
The Role of Curcumin in Cancer: A Focus on the PI3K/Akt Pathway
 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Structure–Activity Relationships of Curcumin
3. Pharmacokinetics of Curcumin and New Delivery Systems
4. PI3K/Akt Signaling and Cancer
5. Curcumin and PI3K/Akt/mTOR Signaling
6. Curcumin against the PI3K/Akt Pathway in Different Malignancies
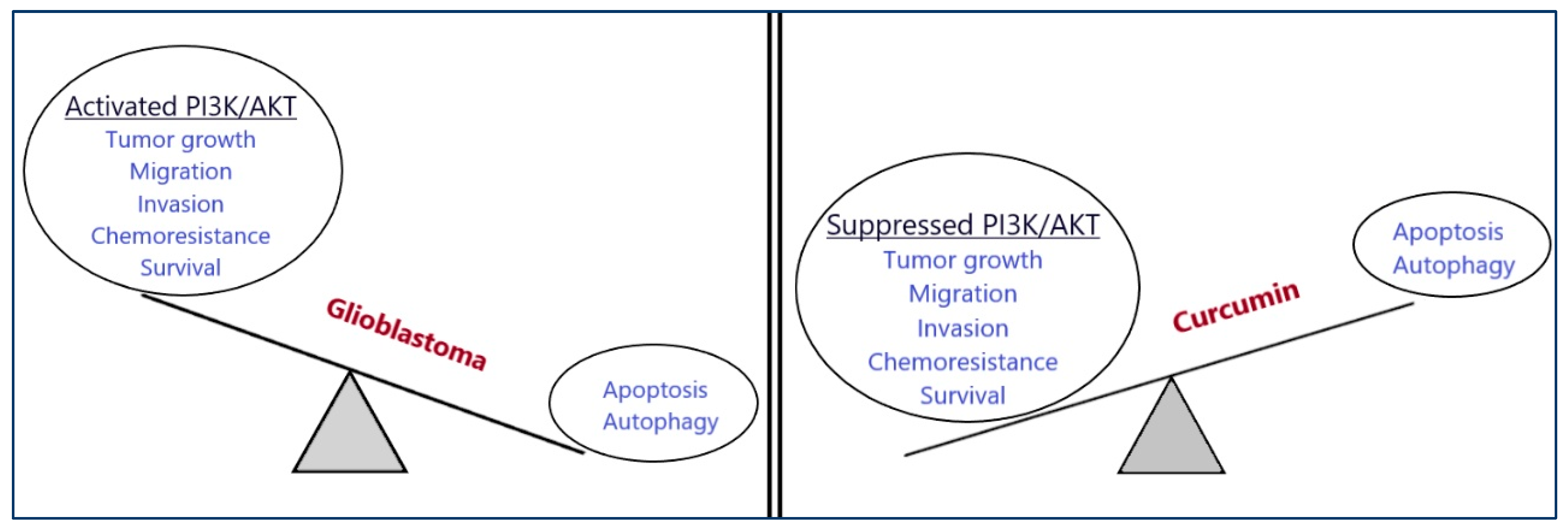
6.1. Glioblastoma
6.2. Lung Cancer
6.3. Breast Cancer
6.4. Prostate Cancer
6.5. Colorectal Cancer
6.6. Thyroid Cancer
6.7. Leukemia
7. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Motofei, I.G. Biology of cancer; from cellular and molecular mechanisms to developmental processes and adaptation. Semin. Cancer Biol. 2022, 86, 600–615. [Google Scholar] [CrossRef] [PubMed]
- Piña-Sánchez, P.; Chávez-González, A.; Ruiz-Tachiquín, M.; Vadillo, E.; Monroy-García, A.; Montesinos, J.J.; Grajales, R.; Gutiérrez de la Barrera, M.; Mayani, H. Cancer Biology, Epidemiology, and Treatment in the 21st Century: Current Status and Future Challenges From a Biomedical Perspective. Cancer Control 2021, 28, 10732748211038735. [Google Scholar] [CrossRef] [PubMed]
- Yip, H.Y.K.; Papa, A. Signaling Pathways in Cancer: Therapeutic Targets, Combinatorial Treatments, and New Developments. Cells 2021, 10, 659. [Google Scholar] [CrossRef] [PubMed]
- Martini, M.; De Santis, M.C.; Braccini, L.; Gulluni, F.; Hirsch, E. PI3K/AKT signaling pathway and cancer: An updated review. Ann. Med. 2014, 46, 372–383. [Google Scholar] [CrossRef] [PubMed]
- Glaviano, A.; Foo, A.S.C.; Lam, H.Y.; Yap, K.C.H.; Jacot, W.; Jones, R.H.; Eng, H.; Nair, M.G.; Makvandi, P.; Geoerger, B.; et al. PI3K/AKT/mTOR signaling transduction pathway and targeted therapies in cancer. Mol. Cancer 2023, 22, 138. [Google Scholar] [CrossRef] [PubMed]
- Stefani, C.; Miricescu, D.; Stanescu-Spinu, I.I.; Nica, R.I.; Greabu, M.; Totan, A.R.; Jinga, M. Growth Factors, PI3K/AKT/mTOR and MAPK Signaling Pathways in Colorectal Cancer Pathogenesis: Where Are We Now? Int. J. Mol. Sci. 2021, 22, 10260. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kwok-Shing, N.P.; Kucherlapati, M.; Chen, F.; Liu, Y.; Tsang, Y.H.; de Velasco, G.; Jeong, K.J.; Akbani, R.; Hadjipanayis, A.; et al. A Pan-Cancer Proteogenomic Atlas of PI3K/AKT/mTOR Pathway Alterations. Cancer Cell 2017, 31, 820–832. [Google Scholar] [CrossRef]
- Li, Q.; Li, Z.; Luo, T.; Shi, H. Targeting the PI3K/AKT/mTOR and RAF/MEK/ERK pathways for cancer therapy. Mol. Biomed. 2022, 3, 47. [Google Scholar] [CrossRef]
- Fleming-de-Moraes, C.D.; Rocha, M.R.; Tessmann, J.W.; de Araujo, W.M.; Morgado-Diaz, J.A. Crosstalk between PI3K/Akt and Wnt/β-catenin pathways promote colorectal cancer progression regardless of mutational status. Cancer Biol. Ther. 2022, 23, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mundekkad, D.; Cho, W.C. Applications of Curcumin and Its Nanoforms in the Treatment of Cancer. Pharmaceutics 2023, 15, 2223. [Google Scholar] [CrossRef] [PubMed]
- Kah, G.; Chandran, R.; Abrahamse, H. Curcumin a Natural Phenol and Its Therapeutic Role in Cancer and Photodynamic Therapy: A Review. Pharmaceutics 2023, 15, 639. [Google Scholar] [CrossRef] [PubMed]
- Kyritsis, A.P.; Bondy, M.L.; Levin, V.A. Modulation of glioma risk and progression by dietary nutrients and antiinflammatory agents. Nutr. Cancer 2011, 63, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Hassanizadeh, S.; Shojaei, M.; Bagherniya, M.; Orekhov, A.N.; Sahebkar, A. Effect of nano-curcumin on various diseases: A comprehensive review of clinical trials. Biofactors 2023, 49, 512–533. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.T.; Newmark, H.L.; Frenkel, K. Inhibitory effects of curcumin on tumorigenesis in mice. J. Cell. Biochem. Suppl. 1997, 27, 26–34. [Google Scholar] [CrossRef]
- Priyadarsini, K.I. Chemical and structural features influencing the biological activity of curcumin. Curr. Pharm. Des. 2013, 19, 2093–2100. [Google Scholar] [PubMed]
- Nelson, K.M.; Dahlin, J.L.; Bisson, J.; Graham, J.; Pauli, G.F.; Walters, M.A. The Essential Medicinal Chemistry of Curcumin. J. Med. Chem. 2017, 60, 1620–1637. [Google Scholar] [CrossRef] [PubMed]
- Zoi, V.; Galani, V.; Tsekeris, P.; Kyritsis, A.P.; Alexiou, G.A. Radiosensitization and Radioprotection by Curcumin in Glioblastoma and Other Cancers. Biomedicines 2022, 10, 312. [Google Scholar] [CrossRef]
- Dhillon, N.; Aggarwal, B.B.; Newman, R.A.; Wolff, R.A.; Kunnumakkara, A.B.; Abbruzzese, J.L.; Ng, C.S.; Badmaev, V.; Kurzrock, R. Phase II Trial of Curcumin in Patients with Advanced Pancreatic Cancer. Clin. Cancer Res. 2008, 14, 4491–4499. [Google Scholar] [CrossRef]
- Sharma, R.A.; McLelland, H.R.; Hill, K.A.; Ireson, C.R.; Euden, S.A.; Manson, M.M.; Pirmohamed, M.; Marnett, L.J.; Gescher, A.J.; Steward, W.P. Pharmacodynamic and pharmacokinetic study of oral Curcuma extract in patients with colorectal cancer. Clin. Cancer Res. 2001, 7, 1894–1900. [Google Scholar]
- Vareed, S.K.; Kakarala, M.; Ruffin, M.T.; Crowell, J.A.; Normolle, D.P.; Djuric, Z.; Brenner, D.E. Pharmacokinetics of curcumin conjugate metabolites in healthy human subjects. Cancer Epidemiol. Biomark. Prev. 2008, 17, 1411–1417. [Google Scholar] [CrossRef] [PubMed]
- Begum, A.N.; Jones, M.R.; Lim, G.P.; Morihara, T.; Kim, P.; Heath, D.D.; Rock, C.L.; Pruitt, M.A.; Yang, F.; Hudspeth, B.; et al. Curcumin structure-function, bioavailability, and efficacy in models of neuroinflammation and Alzheimer’s disease. J. Pharmacol. Exp. Ther. 2008, 326, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Schiborr, C.; Eckert, G.P.; Rimbach, G.; Frank, J. A validated method for the quantification of curcumin in plasma and brain tissue by fast narrow-bore high-performance liquid chromatography with fluorescence detection. Anal. Bioanal. Chem. 2010, 397, 1917–1925. [Google Scholar] [CrossRef]
- Sohn, S.I.; Priya, A.; Balasubramaniam, B.; Muthuramalingam, P.; Sivasankar, C.; Selvaraj, A.; Valliammai, A.; Jothi, R.; Pandian, S. Biomedical Applications and Bioavailability of Curcumin-An Updated Overview. Pharmaceutics 2021, 13, 2102. [Google Scholar] [CrossRef]
- Kanai, M.; Otsuka, Y.; Otsuka, K.; Sato, M.; Nishimura, T.; Mori, Y.; Kawaguchi, M.; Hatano, E.; Kodama, Y.; Matsumoto, S.; et al. Phase I study investigating the safety and pharmacokinetics of highly bioavailable curcumin (Theracurmin) in cancer patients. Cancer Chemother. Pharmacol. 2013, 71, 1521–1530. [Google Scholar] [CrossRef]
- Garcea, G.; Berry, D.P.; Jones, D.J.; Singh, R.; Dennison, A.R.; Farmer, P.B.; Sharma, R.A.; Steward, W.P.; Gescher, A.J. Consumption of the putative chemopreventive agent curcumin by cancer patients: Assessment of curcumin levels in the colorectum and their pharmacodynamic consequences. Cancer Epidemiol. Biomark. Prev. 2005, 14, 120–125. [Google Scholar] [CrossRef]
- Garcea, G.; Jones, D.J.; Singh, R.; Dennison, A.R.; Farmer, P.B.; Sharma, R.A.; Steward, W.P.; Gescher, A.J.; Berry, D.P. Detection of curcumin and its metabolites in hepatic tissue and portal blood of patients following oral administration. Br. J. Cancer 2004, 90, 1011–1015. [Google Scholar] [CrossRef]
- Sharma, R.A.; Euden, S.A.; Platton, S.L.; Cooke, D.N.; Shafayat, A.; Hewitt, H.R.; Marczylo, T.H.; Morgan, B.; Hemingway, D.; Plummer, S.M.; et al. Phase I clinical trial of oral curcumin: Biomarkers of systemic activity and compliance. Clin. Cancer Res. 2004, 10, 6847–6854. [Google Scholar] [CrossRef]
- Babaei, E.; Sadeghizadeh, M.; Hassan, Z.M.; Feizi, M.A.; Najafi, F.; Hashemi, S.M. Dendrosomal curcumin significantly suppresses cancer cell proliferation in vitro and in vivo. Int. Immunopharmacol. 2012, 12, 226–234. [Google Scholar] [CrossRef]
- Yallapu, M.M.; Khan, S.; Maher, D.M.; Ebeling, M.C.; Sundram, V.; Chauhan, N.; Ganju, A.; Balakrishna, S.; Gupta, B.K.; Zafar, N.; et al. Anti-cancer activity of curcumin loaded nanoparticles in prostate cancer. Biomaterials 2014, 35, 8635–8648. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Zhang, Y.; Han, S.; Chen, Y.; Li, B.; Liao, M.; Chen, W.; Deng, X.; Zhao, J.; Huang, B. Synthesis and in vitro/in vivo anti-cancer evaluation of curcumin-loaded chitosan/poly(butyl cyanoacrylate) nanoparticles. Int. J. Pharm. 2010, 400, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Man, S.; Qiu, H.; Liu, Z.; Zhang, M.; Ma, L.; Gao, W. Curcumin-cyclodextrin complexes enhanced the anti-cancer effects of curcumin. Environ. Toxicol. Pharmacol. 2016, 48, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Senanayake, T.H.; Bohling, A.; Vinogradov, S.V. Targeted nanogel conjugate for improved stability and cellular permeability of curcumin: Synthesis, pharmacokinetics, and tumor growth inhibition. Mol. Pharm. 2014, 11, 3112–3122. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Gong, Z.; Zhou, S.; Yang, S.; Wang, D.; Chen, X.; Wu, J.; Liu, L.; Zhong, S.; Zhao, J.; et al. Liposomal Curcumin Targeting Endometrial Cancer Through the NF-κB Pathway. Cell Physiol. Biochem. 2018, 48, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, X.; Chen, H.; Wu, Q.; Zhao, Q.; Fu, D.; Liu, Q.; Wang, Y. The Antitumour Activity of a Curcumin and Piperine Loaded iRGD-Modified Liposome: In Vitro and In Vivo Evaluation. Molecules 2023, 28, 6532. [Google Scholar] [CrossRef] [PubMed]
- Baldassari, S.; Balboni, A.; Drava, G.; Donghia, D.; Canepa, P.; Ailuno, G.; Caviglioli, G. Phytochemicals and Cancer Treatment: Cell-Derived and Biomimetic Vesicles as Promising Carriers. Pharmaceutics 2023, 15, 1445. [Google Scholar] [CrossRef] [PubMed]
- Jean, S.; Kiger’, A.A. Classes of phosphoinositide 3-kinases at a glance. J. Cell Sci. 2014, 127, 923–928. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Sun, M.M.; Zhang, G.G.; Yang, J.; Chen, K.S.; Xu, W.W.; Li, B. Targeting PI3K/Akt signal transduction for cancer therapy. Signal Transduct. Target. Ther. 2021, 6, 425. [Google Scholar] [CrossRef]
- Hanker, A.B.; Kaklamani, V.; Arteaga, C.L. Challenges for the clinical development of PI3K inhibitors: Strategies to improve their impact in solid tumors. Cancer Disco. 2019, 9, 482–491. [Google Scholar] [CrossRef]
- Kandoth, C.; McLellan, M.; Vandin, F.; Ye, K.; Niu, B.; Lu, C.; Xie, M.; Zhang, Q.; McMichael, J.F.; Wyczalkowski, M.A. Mutational landscape and significance across 12 major cancer types. Nature 2013, 502, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Yi, K.H.; Lauring, J. Recurrent AKT mutations in human cancers: Functional consequences and effects on drug sensitivity. Oncotarget 2016, 7, 4241–4251. [Google Scholar] [CrossRef]
- Rodgers, S.J.; Ferguson, D.T.; Mitchell, C.A.; Ooms, L.M. Regulation of PI3K effector signalling in cancer by the phosphoinositide phosphatases. Biosci. Rep. 2017, 37, BSR20160432. [Google Scholar] [CrossRef] [PubMed]
- Mundi, P.S.; Sachdev, J.; McCourt, C.; Kalinsky, K. AKT in cancer: New molecular insights and advances in drug development. Br. J. Clin. Pharmacol. 2016, 82, 943–956. [Google Scholar] [CrossRef] [PubMed]
- Kalhori, M.R.; Irani, S.; Soleimani, M.; Arefian, E.; Kouhkan, F. The effect of miR-579 on the PI3K/AKT pathway in human glioblastoma PTEN mutant cell lines. J. Cell Biochem. 2019, 120, 16760–16774. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, J.; Li, J.; Geng, H.; Zhou, B.; Zhang, B.; Chen, H. miR-320/ELF3 axis inhibits the progression of breast cancer via the PI3K/AKT pathway. Oncol. Lett. 2020, 19, 3239–3248. [Google Scholar] [CrossRef] [PubMed]
- Castellano, E.; Downward, J. RAS Interaction with PI3K: More Than Just Another Effector Pathway. Genes. Cancer 2011, 2, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Zoi, V.; Galani, V.; Lianos, G.D.; Voulgaris, S.; Kyritsis, A.P.; Alexiou, G.A. The Role of Curcumin in Cancer Treatment. Biomedicines 2021, 9, 1086. [Google Scholar] [CrossRef] [PubMed]
- Beevers, C.S.; Zhou, H.; Huang, S. Hitting the golden TORget: Curcumin’s effects on mTOR signaling. Anticancer. Agents Med. Chem. 2013, 13, 988–994. [Google Scholar] [CrossRef]
- Khan, K.; Quispe, C.; Javed, Z.; Iqbal, M.J.; Sadia, H.; Raza, S.; Irshad, A.; Salehi, B.; Reiner, Z.; Sharifi-Rad, J. Resveratrol, curcumin, paclitaxel and miRNAs mediated regulation of PI3K/Akt/mTOR pathway: Go four better to treat bladder cancer. Cancer Cell Int. 2020, 20, 560. [Google Scholar] [CrossRef]
- Tamaddoni, A.; Mohammadi, E.; Sedaghat, F.; Qujeq, D.; As’Habi, A. The anticancer effects of curcumin via targeting the mammalian target of rapamycin complex 1 (mTORC1) signaling pathway. Pharmacol. Res. 2020, 156, 104798. [Google Scholar] [CrossRef]
- Sun, Z.J.; Chen, G.; Zhang, W.; Hu, X.; Liu, Y.; Zhou, Q.; Zhu, L.X.; Zhao, Y.F. Curcumin dually inhibits both mammalian target of rapamycin and nuclear factor-κB pathways through a crossed phosphatidylinositol 3-kinase/Akt/IκB kinase complex signaling axis in adenoid cystic carcinoma. Mol. Pharmacol. 2011, 79, 106–118. [Google Scholar] [CrossRef]
- Xiao, K.; Jiang, J.; Guan, C.; Dong, C.; Wang, G.; Bai, L.; Sun, J.; Hu, C.; Bai, C. Curcumin induces autophagy via activating the AMPK signaling pathway in lung adenocarcinoma cells. J. Pharmacol. Sci. 2013, 123, 102–109. [Google Scholar] [CrossRef]
- Li, W.; Saud, S.M.; Young, M.R.; Chen, G.; Hua, B. Targeting AMPK for cancer prevention and treatment. Oncotarget 2015, 6, 7365–7378. [Google Scholar] [CrossRef] [PubMed]
- Paquette, M.; El-Houjeiri, L.; Pause, A. mTOR Pathways in Cancer and Autophagy. Cancers 2018, 10, 18. [Google Scholar] [CrossRef]
- Liu, F.; Gao, S.; Yang, Y.; Zhao, X.; Fan, Y.; Ma, W.; Yang, D.; Yang, A.; Yu, Y. Antitumor activity of curcumin by modulation of apoptosis and autophagy in human lung cancer A549 cells through inhibiting PI3K/Akt/mTOR pathway. Oncol. Rep. 2018, 39, 1523–1531. [Google Scholar] [CrossRef] [PubMed]
- Maiti, P.; Scott, J.; Sengupta, D.; Al-Gharaibeh, A.; Dunbar, G.L. Curcumin and Solid Lipid Curcumin Particles Induce Autophagy, but Inhibit Mitophagy and the PI3K-Akt/mTOR Pathway in Cultured Glioblastoma Cells. Int. J. Mol. Sci. 2019, 20, 399. [Google Scholar] [CrossRef] [PubMed]
- He, P.; He, Y.; Ma, J.; Liu, Y.; Liu, C.; Baoping, Y.; Dong, W. Thymoquinone induces apoptosis and protective autophagy in gastric cancer cells by inhibiting the PI3K/Akt/mTOR pathway. Phytother. Res. 2023, 37, 3467–3480. [Google Scholar] [CrossRef]
- Yang, L.; Liu, Y.N.; Gu, Y.; Guo, Q. Deltonin enhances gastric carcinoma cell apoptosis and chemosensitivity to cisplatin via inhibiting PI3K/AKT/mTOR and MAPK signaling. World J. Gastrointest. Oncol. 2023, 15, 1739–1755. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, Y.; Zhang, W.; Wang, Y.; Jao, Y.; Liu, J.; Zhang, M.; He, H. AMPK/mTOR/p70S6K axis prevents apoptosis of Porphyromonas gingivalis-infected gingival epithelial cells via BadSer136 phosphorylation. Apoptosis 2023, 28, 1012–1023. [Google Scholar] [CrossRef]
- Kuttikrishnan, S.; Siveen, K.S.; Prabhu, K.S.; Khan, A.Q.; Ahmed, E.I.; Akhtar, S.; Ali, T.A.; Merhi, M.; Dermime, S.; Steinhoff, M.; et al. Curcumin Induces Apoptotic Cell Death via Inhibition of PI3-Kinase/AKT Pathway in B-Precursor Acute Lymphoblastic Leukemia. Front. Oncol. 2019, 9, 484. [Google Scholar] [CrossRef] [PubMed]
- Kizhakkayil, J.; Thayyullathil, F.; Chathoth, S.; Hago, A.; Patel, M.; Galadari, S. Modulation of curcumin-induced Akt phosphorylation and apoptosis by PI3K inhibitor in MCF-7 cells. Biochem. Biophys. Res. Commun. 2010, 394, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Xu, Y.; Meng, L.; Huang, L.; Sun, H. Curcumin inhibits proliferation and promotes apoptosis of breast cancer cells. Exp. Ther. Med. 2018, 16, 1266–1272. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.W.; Chien, S.Y.; Kuo, S.J.; Tseng, L.M.; Lin, H.Y.; Chi, C.W.; Chen, D.R. The Potential Utility of Curcumin in the Treatment of HER-2-Overexpressed Breast Cancer: An In Vitro and In Vivo Comparison Study with Herceptin. Evid. Based Complement. Alternat. Med. 2012, 2012, 486568. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hang, Y.; Liu, J.; Hou, Y.; Wang, N.; Wang, M. Anticancer effect of curcumin inhibits cell growth through miR-21/PTEN/Akt pathway in breast cancer cell. Oncol. Lett. 2017, 13, 4825–4831. [Google Scholar] [CrossRef]
- Jiang, Q.G.; Li, T.Y.; Liu, D.N.; Zhang, H.T. PI3K/Akt pathway involving into apoptosis and invasion in human colon cancer cells LoVo. Mol. Biol. Rep. 2014, 41, 3359–3367. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.M.; Gulhati, P.; Arrieta, I.; Wang, X.; Uchida, T.; Gao, T.; Evers, B.M. Curcumin inhibits proliferation of colorectal carcinoma by modulating Akt/mTOR signaling. Anticancer Res. 2009, 29, 3185–3190, Erratum in: Anticancer Res. 2009, 29, 4315. [Google Scholar] [PubMed]
- Zanotto-Filho, A.; Braganhol, E.; Edelweiss, M.I.; Behr, G.A.; Zanin, R.; Schröder, R.; Simões-Pires, A.; Battastini, A.M.O.; Moreira, J.C.F. The curry spice curcumin selectively inhibits cancer cells growth in vitro and in preclinical model of glioblastoma. J. Nutr. Biochem. 2012, 23, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, F.; Liao, W.; Yu, L.; Hu, Z.; Li, M.; Xia, H. Curcumin suppresses glioblastoma cell proliferation by p-AKT/mTOR pathway and increases the PTEN expression. Arch. Biochem. Biophys. 2020, 689, 108412. [Google Scholar] [CrossRef]
- Wang, X.; Deng, J.; Yuan, J.; Tang, X.; Wang, Y.; Chen, H.; Liu, Y.; Zhou, L. Curcumin exerts its tumor suppressive function via inhibition of NEDD4 oncoprotein in glioma cancer cells. Int. J. Oncol. 2017, 51, 467–477. [Google Scholar] [CrossRef]
- Hussain, A.R.; Al-Rasheed, M.; Manogaran, P.S.; Al-Hussein, K.A.; Platanias, L.C.; Al Kuraya, K.; Uddin, S. Curcumin induces apoptosis via inhibition of PI3’-kinase/AKT pathway in acute T cell leukemias. Apoptosis 2006, 11, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Li, Y.; Shan, Q.; He, G.; Lin, J.; Gong, Y. Curcumin potentiates the anti-leukemia effects of imatinib by downregulation of the AKT/mTOR pathway and BCR/ABL gene expression in Ph+ acute lymphoblastic leukemia. Int. J. Biochem. Cell Biol. 2015, 65, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Ning, Y.; Zeng, G.; Zhou, C.; Ding, X. Curcumin promotes cell cycle arrest and apoptosis of acute myeloid leukemia cells by inactivating AKT. Oncol. Rep. 2021, 45, 11. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Feng, T.; Liu, X.; Liu, Q. Curcumin inhibits migration and invasion of non-small cell lung cancer cells through up-regulation of miR-206 and suppression of PI3K/AKT/mTOR signaling pathway. Acta Pharm. 2020, 70, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Qiao, F.; Wang, Y.; Xu, Y.; Shang, Y. Curcumin inhibits cell proliferation and induces apoptosis of human non-small cell lung cancer cells through the upregulation of miR-192-5p and suppression of PI3K/Akt signaling pathway. Oncol. Rep. 2015, 34, 2782–2789. [Google Scholar] [CrossRef] [PubMed]
- Endo, H.; Inoue, I.; Masunaka, K.; Tanaka, M.; Yano, M. Curcumin induces apoptosis in lung cancer cells by 14-3-3 protein-mediated activation of Bad. Biosci. Biotechnol. Biochem. 2020, 84, 2440–2447. [Google Scholar] [CrossRef]
- Shankar, S.; Srivastava, R.K. Involvement of Bcl-2 family members, phosphatidylinositol 3’-kinase/AKT and mitochondrial p53 in curcumin (diferulolylmethane)-induced apoptosis in prostate cancer. Int. J. Oncol. 2007, 30, 905–918. [Google Scholar] [CrossRef]
- Yu, S.; Shen, G.; Khor, T.O.; Kim, J.H.; Kong, A.N. Curcumin inhibits Akt/mammalian target of rapamycin signaling through protein phosphatase-dependent mechanism. Mol. Cancer Ther. 2008, 7, 2609–2620. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Qin, J.; Liu, W. Curcumin inhibits the invasion of thyroid cancer cells via down-regulation of PI3K/Akt signaling pathway. Gene 2014, 546, 226–232. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, S.; Cheng, X.; Wu, J.; Wu, L.; Wang, Y.; Wang, X.; Bao, J.; Yu, H. Curcumin induces autophagic cell death in human thyroid cancer cells. Toxicol. In Vitro 2022, 78, 105254. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, S.; Cheng, X.; Wu, J.; Wang, X.; Wu, L.; Yu, H.; Bao, J. Curcumin enhances the membrane trafficking of the sodium iodide symporter and augments radioiodine uptake in dedifferentiated thyroid cancer cells via suppression of the PI3K-AKT signaling pathway. Food Funct. 2021, 12, 8260–8273. [Google Scholar] [CrossRef]
- Pellerino, A.; Caccese, M.; Padovan, M.; Cerretti, G.; Lombardi, G. Epidemiology, risk factors, and prognostic factors of gliomas. Clin. Transl. Imaging 2022, 10, 467–475. [Google Scholar] [CrossRef]
- Wu, W.; Klockow, J.L.; Zhang, M.; Lafortune, F.; Chang, E.; Jin, L.; Wu, Y.; Daldrup-Link, H.E. Glioblastoma multiforme (GBM): An overview of current therapies and mechanisms of resistance. Pharmacol. Res. 2021, 171, 105780. [Google Scholar] [CrossRef]
- Kaba, S.E.; Kyritsis, A.P. Recognition and management of gliomas. Drugs 1997, 53, 235–244. [Google Scholar] [CrossRef]
- Alexiou, G.A.; Vartholomatos, E.I.; Tsamis, K.; Peponi, E.; Markopoulos, G.A.; Papathanasopoulou, V.; Tasiou, I.; Ragos, V.; Tsekeris, P.; Kyritsis, A.P.; et al. Combination treatment for glioblastoma with temozolomide, DFMO and radiation. J. BUON 2019, 24, 397–404. [Google Scholar] [PubMed]
- Zoi, V.; Galani, V.; Vartholomatos, E.; Zacharopoulou, N.; Tsoumeleka, E.; Gkizas, G.; Bozios, G.; Tsekeris, P.; Chousidis, I.; Leonardos, I.; et al. Curcumin and Radiotherapy Exert Synergistic Anti-Glioma Effect In Vitro. Biomedicines 2021, 9, 1562. [Google Scholar] [CrossRef] [PubMed]
- Langhans, J.; Schneele, L.; Trenkler, N.; von Bandemer, H.; Nonnenmacher, L.; Karpel-Massler, G.; Siegelin, M.; Zhou, S.; Halatsch, M.; Debatin, K.; et al. The effects of PI3K-mediated signalling on glioblastoma cell behaviour. Oncogenesis 2017, 6, 398. [Google Scholar] [CrossRef]
- Cantidio, F.S.; Gil, G.O.B.; Queiroz, I.N.; Regalin, M. Glioblastoma-treatment and obstacles. Rep. Pract. Oncol. Radiother. 2022, 27, 744–753. [Google Scholar] [CrossRef]
- Puduvalli, V.K.; Sampath, D.; Bruner, J.M.; Nangia, J.; Xu, R.; Kyritsis, A.P. TRAIL-induced apoptosis in gliomas is enhanced by Akt-inhibition and is independent of JNK activation. Apoptosis 2005, 10, 233–243. [Google Scholar] [CrossRef]
- Yang, J.M.; Schiapparelli, P.; Nguyen, H.N.; Igarashi, A.; Zhang, Q.; Abbadi, S.; Amzel, L.M.; Sesaki, H.; Quiñones-Hinojosa, A.; Iijima, M. Characterization of PTEN mutations in brain cancer reveals that pten mono-ubiquitination promotes protein stability and nuclear localization. Oncogene 2017, 36, 3673–3685. [Google Scholar] [CrossRef]
- Han, F.; Hu, R.; Yang, H.; Liu, J.; Sui, J.; Xiang, X.; Wang, F.; Chu, L.; Song, S. PTEN gene mutations correlate to poor prognosis in glioma patients: A meta-analysis. Onco Targets Ther. 2016, 9, 3485–3492. [Google Scholar] [PubMed]
- Goussia, A.C.; Agnantis, N.J.; Rao, J.S.; Kyritsis, A.P. Cytogenetic and molecular abnormalities in astrocytic gliomas (Review). Oncol. Rep. 2000, 7, 401–412. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research Network. Comprehensive genomic characterization defines human glioblastoma genes and core pathways. Nature 2008, 455, 1061–1068, Erratum in Nature 2013, 494, 506. [Google Scholar]
- Sunayama, J.; Matsuda, K.I.; Sato, A.; Tachibana, K.; Suzuki, K.; Narita, Y.; Shibui, S.; Sakurada, K.; Kayama, T.; Tomiyama, A.; et al. Crosstalk between the PI3K/mTOR and MEK/ERK pathways involved in the maintenance of self-renewal and tumorigenicity of glioblastoma stem-like cells. Stem Cells 2010, 28, 1930–1939. [Google Scholar] [CrossRef] [PubMed]
- Yun, C.W.; Jeon, J.; Go, G.; Lee, J.H.; Lee, S.H. The Dual Role of Autophagy in Cancer Development and a Therapeutic Strategy for Cancer by Targeting Autophagy. Int. J. Mol. Sci. 2020, 22, 179. [Google Scholar] [CrossRef] [PubMed]
- Colella, B.; Faienza, F.; Di Bartolomeo, S. EMT regulation by autophagy: A new perspective in glioblastoma biology. Cancers 2019, 11, 312. [Google Scholar] [CrossRef]
- Yang, K.; Niu, L.; Bai, Y.; Le, W. Glioblastoma: Targeting the autophagy in tumorigenesis. Brain Res. Bull. 2019, 153, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Aoki, H.; Takada, Y.; Kondo, S.; Sawaya, R.; Aggarwal, B.B.; Kondo, Y. Evidence that curcumin suppresses the growth of malignant gliomas in vitro and in vivo through induction of autophagy: Role of Akt and extracellular signal-regulated kinase signaling pathways. Mol. Pharmacol. 2007, 72, 29–39. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, Y.; Sun, X.; He, X.; Wang, M.; Wang, Z.; Wang, Q.; Zhu, R.; Wang, S. Curcumin-Loaded Layered Double Hydroxide Nanoparticles-Induced Autophagy for Reducing Glioma Cell Migration and Invasion. J. Biomed. Nanotechnol. 2016, 12, 2051–2062. [Google Scholar] [CrossRef]
- Ryskalin, L.; Gaglione, A.; Limanaqi, F.; Biagioni, F.; Familiari, P.; Frati, A.; Esposito, V.; Fornai, F. The Autophagy Status of Cancer Stem Cells in Gliobastoma Multiforme: From Cancer Promotion to Therapeutic Strategies. Int. J. Mol. Sci. 2019, 20, 3824. [Google Scholar] [CrossRef]
- Ryskalin, L.; Biagioni, F.; Busceti, C.L.; Lazzeri, G.; Frati, A.; Fornai, F. The Multi-Faceted Effect of Curcumin in Glioblastoma from Rescuing Cell Clearance to Autophagy-Independent Effects. Molecules 2020, 25, 4839. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Armas, M.; Corona, J.C.; Escobar, M.L.; Torres, L.; Ordóñez-Romero, F.; Hernández-Hernández, A.; Arenas-Huertero, F. Paraptosis in human glioblastoma cell line induced by curcumin. Toxicol. In Vitro 2018, 51, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.C.; Lin, Y.F.; Huang, M.Y.; Zhang, X.L.; Wang, P.; Huang, M.Q.; Lu, J.J. Paraptosis: A non-classical paradigm of cell death for cancer therapy. Acta Pharmacol. Sin. 2024, 45, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Alduais, Y.; Zhang, H.; Fan, F.; Chen, J.; Chen, B. Non-small cell lung cancer (NSCLC): A review of risk factors, diagnosis, and treatment. Medicine 2023, 102, e32899. [Google Scholar] [CrossRef] [PubMed]
- Abu Rous, F.; Singhi, E.K.; Sridhar, A.; Faisal, M.S.; Desai, A. Lung Cancer Treatment Advances in 2022. Cancer Investig. 2023, 41, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yan, B.; He, S. Advances and challenges in the treatment of lung cancer. Biomed Pharmacother. 2023, 169, 115891. [Google Scholar] [CrossRef] [PubMed]
- Kastamoulas, M.; Chondrogiannis, G.; Kanavaros, P.; Vartholomatos, G.; Bai, M.; Briasoulis, E.; Arvanitis, D.; Galani, V. Cytokine effects on cell survival and death of A549 lung carcinoma cells. Cytokine 2013, 61, 816–825. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.C. Targeting the PI3K/Akt/mTOR pathway in non-small cell lung cancer (NSCLC). Thorac. Cancer 2020, 11, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Liu, J.; Yang, C.; Ma, J.; Chen, X.; Liu, D.; Zhou, Y.; Zhou, W.; Lin, Y.; Yuan, X. Curcumin and Its Analogs in Non-Small Cell Lung Cancer Treatment: Challenges and Expectations. Biomolecules 2022, 12, 1636. [Google Scholar] [CrossRef]
- Wang, A.; Wang, J.; Zhang, S.; Zhang, H.; Xu, Z.; Li, X. Curcumin inhibits the development of non-small cell lung cancer by inhibiting autophagy and apoptosis. Exp. Ther. Med. 2017, 14, 5075–5080. [Google Scholar] [CrossRef]
- Zhang, W.; Bai, W.; Zhang, W. MiR-21 suppresses the anticancer activities of curcumin by targeting PTEN gene in human non-small cell lung cancer A549 cells. Clin. Transl. Oncol. 2014, 16, 708–713. [Google Scholar] [CrossRef] [PubMed]
- Neal, C.L.; Xu, J.; Li, P.; Mori, S.; Yang, J.; Neal, N.N.; Zhou, X.; Wyszomierski, S.L.; Yu, D. Overexpression of 14-3-3ζ in cancer cells activates PI3K via binding the p85 regulatory subunit. Oncogene 2012, 31, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Cui, L.; Zeng, Y.; Song, W.; Gaur, U.; Yang, M. 14-3-3 Proteins Are on the Crossroads of Cancer, Aging, and Age-Related Neurodegenerative Disease. Int. J. Mol. Sci. 2019, 20, 3518. [Google Scholar] [CrossRef] [PubMed]
- Jiao, D.; Wang, J.; Lu, W.; Tang, X.; Chen, J.; Mou, H.; Chen, Q.Y. Curcumin inhibited HGF-induced EMT and angiogenesis through regulating c-Met dependent PI3K/Akt/mTOR signaling pathways in lung cancer. Mol. Ther. Oncolytics 2016, 3, 16018. [Google Scholar] [CrossRef]
- Zhang, Y.; Xia, M.; Jin, K.; Wang, S.; Wei, H.; Fan, C.; Wu, Y.; Li, X.; Li, X.; Li, G.; et al. Function of the c-Met receptor tyrosine kinase in carcinogenesis and associated therapeutic opportunities. Mol. Cancer 2018, 17, 45. [Google Scholar] [CrossRef]
- Orrantia-Borunda, E.; Anchondo-Nuñez, P.; Acuña-Aguilar, L.E.; Gómez-Valles, F.O.; Ramírez-Valdespino, C.A.; Mayrovitz, H.N. Subtypes of Breast Cancer. In Breast Cancer [Internet]; Mayrovitz, H.N., Ed.; Exon Publications: Brisbane, Australia, 2022; Chapter 3. [Google Scholar]
- Trayes, K.P.; Cokenakes, S.E.H. Breast Cancer Treatment. Am. Fam. Physician 2021, 104, 171–178. [Google Scholar] [PubMed]
- Moo, T.A.; Sanford, R.; Dang, C.; Morrow, M. Overview of Breast Cancer Therapy. PET Clin. 2018, 13, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Miricescu, D.; Totan, A.; Stanescu-Spinu, I.I.; Badoiu, S.C.; Stefani, C.; Greabu, M. PI3K/AKT/mTOR Signaling Pathway in Breast Cancer: From Molecular Landscape to Clinical Aspects. Int. J. Mol. Sci. 2020, 22, 173. [Google Scholar] [CrossRef] [PubMed]
- García-Becerra, R.; Santos-Martínez, N.; Díaz, L.; Camacho, J. Mechanisms of resistance to endocrine therapy in breast cancer: Focus on signaling pathways, miRNAs and genetically based resistance. Int. J. Mol. Sci. 2013, 14, 108–145. [Google Scholar] [CrossRef]
- Jia, T.; Zhang, L.; Duan, Y.; Zhang, M.; Wang, G.; Zhang, J.; Zhao, Z. The differential susceptibilities of MCF-7 and MDA-MB-231 cells to the cytotoxic effects of curcumin are associated with the PI3K/Akt-SKP2-Cip/Kips pathway. Cancer Cell Int. 2014, 14, 126. [Google Scholar] [CrossRef]
- He, C.; Dong, X.; Zhai, B.; Jiang, X.; Dong, D.; Li, B.; Jiang, H.; Xu, S.; Sun, X. MiR-21 mediates sorafenib resistance of hepatocellular carcinoma cells by inhibiting autophagy via the PTEN/Akt pathway. Oncotarget 2015, 6, 28867–28881. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xie, W.; Xie, C.; Huang, C.; Zhu, J.; Liang, Z.; Deng, F.; Zhu, M.; Zhu, W.; Wu, R.; et al. Curcumin modulates miR-19/PTEN/AKT/p53 axis to suppress bisphenol A-induced MCF-7 breast cancer cell proliferation. Phytother. Res. 2014, 28, 1553–1560. [Google Scholar] [CrossRef]
- Rafikova, G.; Gilyazova, I.; Enikeeva, K.; Pavlov, V.; Kzhyshkowska, J. Prostate Cancer: Genetics, Epigenetics and the Need for Immunological Biomarkers. Int. J. Mol. Sci. 2023, 24, 12797. [Google Scholar] [CrossRef]
- Sekhoacha, M.; Riet, K.; Motloung, P.; Gumenku, L.; Adegoke, A.; Mashele, S. Prostate Cancer Review: Genetics, Diagnosis, Treatment Options, and Alternative Approaches. Molecules 2022, 27, 5730. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, M.; Taheriazam, A.; Daneii, P.; Hassanpour, A.; Kakavand, A.; Rezaei, S.; Hejazi, E.S.; Aboutalebi, M.; Gholamrezaie, H.; Saebfar, H.; et al. Targeting PI3K/Akt signaling in prostate cancer therapy. J. Cell Commun. Signal. 2023, 17, 423–443. [Google Scholar] [CrossRef]
- Shorning, B.Y.; Dass, M.S.; Smalley, M.J.; Pearson, H.B. The PI3K-AKT-mTOR Pathway and Prostate Cancer: At the Crossroads of AR, MAPK, and WNT Signaling. Int. J. Mol. Sci. 2020, 21, 4507. [Google Scholar] [CrossRef] [PubMed]
- Ferraldeschi, R.; Welti, J.; Luo, J.; Attard, G.; de Bono, J.S. Targeting the androgen receptor pathway in castration-resistant prostate cancer: Progresses and prospects. Oncogene 2015, 34, 1745–1757. [Google Scholar] [CrossRef] [PubMed]
- Katta, S.; Srivastava, A.; Thangapazham, R.L.; Rosner, I.L.; Cullen, J.; Li, H.; Sharad, S. Curcumin-Gene Expression Response in Hormone Dependent and Independent Metastatic Prostate Cancer Cells. Int. J. Mol. Sci. 2019, 20, 4891. [Google Scholar] [CrossRef]
- Duan, B.; Zhao, Y.; Bai, J.; Wang, J.; Duan, X.; Luo, X.; Zhang, R.; Pu, Y.; Kou, M.; Lei, J.; et al. Colorectal Cancer: An Overview. In Gastrointestinal Cancers [Internet]; Morgado-Diaz, J.A., Ed.; Exon Publications: Brisbane, Australia, 2022; Chapter 1. [Google Scholar]
- Hossain, M.S.; Karuniawati, H.; Jairoun, A.A.; Urbi, Z.; Ooi, J.; John, A.; Lim, Y.C.; Kibria, K.M.K.; Mohiuddin, A.K.M.; Ming, L.C.; et al. Colorectal Cancer: A Review of Carcinogenesis, Global Epidemiology, Current Challenges, Risk Factors, Preventive and Treatment Strategies. Cancers 2022, 14, 1732. [Google Scholar] [CrossRef]
- Danielsen, S.A.; Eide, P.W.; Nesbakken, A.; Guren, T.; Leithe, E.; Lothe, R.A. Portrait of the PI3K/AKT pathway in colorectal cancer. Biochim. Biophys. Acta 2015, 1855, 104–121. [Google Scholar] [CrossRef]
- Narayanankutty, A. PI3K/Akt/mTOR Pathway as a Therapeutic Target for Colorectal Cancer: A Review of Preclinical and Clinical Evidence. Curr. Drug Targets 2019, 20, 1217–1226. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Sun, X.; Wang, R.; Guo, Y.; Xu, M. Effects of curcumin on 5-fluorouracil resistance of colon cancer cells through the PI3K/AKT/mTOR pathway via MACC1. Eur. J. Int. Med. 2022, 56, 102202. [Google Scholar] [CrossRef]
- Chen, L.; Li, W.F.; Wang, H.X.; Zhao, H.N.; Tang, J.J.; Wu, C.J.; Lu, L.T.; Liao, W.Q.; Lu, X.C. Curcumin cytotoxicity is enhanced by PTEN disruption in colorectal cancer cells. World J. Gastroenterol. 2013, 19, 6814–6824. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Anastasopoulou, C.; Chandran, C.; Cassaro, S. Thyroid Cancer. In StatPearls [Internet]; [Updated 2023 May 1]; StatPearls Publishing: Treasure Island, FL, USA, 2024. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK459299/ (accessed on 1 January 2024).
- Nabhan, F.; Dedhia, P.H.; Ringel, M.D. Thyroid cancer, recent advances in diagnosis and therapy. Int. J. Cancer 2021, 149, 984–992. [Google Scholar] [CrossRef] [PubMed]
- Regalbuto, C.; Frasca, F.; Pellegriti, G.; Malandrino, P.; Marturano, I.; Di Carlo, I.; Pezzino, V. Update on thyroid cancer treatment. Future Oncol. 2012, 8, 1331–1348. [Google Scholar] [CrossRef]
- Faustino, A.; Couto, J.P.; Pópulo, H.; Rocha, A.S.; Pardal, F.; Cameselle-Teijeiro, J.M.; Lopes, J.M.; Sobrinho-Simões, M.; Soares, P. mTOR pathway overactivation in BRAF mutated papillary thyroid carcinoma. J. Clin. Endocrinol. Metab. 2012, 97, 1139–1149. [Google Scholar] [CrossRef] [PubMed]
- Nozhat, Z.; Hedayati, M. PI3K/AKT Pathway and Its Mediators in Thyroid Carcinomas. Mol. Diagn. Ther. 2016, 20, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yu, J.; Xie, R.; Chen, W.; Lv, Y. Combinatorial anticancer effects of curcumin and sorafenib towards thyroid cancer cells via PI3K/Akt and ERK pathways. Nat. Prod. Res. 2016, 30, 1858–1861. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Chan, S.C.; Ngai, C.H.; Lok, V.; Zhang, L.; Lucero-Prisno, D.E.; Xu, W.; Zheng, Z.J.; Elcarte, E.; Withers, M.; et al. Disease Burden, Risk Factors, and Trends of Leukaemia: A Global Analysis. Front. Oncol. 2022, 12, 904292. [Google Scholar] [CrossRef]
- Malard, F.; Mohty, M. Acute lymphoblastic leukaemia. Lancet 2020, 395, 1146–1162. [Google Scholar] [CrossRef]
- Shiraz, P.; Payne, K.J.; Muffly, L. The Current Genomic and Molecular Landscape of Philadelphia-like Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2020, 21, 2193. [Google Scholar] [CrossRef]
- Wang, L.; Fortney, J.E.; Gibson, L.F. Stromal cell protection of B-lineage acute lymphoblastic leukemic cells during chemotherapy requires active Akt. Leuk. Res. 2004, 28, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, V.E.; Nichols, C.; Kim, H.N.; Gang, E.J.; Kim, Y.M. Targeting PI3K Signaling in Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2019, 20, 412. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Geng, Q.R.; Wang, L.; Lu, Y. Curcumin potentiates antitumor activity of L-asparaginase via inhibition of the AKT signaling pathway in acute lymphoblastic leukemia. Leuk. Lymphoma 2012, 53, 1376–1382. [Google Scholar] [CrossRef] [PubMed]
- Hus, I.; Puła, B.; Robak, T. PI3K Inhibitors for the Treatment of Chronic Lymphocytic Leukemia: Current Status and Future Perspectives. Cancers 2022, 14, 1571. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.K.; Kay, N.E.; Secreto, C.R.; Shanafelt, T.D. Curcumin inhibits prosurvival pathways in chronic lymphocytic leukemia B cells and may overcome their stromal protection in combination with EGCG. Clin. Cancer Res. 2009, 15, 1250–1258. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lei, H.; Wei, X.; Wei, Y.; Feng, R.; Guo, X. Curcumin Enhances Ibrutinib Induced Killing Effect Against TP53-Mutated Chronic Lymphocytic Leukemia Cells In Vitro. Blood 2023, 142, 5792. [Google Scholar] [CrossRef]
- Nepstad, I.; Hatfield, K.J.; Grønningsæter, I.S.; Reikvam, H. The PI3K-Akt-mTOR Signaling Pathway in Human Acute Myeloid Leukemia (AML) Cells. Int. J. Mol. Sci. 2020, 21, 2907. [Google Scholar] [CrossRef]
- Wu, J.C.; Lai, C.S.; Badmaev, V.; Nagabhushanam, K.; Ho, C.T.; Pan, M.H. Tetrahydrocurcumin, a major metabolite of curcumin, induced autophagic cell death through coordinative modulation of PI3K/Akt-mTOR and MAPK signaling pathways in human leukemia HL-60 cells. Mol. Nutr. Food Res. 2011, 55, 1646–1654. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cancer | No. of Patients | Formulation, Dosage, and Delivery Scheme | Results | Ref. |
|---|---|---|---|---|
| Pancreatic or biliary tract cancer | 16 |
| Peak plasma curcumin levels 2 h after Theracurmin administration were 324 ng/mL at level 1 and 440 ng/mL at level 2 | [26] |
| Colorectal cancer | 12 |
| Tissue levels of curcumin 6 h after the last dosage were 0.9 ± 0.4, 6.7 ± 1.6, and 7.7 ± 1.8 for dose levels 1, 2, and 3, respectively | [27] |
| Hepatic metastatic disease from primary colorectal cancer | 12 |
| No quantifiable levels of curcumin in plasma of patients | [28] |
| Adenocarcinoma of the colon or rectum | 15 |
|
| [21] |
| Adenocarcinoma of the colon or rectum | 15 |
|
| [29] |
| Pancreatic cancer | 25 |
|
| [20] |
| Strategy | Curcumin Concentration | Delivery Method | In Vivo Anti-Cancer Results | % Success | Ref. |
|---|---|---|---|---|---|
| Dendrosomal curcumin | 12.5 mg/kg body weight daily for 3 weeks | Intraperitoneal injection | Reduction in tumor size of BALB/c tumor-bearing mice, increase in survival |
| [30] |
| Curcumin-loaded PLGA nanospheres | 25 μg PLGA-CUR NPs once | Intratumor injection | Inhibition of tumor growth in a C4-2 xenograft mouse model | The treated group of mice with C4-2 xenograft tumors showed about 17% tumor growth compared to 38% in the control group | [31] |
| Curcumin–chitosan nanoparticles | 190 mg/kg for 4 weeks (3 times weekly) | Intravenous injection | Inhibition of tumor growth and angiogenesis in HepG2 xenograft mice | The treated group of mice showed a decrease in tumor volume 24 days after the first injection (407.5 mm3 for curcumin-PBCA nanoparticles vs. 936 mm3 for physiological saline) | [32] |
| β-cyclodextrin-curcumin complex | 100 mg/kg in 10 mL of 0.9% sodium chloride daily for 14 days | Oral administration | Reductions in tumor volume and growth rate of tumors in a mouse hepatoma H22 tumor model | The treated group of tumor-bearing mice showed a 34.64% tumor inhibition rate compared to 0% in the control group | [33] |
| Curcumin nanogel conjugate | 6 mg/kg twice every week | Intraperitoneal injection | Inhibition of tumor growth in a MiaPaCa-2 xenograft murine model | The treated group of mice showed a 15-fold decrease in the mean tumor volume on Day 49 compared to the control group | [34] |
| Liposomal curcumin (LC) | 15, 30, 50, or 80 µM of LC for 6 h | Oral administration (incubation with LC) | Suppression of tumor growth in a zebrafish transplantation tumor model of endometrial cancer | The LC-treated group of zebrafish showed decrease in tumor volume by around 2-fold compared to the control 3 days after treatment at 50 μΜ LC | [35] |
| Liposomal co-delivery of curcumin and piperine | 50 mg/kg once per day for 20 days | Intraperitoneal injection | Tumor growth inhibition and reduction in tumor volume in subcutaneous A549 tumor-bearing mice | The treated group of tumor-bearing mice showed a 41.04% tumor inhibition rate compared to the control group | [36] |
| Curcumin in bovine-milk-derived exosomes | 20 mg/kg curcumin and 80 mg/kg exosomes 3 times per week | Oral gavage | Tumor growth inhibition in human cervical cancer (CaSki) xenografts in nude mice | Treated group showed greater inhibition of tumor growth (61%) compared to that of EXOs alone (21%) | [37] |
| Cancer Type | Curcumin Treatment | Cell Line | Results and Mechanism | Year | Ref |
|---|---|---|---|---|---|
| Breast | 10, 20, 30, 40 μΜ curcumin for 3, 6, 12, and 24 h | MCF-7 MDA-MB-231 | Treatment with curcumin decreased AKT phosphorylation dose- and time-dependently | 2014 | [62] |
| 10 or 30 μΜ curcumin for 12 h | T47D, MCF7, MDA-MB-415, SK-BR-3, MDA-MB-231, MDA-MB-468, BT-20 | Treatment with curcumin inhibited the phosphorylation of Akt, mTOR, and S6 after 12 h of treatment. | 2018 | [63] | |
| 1–25 μg/mL (=2.7–67.9 μΜ) curcumin | MCF-7, MCF-10A, MDA-MB-231, BT-474, SK-BR-3-hr | Treatment with curcumin at high concentrations decreased the expression of p-AKT in BT-474 and SK-BR-3-hr | 2012 | [64] | |
| 0, 1, 2, and 5 µM curcumin for 48 h. | MCF-7 | Treatment with curcumin decreased p-AKT levels and increased PTEN levels dose-dependently | 2017 | [65] | |
| Colorectal | 40, 80, and 122 μΜ curcumin for 72 h | Human colon cancer cell line LoVo | Treatment with curcumin decreased p-AKT levels without affecting the total AKT | 2014 | [66] |
| ♦ 25 μΜ curcumin for 2 h prior to EGF stimulation ♦ 12.5, 25, and 50 μΜ curcumin for 6 h | HCT116 CRC cells | ♦ Curcumin inhibited EGF-stimulated PI3K-Akt-mTOR signaling ♦ Treatment with curcumin reduced mTOR and p-p60S6K expression dose-dependently, whereas p-AKT levels were increased, possibly through the inhibition of PHLPP | 2009 | [67] | |
| Glioblastoma | 5, 30, and 50 μΜ curcumin for 6 h | U138MG, U87, and U373 GBM cells | Treatment with curcumin decreased p-AKT levels dose-dependently | 2012 | [68] |
| ♦ 20 μΜ curcumin for 24 h ♦ 10 and 20 μΜ curcumin for 24 h | U87 and U251 GBM cells | ♦ Treatment with curcumin decreased the phosphorylation of AKT (p-AKT) and mTOR (p-mTOR) in U251 cells ♦ Treatment with 10 or 20 μΜ curcumin increased PTEN expression | 2020 | [69] | |
| 25 μΜ curcumin for 24 h | U87MG cells | Treatment with curcumin resulted in decreased levels of PI3Kp85, pPI3Kp85, total Akt, p-Akt, mTOR, and p-mTOR | 2019 | [57] | |
| 10, 15, and 20 μΜ curcumin for 48 h | SNB19 and A1207 cells | Treatment with curcumin decreased p-AKT through the targeting of NEDD4 | 2017 | [70] | |
| Leukemia | 10, 20, and 40 μΜ curcumin for 24 h | B-Pre-ALL cells (697, REH, RS4;11, and SupB15 cells) | Treatment with curcumin dose-dependently decreased the protein expression of p-AKT, AKT, p-GSK3, GSK3, and p-FOXO1 FOXO1 | 2019 | [61] |
| 20 and 40 μΜ curcumin for 24 h | T-ALL cells (CEM, HSB2, Jurkat and Molt-4) | Treatment with curcumin resulted in the dephosphorylation of AKT and, thus, inactivation of its downstream effectors, FOXO and GSK3 | 2006 | [71] | |
| 30 μM curcumin for 3, 6, 12, and 24 h or 10, 20, and 40 μM curcumin for 24 h | Human Ph+ B-ALL cell line SUP-B15 | Treatment with curcumin dose- and time-dependently decreased the expression of p-AKT, p-mTOR, p-GSK3, and p-P70S6, but had no effect on PTEN | 2015 | [72] | |
| 25 μΜ curcumin for 24 h | AML cell lines (HL-60, ML-2, MOLM-13, OCI-AML3, OCI-AML5, and U937) | Treatment with curcumin decreased the protein expression of p-AKT in OCI-AML5 and ML-2 cells | 2021 | [73] | |
| Lung | 40 μΜ curcumin for 48 h | NSCLC A549 cells | Treatment with curcumin resulted in dose-dependent inhibition of Akt phosphorylation and mTOR phosphorylation | 2018 | [56] |
| 20 μΜ curcumin for 24 h | NSCLC A549 cells | Treatment with curcumin decreased the levels of p-AKT and p-mTOR without affecting the total AKT and total mTOR | 2020 | [74] | |
| 5, 10, and 20 μΜ curcumin for 24 h | NSCLC A549 cells | Treatment with curcumin at higher concentrations decreased the levels of PI3K and AKT | 2015 | [75] | |
| 25, 50, and 100 μΜ curcumin for 48 h | NSCLC A549 cells | Treatment with curcumin at higher concentrations decreased the levels of p-AKT | 2020 | [76] | |
| Prostate | 5, 10, 15, and 20 μΜ curcumin for 24 h | LNCaP, DU145, and PC3 cells | Treatment of LNCaP cells with curcumin reduced the expression of PI3K subunits (p110 and p85) and p-AKT | 2007 | [77] |
| ♦ 10, 20, 30, 40, and 50 μΜ curcumin for 1 h ♦ 40 μΜ curcumin for up to 120 h | PC-3 cells | ♦ Treatment with curcumin inhibited the levels of p-Akt (T308 and S473), p-FoxO1 (S256), p-GSK3β (S9), p-mTOR (S2448/2481), p-p70S6K (T389), and p-S6 (S235/236) dose-dependently ♦ Treatment with curcumin inhibited the levels of the above mentioned proteins time-dependently | 2008 | [78] | |
| Thyroid | 20 μΜ for 48 h | FTC133 cells | Treatment with curcumin decreased PI3K and p-Akt protein expression | 2014 | [79] |
| 12.5, 25, and 50 μΜ curcumin for 24 h | BCPAP cells | Treatment with curcumin reduced the levels of phosphorylated PDK1, AKT, p70S6, p85S6, S6, and 4E-BP1 without affecting the total expression of the corresponding proteins | 2022 | [80] | |
| 12.5, 25, and 50 μΜ curcumin for 24 h | BCPAP and KTC-1 cells | Treatment with curcumin decreased the levels of p-AKT (Thr308 and Ser473), p70S6, p85S6, S6, and 4E-BP1 dose-dependently | 2021 | [81] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zoi, V.; Kyritsis, A.P.; Galani, V.; Lazari, D.; Sioka, C.; Voulgaris, S.; Alexiou, G.A. The Role of Curcumin in Cancer: A Focus on the PI3K/Akt Pathway. Cancers 2024, 16, 1554. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers16081554
Zoi V, Kyritsis AP, Galani V, Lazari D, Sioka C, Voulgaris S, Alexiou GA. The Role of Curcumin in Cancer: A Focus on the PI3K/Akt Pathway. Cancers. 2024; 16(8):1554. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers16081554
Chicago/Turabian StyleZoi, Vasiliki, Athanassios P. Kyritsis, Vasiliki Galani, Diamanto Lazari, Chrissa Sioka, Spyridon Voulgaris, and Georgios A. Alexiou. 2024. "The Role of Curcumin in Cancer: A Focus on the PI3K/Akt Pathway" Cancers 16, no. 8: 1554. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers16081554