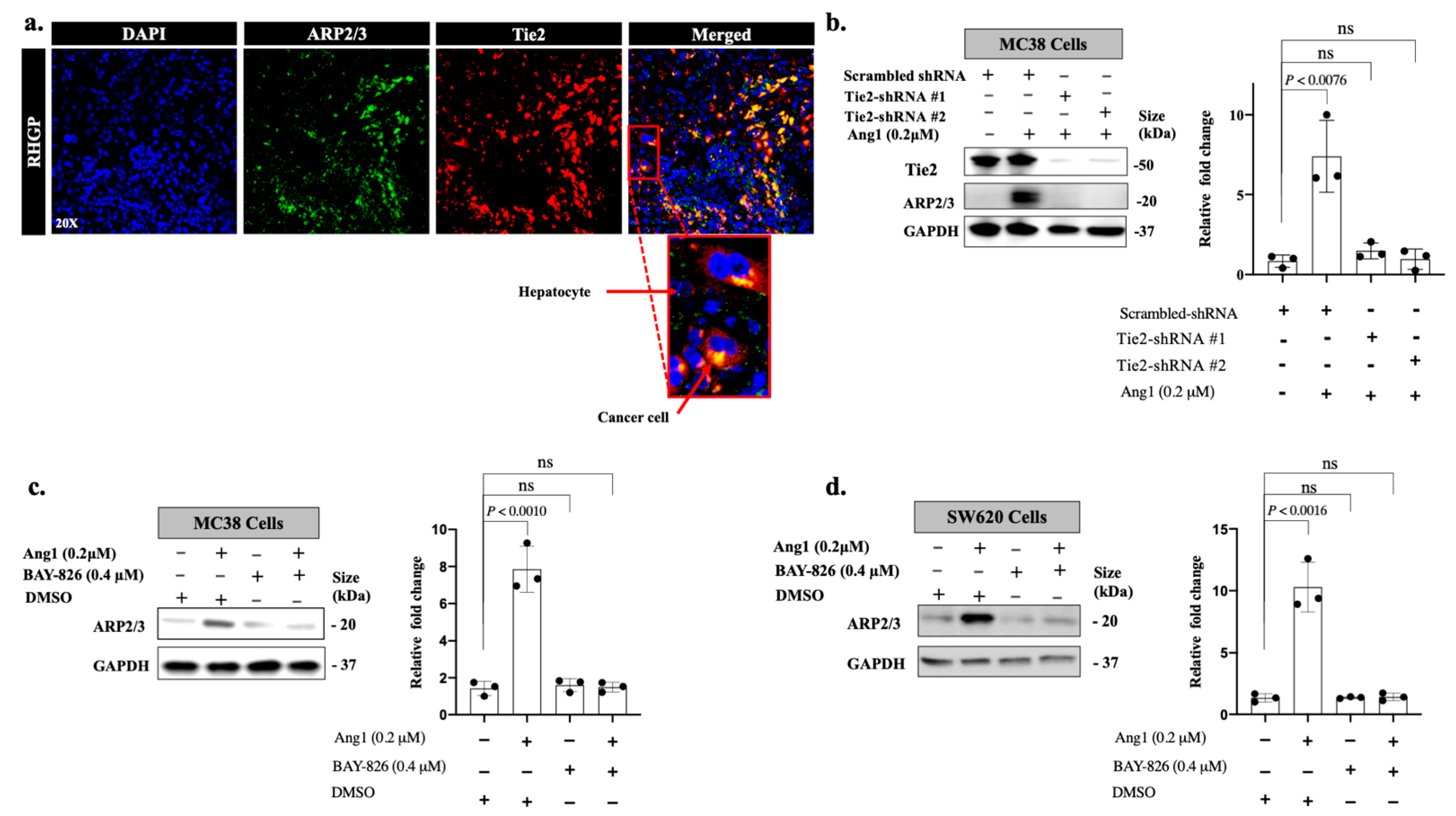
3.2. Ang1 Regulates ARP2/3 Expression through Tie2-PI3K/AKT Pathway
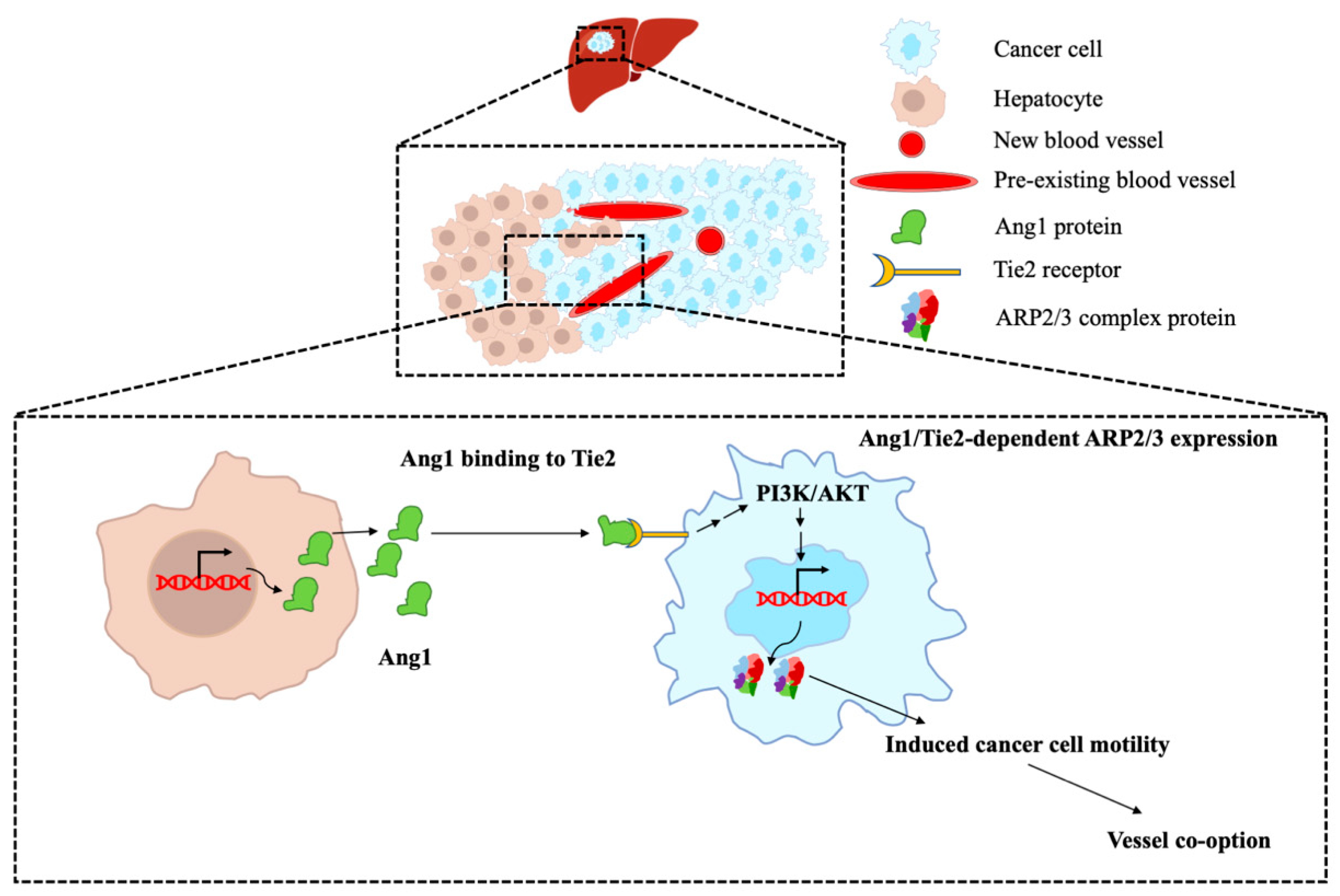
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chow, F.C.; Chok, K.S. Colorectal Liver Metastases: An Update on Multidisciplinary Approach. World J. Hepatol. 2019, 11, 150–172. [Google Scholar] [CrossRef] [PubMed]
- Zarour, L.R.; Anand, S.; Billingsley, K.G.; Bisson, W.H.; Cercek, A.; Clarke, M.F.; Coussens, L.M.; Gast, C.E.; Geltzeiler, C.B.; Hansen, L.; et al. Colorectal Cancer Liver Metastasis: Evolving Paradigms And. Cell. Mol. Gastroenterol. Hepatol. 2017, 3, 163–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomlinson, J.S.; Jarnagin, W.R.; Dematteo, R.P.; Fong, Y.; Kornprat, P.; Gonen, M.; Kemeny, N.; Brennan, M.F.; Blumgart, L.H.; Angelica, M.D. Actual 10-Year Survival After Resection of Colorectal Liver Metastases Defines Cure. J. Clin. Oncol. 2007, 25, 4575–4580. [Google Scholar] [CrossRef] [PubMed]
- Very, N.; Lefebvre, T.; Yazidi-belkoura, I.E. Drug Resistance Related to Aberrant Glycosylation in Colorectal Cancer. Oncotarget 2018, 9, 1380–1402. [Google Scholar] [CrossRef] [Green Version]
- Fusai, G.; Davidson, B.R. Strategies to Increase the Resectability of Liver Metastases from Colorectal Cancer. Dig. Surg. 2003, 20, 481–496. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fedewa, S.A.; Ahnen, D.J.; Meester, R.G.S.; Barzi, A.; Jemal, A. Colorectal Cancer Statistics, 2017. CA Cancer J. Clin. 2017, 67, 177–193. [Google Scholar] [CrossRef]
- Rocha, F.G.; Helton, W.S. Resectability of Colorectal Liver Metastases: An Evolving Definition. HPB 2012, 14, 283–284. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, V.; Lazaris, A.; Mayer, T.Z.; Petrillo, S.K.; Alamri, H.; Rada, M.; Jarrouj, G.; Park, W.; Gao, Z.; McDonald, P.P.; et al. Neutrophils Expressing Lysyl Oxidase-like 4 Protein Are Present in Colorectal Cancer Liver Metastases Resistant to Anti-angiogenic Therapy. J. Pathol. 2020, 251, 213–223. [Google Scholar] [CrossRef]
- Frentzas, S.; Simoneau, E.; Bridgeman, V.L.; Vermeulen, P.B.; Foo, S.; Kostaras, E.; Nathan, M.; Wotherspoon, A.; Gao, Z.; Shi, Y.; et al. Vessel co-option mediates resistance to anti-angiogenic therapy in liver metastases. Nat. Med. 2016, 22, 1294–1302. [Google Scholar] [CrossRef] [Green Version]
- Rada, M.; Kapelanski-Lamoureux, A.; Petrillo, S.; Tabariès, S.; Siegel, P.; Reynolds, A.R.; Lazaris, A.; Metrakos, P. Runt Related Transcription Factor-1 Plays a Central Role in Vessel Co-Option of Colorectal Cancer Liver Metastases. Commun. Biol. 2021, 4, 950. [Google Scholar] [CrossRef]
- Rada, M.; Lazaris, A.; Kapelanski-Lamoureux, A.; Mayer, T.Z.; Metrakos, P. Tumor Microenvironment Conditions That Favor Vessel Co-Option in Colorectal Cancer Liver Metastases: A Theoretical Model. Semin. Cancer Biol. 2021, 71, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Lazaris, A.; Amri, A.; Petrillo, S.K.; Zoroquiain, P.; Ibrahim, N.; Salman, A.; Gao, Z.; Vermeulen, P.B.; Metrakos, P. Vascularization of colorectal carcinoma liver metastasis: Insight into stratification of patients for anti-angiogenic therapies. J. Pathol. Clin. Res. 2018, 4, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.; Rolff, H.C.; Eefsen, R.; Vanier, B. The Morphological Growth Patterns of Colorectal Liver Metastases Are Prognostic for Overall Survival. Mod. Pathol. 2014, 27, 1641–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vermeulen, P.B.; Colpaert, C.; Salgado, R.; Royers, R.; Hellemans, H.; Heuvel, E.V.; Goovaerts, G.; Dirix, L.Y.; Marck, E.V. Liver Metastases from Colorectal Adenocarcinomas Grow in Three Patterns with Different Angiogenesis and Desmoplasia. J. Pathol. 2001, 195, 336–342. [Google Scholar] [CrossRef]
- Bridgeman, V.L.; Vermeulen, P.B.; Foo, S.; Bilecz, A.; Daley, F.; Kostaras, E.; Nathan, M.R.; Wan, E.; Frentzas, S.; Schweiger, T.; et al. Vessel Co-Option Is Common in Human Lung Metastases and Mediates Resistance to Anti-Angiogenic Therapy in Preclinical Lung Metastasis Models. J. Pathol. 2017, 241, 362–374. [Google Scholar] [CrossRef]
- Kuczynski, E.A.; Vermeulen, P.B.; Pezzella, F.; Kerbel, R.S.; Reynolds, A.R. Vessel Co-Option in Cancer. Nat. Rev. Clin. Oncol. 2019, 16, 469–493. [Google Scholar] [CrossRef]
- Ibrahim, N.; Lazaris, A.; Rada, M.; Petrillo, S.; Huck, L.; Hussain, S.; Ouladan, S.; Gao, Z.; Gregorieff, A.; Essalmani, R.; et al. Angiopoietin1 Deficiency in Hepatocytes Affects the Growth of Colorectal Cancer Liver. Cancers 2020, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Fukuhara, S.; Sako, K.; Takenouchi, T.; Kitani, H.; Kume, T.; Koh, G.Y.; Mochizuki, N. Angiopoietin-1/Tie2 Signal Augments Basal Notch Signal Controlling Vascular Quiescence by Inducing Delta-Like 4 Expression through AKT-Mediated Activation of beta-Catenin. J. Biol. Chem. 2011, 286, 8055–8066. [Google Scholar] [CrossRef] [Green Version]
- Holopainen, T.; Huang, H.; Chen, C.; Kyung, E.K.; Zhang, L.; Zhou, F.; Han, W.; Li, C.; Yu, J.; Wu, J.; et al. Angiopoietin-1 Overexpression Modulates Vascular Endothelium to Facilitate Tumor Cell Dissemination and Metastasis Establishment. Cancer Res. 2009, 69, 4656–4664. [Google Scholar] [CrossRef] [Green Version]
- Koblizek, T.I.; Weiss, C.; Yancopoulos, G.D.; Deutsch, U.; Risau, W. Angiopoietin-1 Induces Sprouting Angiogenesis in Vitro. Curr. Biol. 1998, 8, 529–532. [Google Scholar] [CrossRef] [Green Version]
- Robinson, R.C.; Turbedsky, K.; Kaiser, D.A.; Marchand, J.; Higgs, H.N.; Choe, S.; Pollard, T.D. Crystal Structure of Arp2/3 Complex. Science 2001, 294, 1679–1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molinie, N.; Gautreau, X.A. The Arp2/3 Regulatory System and Its Deregulation in Cancer. Physiol. Rev. 2018, 98, 215–238. [Google Scholar] [CrossRef] [PubMed]
- Rada, M.; Tsamchoe, M.; Kapelanski-Lamoureux, A.; Bloom, J.; Petrillo, S.K.; Kim, D.H.; Lazaris, A.; Metrakos, P. Cancer Cells Induce Hepatocytes Apoptosis in Co-Opted Colorectal Cancer Liver Metastatic Lesions. bioRxiv 2021, 0211, 1–21. [Google Scholar] [CrossRef]
- Rada, M.; Tsamchoe, M.; Kapelanski-lamoureux, A.; Hassan, N.; Bloom, J.; Petrillo, S.; Kim, D.H.; Lazaris, A.; Metrakos, P. Cancer Cells Promote Phenotypic Alterations in Hepatocytes at the Edge of Cancer Cell Nests to Facilitate Vessel Co-Option Establishment in Colorectal Cancer Liver Metastases. Cancers 2022, 14, 1318. [Google Scholar] [CrossRef]
- Farra, R.; Dapas, B.; Pozzato, G.; Scaggiante, B.; Agostini, F.; Zennaro, C.; Grassi, M.; Rosso, N.; Giansante, C.; Fiotti, N.; et al. Effects of E2F1—Cyclin E1–E2 Circuit down Regulation in Hepatocellular Carcinoma. Dig. Liver Dis. 2011, 43, 1006–1014. [Google Scholar] [CrossRef]
- Zhang, Q.; Hou, X.; Evans, B.J.; Vanblaricom, J.L.; Weroha, S.J.; Cliby, W.A. LY2157299 Monohydrate, a TGF-ΒR1 Inhibitor, Suppresses Tumor Growth and Ascites Development in Ovarian Cancer. Cancers 2018, 10, 260. [Google Scholar] [CrossRef] [Green Version]
- Kitajima, D.; Kasamatsu, A.; Nakashima, D.A.I.; Miyamoto, I.; Kimura, Y. Evidence for Critical Role of Tie2/Ang1 Interaction in Metastatic Oral Cancer. Oncol. Lett. 2018, 15, 7237–7242. [Google Scholar] [CrossRef]
- Cascone, T.; Heymach, J.V.; Cancer, A. Targeting the Angiopoietin / Tie2 Pathway: Cutting Tumor Vessels With a Double-Edged Sword? J. Clin. Oncol. 2012, 30, 441–444. [Google Scholar] [CrossRef]
- Martin, V.; Liu, D.; Fueyo, J.; Gomez-Manzano, C. Tie2: A Journey from Normal Angiogenesis to Cancer and Beyond. Histol. Histopathol. 2008, 23, 773–780. [Google Scholar] [CrossRef]
- Schneider, H.; Szabo, E.; Machado, R.A.; Broggini-Tenzer, A.; Walter, A.; Lobell, M.; Heldmann, D.; Sussmeier, F.; Grunewald, S.; Weller, M. Novel TIE-2 Inhibitor BAY-826 Displays in Vivo Efficacy in Experimental Syngeneic Murine Glioma Models. J. Neurochem. 2017, 140, 170–182. [Google Scholar] [CrossRef]
- Lelievre, E.; Bourbon, P.; Duan, L.; Nussbaum, R.L.; Fong, G. Deficiency in the p110alpha subunit of PI3K results in diminished Tie2 expression and Tie2(-/-)-like vascular defects in mice. Blood 2005, 105, 3935–3939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, K.E.; Li, J.; Li, X.; Chang, S.H.I.; Zhang, Z. Ang1/Tie2 Induces Cell Proliferation and Migration in Human Papillary Thyroid Carcinoma via the PI3K/AKT Pathway. Oncol. Lett. 2018, 15, 1313–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING V11: Protein—Protein Association Networks with Increased Coverage, Supporting Functional Discovery in Genome-Wide Experimental Datasets. Nucleic Acids Res. 2019, 47, 607–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Kuramitsu, Y.; Baron, B.; Kitagawa, T.; Tokuda, K.; Akada, J.; Maehara, S.; Maehara, Y.; Nakamura, K. PI3K Inhibitor LY294002, as Opposed to Wortmannin, Enhances AKT Phosphorylation in Gemcitabine-Resistant Pancreatic Cancer Cells. Int. J. Oncol. 2017, 50, 606–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nallanthighal, S.; Rada, M.; Heiserman, J.P.; Cha, J.; Sage, J.; Zhou, B.; Yang, W.; Hu, Y.; Korgaonkar, C.; Hanos, C.T.; et al. Inhibition of Collagen XI Alpha 1-Induced Fatty Acid Oxidation Triggers Apoptotic Cell Death in Cisplatin-Resistant Ovarian Cancer. Cell Death Dis. 2020, 11, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rada, M.; Nallanthighal, S.; Cha, J.; Ryan, K.; Sage, J.; Eldred, C.; Ullo, M.; Orsulic, S.; Cheon, D.-J. Inhibitor of Apoptosis Proteins (IAPs) Mediate Collagen Type XI Alpha 1-Driven Cisplatin Resistance in Ovarian Cancer. Oncogene 2018, 37, 4809–4820. [Google Scholar] [CrossRef]
- Kuczynski, E.A.; Yin, M.; Bar-Zion, A.; Lee, C.R.; Butz, H.; Man, S.; Daley, F.; Vermeulen, P.B.; Yousef, G.M.; Foster, F.S.; et al. Co-Option of Liver Vessels and Not Sprouting Angiogenesis Drives Acquired Sorafenib Resistance in Hepatocellular Carcinoma. J. Natl. Cancer Inst. 2016, 108, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, J.I.; Shields, D.J.; Barillas, S.G.; Acevedo, L.M.; Murphy, E.; Haung, J.; Scheppeke, L.; Stockmann, C.; Johnson, R.S.; Angle, N.; et al. A Role for VEGF as a Negative Regulator of Pericyte Function and Vessel Maturation. Nature 2008, 456, 809–813. [Google Scholar] [CrossRef]
- Hendrix, M.J.C.; Seftor, E.A.; Hess, A.R.; Seftor, R.E.B. Vasculogenic Mimicry and Tumour-Cell Plasticity: Lessons from Melanoma. Nat. Rev. Cancer 2003, 3, 411–421. [Google Scholar] [CrossRef]
- Gotink, K.; Broxterman, H.; Labots, M.; de Haas, R.; Dekker, H.; Honeywell, R.; Rudek, M.; Beerepoot, L.; Musters, R.; Jansen, G.; et al. Lysosomal Sequestration of Sunitinib: A Novel Mechanism of Drug Resistance. Clin. Cancer Res. 2011, 17, 7337–7346. [Google Scholar] [CrossRef] [Green Version]
- Croci, D.O.; Cerliani, J.P.; Dalotto-Moreno, T.; Méndez-Huergo, S.P.; Mascanfroni, I.D.; Dergan-Dylon, S.; Toscano, M.A.; Caramelo, J.J.; García-Vallejo, J.J.; Ouyang, J.; et al. Glycosylation-Dependent Lectin-Receptor Interactions Preserve Angiogenesis in Anti-VEGF Refractory Tumors. Cell 2014, 156, 744–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donnem, T.; Hu, J.; Ferguson, M.; Adighibe, O.; Snell, C.; Harris, A.L.; Gatter, K.C.; Pezzella, F. Vessel Co-Option in Primary Human Tumors and Metastases: An Obstacle to Effective Anti-Angiogenic Treatment? Cancer Med. 2013, 2, 427–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuczynski, E.A.; Kerbel, R.S. Implications of Vessel Co-Option in Sorafenib-Resistant Hepatocellular Carcinoma. Chin. J. Cancer 2016, 35, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audero, E.; Cascone, I.; Zanon, I.; Previtali, S.C.; Piva, R.; Schiffer, D.; Bussolino, F. Expression of Angiopoietin-1 in Human Glioblastomas Regulates Tumor-Induced Angiogenesis in Vivo and in Vitro Studies. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 536–541. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.; Chen, N.; Yang, D.; Crane, J.; Huang, B.; Dong, R.; Yi, X.; Guo, J.; Cai, J.; Wang, Z. Cervical Cancer Cell-Derived Angiopoietins Promote Tumor Progression. Tumor Biol. 2017, 39, 1010428317711658. [Google Scholar] [CrossRef] [Green Version]
- Duran, C.L.; Borriello, L.; Karagiannis, G.S.; Entenberg, D.; Oktay, M.H.; Condeelis, J.S. Targeting Tie2 in the Tumor Microenvironment: From Angiogenesis to Dissemination. Cancers 2021, 13, 5730. [Google Scholar] [CrossRef]
- Sopo, M.; Sallinen, H.; Hämäläinen, K.; Kivelä, A.; Ylä- Herttuala, S.; Kosma, V.M.; Keski-Nisula, L.; Anttila, M. High Expression of Tie-2 Predicts Poor Prognosis in Primary High Grade Serous Ovarian Cancer. PLoS ONE 2020, 15, e0241484. [Google Scholar] [CrossRef]
- Yang, W.J.; Hao, Y.X.; Yang, X.; Fu, X.L.; Shi, Y.; Yue, H.L.; Yin, P.; Dong, H.L.; Yu, P.W. Overexpression of Tie2 Is Associated with Poor Prognosis in Patients with Gastric Cancer. Oncol. Lett. 2018, 15, 8027–8033. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.B.; Shifat, R.; Johnson, D.G.; Bedford, M.T.; Gabrusiewicz, K.R.; Cortes-Santiago, N.; Luo, X.; Lu, Z.; Ezhilarasan, R.; Sulman, E.P.; et al. TIE2-Mediated Tyrosine Phosphorylation of H4 Regulates DNA Damage Response by Recruiting ABL1. Sci. Adv. 2016, 2, e1501290. [Google Scholar] [CrossRef] [Green Version]
- Al Bandar, M.H.; Kim, N.K. Perspective on Therapeutic Application of Selective Internal Radiotherapy in Colorectal Cancer Liver Metastasis. Transl. Cancer Res. 2016, 5 (Suppl. 4), S772–S777. [Google Scholar] [CrossRef]
- Li, B.; Xu, W.W.; Lam, A.K.Y.; Wang, Y.; Hu, H.F.; Guan, X.Y.; Qin, Y.R.; Saremi, N.; Tsao, S.W.; He, Q.Y.; et al. Significance of PI3K/AKT Signaling Pathway in Metastasis of Esophageal Squamous Cell Carcinoma and Its Potential as a Target for Anti-Metastasis Therapy. Oncotarget 2017, 8, 38755–38766. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Nie, J.; Ma, X.; Wei, Y.; Peng, Y.; Wei, X. Targeting PI3K in Cancer: Mechanisms and Advances in Clinical Trials 06 Biological Sciences 0601 Biochemistry and Cell Biology. Mol. Cancer 2019, 18, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, I.; Kim, H.G.; So, J.N.; Kim, J.H.; Kwak, H.J.; Koh, G.Y. Angiopoietin-1 Regulates Endothelial Cell Survival through the Phosphatidylinositol 3′-Kinase/Akt Signal Transduction Pathway. Circ. Res. 2000, 86, 24–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Liu, N.F. The Role of Ang/Tie Signaling in Lymphangiogenesis. Lymphology 2010, 43, 59–72. [Google Scholar] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rada, M.; Kapelanski-Lamoureux, A.; Tsamchoe, M.; Petrillo, S.; Lazaris, A.; Metrakos, P. Angiopoietin-1 Upregulates Cancer Cell Motility in Colorectal Cancer Liver Metastases through Actin-Related Protein 2/3. Cancers 2022, 14, 2540. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14102540
Rada M, Kapelanski-Lamoureux A, Tsamchoe M, Petrillo S, Lazaris A, Metrakos P. Angiopoietin-1 Upregulates Cancer Cell Motility in Colorectal Cancer Liver Metastases through Actin-Related Protein 2/3. Cancers. 2022; 14(10):2540. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14102540
Chicago/Turabian StyleRada, Miran, Audrey Kapelanski-Lamoureux, Migmar Tsamchoe, Stephanie Petrillo, Anthoula Lazaris, and Peter Metrakos. 2022. "Angiopoietin-1 Upregulates Cancer Cell Motility in Colorectal Cancer Liver Metastases through Actin-Related Protein 2/3" Cancers 14, no. 10: 2540. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14102540