High-Level Production of Succinic Acid from Crude Glycerol by a Wild Type Organism

Thünen-Institute of Agricultural Technology, Bundesallee 47, 38116 Braunschweig, Germany
*
Author to whom correspondence should be addressed.
Catalysts 2020, 10(5), 470; https://0-doi-org.brum.beds.ac.uk/10.3390/catal10050470
Submission received: 26 March 2020
/
Revised: 16 April 2020
/
Accepted: 23 April 2020
/
Published: 25 April 2020
(This article belongs to the Special Issue New Glycerol Upgrading Processes)
Abstract
:With the transition to the bio-based economy, it is becoming increasingly important for the chemical industry to obtain basic chemicals from renewable raw materials. Succinic acid, one of the most important bio-based building block chemicals, is used in the food and pharmaceutical industries, as well as in the field of bio-based plastics. An alternative process for the bio-based production of succinic acid was the main objective of this study, focusing on the biotechnological production of succinic acid using a newly isolated organism. Pure glycerol compared to crude glycerol, at the lowest purity, directly from a biodiesel plant side stream, was successfully converted. A maximum final titer of 117 g L−1 succinic acid and a yield of 1.3 g g−1 were achieved using pure glycerol and 86.9 g L−1 succinic acid and a yield of 0.9 g g−1 using crude glycerol. Finally, the succinic acid was crystallized, achieving maximum yield of 95% and a purity of up to 99%.
1. Introduction
Growing environmental awareness and the finiteness of fossil raw materials require a switch to renewable products, even for basic building blocks of the chemical industry. Succinic acid is a dicarboxylic acid and is one of the "Top 10" chemicals with the potential to be produced from renewable resources [1]. Succinic acid offers great potential as a platform chemical in the application areas of bio-based plastics, solvents, plasticizers, adhesives, and resins. It also serves as a pH regulator and additive in the food industry. In the agricultural and pharmaceutical sectors, succinic acid is used as an additive in agricultural foods, such as animal feed, and in the preparation of medicines [2,3,4,5,6].
Industrially, succinic acid is currently produced petrochemically from maleic acid and is thus based on petroleum [7]. With the transition to a bio-based economy, it is becoming more important for the chemical industry to produce basic chemicals from renewable raw materials. Therefore, there is a great need for an alternative biotechnical production of succinic acid with a suited renewable raw material as a substrate. Preferably, the substrate should not compete with the food and feed industry. Already, today, there are 800 million starving people in the world, and 2 billion people are malnourished. Together, they are about 30% of the world population, and this will continue to grow in the future [8].
For this reason, new concepts are necessary for biotechnologically produced basic building blocks of the chemical industry. Crude glycerol is a side-product of biodiesel production. The production of 100 L of biodiesel produces 10 L of crude glycerol that can be used directly or after further purification steps up to pharmaceutical grade [9].
Succinic acid can be produced by fermentation as it is an intermediate of the tricarboxylic acid cycle (TCA, citrate cycle) and one of the end products of anaerobic metabolism. The fermentative succinic acid production is carried out both with wild-type strains and with genetically modified microorganisms. Among the most promising wild-type strains are Actinobacillus succinogenes, Basfia succiniciproducens, and Mannheimia succiniciproducens isolated from the rumen of ruminants [10]. Furthermore, the model organism Escherichia coli has been genetically modified so that it is of interest for biotechnological succinic acid production. A variety of substrates, such as glucose (starch, cellulose, hydrolysates), sucrose (sugarcane, turnip), maltose (starch), lactose (whey), mannitol (starch), xylose (hemicellulose hydrolysates), and glycerol, are described as usable fermentation substrates. Many strains require carbon dioxide for their growth, which serves as a co-substrate during the production of succinic acid [11]. Under anaerobic conditions, in the reductive branch of the TCA, carbon dioxide is fixed [12,13,14]. Phosphoenolpyruvate carboxylase (PEPC) acts as an essential anaplerotic enzyme under anaerobic conditions. The activity of PEPC determines the carbon flux from phosphoenolpyruvate (PEP) to oxalacetate or to pyruvate. For each C3 molecule, one CO2 molecule is fixed from the air, thereby forming the C4 building block, oxaloacetate, which is subsequently converted via the reductive path of the TCA cycle and the intermediates—malate and fumarate—to the target product succinic acid [15]. Representative examples of bacteria in which succinic acid production has already been commercialized are Saccharomyces cerevisiae, Pichia kudriavzevii, E. coli, M. succiniciproducens, B. succiniciproducens, A. succinogenes, and Corynebacterium glutamicum [16]. Glucose (C6 molecule) and glycerol (C3 molecule) mostly serve as carbon sources. They differ not only in the number of carbon atoms but also in their oxidation states. By using a more reduced substrate, e.g., glycerol, the availability of intracellular nicotinamide adenine dinucleotide (NADH) is increased, and more reduction equivalents are provided for the synthesis of succinic acid. The cofactor pair NADH / NAD+ has a central role in microbial catabolism. Using NAD+, a carbon source is oxidized, and reduction equivalents in the form of NADH are created. The oxidation of NADH to NAD+ is particularly important for cell growth since it achieves a redox equilibrium [17].
Compared to the petrochemical succinic acid production, the usability of renewable raw materials, as well as the carbon dioxide fixation, are the major advantages. A significantly better CO2 balance and a 30–40% lower energy consumption compared to conventional succinic acid obtained from fossil raw materials have led to the commercialization of bio-based succinic acid in recent years [18]. However, the current situation shows that the technologies developed are not yet competitive. The price of oil and political decisions by governments are only two uncertainties in the complex structure that determine the economic success of a bio-based product [19]. Nevertheless, to establish a successful industrial process for the production of biotechnologically produced succinic acid, the following parameters are necessary [2]: selection or development of a production strain, selection of the raw material, specific productivity of the bioreactors, as well as the development of an efficient process for the recovery of succinic acid.
Thus, the newly isolated organism able to convert glycerol, as well as crude glycerol, are the decisive motivation for research on this biotechnological process.
In the present work, crude glycerol, compared to pure glycerol, was tested as a substrate for biotechnological production of succinic acid, using a newly isolated organism. For this purpose, various preliminary experiments were carried out in laboratory glass bottles, followed by investigations in 350 mL parallel bioreactors. In addition, the purity of the isolated succinic acid was determined.
2. Results
2.1. Reference Cultivation with Pure Glycerol
The reference cultivation with the newly isolated organism named Isolate AKR177 was carried out with the Gly medium using pure glycerol as substrate, a cultivation temperature of 32 °C, and 150 rpm in 100 mL laboratory glass bottles (Figure 1). The reference cultivation and all further cultivations were inoculated with 2% inoculum from the second preculture using pure glycerol as a substrate. The pH was buffered by the initial addition of magnesium carbonate and was not regulated during the cultivation. Within the first 24 h, regardless of the initial glycerol concentration, the pH increased slightly from 7.7 to 7.9 due to the fact that the carbonate ions react with water to form bicarbonate and hydroxide ions. In the further cultivation process, however, the pH dropped slightly due to the metabolization of glycerol to organic acids, and the carbonate ions were consumed to buffer the system. The formation of the product, as well as the growth (optical density – OD (605 nm)), exhibited approximately a sigmoidal course. Within the first 24 h, there was a short lag phase in which the substrate was used for biomass production. This was followed by a phase in which succinic acid, as well as the biomass, were formed in parallel to the consumption of glycerol. Overall, the optical density increased to 29 (50 g L−1 initial substrate) and 38 (85 g L−1 initial substrate) and was more or less constant at the end of the cultivation.
Using an initial substrate concentration of 50 g L−1, the batch fermentation was finished after 5 d. The glycerol was completely consumed, and a final concentration of 49.6 g L−1 succinic acid (SA) and 3.2 g L−1 lactic acid (LA) was formed. Using an initial concentration of 85 g L−1 pure glycerol, after 15 d, the substrate glycerol was completely consumed. Based on the substrate curve, it could be assumed that it was used up earlier, but this was not documented due to the lack of sampling. The final concentration of succinic acid was 88.8 g L−1, and the concentration of lactic acid was 6.3 g L−1. Regardless of the initial glycerol concentration, the production of lactic acid started after 48 h and increased slightly and linearly. Besides, with lactic acid, no further side products were formed. The maximum glycerol consumption rate was 0.7 g L−1 h−1 using initial 50 g L−1 glycerol compared to 0.37 g L−1 h−1 with 85 g L−1 initial glycerol. Therefore, the maximum productivity (Table 1) was also higher, using an initial 50 g L−1 glycerol. The yield at the end of the cultivation was comparable with 0.9 and 0.94 g g−1.
2.2. Cultivation with Crude Glycerol
2.2.1. Cultivation with Crude Glycerol in 100 mL Laboratory Glass Bottles
The comparison between pure and crude glycerol in 100 mL laboratory glass bottles is shown in Figure 2. The microorganism was able to convert an initial concentration of 50 g L−1 crude glycerol in the same way as pure glycerol. There were no significant differences in glucose consumption rate, productivity, or final titers of succinic acid (Table 2). During the first 20 h, a lag phase existed where the organism adapted to the cultivation conditions, and biomass was formed. Afterward, the substrate uptake rate increased, which could be seen from the higher glycerol consumption rate. At the same time, the productivity of succinic acid increased. After 96 h, the productivity decreased, and, after 120 h, the glycerol was completely consumed. However, glycerol was expected to be depleted earlier, but this could not be documented due to the lack of sampling. This was also supported by decreasing productivity at the end of cultivation. Besides, during the cultivation with the crude glycerol, only 2.2 g L−1 lactic acid was formed as a sole side product.
Cultivation using that scale and an initial substrate concentration of 50 g L−1 resulted in a similar behavior independent from the purity of the glycerol. Therefore, it was of great interest to have an increased initial substrate concentration. For this purpose, a scale-up was carried out in the parallel bioreactor system using higher initial glycerol concentrations under pH-controlled condition and fed-batch mode.
2.2.2. Cultivation with Crude Glycerol in the 350 mL Bioreactor System Compared to Pure Glycerol
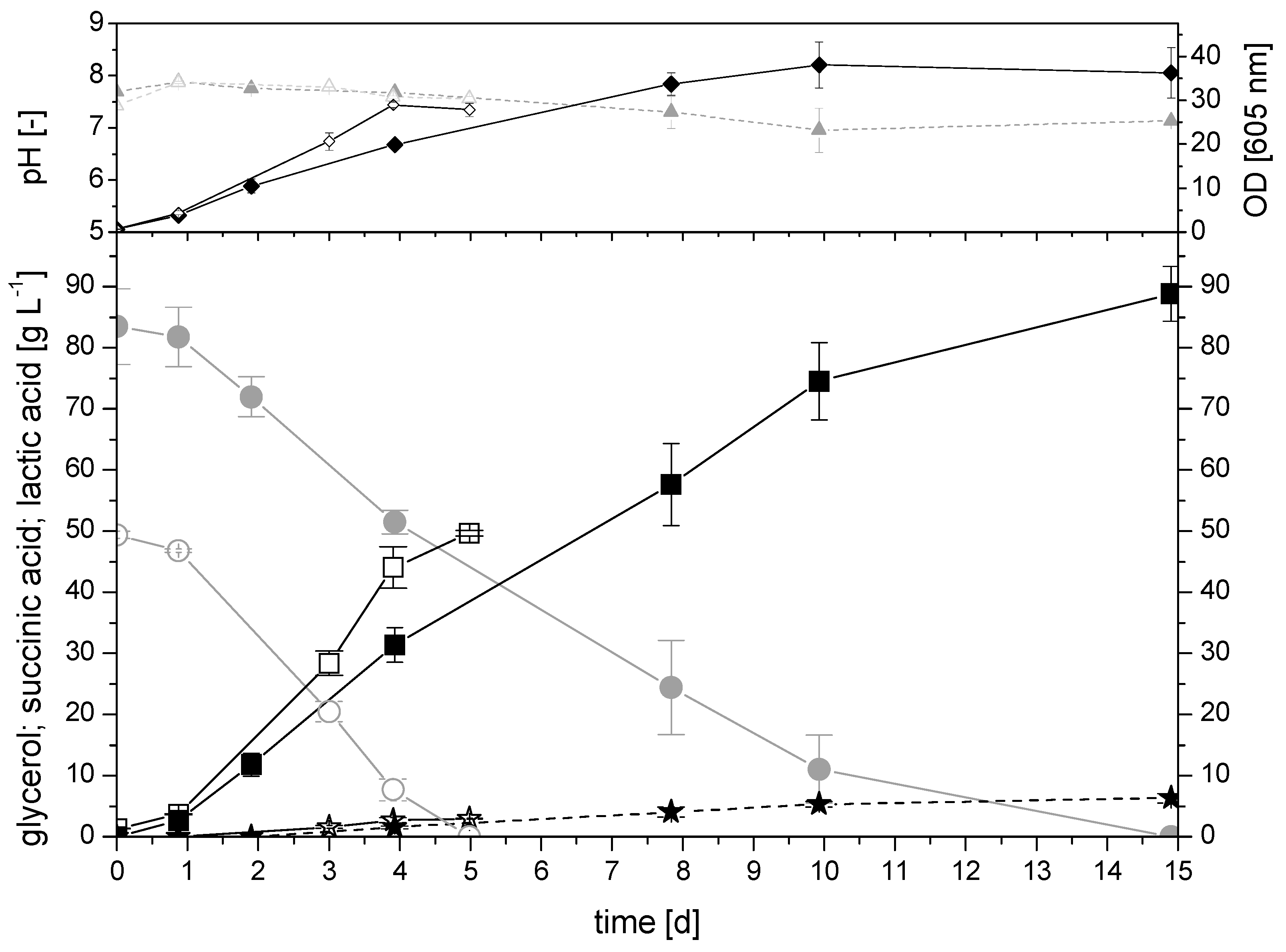
In order to transfer the cultivation in a bioreactor scale under pH control and to switch to a fed-batch mode, the bioreactor system (DASbox) was used. The comparison between pure and crude glycerol in the 350 mL bioreactor system is shown in Figure 3.
Using pure glycerol, the first batch phase (the first 4 days) was started with an initial 67 g L−1 pure glycerol and an unregulated pH using magnesium carbonate as a buffer. The pH dropped from 7.7 to a pH value of 6.8. Comparable to the cultivation in the laboratory glass bottles, the data showed an initial lag phase within the first 24 h. Thereafter, the concentration of succinic acid increased linearly until the substrate was almost consumed, new glycerol was fed after 4 days, and the cultivation strategy was changed with the beginning of that second batch phase. The pH control to a pH value of 7.3 using 2M Na2CO3 was started. Sodium carbonate was used since it is readily soluble in water, in contrast to magnesium carbonate, and can, therefore, be well dosed as a solution. Additionally, the gassing strategy was changed during the second batch phase. During the first batch phase, the reactor was fumigated with a constant volume flow of 4.5 L h−1 and a composition of 50% CO2 and 50% air. During the second batch phase, the reactor was fumigated with a flow rate of 4.5 L h−1 once a day for 13 min with 95% air and 5% CO2. The substrate feed resulted in a concentration of 55 g L−1 glycerol. The microorganisms were in the stationary phase, and the productivity decreased during that second batch phase. After the substrate was almost depleted, the second feed of glycerol was conducted after 167 h (7 days).
In the third batch phase, the glycerol consumption rate and the productivity of succinic acid production were significantly lower than in the previous two phases. The measured final concentration of succinic acid was 117 g L−1 (Table 3). The lactic acid concentration increased approximately linearly, during the cultivation, to a final titer of 8.6 g L−1. Over the entire cultivation process, a total volume of 40 mL of the pH correction agent was added. At an initial volume of 140 mL, this was a volume increase of 28.5%. Of course, this significant dilution had to be taken into account for calculating the yield. However, the substrate and product concentrations, shown in Figure 3, were the measured values. The cultivation was stopped before the glycerol was consumed completely in the third batch phase. With 1.3 g g−1, a yield above 1 g g−1 could be achieved.
Compared to the cultivation with crude glycerol in the 100 mL laboratory glass bottle and an initial glycerol concentration of 64 g L−1, the behavior in the 350 mL bioreactor system was not independent of the purity grade of the glycerol (Figure 3). The maximum glycerol consumption rate using pure glycerol was 1.03 g L−1h−1 and, thus, significantly higher compared to crude glycerol with a maximum glycerol consumption rate of 0.59 g L−1h−1. The first batch was carried out for 6 days with crude glycerol as a substrate, and the reactor was fumigated with a constant volume flow of 4.5 L h−1 and a composition of 50% CO2 and 50% air. After the glycerol was nearly consumed, 5 mL crude glycerol was fed, increasing the concentration to 26 g L−1. At the same time, the fumigation was switched to a flow rate of 4.5 L h−1 once a day for 13 min with 95% air and 5% CO2, and the pH control was initiated with 2M Na2CO3 to a pH value of 7.3. After 8 days, the glycerol was nearly consumed during the second batch, and 5 mL crude glycerol was added a second time. After 11 days, the glycerol consumption decreased, and the cultivation was stopped. The maximum productivity during the cultivation using crude glycerol was 0.55 g L−1 h−1 and 0.69 g L−1 h−1 using pure glycerol. The yield was also lower using crude glycerol, but nevertheless, a yield of 0.9 g g −1 and a final titer of 86.9 g L−1 succinic acid were achieved (Table 3).
2.3. Crystallization and Purity Determination
At the end of the fermentation, the fermentation broth mainly consists of the produced succinic acid as succinate salt, the by-product lactate, residual glycerol, and the cells. An overview of the downstream recovery and purification steps for biologically derived succinic acid is given by Nghiem et al. [20]. In order to isolate the produced succinic acid from the fermentation broth, cooling crystallization was performed similarly to the procedure described by Li et al. [21]. Therefore, the fermentation broth was first centrifuged to remove the cells. The supernatant was acidified with H2SO4 to pH 2 in order to convert the readily soluble succinates to the less soluble succinic acid. Storing the acidified supernatant for 12 h at 4 °C induced succinic acid crystallization. At 4 °C, the solubility of succinic acid is only 30 g L−1, whereas the by-product lactic acid, as well as glycerol, are completely miscible with water and, accordingly, remain in solution [21]. Crystallization was carried out twice for fermentation broths using pure glycerol and crude glycerol as substrate, respectively.
By that procedure, 57–70% of the produced succinic acid could be isolated as crystals, whereas 26–32% of the total succinic acid remained in the mother liquor. A small portion of succinic acid (5–17%) was lost due to wall effects on that small scale. The crystallization yield did not depend on the type of the substrate. Depending on the substrate, the crystals differed in their color and shape. The crystals obtained from the culture broth using pure glycerol appeared white with a fine needle-like shape. The crystals obtained from the culture broth using crude glycerol were slightly colored (brownish-orange) by impurities originating from the crude glycerol and exhibited a coarser structure. Thus, an additional adsorption step by activated carbon should be installed prior to crystallization. Nevertheless, the purity of the crystals was determined. Therefore, the crystals were resolved, and the solution was measured by high-performance liquid chromatography (HPLC). The purity was between 95% and 99.9%, no matter which substrate was used. These results were comparable to those of Li et al., who reported a 70% succinic acid yield and a purity of 90% from a very different cultivation process [21].
3. Discussion
This study demonstrated that pure glycerol, as well as crude glycerol, could be successfully and efficiently converted as a carbon source to succinic acid. With respect to the economic feasibility (based on three parameters: cost of biomass per ton, the availability of the biomass in tons, and the yield in grams per gram), glycerol is listed as a starting material for the bio-based succinic acid production in rank 2 of 15 renewable biomass and carbon sources [22]. Glycerol is a C3 body, and succinic acid is a C4 body; therefore, the maximum yield via the oxidative pathway of the TCA is 0.75 mol mol−1 or 0.78 g g−1. However, if the reductive pathway of the TCA and thus CO2 fixation is activated, a yield of 1.3 g g−1 is achievable. In contrast, succinic acid production based on glucose, and without taking biomass formation in the account, is not balanced in terms of redox status and only achieves a theoretical maximum yield of 1.1 g g−1 without external reducing agent [22,23]. Accordingly, glycerol is a higher reduced carbon source than glucose, so that the conversion of 1 mol glycerol to pyruvate produces 2 mol NADH. Due to the higher degree of reduction, it is obvious that glycerol is favored to form reduced products, such as succinic acid.
Regardless of the purity of the glycerol, the fixation of CO2 was presented in this study and led to a yield of 0.9 g g−1 for crude glycerol and 1.3 g g−1 for pure glycerol, which corresponded to the theoretical maximum yield. The biotechnological production of succinic acid was successfully transferred to the bioreactor scale. The glycerol consumption rate was increased from 0.7 g L−1 h−1 using initial 50 g L−1 glycerol and 100 mL laboratory glass bottle to 1.03 g L−1 h−1 using initial 67 g L−1 glycerol and the bioreactor system. The maximum final titer of 117 g L−1 succinic acid and yield of 1.3 g g−1 were achieved with pure glycerol as a substrate. Further on, it was shown in this study that the organism was very well suited to convert crude glycerol as a substrate for succinic acid production. The crude glycerol was directly used without additional purification steps. Using crude glycerol, the maximum substrate consumption rate, as well as the maximum productivity, were slightly lower compared to pure glycerol, but 87 g L−1 succinic acid was successfully produced with a yield of 0.9 g g−1. Probably, the lower final titer and glycerol consumption rate were caused by impurities present in the crude glycerol, such as fatty acids, heavy metal ions, or salts, which has to be investigated in further experiments. But so far, the achieved values presented in this work are among the highest values reported in the literature using wild-type strains in the biotechnological production of succinic acid. A selection of examples for biotechnological production of succinic acid using different wild-type strains, as well as different types of carbon sources, is given in Table 4. Established microorganisms for the production of succinic acid are Actinobacillus succinogenes, Corynebacterium glutamicum, Anaerobiospirillum succiniciproducens, Mannheimia succiniciproducens, Basfia succiniciproducens, Yarrowia lipolytica, or Escherichia coli. However, Corynebacterium glutamicum, Anaerobiospirillum succiniciproducens, Mannheimia succiniciproducens, Basfia succiniciproducens, Yarrowia lipolytica, as well as Escherichia coli, are mainly used in the form of genetically-modified organisms [12,24,25,26,27,28,29,30,31].
The highest final titer of 198 g L−1 succinic acid is achieved with glycerol and a genetically-modified (GMO) Yarrowia lipolytica [24]. The investigations regarding crude glycerol as sole carbon sources have also been carried out. According to Li et. al (2018), the treatment of crude glycerol by activated carbon effectively released the inhibition on the glycerol consumption and succinate production of the genetically engineered E. coli strains, and the fermentation result of the treated crude glycerol was with 566.0 mM succinic acid compared to the usage of pure glycerol [31]. Gao et al. (2016) genetically modified Y. lipolytica, which resulted in 43 g L−1 succinic acid production from crude glycerol. Using the fed-batch strategy in 2.5 L fermenter, up to 160 g L−1 succinic acid was yielded [25].
Based on the metabolic pathways, different strategies of metabolic engineering could be used to improve the production of succinic acid in genetically-modified microorganisms [15]. The use of GMOs is not widespread in Europe, so it is our intention to reach comparable production levels of succinic acid with a wild-type strain and a beneficial substrate. Succinate is the key intermediate of the TCA cycle and overproduced by some wild-type strains as an extracellular final fermentation product. Although the wild-type strains can already produce high concentrations of succinic acid, the high costs for the cultivation media prevent commercialization. So far, little attention has been paid to the usage of glycerol as a sole carbon source combined with a wild type strain. Vlysidis et al. (2011) used the wild type strain Actinobacillus succinogenes and achieved 29.3 g L−1 succinic acid in batch fermentation and glycerol as the sole carbon source [36]. In comparison with the literature, it could be shown in our study that very high final titers, up to 47 g L−1 succinic acid in batch and 87 g L−1 succinic acid in fed-batch cultivation, were achieved with crude glycerol as substrate. This crude glycerol, a side-product of rapeseed biodiesel production, was very successfully converted without further purification steps, and thus the costs of the biotechnological succinic acid production could be reduced.
4. Materials and Methods
4.1. Microorganism
The experiments were performed with a self-isolated culture named Isolate AKR177, which is safety deposited at the DSMZ (German Collection of Microorganisms and Cell Cultures, Braunschweig, Germany). The Isolate AKR177 was cultivated on selective culture media for Actinomycetes (ISP media). The DNA was extracted from this culture, and the 16S rRNA gene could be amplified by polymerase chain reaction (PCR) with selective primers. These amplicons were then sequenced, and a strain of the class Actinobacteria was identified as the production strain [38].
4.2. Media composition and Chemicals
If not mentioned otherwise, the production media was the Gly medium containing 50 g L−1 glycerol, 5 g L−1 yeast extract, 5 g L−1 tryptone/peptone ex casein, 1.5 g L−1 K2HPO4, 1 g L-1 NaCl, 1 g L−1 (NH4)2SO4, 0.265 g L−1 CaCl2·2H2O, 0.094 g L−1 MgCl2, 9 mL 1 M H3PO4, 30 mL 1 M HCl, and 50 g L−1 MgCO3. All components were prepared separately as stock solutions. MgCO3 was weighed in the glassware and autoclaved separately.
All media components were p.a. quality and purchased from Merck (Darmstadt, Germany), Sigma-Aldrich (St. Louis, MO, USA), AppliChem GmbH (Darmstadt, Germany), VWR International GmbH (Darmstadt, Germany), or Roth (Karlsruhe, Germany). Glycerol was added to the medium as a carbon source, either as pure glycerol (≥98%) or crude glycerol K501 provided by Biopetrol Schwarzheide GmbH (Schwarzheide, Germany). The crude glycerol originated from transesterification of rapeseed oil and was the least processed stage of the rapeseed biodiesel production. The crude glycerol K501 contained 48%(wt/wt) glycerol, pH value of 6.2, 10 ppm chloride, an ash content of 3.4%, and the fatty acid content was not determined.
4.3. Cultivation
Precultures were conducted in 100 mL laboratory glass bottles (Borosilicatglas DIN GL 45, SIMAX®, Bohemia Cristal Handelsgesellschaft mbH, Selb, Germany). The laboratory glass bottles were sealed with a silicone rubber septum (thickness: 3 mm, diameter: 42 mm, Roth, Karlsruhe, Germany) with a cannula (diameter: 1.6 mm, length: 15 mm; HSW-PREMIUM®, Henke-Sass, Wolf GmbH, Tuttlingen, Germany). A 50 mL Gly medium, with 20 g L−1 glycerol and 28 g L −1 MgCO3, was used for the preculture. A 0.5 mL glycerol stock culture (cryoculture) (corresponding to 1% total volume) was used for the inoculation. The precultures were cultivated at 32 °C and 150 rpm (25 mm shaking diameter) for 72 h.
The second precultures were prepared from the precultures in 100 mL laboratory glass bottles (equipped with septum and cannula). For the second precultures, 50 mL Gly medium, with 50 g L−1 glycerol and 50 g L −1 MgCO3, were inoculated with 1 mL preculture (corresponds to 2% total volume) and cultivated at 32 °C and 150 rpm (25 mm shaking diameter) for 72 h.
The main cultures were cultivated in 100 mL laboratory glass bottles (equipped with septum and cannula) with 50 mL Gly medium containing 50/88.8 g L−1 glycerol and 50/88.8 g L −1 MgCO3, and they were inoculated with 1 mL of the second preculture (corresponds to 2% total volume). The cultivation was carried out at 32 °C and at 150 rpm (25 mm shaking diameter). To generate valuable data, one milliliter of the sample was taken daily under sterile conditions. The sample was examined for its optical density, pH value, and its substrate and product concentration. All cultivations were carried out in triplicates.
The cultivation in bioreactors was conducted in four parallel 350 mL-bioreactors, equipped with a Rushton impeller (6 blades) (model SR0250ODLS, DASGIP GmbH, Jülich, Germany), and a working volume of 140 mL Gly medium. DASGIP Control software (DASGIP GmbH, Jülich, Germany) was used for the regulation of gassing, temperature, pH-value, and stirring rate, as well as recording the data. During the first batch phase, MgCO3 was added equimolar to the initial glycerol concentration to regulate the pH value. In order to ensure microaerophilic conditions, the reactor was sealed airtight with seals. The cultivation started after inoculation with 2% of the working volume (corresponds to 2.8 mL) of the second preculture. During the first batch phase, the headspace was fumigated with 50% CO2 and 50% air at a flow rate of 4.5 L h −1. After the glycerol was almost used up, a feed with glycerol was carried out. Parallel to the supplementation, with the substrate, gassing was changed. The reactor was fumigated once a day for 13 min with 95% air and 5% CO2 at a flow rate of 4.5 L h −1. During that second cultivation phase, the pH regulation to pH 7.3 was carried out with 2 M Na2CO3. After the glycerol was almost used up again, a second feed with glycerol was carried out under the same pH and gassing strategy. The experiments were performed at 32 °C, 400 rpm, and a filling volume of 140 mL. All cultivations were carried out in duplicates.
4.4. Analytical Methods
The biomass formation was determined using the optical density OD605 (T80 UV/VIS Spectrophotometer, PG Instruments Limited, Lutterworth, Great Britain). The absorbance of the sample diluted with 1 M HCl was measured at a wavelength of 605 nm.
Substrate (glycerol), product (succinic acid), and by-products (lactic acid) in the sample supernatant were identified and quantified using a Shimadzu HPLC (Shimadzu Corp., Kyoto, Japan) with an HPX-87H column (BioRad, Munich, Germany) with a refractive index detector (RI) and UV detector at 210 nm. For the HPLC analysis, a sample of 100 μL was diluted with 900 μL 50 mM H2SO4. The samples were centrifuged at 12,000 rpm for 7 min at 4 °C, and the supernatant was used for further dilutions and analysis using the HPLC. The column of the HPLC was tempered at 40 °C, and as a mobile phase, a 5 mM H2SO4 solution at a flow rate of 0.7 mL min−1 was used.
4.5. Crystallization and Purity Determination
In order to obtain succinic acid in crystalline form, the fermentation broth was first centrifuged at 8500 rpm for 15 min at 10 °C. The supernatant was then acidified with sulfuric acid (96 % H2SO4) to achieve a pH of about 2 and stored at 4 °C for 12 h. During this time, the crystallization took place. The crystals were then separated from the solution by filtration (Black ribbon MN 640 w, Macherey Nagel, Düren, Germany). The filtrate was then diluted 1:100 with 50 mM H2SO and analyzed by HPLC. The solid (crystals) was dried at 105 °C to constant weight for at least 48 h. The weight was determined gravimetrically. A defined weight of the crystals was redissolved in ultrapure water (Millipore, Merck KGaA, Darmstadt, Germany). The resulting solution was analyzed using HPLC to determine the purity of the crystals.
Author Contributions
A.K. and L.H. designed the experiments. L.H. performed the experiments. S.B. and K.G. established the methodology. U.P. and A.K. supervised the experiments. A.K. conducted the original draft preparation. A.K. and U.P. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the German Federal Ministry of Food and Agriculture, following a decision of the German Bundestag, via the Agency of Renewable Resources (Grant No. 22036214).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Bozell, J.J.; Petersen, G.R. Technology development for the production of biobased products from biorefinery carbohydrates—The us department of energy’s “top 10” revisited. Green Chem. 2010, 12, 539. [Google Scholar] [CrossRef]
- Ferone, M.; Raganati, F.; Ercole, A.; Olivieri, G.; Salatino, P.; Marzocchella, A. Continuous succinic acid fermentation by Actinobacillus succinogenes in a packed-bed biofilm reactor. Biotechnol. Biofuels 2018, 11, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKinlay, J.B.; Vieille, C.; Zeikus, J.G. Prospects for a bio-based succinate industry. Appl. Microbiol. Biotechnol. 2007, 76, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Bechthold, I.; Bretz, K.; Kabasci, S.; Kopitzky, R.; Springer, A. Succinic acid: A new platform chemical for biobased polymers from renewable resources. Chem. Eng. Technol. 2008, 31, 647–654. [Google Scholar] [CrossRef]
- Bradfield, M.F.; Mohagheghi, A.; Salvachua, D.; Smith, H.; Black, B.A.; Dowe, N.; Beckham, G.T.; Nicol, W. Continuous succinic acid production by Actinobacillus succinogenes on xylose-enriched hydrolysate. Biotechnol. Biofuels 2015, 8, 181. [Google Scholar] [CrossRef] [Green Version]
- Cukalovic, A.; Stevens, C.V. Feasibility of production methods for succinic acid derivatives: A marriage of renewable resources and chemical technology. Biofuels Bioprod. Biorefin. 2008, 2, 505–529. [Google Scholar] [CrossRef]
- Cornils, B.; Lappe, P. Ullmann’s Encyclopedia of Industrial Chemistry; Wiley-VCH Verlag: Weinheim, Germany, 2010; pp. 287–304. [Google Scholar]
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2019. Safeguarding Against Economic Slowdowns and Downturns; Licence: CC BY-NC-SA 3.0 IGO; FAO: Rome, Italy, 2019. [Google Scholar]
- Yazdani, S.S.; Gonzalez, R. Anaerobic fermentation of glycerol: A path to economic viability for the biofuels industry. Curr. Opin. Biotechnol. 2007, 18, 213–219. [Google Scholar] [CrossRef]
- Pateraki, C.; Patsalou, M.; Vlysidis, A.; Kopsahelis, N.; Webb, C.; Koutinas, A.A.; Koutinas, M. Actinobacillus succinogenes: Advances on succinic acid production and prospects for development of integrated biorefineries. Biochem. Eng. J. 2016, 112, 285–303. [Google Scholar] [CrossRef]
- Herselman, J.; Bradfield, M.F.A.; Vijayan, U.; Nicol, W. The effect of carbon dioxide availability on succinic acid production with biofilms of Actinobacillus succinogenes. Biochem. Eng. J. 2017, 117, 218–225. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Lee, S.Y. Production of succinic acid by bacterial fermentation. Enzyme Microb. Technol. 2006, 39, 352–361. [Google Scholar] [CrossRef]
- Cheng, K.-K.; Zhao, X.-B.; Zeng, J.; Zhang, J.-A. Biotechnological production of succinic acid: Current state and perspectives. Biofuels Bioprod. Biorefin. 2012, 6, 302–318. [Google Scholar] [CrossRef]
- Chen, Y.; Nielsen, J. Advances in metabolic pathway and strain engineering paving the way for sustainable production of chemical building blocks. Curr. Opin. Biotechnol. 2013, 24, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Guo, F.; Zhang, S.; Zhang, W.; Yang, Q.; Dong, W.; Jiang, M.; Ma, J.; Xin, F. Bio-based succinic acid: An overview of strain development, substrate utilization, and downstream purification. Biofuels Bioprod. Biorefin. 2020. [Google Scholar] [CrossRef]
- Tan, J.P.; Jahim, J.M.; Harun, S.; Wu, T.Y. Overview of the potential of bio-succinic acid production from oil palm fronds. J. Phys. Sci. 2017, 28, 53–72. [Google Scholar]
- Ahn, J.H.; Jang, Y.-S.; Lee, S.Y. Production of succinic acid by metabolically engineered microorganisms. Curr. Opin. Biotechnol. 2016, 42, 54–66. [Google Scholar] [CrossRef]
- Li, J.; Jiang, M.; Chen, K.; Shang, L.; Wei, P.; Ying, H.; Ye, Q.; Ouyang, P.; Chang, H. Enhanced production of succinic acid by Actinobacillus succinogenes with reductive carbon source. Process Biochem. 2010, 45, 980–985. [Google Scholar] [CrossRef]
- Biobasierte Chemikalien|chemanager-online.com—Chemie und Life Science. Available online: https://www.chemanager-online.com/themen/chemikalien-distribution/biobasierte-chemikalien (accessed on 12 August 2019).
- Nghiem, N.P.; Kleff, S.; Schwegmann, S. Succinic acid: Technology development and commercialization. Fermentation 2017, 3, 26. [Google Scholar] [CrossRef]
- Li, Q.; Wang, D.; Wu, Y.; Li, W.L.; Zhang, Y.J.; Xing, J.M.; Su, Z.G. One step recovery of succinic acid from fermentation broths by crystallization. Sep. Purif. Technol. 2010, 72, 294–300. [Google Scholar] [CrossRef]
- Tan, J.P.; Md Jahim, J.; Wu, T.Y.; Harun, S.; Kim, B.H.; Mohammad, A.W. Insight into bio-mass as a renewable carbon source for the production of succinic acid and the factors affecting the metabolic flux toward higher succinate yield. Industrial & Engineering. Chem. Res. 2014, 53, 16123–16134. [Google Scholar]
- Blankschien, M.D.; Clomburg, J.M.; Gonzalez, R. Metabolic engineering of Escherichia coli for the production of succinate from glycerol. Metab. Eng. 2010, 12, 409–419. [Google Scholar] [CrossRef]
- Li, C.; Yang, X.; Gao, S.; Wang, H.; Lin, C.S.K. High efficiency succinic acid production from glycerol via in situ fibrous bed bioreactor with an engineered Yarrowia lipolytica. Bioresour. Technol. 2017, 225, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.J.; Yang, X.F.; Wang, H.M.; Rivero, C.P.; Li, C.; Cui, Z.Y.; Qi, Q.S.; Lin, C.S.K. Robust succinic acid production from crude glycerol using engineered Yarrowia lipolytica. Biotechnol. Biofuels 2016, 9, 179. [Google Scholar] [CrossRef]
- Yocum, R.R.; Hermann, T.; Hu, X. Fermentation of Glycerol to Organic Acids. U.S. Patent 2014 0234923 A1, 21 August 2014. [Google Scholar]
- Wang, C.; Zhang, H.; Cai, H.; Zhou, Z.; Chen, Y.; Chen, Y.; Ouyang, P. Succinic acid production from corn cob hydrolysates by genetically engineered Corynebacterium glutamicum. Appl. Biochem. Biotechnol. 2014, 172, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, H.; Haefner, S.; Von Abendroth, G.; Hollman, R.; Raddatz, A.; Ernst, H.; Gurski, H. Novel Microbial Succinic Acid Producers and Purification of Succinic Acid. U.S. Patent 2014 0127764 A1, 8 Jan 2014. [Google Scholar]
- Okino, S.; Noburyu, R.; Suda, M.; Jojima, T.; Inui, M.; Yukawa, H. An efficient succinic acid production process in a metabolically engineered Corynebacterium glutamicum strain. Appl. Microbiol. Biotechnol. 2008, 81, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Vemuri, G.N.; Eiteman, M.A.; Altman, E. Succinate production in dual-phase Escherichia coli fermentations depends on the time of transition from aerobic to anaerobic conditions. J. Ind. Microbiol. Biotechnol. 2002, 28, 325–332. [Google Scholar] [CrossRef]
- Li, Q.; Huang, B.; He, Q.; Lu, J.; Li, X.; Li, Z.; Wu, H.; Ye, Q. Production of succinate from simply purified crude glycerol by engineered Escherichia coli using two-stage fermentation. Bioresour. Bioprocess. 2018, 5, 41. [Google Scholar] [CrossRef]
- Guettler, M.V.; Rumler, D.; Jain, M.K. Actinobacillus succinogenes sp. Nov., a novel succinic-acid-producing strain from the bovine rumen. Int. J. Syst. Bacteriol. 1999, 49, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Bradfield, M.F.A.; Nicol, W. Continuous succinic acid production from xylose by Actinobacillus succinogenes. Bioproc. Biosyst. Eng. 2016, 39, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Stellmacher, R.; Hangebrauk, J.; von Abendroth, G.; Scholten, E.; Wittmann, C. Fermentative production of succinic acid with Basfia succiniciproducens. Chem. Ing. Tech. 2010, 82, 1223–1229. [Google Scholar] [CrossRef]
- Scholten, E.; Dagele, D. Succinic acid production by a newly isolated bacterium. Biotechnol. Lett. 2008, 30, 2143–2146. [Google Scholar] [CrossRef]
- Vlysidis, A.; Binns, M.; Webb, C.; Theodoropoulos, C. Glycerol utilisation for the production of chemicals: Conversion to succinic acid, a combined experimental and computational study. Biochem. Eng. J. 2011, 58–59, 1–11. [Google Scholar] [CrossRef]
- Shen, N.; Qin, Y.; Wang, Q.; Liao, S.; Zhu, J.; Zhu, Q.; Mi, H.; Adhikari, B.; Wei, Y.; Huang, R. Production of succinic acid from sugarcane molasses supplemented with a mixture of corn steep liquor powder and peanut meal as nitrogen sources by Actinobacillus succinogenes. Lett. Appl. Microbiol. 2015, 60, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Bromann, S.; Goy, K.; Kuenz, A.; Prüße, U.; Willke, T. Schlussbericht zum Vorhaben Biobst—Biotechnische Herstellung von Bernsteinsäure aus Nachwachsenden Rohstoffen (Biotechnological Production of Succinic Acid from Renewable Resources); Laufzeit: 01.09.2015 bis 15.10.2018; Johann Heinrich von Thünen-Institut: Braunschweig, Germany, 2019. [Google Scholar]
Figure 1.
Reference cultivation in 100 mL laboratory glass bottle: batch fermentation with initial 50 g L−1 pure glycerol (open symbols) and initial 85 g L−1 pure glycerol (closed symbols); succinic acid concentration (square, grams per liter), glycerol concentration (circle, grams per liter), lactic acid concentration (stars, grams per liter), and formation of biomass (diamond, optical density at 605 nm), pH (triangle); Isolate AKR177, Gly medium, MgCO3, 32 °C, 150 rpm.
Figure 1.
Reference cultivation in 100 mL laboratory glass bottle: batch fermentation with initial 50 g L−1 pure glycerol (open symbols) and initial 85 g L−1 pure glycerol (closed symbols); succinic acid concentration (square, grams per liter), glycerol concentration (circle, grams per liter), lactic acid concentration (stars, grams per liter), and formation of biomass (diamond, optical density at 605 nm), pH (triangle); Isolate AKR177, Gly medium, MgCO3, 32 °C, 150 rpm.

Figure 2.
Cultivation in 100 mL laboratory glass bottle: batch fermentation with initial 49.4 g L−1 pure glycerol (open symbols) and 47.7 g L−1 crude glycerol (closed symbols); succinic acid concentration (square, grams per liter), glycerol concentration (circle, grams per liter), lactic acid concentration (stars, grams per liter), and formation of biomass (diamond, optical density at 605 nm), pH (triangle); Isolate AKR177, Gly medium, MgCO3, 32 °C, 150 rpm.
Figure 2.
Cultivation in 100 mL laboratory glass bottle: batch fermentation with initial 49.4 g L−1 pure glycerol (open symbols) and 47.7 g L−1 crude glycerol (closed symbols); succinic acid concentration (square, grams per liter), glycerol concentration (circle, grams per liter), lactic acid concentration (stars, grams per liter), and formation of biomass (diamond, optical density at 605 nm), pH (triangle); Isolate AKR177, Gly medium, MgCO3, 32 °C, 150 rpm.

Figure 3.
Cultivation in 350 mL bioreactor system: fed-batch fermentation with initial 67 g L−1 pure glycerol (open symbols) and 64 g L−1 crude glycerol (closed symbols); succinic acid concentration (square), glycerol concentration (circle), lactic acid concentration (stars), and formation of biomass (diamond, optical density at 605 nm), pH (triangle); Isolate AKR177; Gly medium; MgCO3; after 1st substrate feeding, pH control activated to a pH value of 7.3 using 2M Na2CO3; 32 °C; 400 rpm.
Figure 3.
Cultivation in 350 mL bioreactor system: fed-batch fermentation with initial 67 g L−1 pure glycerol (open symbols) and 64 g L−1 crude glycerol (closed symbols); succinic acid concentration (square), glycerol concentration (circle), lactic acid concentration (stars), and formation of biomass (diamond, optical density at 605 nm), pH (triangle); Isolate AKR177; Gly medium; MgCO3; after 1st substrate feeding, pH control activated to a pH value of 7.3 using 2M Na2CO3; 32 °C; 400 rpm.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Results of the reference cultivation in 100 mL laboratory glass bottle: Isolate AKR177, Gly medium with initial 50 g L−1 and 85 g L−1 pure glycerol, MgCO3, 32 °C, 150 rpm.
Table 1.
Results of the reference cultivation in 100 mL laboratory glass bottle: Isolate AKR177, Gly medium with initial 50 g L−1 and 85 g L−1 pure glycerol, MgCO3, 32 °C, 150 rpm.
| Reference Cultivation in 100 mL Laboratory Glass Bottle | ||
|---|---|---|
| Initial 50 g L−1 Substrate | Initial 85 g L−1 Substrate | |
| CEnd, SA / g L−1 | 49.4 | 88.8 |
| CEnd, LA / g L−1 | 3.2 | 6.3 |
| Pmax / g L−1 h−1 | 0.76 | 0.31 |
| P / g L−1 h−1 | 0.37 | 0.22 |
| Y1 / g g−1 | 0.9 | 0.94 |
1 produced succinic acid per consumed glycerol, including the dilution or concentration factor; final concentration (CEnd); maximum productivity (Pmax); productivity (P); yield (Y).
Table 2.
Cultivation in 100 mL laboratory glass bottle: batch fermentation with initial 50 g L−1 pure glycerol and crude glycerol; Isolate AKR177, Gly medium, MgCO3, 32 °C, 150 rpm.
Table 2.
Cultivation in 100 mL laboratory glass bottle: batch fermentation with initial 50 g L−1 pure glycerol and crude glycerol; Isolate AKR177, Gly medium, MgCO3, 32 °C, 150 rpm.
| Pure Glycerol | Crude Glycerol | |
|---|---|---|
| CEnd, SA / g L−1 | 49.4 | 46.8 |
| CEnd, LA / g L−1 | 3.2 | 2.3 |
| Pmax / g L−1 h−1 | 0.76 | 0.62 |
| P / g L−1 h−1 | 0.37 | 0.35 |
| Y1 / g g−1 | 0.9 | 0.88 |
1 produced succinic acid per consumed glycerol, including the dilution or concentration factor.
Table 3.
Cultivation in the 350 mL bioreactor system: Isolate AKR177; Gly medium with initial 67 g L−1 pure or crude glycerol; after 1st substrate feeding, pH control activated to a pH value of 7.3 using 2M Na2CO3; MgCO3; 32 °C; 400 rpm.
Table 3.
Cultivation in the 350 mL bioreactor system: Isolate AKR177; Gly medium with initial 67 g L−1 pure or crude glycerol; after 1st substrate feeding, pH control activated to a pH value of 7.3 using 2M Na2CO3; MgCO3; 32 °C; 400 rpm.
| Pure Glycerol | Crude Glycerol | |
|---|---|---|
| CEnd, SA / g L−1 | 117 | 86.9 |
| CEnd, LA / g L−1 | 8.6 | 6 |
| Pmax / g L−1 h−1 | 0.69 | 0.55 |
| P / g L−1 h−1 | 0.34 | 0.33 |
| Y1 / g g−1 | 1.3 | 0.9 |
1 produced succinic acid per consumed glycerol, including the dilution or concentration factor.
Table 4.
Literature data on succinic acid production, yield, and productivity using different microorganisms and carbon sources using wild-type strains.
Table 4.
Literature data on succinic acid production, yield, and productivity using different microorganisms and carbon sources using wild-type strains.
| Microorganism | Substrate | Succinic Acid | Yield | Productivity | Reference |
|---|---|---|---|---|---|
| (g L−1) | (g g−1) | (g L−1h−1) | |||
| Actinobacillus succinogenes 130Z | Glucose | 106 | 0.8 | 1.34 | [32] |
| Xylose1 | 29.4 | 0.68 | 3.41 | [33] | |
| Basfia succiniciproducens DD1 | Glucose | 31.7 | 0.63 | 1.32 | [34] |
| Basfia succiniciproducens DD1 | Crude glycerol | 8.4 | 1.2 | 0.9 | [35] |
| Actinobacillus succinogenes ATCC 55618 | Glycerol | 29.3 | 1.23 | 0.27 | [36] |
| Actinobacillus succinogenes GXAS 137 | Sugar cane molasses | 64.7 | 0.86 | 1.4 | [37] |
| Isolate AKR177 | Glycerol | 117 | 1.3 | 0.34 | This study |
| Isolate AKR177 | Crude glycerol | 86.9 | 0.9 | 0.33 | This study |
1 continuous, biofilm reactor (immobilized).
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kuenz, A.; Hoffmann, L.; Goy, K.; Bromann, S.; Prüße, U. High-Level Production of Succinic Acid from Crude Glycerol by a Wild Type Organism. Catalysts 2020, 10, 470. https://0-doi-org.brum.beds.ac.uk/10.3390/catal10050470
AMA Style
Kuenz A, Hoffmann L, Goy K, Bromann S, Prüße U. High-Level Production of Succinic Acid from Crude Glycerol by a Wild Type Organism. Catalysts. 2020; 10(5):470. https://0-doi-org.brum.beds.ac.uk/10.3390/catal10050470
Chicago/Turabian StyleKuenz, Anja, Lisa Hoffmann, Katharina Goy, Sarah Bromann, and Ulf Prüße. 2020. "High-Level Production of Succinic Acid from Crude Glycerol by a Wild Type Organism" Catalysts 10, no. 5: 470. https://0-doi-org.brum.beds.ac.uk/10.3390/catal10050470
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.