A Comprehensive Analysis of Tn and STn Antigen Expression in Esophageal Adenocarcinoma

 , , , , ,
, , , , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Selection and IHC Establishment
2.2. Esophageal Cancer Tissue Microarray
2.3. Immunohistochemistry
2.4. Real-Time qRT-PCR
2.5. Statistical Analysis
3. Results
3.1. Esophageal Adenocarcinoma Express High Levels of Tn and STn Antigen
3.2. Demographic and Histopathological Parameters
3.3. Tn Antigen Immunostaining in Esophageal Cancers
3.4. The Association between Tn Antigens and Histopathological Tumor Phenotype
3.5. STn Antigen Immunostaining in Esophageal Cancers
3.6. The Association between STn Antigens and Histopathological Tumor Phenotype
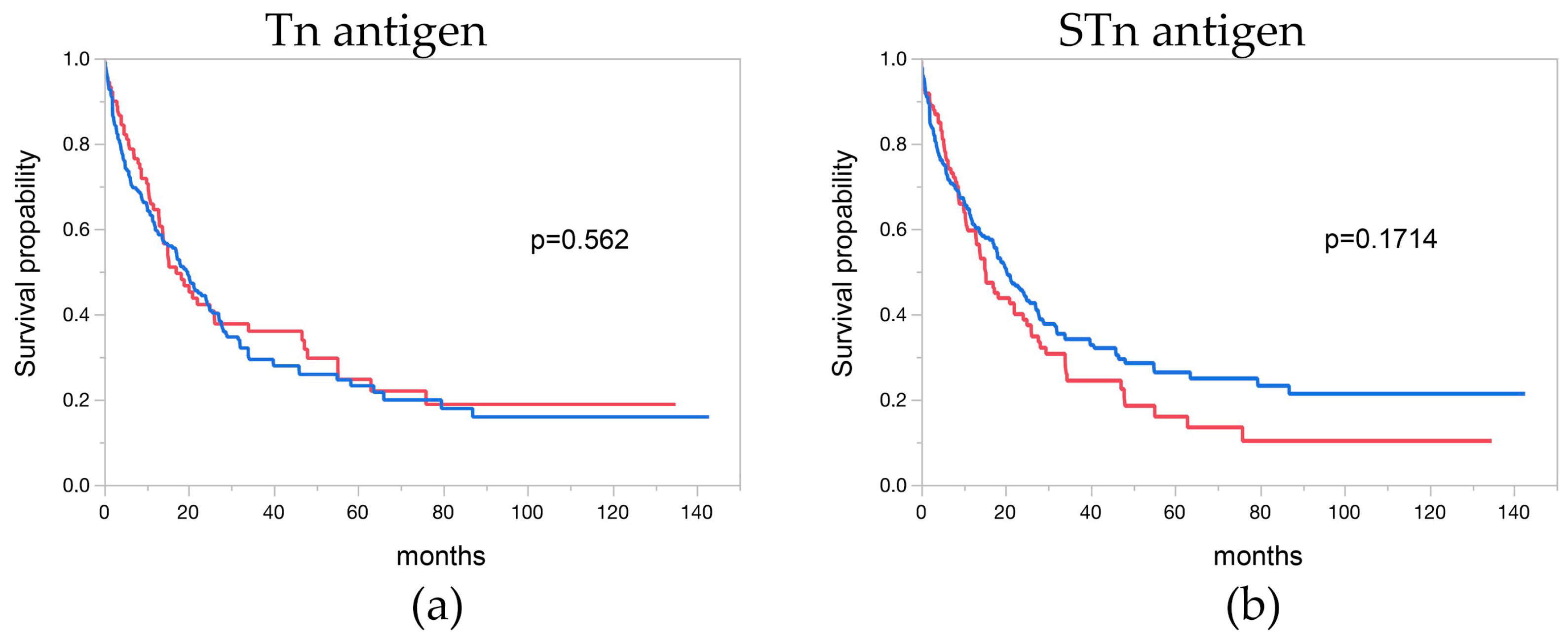
3.7. Tn and STn Antigen and Overall Survival
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Joseph, A.; Raja, S.; Kamath, S.; Jang, S.; Allende, D.; McNamara, M.; Videtic, G.; Murthy, S.; Bhatt, A. Esophageal adenocarcinoma: A dire need for early detection and treatment. Cleve Clin. J. Med. 2022, 89, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, N.J.; Richter, J.E. Barrett’s oesophagus. Lancet 2009, 373, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Domper Arnal, M.J.; Ferrandez Arenas, A.; Lanas Arbeloa, A. Esophageal cancer: Risk factors, screening and endoscopic treatment in Western and Eastern countries. World J. Gastroenterol. 2015, 21, 7933–7943. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Q.; Ma, Y.L.; Qin, Q.; Wang, P.H.; Luo, Y.; Xu, P.F.; Cui, Y. Epidemiology of esophageal cancer in 2020 and projections to 2030 and 2040. Thorac. Cancer 2023, 14, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Obermannova, R.; Alsina, M.; Cervantes, A.; Leong, T.; Lordick, F.; Nilsson, M.; van Grieken, N.C.T.; Vogel, A.; Smyth, E.C.; ESMO Guidelines Committee. Oesophageal cancer: ESMO Clinical Practice Guideline for diagnosis, treatment and follow-up. Ann. Oncol. 2022, 33, 992–1004. [Google Scholar] [CrossRef]
- Springer, G.F. Immunoreactive T and Tn epitopes in cancer diagnosis, prognosis, and immunotherapy. J. Mol. Med. 1997, 75, 594–602. [Google Scholar] [CrossRef]
- Bagdonaite, I.; Pallesen, E.M.H.; Nielsen, M.I.; Bennett, E.P.; Wandall, H.H. Mucin-Type O-GalNAc Glycosylation in Health and Disease. Adv. Exp. Med. Biol. 2021, 1325, 25–60. [Google Scholar] [CrossRef]
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: Mechanisms and clinical implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef]
- Ju, T.; Otto, V.I.; Cummings, R.D. The Tn antigen-structural simplicity and biological complexity. Angew. Chem. Int. Ed. Engl. 2011, 50, 1770–1791. [Google Scholar] [CrossRef]
- Sewell, R.; Backstrom, M.; Dalziel, M.; Gschmeissner, S.; Karlsson, H.; Noll, T.; Gatgens, J.; Clausen, H.; Hansson, G.C.; Burchell, J.; et al. The ST6GalNAc-I sialyltransferase localizes throughout the Golgi and is responsible for the synthesis of the tumor-associated sialyl-Tn O-glycan in human breast cancer. J. Biol. Chem. 2006, 281, 3586–3594. [Google Scholar] [CrossRef]
- Munkley, J. The Role of Sialyl-Tn in Cancer. Int. J. Mol. Sci. 2016, 17, 275. [Google Scholar] [CrossRef] [PubMed]
- Romer, T.B.; Aasted, M.K.M.; Dabelsteen, S.; Groen, A.; Schnabel, J.; Tan, E.; Pedersen, J.W.; Haue, A.D.; Wandall, H.H. Mapping of truncated O-glycans in cancers of epithelial and non-epithelial origin. Br. J. Cancer 2021, 125, 1239–1250. [Google Scholar] [CrossRef]
- Fu, C.; Zhao, H.; Wang, Y.; Cai, H.; Xiao, Y.; Zeng, Y.; Chen, H. Tumor-associated antigens: Tn antigen, sTn antigen, and T antigen. HLA 2016, 88, 275–286. [Google Scholar] [CrossRef]
- Flucke, U.; Zirbes, T.K.; Schroder, W.; Monig, S.P.; Koch, V.; Schmitz, K.; Thiele, J.; Dienes, H.P.; Holscher, A.H.; Baldus, S.E. Expression of mucin-associated carbohydrate core antigens in esophageal squamous cell carcinomas. Anticancer Res. 2001, 21, 2189–2193. [Google Scholar] [PubMed]
- Radhakrishnan, P.; Dabelsteen, S.; Madsen, F.B.; Francavilla, C.; Kopp, K.L.; Steentoft, C.; Vakhrushev, S.Y.; Olsen, J.V.; Hansen, L.; Bennett, E.P.; et al. Immature truncated O-glycophenotype of cancer directly induces oncogenic features. Proc. Natl. Acad. Sci. USA 2014, 111, E4066–E4075. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, B.T.; Picksak, A.S.; Kwiatkowski, M.; Grupp, K.; Jucker, M.; Bachmann, K.; Mercanoglu, B.; Izbicki, J.R.; Kahlert, C.; Bockhorn, M.; et al. Truncated O-GalNAc glycans impact on fundamental signaling pathways in pancreatic cancer. Glycobiology 2021, cwab088. [Google Scholar] [CrossRef]
- Werther, J.L.; Rivera-MacMurray, S.; Bruckner, H.; Tatematsu, M.; Itzkowitz, S.H. Mucin-associated sialosyl-Tn antigen expression in gastric cancer correlates with an adverse outcome. Br. J. Cancer 1994, 69, 613–616. [Google Scholar] [CrossRef]
- Yamada, T.; Watanabe, A.; Yamada, Y.; Shino, Y.; Tanase, M.; Yamashita, J.; Miwa, T.; Nakano, H. Sialosyl Tn antigen expression is associated with the prognosis of patients with advanced gastric cancer. Cancer 1995, 76, 1529–1536. [Google Scholar] [CrossRef]
- Cotton, S.; Azevedo, R.; Gaiteiro, C.; Ferreira, D.; Lima, L.; Peixoto, A.; Fernandes, E.; Neves, M.; Neves, D.; Amaro, T.; et al. Targeted O-glycoproteomics explored increased sialylation and identified MUC16 as a poor prognosis biomarker in advanced-stage bladder tumours. Mol. Oncol. 2017, 11, 895–912. [Google Scholar] [CrossRef]
- Akita, K.; Yoshida, S.; Ikehara, Y.; Shirakawa, S.; Toda, M.; Inoue, M.; Kitawaki, J.; Nakanishi, H.; Narimatsu, H.; Nakada, H. Different levels of sialyl-Tn antigen expressed on MUC16 in patients with endometriosis and ovarian cancer. Int. J. Gynecol. Cancer 2012, 22, 531–538. [Google Scholar] [CrossRef]
- Hashiguchi, Y.; Kasai, M.; Fukuda, T.; Ichimura, T.; Yasui, T.; Sumi, T. Serum Sialyl-Tn (STN) as a Tumor Marker in Patients with Endometrial Cancer. Pathol. Oncol. Res. 2016, 22, 501–504. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Ogawa, H.; Nakanishi, K.; Tanizawa, O.; Karino, K.; Endo, J. Clinical value of sialyl Tn antigen in patients with gynecologic tumors. Obstet Gynecol. 1990, 75, 1032–1036. [Google Scholar]
- Kobayashi, H.; Terao, T.; Kawashima, Y. Sialyl Tn as a prognostic marker in epithelial ovarian cancer. Br. J. Cancer 1992, 66, 984–985. [Google Scholar] [CrossRef] [PubMed]
- Nanashima, A.; Yamaguchi, H.; Nakagoe, T.; Matsuo, S.; Sumida, Y.; Tsuji, T.; Sawai, T.; Yamaguchi, E.; Yasutake, T.; Ayabe, H. High serum concentrations of sialyl Tn antigen in carcinomas of the biliary tract and pancreas. J. Hepatobiliary Pancreat Surg. 1999, 6, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Nishimura, G.; Nonomura, A.; Miwa, K.; Miyazaki, I. Serological studies on CEA, CA 19-9, STn and SLX in colorectal cancer. Hepatogastroenterology 1999, 46, 914–919. [Google Scholar]
- Nakagoe, T.; Sawai, T.; Tsuji, T.; Jibiki, M.A.; Nanashima, A.; Yamaguchi, H.; Yasutake, T.; Ayabe, H.; Arisawa, K.; Ishikawa, H. Predictive factors for preoperative serum levels of sialy Lewis(x), sialyl Lewis(a) and sialyl Tn antigens in gastric cancer patients. Anticancer Res. 2002, 22, 451–458. [Google Scholar]
- Imada, T.; Rino, Y.; Takahashi, M.; Hatori, S.; Shiozawa, M.; Amano, T.; Kondo, J. Serum CA 19-9, SLX, STN and CEA levels of the peripheral and the draining venous blood in gastric cancer. Hepatogastroenterology 1999, 46, 2086–2090. [Google Scholar]
- Genega, E.M.; Hutchinson, B.; Reuter, V.E.; Gaudin, P.B. Immunophenotype of high-grade prostatic adenocarcinoma and urothelial carcinoma. Mod. Pathol. 2000, 13, 1186–1191. [Google Scholar] [CrossRef]
- Arai, T.; Fujita, K.; Fujime, M.; Irimura, T. Expression of sialylated MUC1 in prostate cancer: Relationship to clinical stage and prognosis. Int. J. Urol. 2005, 12, 654–661. [Google Scholar] [CrossRef]
- Reichelt, U.; Duesedau, P.; Tsourlakis, M.; Quaas, A.; Link, B.C.; Schurr, P.G.; Kaifi, J.T.; Gros, S.J.; Yekebas, E.F.; Marx, A.; et al. Frequent homogeneous HER-2 amplification in primary and metastatic adenocarcinoma of the esophagus. Mod. Pathol. 2007, 20, 120–129. [Google Scholar] [CrossRef]
- Grupp, K.; Uzunoglu, F.G.; Melling, N.; Hofmann, B.; El Gammal, A.T.; Groteluschen, R.; Heumann, A.; Bellon, E.; Reeh, M.; Wolters-Eisfeld, G.; et al. FOXO1 overexpression and loss of pSerine256-FOXO1 expression predicts clinical outcome in esophageal adenocarcinomas. Sci. Rep. 2018, 8, 17370. [Google Scholar] [CrossRef] [PubMed]
- Dancau, A.M.; Simon, R.; Mirlacher, M.; Sauter, G. Tissue Microarrays. Methods Mol. Biol. 2016, 1381, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Bosman, F.T.; World Health Organization; International Agency for Research on Cancer. WHO Classification of Tumours of the Digestive System, 4th ed.; International Agency for Research on Cancer: Lyon, France, 2010; p. 417. [Google Scholar]
- Sobin, L.H.; Gospodarowicz, M.K.; Wittekind, C.; International Union against Cancer. TNM Classification of Malignant Tumours, 7th ed.; Wiley-Blackwell: Chichester, UK; Hoboken, NJ, USA, 2010; p. 66. [Google Scholar]
- Steentoft, C.; Vakhrushev, S.Y.; Joshi, H.J.; Kong, Y.; Vester-Christensen, M.B.; Schjoldager, K.T.; Lavrsen, K.; Dabelsteen, S.; Pedersen, N.B.; Marcos-Silva, L.; et al. Precision mapping of the human O-GalNAc glycoproteome through SimpleCell technology. EMBO J. 2013, 32, 1478–1488. [Google Scholar] [CrossRef] [PubMed]
- Aryal, R.P.; Ju, T.; Cummings, R.D. Tight complex formation between Cosmc chaperone and its specific client non-native T-synthase leads to enzyme activity and client-driven dissociation. J. Biol. Chem. 2012, 287, 15317–15329. [Google Scholar] [CrossRef]
- Ju, T.; Xia, B.; Aryal, R.P.; Wang, W.; Wang, Y.; Ding, X.; Mi, R.; He, M.; Cummings, R.D. A novel fluorescent assay for T-synthase activity. Glycobiology 2011, 21, 352–362. [Google Scholar] [CrossRef]
- Wang, Y.; Liao, X.; Ye, Q.; Huang, L. Clinic implication of MUC1 O-glycosylation and C1GALT1 in esophagus squamous cell carcinoma. Sci. China Life Sci. 2018, 61, 1389–1395. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.H.; Hu, R.H.; Huang, M.J.; Lai, I.R.; Chen, C.H.; Lai, H.S.; Wu, Y.M.; Huang, M.C. C1GALT1 promotes invasive phenotypes of hepatocellular carcinoma cells by modulating integrin beta1 glycosylation and activity. PLoS ONE 2014, 9, e94995. [Google Scholar] [CrossRef]
- Wolters-Eisfeld, G.; Mercanoglu, B.; Hofmann, B.T.; Wolpers, T.; Schnabel, C.; Harder, S.; Steffen, P.; Bachmann, K.; Steglich, B.; Schrader, J.; et al. Loss of complex O-glycosylation impairs exocrine pancreatic function and induces MODY8-like diabetes in mice. Exp. Mol. Med. 2018, 50, 1–13. [Google Scholar] [CrossRef]
- Mi, R.; Song, L.; Wang, Y.; Ding, X.; Zeng, J.; Lehoux, S.; Aryal, R.P.; Wang, J.; Crew, V.K.; van Die, I.; et al. Epigenetic silencing of the chaperone Cosmc in human leukocytes expressing tn antigen. J. Biol. Chem. 2012, 287, 41523–41533. [Google Scholar] [CrossRef]
- Zeng, J.; Mi, R.; Wang, Y.; Li, Y.; Lin, L.; Yao, B.; Song, L.; van Die, I.; Chapman, A.B.; Cummings, R.D.; et al. Promoters of Human Cosmc and T-synthase Genes Are Similar in Structure, Yet Different in Epigenetic Regulation. J. Biol. Chem. 2015, 290, 19018–19033. [Google Scholar] [CrossRef]
- Thomas, D.; Sagar, S.; Caffrey, T.; Grandgenett, P.M.; Radhakrishnan, P. Truncated O-glycans promote epithelial-to-mesenchymal transition and stemness properties of pancreatic cancer cells. J. Cell. Mol. Med. 2019, 23, 6885–6896. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, B.T.; Schluter, L.; Lange, P.; Mercanoglu, B.; Ewald, F.; Folster, A.; Picksak, A.S.; Harder, S.; El Gammal, A.T.; Grupp, K.; et al. COSMC knockdown mediated aberrant O-glycosylation promotes oncogenic properties in pancreatic cancer. Mol. Cancer 2015, 14, 109. [Google Scholar] [CrossRef] [PubMed]
- Dombek, G.E.; Ore, A.S.; Cheng, J.; Matsumoto, Y.; Glickman, J.N.; Fleishman, A.; Heimburg-Molinaro, J.; Poylin, V.Y.; Fabrizio, A.; Cataldo, T.; et al. Immunohistochemical analysis of Tn antigen expression in colorectal adenocarcinoma and precursor lesions. BMC Cancer 2022, 22, 1281. [Google Scholar] [CrossRef] [PubMed]
- RodrIguez, E.; Schetters, S.T.T.; van Kooyk, Y. The tumour glyco-code as a novel immune checkpoint for immunotherapy. Nat. Rev. Immunol. 2018, 18, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Stowell, S.R.; Ju, T.; Cummings, R.D. Protein glycosylation in cancer. Annu. Rev. Pathol. 2015, 10, 473–510. [Google Scholar] [CrossRef]
- Peixoto, A.; Relvas-Santos, M.; Azevedo, R.; Santos, L.L.; Ferreira, J.A. Protein Glycosylation and Tumor Microenvironment Alterations Driving Cancer Hallmarks. Front. Oncol. 2019, 9, 380. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, C.; Radhakrishnan, P. The (Sialyl) Tn antigen: Contributions to immunosuppression in gastrointestinal cancers. Front. Oncol. 2022, 12, 1093496. [Google Scholar] [CrossRef]
- Dusoswa, S.A.; Verhoeff, J.; Abels, E.; Mendez-Huergo, S.P.; Croci, D.O.; Kuijper, L.H.; de Miguel, E.; Wouters, V.; Best, M.G.; Rodriguez, E.; et al. Glioblastomas exploit truncated O-linked glycans for local and distant immune modulation via the macrophage galactose-type lectin. Proc. Natl. Acad. Sci. USA 2020, 117, 3693–3703. [Google Scholar] [CrossRef]
- Gonzalez-Gil, A.; Schnaar, R.L. Siglec Ligands. Cells 2021, 10, 1260. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Jia, N.; Heimburg-Molinaro, J.; Cummings, R.D. Targeting Tn-positive tumors with an afucosylated recombinant anti-Tn IgG. Sci. Rep. 2023, 13, 5027. [Google Scholar] [CrossRef]
- Prendergast, J.M.; Galvao da Silva, A.P.; Eavarone, D.A.; Ghaderi, D.; Zhang, M.; Brady, D.; Wicks, J.; DeSander, J.; Behrens, J.; Rueda, B.R. Novel anti-Sialyl-Tn monoclonal antibodies and antibody-drug conjugates demonstrate tumor specificity and anti-tumor activity. MAbs 2017, 9, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Posey, A.D., Jr.; Schwab, R.D.; Boesteanu, A.C.; Steentoft, C.; Mandel, U.; Engels, B.; Stone, J.D.; Madsen, T.D.; Schreiber, K.; Haines, K.M.; et al. Engineered CAR T Cells Targeting the Cancer-Associated Tn-Glycoform of the Membrane Mucin MUC1 Control Adenocarcinoma. Immunity 2016, 44, 1444–1454. [Google Scholar] [CrossRef] [PubMed]
- Lodewijk, I.; Duenas, M.; Paramio, J.M.; Rubio, C. CD44v6, STn & O-GD2: Promising tumor associated antigens paving the way for new targeted cancer therapies. Front. Immunol. 2023, 14, 1272681. [Google Scholar] [CrossRef] [PubMed]
- Raglow, Z.; McKenna, M.K.; Bonifant, C.L.; Wang, W.; Pasca di Magliano, M.; Stadlmann, J.; Penninger, J.M.; Cummings, R.D.; Brenner, M.K.; Markovitz, D.M. Targeting glycans for CAR therapy: The advent of sweet CARs. Mol. Ther. 2022, 30, 2881–2890. [Google Scholar] [CrossRef]
- Munkley, J. Aberrant Sialylation in Cancer: Therapeutic Opportunities. Cancers 2022, 14, 4248. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameter | EACs (n = 501) | |
|---|---|---|
| Sex | male | 425 |
| female | 76 | |
| Tumor stage | pT1 | 108 |
| pT2 | 87 | |
| pT3 | 278 | |
| pT4 | 28 | |
| Lymph node metastasis | pN0 | 135 |
| pN1 | 172 | |
| pN2 | 95 | |
| pN3 | 99 | |
| UICC | I | 110 |
| II | 87 | |
| III | 229 | |
| IV | 75 | |
| Distant Metastasis | M0 | 421 |
| M1 | 80 | |
| Grading | G1 | 14 |
| G2 | 318 | |
| G3 | 169 |
| Tn Antigen (CD175) | ||||
|---|---|---|---|---|
| Parameter | n Evaluable | Negative (%) | Positive (%) | p Value |
| All cancers | 311 | 28.9 | 71.1 | |
| Tumor stage | ||||
| pT1 | 51 | 31.4 | 68.6 | 0.0266 |
| pT2 | 58 | 39.7 | 60.3 | |
| pT3 | 182 | 26.4 | 73.6 | |
| pT4 | 17 | 5.9 | 94.1 | |
| Lymph node metastasis | ||||
| pN0 | 76 | 32.9 | 67.1 | 0.2877 |
| pN1 | 107 | 28.0 | 72.0 | |
| pN2 | 56 | 33.9 | 66.1 | |
| pN3 | 60 | 20.00 | 80.00 | |
| UICC | ||||
| I | 55 | 30.9 | 69.1 | 0.0732 |
| II | 51 | 41.2 | 58.8 | |
| III | 157 | 26.1 | 73.9 | |
| IV | 40 | 17.50 | 82.50 | |
| Distant metastasis | ||||
| M0 | 259 | 29.7 | 70.3 | 0.2579 |
| M1 | 46 | 21.7 | 78.3 | |
| Resection margin | ||||
| R0 | 219 | 29.7 | 70.3 | 0.7256 |
| R1 | 72 | 25.00 | 75.00 | |
| R2 | 8 | 25.00 | 75.00 | |
| Grading | ||||
| G1 | 10 | 20.00 | 80.00 | 0.2463 |
| G2 | 108 | 34.3 | 65.7 | |
| G3 | 183 | 25.7 | 74.3 | |
| STn Antigen (CD175s) | ||||
|---|---|---|---|---|
| Parameter | n Evaluable | Negative (%) | Positive (%) | p Value |
| All cancers | 353 | 29.5 | 70.5 | |
| Tumor stage | ||||
| pT1 | 59 | 25.4 | 74.6 | 0.1218 |
| pT2 | 63 | 39.7 | 60.3 | |
| pT3 | 208 | 28.8 | 71.2 | |
| pT4 | 20 | 15.0 | 85.0 | |
| Lymph node metastasis | ||||
| pN0 | 86 | 26.7 | 73.3 | 0.8868 |
| pN1 | 117 | 30.8 | 69.2 | |
| pN2 | 62 | 32.3 | 67.7 | |
| pN3 | 76 | 28.9 | 71.1 | |
| UICC | ||||
| I | 62 | 25.8 | 74.2 | 0.581 |
| II | 56 | 35.7 | 64.3 | |
| III | 178 | 30.3 | 69.7 | |
| IV | 48 | 25.0 | 75.0 | |
| Distant metastasis | ||||
| M0 | 293 | 30.4 | 69.9 | 0.5062 |
| M1 | 54 | 25.9 | 74.1 | |
| Resection margin | ||||
| R0 | 248 | 30.2 | 69.8 | 0.7611 |
| R1 | 85 | 27.1 | 72.9 | |
| R2 | 8 | 37.5 | 62.5 | |
| Grading | ||||
| G1 | 12 | 25.0 | 75.0 | 0.8482 |
| G2 | 120 | 27.5 | 72.5 | |
| G3 | 210 | 30.0 | 70.0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mercanoglu, B.; Karstens, K.-F.; Giannou, A.D.; Meiners, J.; Lücke, J.; Seeger, P.; Brackrock, V.; Güngör, C.; Izbicki, J.R.; Bockhorn, M.; et al. A Comprehensive Analysis of Tn and STn Antigen Expression in Esophageal Adenocarcinoma. Cancers 2024, 16, 240. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers16020240
Mercanoglu B, Karstens K-F, Giannou AD, Meiners J, Lücke J, Seeger P, Brackrock V, Güngör C, Izbicki JR, Bockhorn M, et al. A Comprehensive Analysis of Tn and STn Antigen Expression in Esophageal Adenocarcinoma. Cancers. 2024; 16(2):240. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers16020240
Chicago/Turabian StyleMercanoglu, Baris, Karl-Frederick Karstens, Anastasios D. Giannou, Jan Meiners, Jöran Lücke, Philipp Seeger, Vera Brackrock, Cenap Güngör, Jakob R. Izbicki, Maximilian Bockhorn, and et al. 2024. "A Comprehensive Analysis of Tn and STn Antigen Expression in Esophageal Adenocarcinoma" Cancers 16, no. 2: 240. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers16020240