
Enriched Riceberry Bran Oil Exerts Chemopreventive Properties through Anti-Inflammation and Alteration of Gut Microbiota in Carcinogen-Induced Liver and Colon Carcinogenesis in Rats
 , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. RBBO Sample
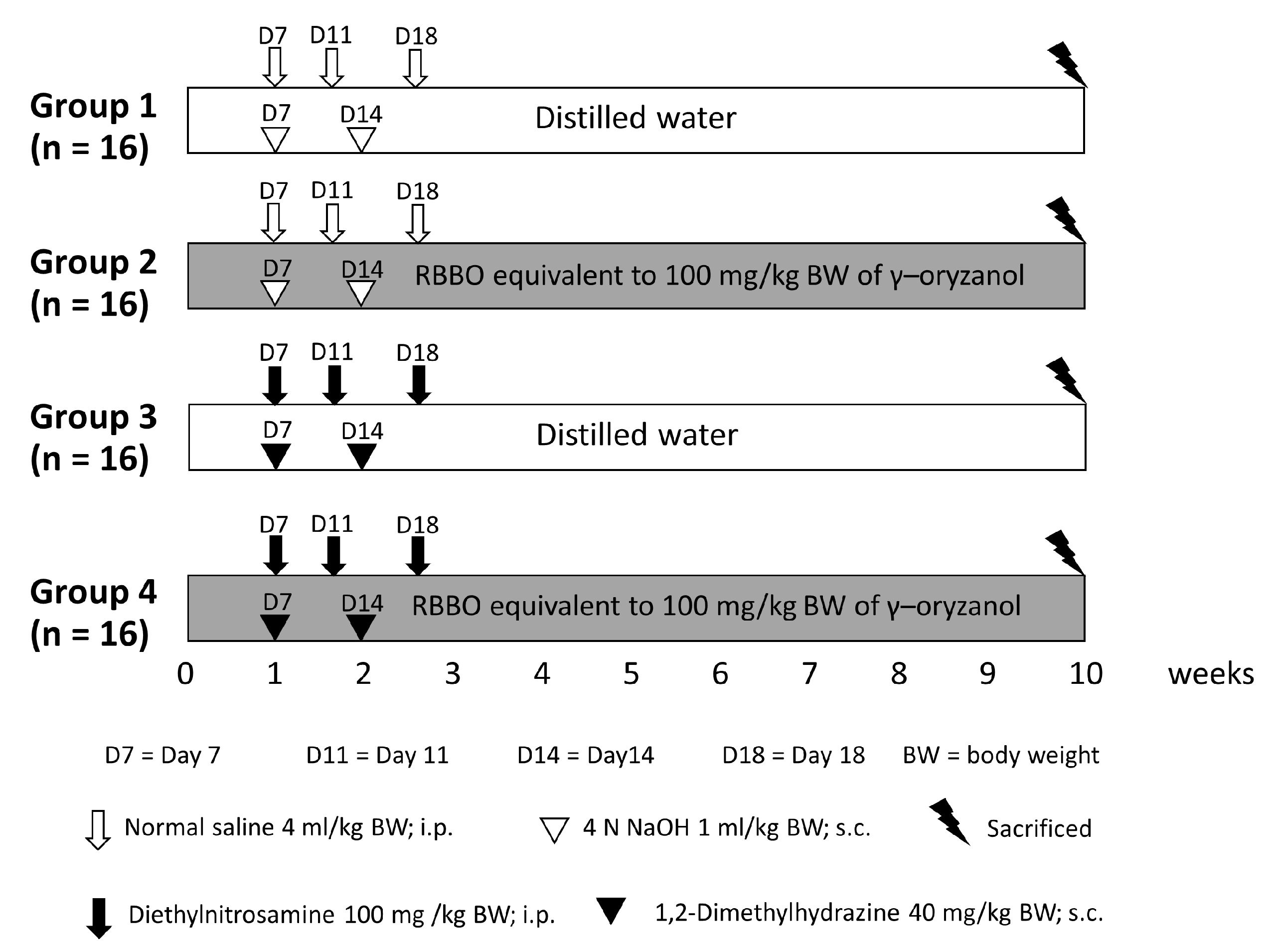
2.3. Animals and Experimental Protocol
2.4. Determination of Preneoplastic Lesions in Colon and Liver Tissues
2.5. Immunohistochemistry of Proliferation Cell Nuclear Antigen (PCNA)
2.6. Terminal Deoxynucleotidyltransferase (TdT)–dUTP Nick End Labeling (TUNEL) Assay
2.7. Determination of Pro-Inflammatory Cytokine Gene Expression by Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.8. Measurement of SCFA in Rat Feces
2.9. Analysis of Composition of Fecal Intestinal Microbiota in Rat
2.10. Statistical Analysis
3. Results
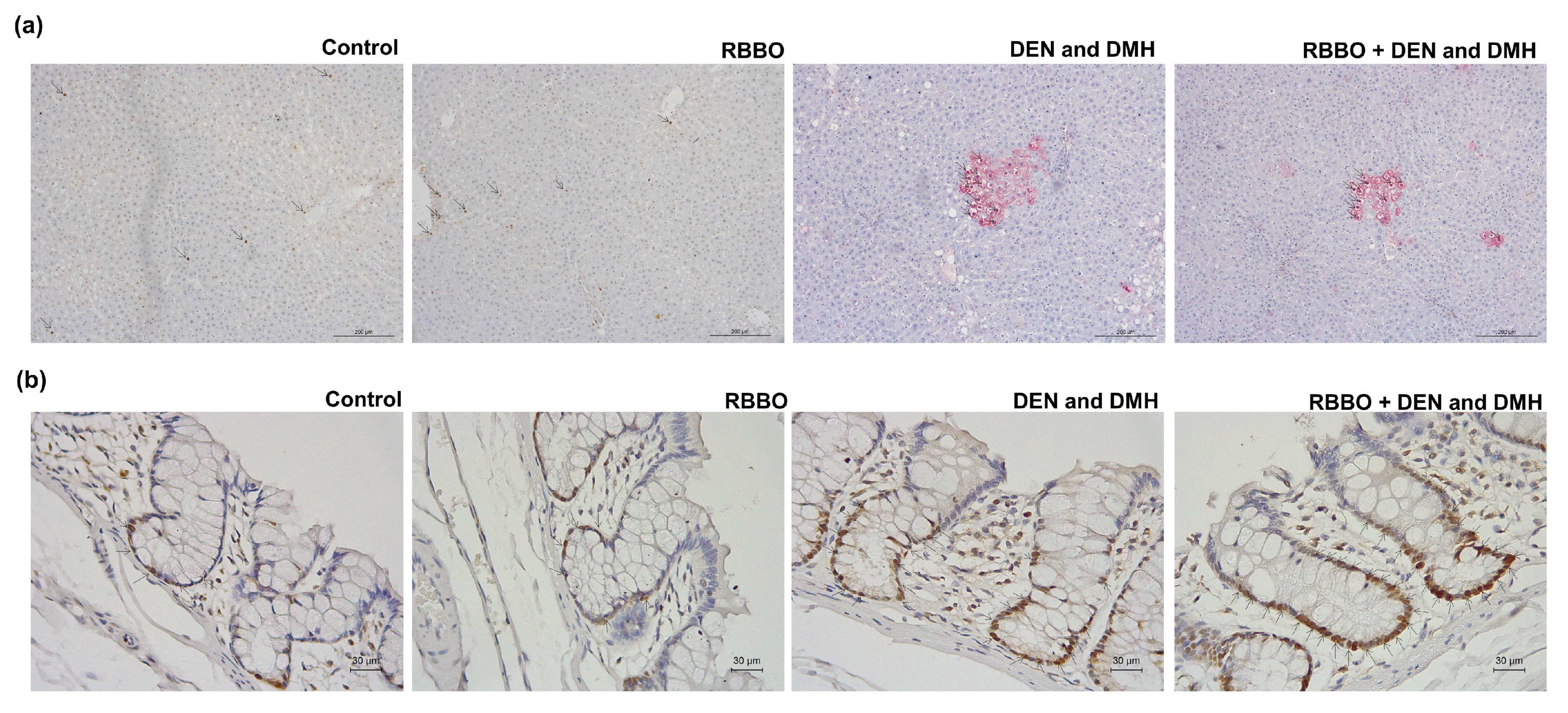
3.1. Effect of RBBO on Preneoplastic Lesions of Liver and Colorectal Carcinogenesis in Rats
3.2. Inhibitory Mechanism of RBBO Involved in Cell Proliferation and Apoptosis in Liver and Colon Tissues of DEN- and DMH-Initiated Rats
3.3. Effect of RBBO on the Expression of Pro-Inflammatory Genes in the Livers and Colons of Rats
3.4. Effect of RBBO on Fecal SCFAs Production in Rats
3.5. Effect of RBBO on Bacterial Profile in Rats
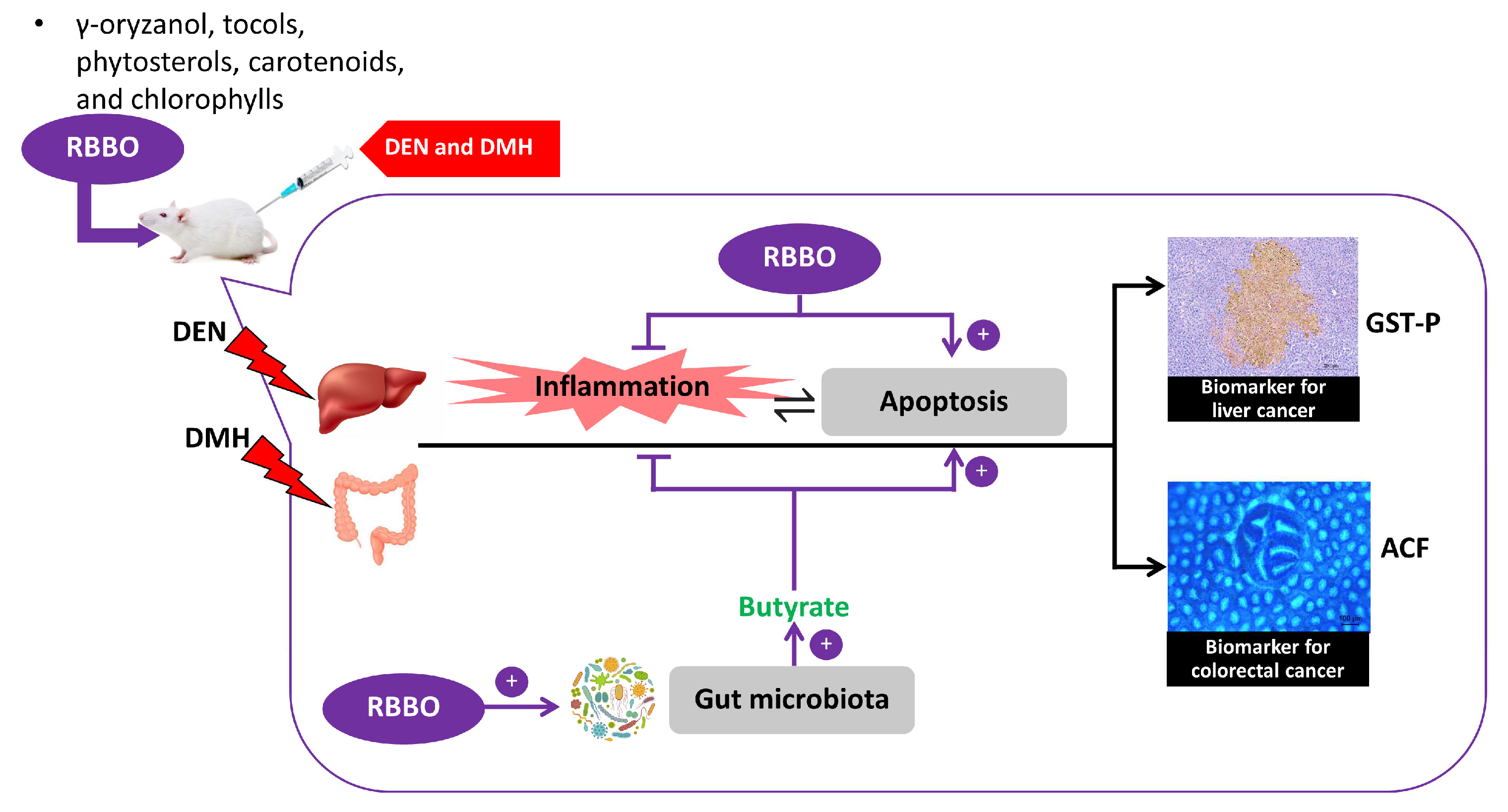
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACF | aberrant crypt foci |
| ALT | alanine aminotransferase |
| AP-1 | activator protein 1 |
| AST | aspartate aminotransferase |
| BW | body weight |
| Ccnd1 | cyclin D1 |
| c-JUN | Jun proto-oncogene |
| COX-2 | cyclooxygenase-2 |
| cDNA | complementary deoxyribonucleic acid |
| DAB | 3,3′-diaminobenzidine |
| DEN | diethylnitrosamine |
| DMH | 1,2-dimethylhydrazine |
| DNA | deoxyribonucleic acid |
| FasL | Fas ligand |
| F/B | Firmicutes/Bacteroidetes |
| FID | flame ionization detector |
| GC | gas chromatography |
| GST-P | glutathione S-transferase placental form |
| H&E | hematoxylin and eosin |
| HFD | High-fat diet |
| IBD | inflammatory bowel disease |
| IL-1β | interleukin-1 beta |
| IL-6 | interleukin-6 |
| iNOS | inducible nitric oxide synthase |
| i.p. | intraperitoneal |
| min | minute |
| mRNA | messenger ribonucleic acid |
| MUFAs | monounsaturated fatty acids |
| NF-κB | nuclear factor kappa B |
| PCNA | proliferation cell nuclear antigen |
| PCR | polymerase chain reaction |
| PUFAs | polyunsaturated fatty acids |
| qRT-PCR | quantitative reverse transcription polymerase chain reaction |
| RBBO | Riceberry bran oil |
| RNA | ribonucleic acid |
| s.c. | subcutaneous |
| SCFA | short-chain fatty acids |
| sec | second |
| TNF-α | tumor necrosis factor-alpha |
| TUNEL | terminal deoxynucleotidyltransferase–deoxyuridine triphosphate nick end labeling |
References
- Oliveira, P.A.; Colaço, A.; Chaves, R.; Guedes-Pinto, H.; De-La-Cruz, P.L.F.; Lopes, C. Chemical carcinogenesis. An. Acad. Bras. Cienc. 2007, 79, 593–616. [Google Scholar] [CrossRef]
- Nasri, H.; Baradaran, A.; Shirzad, H.; Rafieian-Kopaei, M. New concepts in nutraceuticals as alternative for pharmaceuticals. Int. J. Prev. Med. 2014, 5, 1487–1499. [Google Scholar]
- Futakuchi, M.; Cheng, J.L.; Hirose, M.; Kimoto, N.; Cho, Y.M.; Iwata, T.; Kasai, M.; Tokudome, S.; Shirai, T. Inhibition of conjugated fatty acids derived from safflower or perilla oil of induction and development of mammary tumors in rats induced by 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP). Cancer Lett. 2002, 178, 131–139. [Google Scholar] [CrossRef]
- Vafa, M.; Haghighat, N.; Moslehi, N.; Eghtesadi, S.; Heydari, I. Effect of tocotrienols enriched canola oil on glycemic control and oxidative status in patients with type 2 diabetes mellitus: A randomized double-blind placebo-controlled clinical trial. J. Res. Med. Sci. 2015, 20, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Silva Figueiredo, P.; Carla Inada, A.; Marcelino, G.; Maiara Lopes Cardozo, C.; de Cássia Freitas, K.; de Cássia Avellaneda Guimarães, R.; Pereira de Castro, A.; Aragão do Nascimento, V.; Aiko Hiane, P. Fatty acids consumption: The role metabolic aspects involved in obesity and its associated disorders. Nutrients 2017, 9, 1158. [Google Scholar] [CrossRef]
- Sellem, L.; Srour, B.; Guéraud, F.; Pierre, F.; Kesse-Guyot, E.; Fiolet, T.; Lavalette, C.; Egnell, M.; Latino-Martel, P.; Fassier, P.; et al. Saturated, mono- and polyunsaturated fatty acid intake and cancer risk: Results from the French prospective cohort NutriNet-Santé. Eur. J. Nutr. 2019, 58, 1515–1527. [Google Scholar] [CrossRef]
- Guasch-Ferré, M.; Zong, G.; Willett, W.C.; Zock, P.L.; Wanders, A.J.; Hu, F.B.; Sun, Q. Associations of monounsaturated fatty acids from plant and animal sources with total and cause-specific mortality in two US prospective cohort studies. Circ. Res. 2019, 124, 1266–1275. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef]
- Romagnolo, D.F.; Donovan, M.G.; Doetschman, T.C.; Selmin, O.I. n-6 Linoleic acid induces epigenetics alterations associated with colonic inflammation and cancer. Nutrients 2019, 11, 171. [Google Scholar] [CrossRef]
- Wang, W.; Yang, J.; Nimiya, Y.; Lee, K.S.S.; Sanidad, K.; Qi, W.; Sukamtoh, E.; Park, Y.; Liu, Z.; Zhang, G. ω-3 Polyunsaturated fatty acids and their cytochrome P450-derived metabolites suppress colorectal tumor development in mice. J. Nutr. Biochem. 2017, 48, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef]
- Qiao, Y.; Sun, J.; Ding, Y.; Le, G.; Shi, Y. Alterations of the gut microbiota in high-fat diet mice is strongly linked to oxidative stress. Appl. Microbiol. Biotechnol. 2013, 97, 1689–1697. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wei, H.; Zhou, Y.; Szeto, C.H.; Li, C.; Lin, Y.; Coker, O.O.; Lau, H.C.H.; Chan, A.W.H.; Sung, J.J.Y.; et al. High-fat diet promotes colorectal tumorigenesis through modulating gut microbiota and metabolites. Gastroenterology 2022, 162, 135–149.e132. [Google Scholar] [CrossRef]
- Zhang, X.; Coker, O.O.; Chu, E.S.; Fu, K.; Lau, H.C.H.; Wang, Y.-X.; Chan, A.W.H.; Wei, H.; Yang, X.; Sung, J.J.Y.; et al. Dietary cholesterol drives fatty liver-associated liver cancer by modulating gut microbiota and metabolites. Gut 2021, 70, 761–774. [Google Scholar] [CrossRef]
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 577–589. [Google Scholar] [CrossRef]
- Wongwaiwech, D.; Weerawatanakorn, M.; Tharatha, S.; Ho, C.T. Comparative study on amount of nutraceuticals in by-products from solvent and cold pressing methods of rice bran oil processing. J. Food Drug Anal. 2019, 27, 71–82. [Google Scholar] [CrossRef]
- Xu, Z.; Godber, J.S. Antioxidant activities of major components of γ-oryzanol from rice bran using a linoleic acid model. J. Am. Oil Chem. Soc. 2001, 78, 645. [Google Scholar] [CrossRef]
- Posuwan, J.; Prangthip, P.; Leardkamolkarn, V.; Yamborisut, U.; Surasiang, R.; Charoensiri, R.; Kongkachuichai, R. Long-term supplementation of high pigmented rice bran oil (Oryza sativa L.) on amelioration of oxidative stress and histological changes in streptozotocin-induced diabetic rats fed a high fat diet; Riceberry bran oil. Food Chem. 2013, 138, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Leardkamolkarn, V.; Thongthep, W.; Suttiarporn, P.; Kongkachuichai, R.; Wongpornchai, S.; Wanavijitr, A. Chemopreventive properties of the bran extracted from a newly-developed Thai rice: The Riceberry. Food Chem. 2011, 125, 978–985. [Google Scholar] [CrossRef]
- Suttiarporn, P.; Chumpolsri, W.; Mahatheeranont, S.; Luangkamin, S.; Teepsawang, S.; Leardkamolkarn, V. Structures of phytosterols and triterpenoids with potential anti-cancer activity in bran of black non-glutinous rice. Nutrients 2015, 7, 1672–1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arya, A.; Arya, S.; Arya, M. Chemical Carcinogen and Cancer Risk: An overview. J. Chem. Pharm. Res. 2011, 3, 621–631. [Google Scholar]
- Punvittayagul, C.; Chariyakornkul, A.; Jarukamjorn, K.; Wongpoomchai, R. Protective Role of Vanillic Acid against Diethylnitrosamine- and 1,2-Dimethylhydrazine-Induced Hepatocarcinogenesis in Rats. Molecules 2021, 26, 2718. [Google Scholar] [CrossRef]
- Chariyakornkul, A.; Inboot, N.; Taya, S.; Wongpoomchai, R. Low-polar extract from seed of Cleistocalyx nervosum var. paniala modulates initiation and promotion stages of chemically-induced carcinogenesis in rats. Biomed. Pharmacother. 2021, 133, 110963. [Google Scholar] [CrossRef] [PubMed]
- Punvittayagul, C.; Chariyakornkul, A.; Chewonarin, T.; Jarukamjorn, K.; Wongpoomchai, R. Augmentation of diethylnitrosamine-induced early stages of rat hepatocarcinogenesis by 1,2-dimethylhydrazine. Drug Chem. Toxicol. 2019, 42, 641–648. [Google Scholar] [CrossRef]
- Phannasorn, W.; Chariyakornkul, A.; Sookwong, P.; Wongpoomchai, R. Comparative studies on the hepatoprotective effect of white and coloured rice bran oil against acetaminophen-induced oxidative stress in mice through antioxidant- and xenobiotic-metabolizing systems. Oxid. Med. Cell. Longev. 2021, 2021, 5510230. [Google Scholar] [CrossRef] [PubMed]
- Bird, R.P. Observation and quantification of aberrant crypts in the murine colon treated with a colon carcinogen: Preliminary findings. Cancer Lett. 1987, 37, 147–151. [Google Scholar] [CrossRef]
- Thumvijit, T.; Taya, S.; Punvittayagul, C.; Peerapornpisal, Y.; Wongpoomchai, R. Cancer chemopreventive effect of Spirogyra neglecta (Hassall) Kutzing on diethylnitrosamine-induced hepatocarcinogenesis in rats. Asian Pac. J. Cancer Prev. 2014, 15, 1611–1616. [Google Scholar] [CrossRef]
- Dokkaew, A.; Punvittayagul, C.; Insuan, O.; Limtrakul Dejkriengkraikul, P.; Wongpoomchai, R. Protective effects of defatted sticky rice bran extracts on the early stages of hepatocarcinogenesis in rats. Molecules 2019, 24, 2142. [Google Scholar] [CrossRef] [PubMed]
- Calik, A.; Ergün, A. Effect of lactulose supplementation on growth performance, intestinal histomorphology, cecal microbial population, and short-chain fatty acid composition of broiler chickens. Poult. Sci. 2015, 94, 2173–2182. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Benetou, V.; Lagiou, A.; Lagiou, P. Chemoprevention of cancer: Current evidence and future prospects. F1000Res 2015, 4, 916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenwald, P.; Clifford, C.K.; Milner, J.A. Diet and cancer prevention. Eur. J. Cancer 2001, 37, 948–965. [Google Scholar] [CrossRef]
- Nam, S.H.; Choi, S.P.; Kang, M.Y.; Koh, H.J.; Kozukue, N.; Friedman, M. Bran extracts from pigmented rice seeds inhibit tumor promotion in lymphoblastoid B cells by phorbol ester. Food Chem. Toxicol. 2005, 43, 741–745. [Google Scholar] [CrossRef]
- Nam, S.H.; Choi, S.P.; Kang, M.Y.; Koh, H.J.; Kozukue, N.; Friedman, M. Antioxidative activities of bran extracts from twenty one pigmented rice cultivars. Food Chem. 2006, 94, 613–620. [Google Scholar] [CrossRef]
- Moon, S.H.; Kim, D.; Shimizu, N.; Okada, T.; Hitoe, S.; Shimoda, H. Ninety-day oral toxicity study of rice-derived γ-oryzanol in Sprague-Dawley rats. Toxicol. Rep. 2017, 4, 9–18. [Google Scholar] [CrossRef]
- Fujiwara, S.; Sakurai, S.; Sugimoto, I.; Awata, N. Absorption and Metabolism of γ-Oryzanol in Rats. Chem. Pharm. Bull. (Tokyo) 1983, 31, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Punvittayagul, C.; Sankam, P.; Taya, S.; Wongpoomchai, R. Anticlastogenicity and anticarcinogenicity of purple rice extract in rats. Nutr. Cancer 2016, 68, 646–653. [Google Scholar] [CrossRef]
- Chuang, S.E.; Kuo, M.L.; Hsu, C.H.; Chen, C.R.; Lin, J.K.; Lai, G.M.; Hsieh, C.Y.; Cheng, A.L. Curcumin-containing diet inhibits diethylnitrosamine-induced murine hepatocarcinogenesis. Carcinogenesis 2000, 21, 331–335. [Google Scholar] [CrossRef]
- Grab, D.J.; Zedeck, M.S. Organ-specific Effects of the Carcinogen Methylazoxymethanol Related to Metabolism by Nicotinamide Adenine Dinucleotide-dependent Dehydrogenases. Cancer Res. 1977, 37, 4182–4189. [Google Scholar]
- Hirose, M.; Thamavit, W.; Asamoto, M.; Osawa, T.; Ito, N. Inhibition of glutathione S-transferase p type-positive foci development by linolic acid hydroperoxides and their secondary oxidative products in a rat in vivo midterm test for liver carcinogens. Toxicol. Lett. 1986, 32, 51–58. [Google Scholar] [CrossRef]
- Husain, K.; Zhang, A.; Shivers, S.; Davis-Yadley, A.; Coppola, D.; Yang, C.S.; Malafa, M.P. Chemoprevention of azoxymethane-induced colon carcinogenesis by delta-tocotrienol. Cancer Prev. Res. 2019, 12, 357–366. [Google Scholar] [CrossRef]
- Iqbal, J.; Minhajuddin, M.; Beg, Z.H. Suppression of diethylnitrosamine and 2-acetylaminofluorene-induced hepatocarcinogenesis in rats by tocotrienol-rich fraction isolated from rice bran oil. Eur. J. Cancer Prev. 2004, 13, 515–520. [Google Scholar] [CrossRef]
- Kim, S.P.; Kang, M.Y.; Nam, S.H.; Friedman, M. Dietary rice bran component γ-oryzanol inhibits tumor growth in tumor-bearing mice. Mol. Nutr. Food Res. 2012, 56, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Janezic, S.A.; Rao, A.V. Dose-dependent effects of dietary phytosterol on epithelial cell proliferation of the murine colon. Food Chem. Toxicol. 1992, 30, 611–616. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef]
- Khan, R.; Rehman, M.U.; Khan, A.Q.; Tahir, M.; Sultana, S. Glycyrrhizic acid suppresses 1,2-dimethylhydrazine-induced colon tumorigenesis in Wistar rats: Alleviation of inflammatory, proliferation, angiogenic, and apoptotic markers. Environ. Toxicol. 2018, 33, 1272–1283. [Google Scholar] [CrossRef]
- Mansour, D.F.; Abdallah, H.M.I.; Ibrahim, B.M.M.; Hegazy, R.R.; Esmail, R.S.E.; Abdel-Salam, L.O. The carcinogenic agent diethylnitrosamine induces early oxidative stress, inflammation and proliferation in rat liver, stomach and colon: Protective effect of ginger extract. Asian Pac. J. Cancer Prev. 2019, 20, 2551–2561. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Yu, S.; Park, H.J.; Jung, J.; Go, G.-W.; Kim, W. Rice bran oil ameliorates inflammatory responses by enhancing mitochondrial respiration in murine macrophages. PLoS ONE 2019, 14, e0222857. [Google Scholar] [CrossRef] [PubMed]
- Yamada, A.; Arakaki, R.; Saito, M.; Kudo, Y.; Ishimaru, N. Dual role of Fas/FasL-mediated signal in peripheral immune tolerance. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Campbell, S.E.; Stone, W.L.; Lee, S.; Whaley, S.; Yang, H.; Qui, M.; Goforth, P.; Sherman, D.; McHaffie, D.; Krishnan, K. Comparative effects of RRR-alpha- and RRR-gamma-tocopherol on proliferation and apoptosis in human colon cancer cell lines. BMC Cancer 2006, 6, 13. [Google Scholar] [CrossRef]
- Sakai, M.; Okabe, M.; Yamasaki, M.; Tachibana, H.; Yamada, K. Induction of apoptosis by tocotrienol in rat hepatoma dRLh-84 cells. Anticancer Res. 2004, 24, 1683–1688. [Google Scholar] [PubMed]
- Greenhalgh, K.; Meyer, K.M.; Aagaard, K.M.; Wilmes, P. The human gut microbiome in health: Establishment and resilience of microbiota over a lifetime. Environ. Microbiol. 2016, 18, 2103–2116. [Google Scholar] [CrossRef] [PubMed]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes ratio: A relevant marker of gut dysbiosis in obese patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef]
- Sun, T.; Liu, S.; Zhou, Y.; Yao, Z.; Zhang, D.; Cao, S.; Wei, Z.; Tan, B.; Li, Y.; Lian, Z.; et al. Evolutionary biologic changes of gut microbiota in an ’adenoma-carcinoma sequence’ mouse colorectal cancer model induced by 1, 2-Dimethylhydrazine. Oncotarget 2017, 8, 444–457. [Google Scholar] [CrossRef] [PubMed]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef]
- McNabney, S.M.; Henagan, T.M. Short chain fatty acids in the colon and peripheral tissues: A focus on butyrate, colon cancer, obesity and insulin resistance. Nutrients 2017, 9, 1348. [Google Scholar] [CrossRef]
- Zhang, J.; Song, L.; Wang, Y.; Liu, C.; Zhang, L.; Zhu, S.; Liu, S.; Duan, L. Beneficial effect of butyrate-producing Lachnospiraceae on stress-induced visceral hypersensitivity in rats. J. Gastroenterol. Hepatol. 2019, 34, 1368–1376. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.T.; Kantilal, H.K.; Davamani, F. The mechanism of bacteroides fragilis toxin contributes to colon cancer formation. Malays. J. Med. Sci. 2020, 27, 9–21. [Google Scholar] [CrossRef]
- Wu, Y.; Jiao, N.; Zhu, R.; Zhang, Y.; Wu, D.; Wang, A.-J.; Fang, S.; Tao, L.; Li, Y.; Cheng, S.; et al. Identification of microbial markers across populations in early detection of colorectal cancer. Nat. Commun. 2021, 12, 3063. [Google Scholar] [CrossRef] [PubMed]
- Ewaschuk, J.B.; Walker, J.W.; Diaz, H.; Madsen, K.L. Bioproduction of conjugated linoleic acid by probiotic bacteria occurs in vitro and in vivo in mice. J. Nutr. 2006, 136, 1483–1487. [Google Scholar] [CrossRef] [PubMed]
- Yasui, Y.; Suzuki, R.; Kohno, H.; Miyamoto, S.; Beppu, F.; Hosokawa, M.; Miyashita, K.; Tanaka, T. 9trans,11trans conjugated linoleic acid inhibits the development of azoxymethane-induced colonic aberrant crypt foci in rats. Nutr. Cancer 2007, 59, 82–91. [Google Scholar] [CrossRef] [PubMed]
- So, W.K.; Law, B.M.; Law, P.T.; Chan, C.W.; Chair, S.Y. Current Hypothesis for the Relationship between Dietary Rice Bran Intake, the Intestinal Microbiota and Colorectal Cancer Prevention. Nutrients 2016, 8, 569. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.X.; Williams, P.N.; Carey, A.M.; Zhu, Y.G.; Deacon, C.; Raab, A.; Feldmann, J.; Islam, R.M.; Meharg, A.A. Inorganic arsenic in rice bran and its products are an order of magnitude higher than in bulk grain. Environ. Sci. Technol. 2008, 42, 7542–7546. [Google Scholar] [CrossRef] [PubMed]
- Jariwalla, R.J. Rice-bran products: Phytonutrients with potential applications in preventive and clinical medicine. Drugs Exp. Clin. Res. 2001, 27, 17–26. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| tumor necrosis factor-alpha (TNF-α) | AAATGGCCCTCTCATCAGTCC | TCTGCTTGGTGGTTTGCTACGAC |
| Interleukin-6 (IL-6) | TGATGGATGCTTCCAAACTG | GAGCATTGGAAGTTGGGG TA |
| Interleukin-1 beta (IL-1β) | CACCTCTCAAGCAGAGCACAG | GGGTTCCATGGTGAAGTCAAC |
| inducible nitric oxide synthase (iNOS) | CAGGTGCTATTCCCAGCCCAACA | CATTCTGTGCAGTCCCAGTGAGGAA |
| nuclear factor kappa B (NF-κB) | GGCATGCGTTTCCGTTACAA | TGATCTTGATGGTGGGGTGC |
| β-actin | ACAGGATGCAGAAGGAGATTAC | AGAGTGAGGCCAGGATAGA |
| Treatment | Final Body Weight (g) | Liver Function Test (Unit/L) | |
|---|---|---|---|
| AST Activity | ALT Activity | ||
| Control | 397.0 ± 6.6 | 60 ± 1.7 | 27 ± 1.4 |
| RBBO | 387.0 ± 3.7 | 67 ± 6.1 | 27 ± 3.5 |
| DEN and DMH | 369.4 ± 10.0 * | 75 ± 2.8 * | 40 ± 2.5 * |
| RBBO + DEN and DMH | 352.5 ± 10.6 | 81 ± 4.8 | 49 ± 2.9 |
| Treatment | Number of PCNA Positive Cells in Hepatocytes | % Relative (PCNA + Cells/Total Colonocytes) | |
|---|---|---|---|
| PCNA+/1000GST-P+ Cells | PCNA+/1000 Surrounding Cells | ||
| Control | ND | 5.81 ± 1.39 | 15.31 ± 6.00 |
| RBBO | ND | 4.69 ± 2.13 | 15.52 ± 4.00 |
| DEN and DMH | 47.29 ± 7.98 | 13.58 ± 5.38 * | 46.87 ± 5.38 * |
| RBBO + DEN and DMH | 45.10 ± 8.29 | 12.75 ± 3.68 | 42.68 ± 3.68 |
| Treatment | Number of TUNEL Positive Cells in Hepatocytes | % Relative (TUNEL + Cells/Total Colonocytes) | |
|---|---|---|---|
| TUNEL+/1000GST-P+ Cells | TUNEL+/1000 Surrounding Cells | ||
| Control | ND | 21.47 ± 4.67 | 43.89 ± 8.57 |
| RBBO | ND | 26.85 ± 2.73 | 44.72 ± 7.48 |
| DEN and DMH | 79.86 ± 3.93 | 56.61 ± 5.38 * | 53.92 ± 3.61 * |
| RBBO + DEN and DMH | 91.32 ± 4.97 # | 73.28 ± 6.02 # | 64.84 ± 4.37 # |
| SCFA (µ mole/g Feces) | Treatment | |||
|---|---|---|---|---|
| Control | RBBO | DEN and DMH | RBBO + DEN and DMH | |
| Acetate | 46.34 ± 3.53 | 36.44 ± 6.20 * | 42.19 ± 4.92 | 39.72 ± 2.37 # |
| Propionate | 7.41 ± 1.31 | 6.64 ± 3.61 | 7.78 ± 2.29 | 7.36 ± 0.72 |
| Butyrate | 1.65 ± 0.11 | 2.15 ± 0.76 * | 1.16 ± 0.22 * | 2.09 ± 0.33 # |
| Isobutyrate | 10.77 ± 2.91 | 6.64 ± 2.64 * | 8.38 ± 1.06 * | 10.63 ± 2.59 |
| Valerate | 0.84 ± 0.22 | 0.74 ± 0.23 | 0.58 ± 0.10 * | 1.05 ± 0.43 # |
| Isovalerate | 2.48 ± 0.30 | 2.44 ± 0.43 | 1.64 ± 0.37 * | 1.97 ± 0.66 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phannasorn, W.; Pharapirom, A.; Thiennimitr, P.; Guo, H.; Ketnawa, S.; Wongpoomchai, R. Enriched Riceberry Bran Oil Exerts Chemopreventive Properties through Anti-Inflammation and Alteration of Gut Microbiota in Carcinogen-Induced Liver and Colon Carcinogenesis in Rats. Cancers 2022, 14, 4358. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14184358
Phannasorn W, Pharapirom A, Thiennimitr P, Guo H, Ketnawa S, Wongpoomchai R. Enriched Riceberry Bran Oil Exerts Chemopreventive Properties through Anti-Inflammation and Alteration of Gut Microbiota in Carcinogen-Induced Liver and Colon Carcinogenesis in Rats. Cancers. 2022; 14(18):4358. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14184358
Chicago/Turabian StylePhannasorn, Warunyoo, Aroonrat Pharapirom, Parameth Thiennimitr, Huina Guo, Sunantha Ketnawa, and Rawiwan Wongpoomchai. 2022. "Enriched Riceberry Bran Oil Exerts Chemopreventive Properties through Anti-Inflammation and Alteration of Gut Microbiota in Carcinogen-Induced Liver and Colon Carcinogenesis in Rats" Cancers 14, no. 18: 4358. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14184358