Integrative Structural and Computational Biology of Phytases for the Animal Feed Industry

 ,
,
Abstract
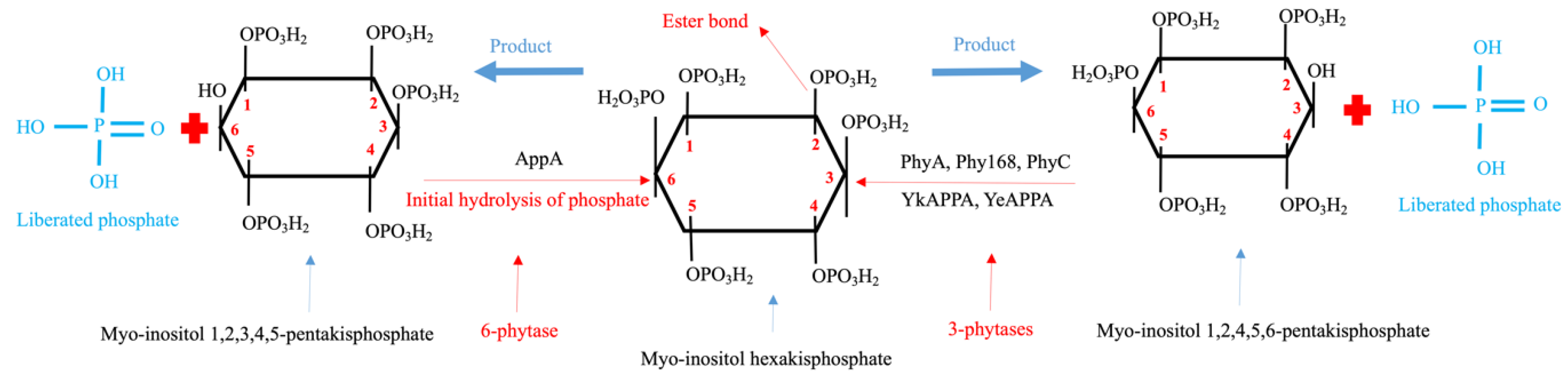
:1. Introduction
2. Modifications of Phytases
2.1. Improved Thermostability in Phytase
2.1.1. Aspergillus Phytases
2.1.2. Escherichia coli Phytase
2.1.3. Bacillus Phytases
2.1.4. Yersinia Phytases
2.1.5. Other Phytases
2.2. Improved Acid Stability
2.2.1. Aspergillus Phytase
2.2.2. Escherichia coli Phytase
2.2.3. Yersinia Phytases
2.3. Improved Proteolytic Stability
2.3.1. Yersinia Phytases
2.3.2. Escherichia coli Phytase
2.4. Improved Catalytic Efficiency and Specific Activity
2.4.1. Aspergillus Phytases
2.4.2. Escherichia coli Phytase
2.4.3. Bacillus Phytases
2.4.4. Yersinia Phytases
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sparvoli, F.; Cominelli, E. Seed biofortification and phytic acid reduction: A conflict of interest for the plant? Plants 2015, 4, 728–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushik, G. Applied Environmental Biotechnology: Present Scenario and Future Trends; Springer: New Delhi, India, 2015; pp. 1–167. [Google Scholar] [CrossRef]
- Gupta, R.K.; Gangoliya, S.S. Reduction of phytic acid and enhancement of bioavailable micronutrients in food grains. J. Food Sci. Technol. 2015, 52, 676–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Hack, M.E.A.; Alagawany, M.; Arif, M.; Emam, M.; Saeed, M.; Arain, M.A.; Siyal, F.A.; Patra, A.; Elnesr, S.S.; Khan, R.U. The uses of microbial phytase as a feed additive in poultry nutrition—A review. Ann. Anim. Sci. 2018, 18, 639–658. [Google Scholar] [CrossRef] [Green Version]
- Dersjant-Li, Y.; Awati, A.; Schulze, H.; Partridge, G. Phytase in non-ruminant animal nutrition: A critical review on phytase activities in the gastrointestinal tract and influencing factors. J. Sci. Food Agric. 2015, 95, 878–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afinah, S.; Yazid, A.M.; Anis Shobirin, M.H.; Shuhaimi, M. Review Article Phytase: Application in food industry. Int. Food Res. J. 2010, 17, 13–21. [Google Scholar]
- Lim, D.; Golovan, S.; Forsberg, C.W.; Jia, Z. Crystal structures of Escherichia coli phytase and its complex with phytate. Nat. Struct. Biol. 2000, 7, 108–113. [Google Scholar] [CrossRef]
- Shin, S.; Ha, N.C.; Oh, B.C.; Oh, T.K.; Oh, B.H. Enzyme mechanism and catalytic property of β propeller phytase. Structure 2001, 9, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Chanderman, A.; Makolomakwa, M.; Perumal, K.; Singh, S. Microbial production of phytases for combating environmental phosphate pollution and other diverse applications. Crit. Rev. Environ. Sci. Technol. 2016, 46, 556–591. [Google Scholar] [CrossRef] [Green Version]
- Madsen, C.K.; Brinch-pedersen, H. Molecular Advances on Phytases in Barley and Wheat. Int. J. Mol. Sci. 2019, 20, 2459. [Google Scholar] [CrossRef] [Green Version]
- Troxell, B.; Petri, N.; Daron, C.; Pereira, R.; Mendoza, M.; Hassan, H.M.; Koci, M.D. Poultry body temperature contributes to invasion control through reduced expression of Salmonella pathogenicity island 1 genes in Salmonella enterica serovars typhimurium and enteritidis. Appl. Environ. Microbiol. 2015, 81, 8192–8201. [Google Scholar] [CrossRef] [Green Version]
- Outchkourov, N.; Petkov, S. Phytases for Feed Applications. In Industrial Enzyme Applications; Vogel, A., Ed.; Wiley-VCH: Weinheim, Germany, 2019; pp. 255–285. [Google Scholar] [CrossRef]
- Mrudula Vasudevan, U.; Jaiswal, A.K.; Krishna, S.; Pandey, A. Thermostable phytase in feed and fuel industries. Bioresour. Technol. 2019, 278, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, M.R.; Ravindran, V.; Svihus, B. Pelleting of broiler diets: An overview with emphasis on pellet quality and nutritional value. Anim. Feed Sci. Technol. 2013, 179, 1–23. [Google Scholar] [CrossRef]
- Modarres, H.P.; Mofrad, M.R.; Sanati-Nezhad, A. Protein thermostability engineering. RSC Adv. 2016, 6, 115252–115270. [Google Scholar] [CrossRef]
- Jimenez, A.; Miriam, R.; Merino, V.F. Tailoring Proteins to Re-Evolve Nature: A Short Review. Mol. Biotechnol. 2018, 60, 946–974. [Google Scholar] [CrossRef]
- Liu, S.Y.; Selle, P.H.; Cowieson, A.J. Influence of conditioning temperatures on amino acid digestibility coefficients at four small intestinal sites and their dynamics with starch and nitrogen digestion in sorghum-based broiler diets. Anim. Feed Sci. Technol. 2013, 185, 85–93. [Google Scholar] [CrossRef]
- Noorbatcha, I.A.; Sultan, A.M.; Salleh, H.M.; Amid, A. Understanding thermostability factors of aspergillus niger PhyA phytase: A molecular dynamics study. Protein J. 2013, 32, 309–316. [Google Scholar] [CrossRef]
- Liao, Y.; Li, C.M.; Chen, H.; Wu, Q.; Shan, Z.; Han, X.Y. Site-directed mutagenesis improves the thermostability and catalytic efficiency of aspergillus niger N25 phytase mutated by I44E and T252R. Appl. Biochem. Biotechnol. 2013, 171, 900–915. [Google Scholar] [CrossRef]
- Hesampour, A.; Ehsan, S.; Siadat, R. Enhancement of Thermostability and Kinetic Efficiency of Aspergillus niger PhyA Phytase by Site-Directed Mutagenesis. Appl. Biochem. Biotechnol. 2014. [Google Scholar] [CrossRef]
- Fonseca-Maldonado, R.; Maller, A.; Bonneil, E.; Thibault, P.; Botelho-Machado, C.; Ward, R.J.; Polizeli, M.D.L.T.D.M. Biochemical properties of glycosylation and characterization of a histidine acid phosphatase (phytase) expressed in Pichia pastoris. Protein Expr. Purif. 2014, 99, 43–49. [Google Scholar] [CrossRef]
- Kumar, K.; Patel, K.; Agrawal, D.C.; Khire, J.M. Insights into the unfolding pathway and identification of thermally sensitive regions of phytase from Aspergillus niger by molecular dynamics simulations. J. Mol. Model. 2015, 21. [Google Scholar] [CrossRef]
- Han, N.; Miao, H.; Yu, T.; Xu, B.; Yang, Y.; Wu, Q.; Zhang, R. Enhancing thermal tolerance of Aspergillus niger PhyA phytase directed by structural comparison and computational simulation. BMC Biotechnol. 2018, 18, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Zeng, M.; Wu, Z.F.; Chen, H.; Wang, H.N.; Wu, Q.; Shan, Z.; Han, X.Y. Improving Phytase Enzyme Activity in a Recombinant phyA Mutant Phytase from Aspergillus niger N25 by Error-Prone PCR. Appl. Biochem. Biotechnol. 2012, 166, 549–562. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Jin, W.; Sun, R.; Liao, Y.; Zhen, T.; Chen, H.; Wu, Q.; Gou, L. Improved thermostability and enzyme activity of a recombinant phyA mutant phytase from Aspergillus niger N25 by directed evolution and site- directed mutagenesis. Enzyme Microb. Technol. 2018, 108, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Fei, B.; Xu, H.; Cao, Y.; Ma, S. A multi-factors rational design strategy for enhancing the thermostability of Escherichia coli AppA phytase. J. Ind. Microbiol. Biotechnol. 2013, 40, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Fei, B.; Xu, H.; Zhang, F.; Li, X.; Ma, S.; Cao, Y.; Xie, J.; Qiao, D.; Cao, Y. Relationship between Escherichia coli AppA phytase’s thermostability and salt bridges. J. Biosci. Bioeng. 2013, 115, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Fei, B.; Cao, Y.; Xu, H.; Li, X.; Song, T.; Fei, Z.; Qiao, D.; Cao, Y. AppA C-terminal plays an important role in its thermostability in escherichia coli. Curr. Microbiol. 2013, 66, 374–378. [Google Scholar] [CrossRef]
- Xi, M.Y.; Wei, W. Improving the thermostability of Escherichia coli phytase, appA, by enhancement of glycosylation. Biotechnol. Lett. 2013, 35, 1669–1676. [Google Scholar] [CrossRef]
- Wu, T.H.; Chen, C.C.; Cheng, Y.S.; Ko, T.P.; Lin, C.Y.; Lai, H.L.; Huang, T.Y.; Liu, J.R.; Guo, R.T. Improving specific activity and thermostability of Escherichia coli phytase by structure-based rational design. J. Biotechnol. 2014, 175, 1–6. [Google Scholar] [CrossRef]
- Wang, X.; Yao, M.; Yang, B.; Fu, Y.; Hu, F.; Liang, A. Enzymology and thermal stability of phytase appA mutants. RSC Adv. 2015, 5, 43863–43872. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.; Gao, S.; Zhu, L.; Li, W.; Tian, X.; Liu, Y. Site-directed mutagenesis and thermal stability analysis of phytase from Escherichia coli. Biosci. Biotechnol. Res. Commun. 2016, 9, 357–365. [Google Scholar] [CrossRef]
- Wang, X.; Du, J.; Zhang, Z.Y.; Fu, Y.J.; Wang, W.M.; Liang, A.H. A rational design to enhance the resistance of Escherichia coli phytase appA to trypsin. Appl. Microbiol. Biotechnol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Fakhravar, A.; Hesampour, A. Rational design-based engineering of a thermostable phytase by site- directed mutagenesis. Mol. Biol. Rep. 2018. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, X.; Gai, Y.; Sun, Y.; Zhang, D. Evolution of E. coli Phytase for Increased Thermostability Guided by Rational Parameters. J. Microbiol. Biotechnol. 2019, 29, 419–428. [Google Scholar] [CrossRef] [Green Version]
- Farhat-Khemakhem, A.; Ali, M.B.; Boukhris, I.; Khemakhem, B.; Maguin, E.; Bejar, S.; Chouayekh, H. Crucial role of Pro 257 in the thermostability of Bacillus phytases: Biochemical and structural investigation. Int. J. Biol. Macromol. 2013, 54, 9–15. [Google Scholar] [CrossRef]
- Xu, W.; Wang, Z.; Shao, R. Site-directed Mutagenesis of a Neutral Phytase from Bacillus amyloliquefaciens: Influencing Activity and Stability. Adv. Mater. Res. 2014, 1034, 271–278. [Google Scholar] [CrossRef]
- Xu, W.; Shao, R.; Wang, Z.; Yan, X. Improving the Neutral Phytase Activity from Bacillus amyloliquefaciens DSM 1061 by Site-Directed Mutagenesis. Appl. Biochem. Biotechnol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Boukhris, I.; Farhat-khemakhem, A.; Blibech, M.; Bouchaala, K.; Chouayekh, H. Characterization of an extremely salt-tolerant and thermostable phytase from Bacillus amyloliquefaciens US573. Int. J. Biol. Macromol. 2015, 80, 581–587. [Google Scholar] [CrossRef]
- Chen, W.; Ye, L.; Guo, F.; Lv, Y.; Yu, H. Enhanced activity of an alkaline phytase from Bacillus subtilis 168 in acidic and neutral environments by directed evolution. Biochem. Eng. J. 2015, 98, 137–143. [Google Scholar] [CrossRef]
- Shivange, A.V.; Dennig, A.; Schwaneberg, U. Multi-site saturation by OmniChange yields a pH- and thermally improved phytase. J. Biotechnol. 2014, 170, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Shivange, A.V.; Roccatano, D.; Schwaneberg, U. Iterative key-residues interrogation of a phytase with thermostability increasing substitutions identified in directed evolution. Appl. Microbiol. Biotechnol. 2016, 100, 227–242. [Google Scholar] [CrossRef]
- Niu, C.; Luo, H.; Shi, P.; Huang, H.; Wang, Y.; Yang, P.; Yao, B. N-glycosylation improves the pepsin resistance of histidine acid phosphatase phytases by enhancing their stability at acidic pHs and reducing Pepsin’s Accessibility to its cleavage sites. Appl. Environ. Microbiol. 2016, 82, 1004–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, C.; Yang, P.; Huang, H.; Wang, Y.; Yao, B. Engineering of Yersinia Phytases to Improve Pepsin and Trypsin Resistance and Thermostability and Application Potential in the Food and Feed Industry. J. Agric. Food Chem. 2017. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Wu, X.; Xie, L.; Huang, Z. Identification and characterization of a mesophilic phytase highly resilient to high-temperatures from a fungus-garden associated metagenome. Appl. Microbiol. Biotechnol. 2015. [Google Scholar] [CrossRef]
- Tan, H.; Miao, R.; Liu, T.; Cao, X.; Wu, X.; Xie, L.; Huang, Z.; Peng, W.; Gan, B. Enhancing the thermal resistance of a novel acidobacteria-derived phytase by engineering of disulfide bridges. J. Microbiol. Biotechnol. 2016, 26, 1717–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravindran, V. Feed enzymes: The science, practice, and metabolic realities. J. Appl. Poult. Res. 2013, 22, 628–636. [Google Scholar] [CrossRef]
- Zhou, S.; Liu, Z.; Xie, W.; Yu, Y.; Ning, C.; Yuan, M.; Mou, H. Improving catalytic efficiency and maximum activity at low pH of Aspergillus neoniger phytase using rational design. Int. J. Biol. Macromol. 2019, 131, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
- Bryan, D.D.; Classen, H.L. In Vitro Methods of Assessing Protein Quality for Poultry. Animals 2020, 10, 551. [Google Scholar] [CrossRef] [Green Version]
- Niu, C.; Yang, P.; Luo, H.; Huang, H.; Wang, Y.; Yao, B. Engineering the residual side chains of HAP phytases to improve their pepsin resistance and catalytic efficiency. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Source | Phytase | Approaches | Location of Mutation | Contributing Factors in Thermostability | References |
|---|---|---|---|---|---|---|
| 1 | Aspergillus niger | PhyA | Computational design | Loops | H-bond, salt bridge | Noorbatchaet al., 2013 |
| 2 | Escherichia coli | AppA | Site-directed mutagenesis | α/β-domain | H-bond, Salt bridges | Fei et al., 2013 |
| 3 | Escherichia coli | AppA | Site-directed mutagenesis | Loops | Salt bridges | Fei et al., 2013 |
| 4 | Escherichia coli | AppA | Site-directed mutagenesis | α-helixes and concave surface between the two b-sheets | N-glycosylation | Yao et al., 2013 |
| 5 | Aspergillus niger N25 Phytase | PhyA | Site-directed mutagenesis | Loops | H-bond, salt bridge | Liao et al., 2013 |
| 6 | Escherichia coli | AppA | Site-directed mutagenesis | Deletion of C- terminal (coil including seven amino acids) | Deletion of C-terminal | Fei et al., 2013 |
| 7 | Bacillus subtilis US417 | PHYUS417 | Site-directed mutagenesis | Loop | Role of P257 | Farhat et al., 2013 |
| 8 | Aspergillus niger | PhyA | Site-directed mutagenesis | Loop | H-bond | Hesampour et al., 2014 |
| 9 | Yersinia mollaretii | Ymphytase | Multi-site saturation | Next to the active site loop, surface, α-domain, α/β-domain | H-bond, aromatic–aromatic interaction | Shivange et al., 2014 |
| 10 | Escherichia coli | AppA | Site-directed mutagenesis | Loops | N- glycosylation | Wu et al., 2014 |
| 11 | Aspergillus japonicus C03 | PhyA | Through expression in Pichia pastoris | - | N- glycosylation | Fonseca et al., 2014 |
| 12 | Bacillus amyloliquefaciens | PhyC | Site-directed mutagenesis | Surface of phytase | Replacement Asp with Glu | Xu et al., 2014 |
| 13 | Aspergillus niger | PhyA | Molecular dynamic simulations study | Loops are most fluctuated regions | H-bond, salt bridges | Kapil Kumar et al., 2015 |
| 14 | Bacillus amyloliquefaciens DSM 1061 | PhyC | Site-directed mutagenesis | Surface of phytase | Replacement Asp and Ser with Glu | Wei Xu et al., 2015 |
| 15 | Bacillus amyloliquefaciens US573 | PHY US573 | Compared to TS-Phy from B. amyloliquefaciens DS11 | Loop | Replacement Ala -346 with Val, increases rigidity of loop | Boukhris et al., 2015 |
| 16 | Insect-cultivated fungus gardens | PhyXT52 | Site-directed mutagenesis | - | Salt bridges, disulfide bonds | Tan et al., 2015 |
| 17 | Bacillus subtilis 168 | Phy168 | Directed evolution, site saturation mutagenesis | Loop | H-bond | Chen et al., 2015 |
| 18 | Escherichia coli | AppA | Multipoint site-directed mutagenesis | Phytase surface | N-glycosylation, increased hydrophobic surface | Wang et al., 2015 |
| 19 | Yersinia mollaretii | Ymphytase | Iterative Key- residues interrogation | Helix, close to active site, phytase surface | H-bonds, salt bridge | Shivange et al., 2016 |
| 20 | Acidobacteria | PhyA | Site-directed mutagenesis | Disulfide bridges | Tan et al., 2016 | |
| 21 | Yersinia kristensenii | YkAPPA | Site-directed mutagenesis | Exposed surface of phytase | N- glycosylation | Niu et al., 2016 |
| 22 | Escherichia coli | AppA | Site-directed mutagenesis | On the surface of the pocket edge | Replacement Lys with Glu | Zhang et al., 2016 |
| 23 | Yersinia kristensenii | YkAPPA | Site-directed mutagenesis | Phytase surface | Increasing the positive charge, decreasing residual size and hydrophobicity | Niu et al., 2017 |
| 24 | Aspergillus niger | PhyA | Site-directed mutagenesis | - | H-bond | Han et al., 2018 |
| 25 | Escherichia coli | AppA | Site-directed mutagenesis | Loop | H-bond | Wang et al., 2018 |
| 26 | Aspergillus niger N25 | phyA | Directed evolution, Site-directed mutagenesis | Loop, C terminus of an α-helix | H-bond, salt bridge | Tang et al., 2018 |
| 27 | Escherichia coli | AppA | Site-directed mutagenesis | - | Hydrophobic interactions | Fakhravar and Hesampour, 2018 |
| 28 | Escherichia coli | AppA | Iterative Mutagenesis | Surface and loops and β-sheet | Increased electrostatic interaction through introducing hydrophobic and aromatic amino acids | Li et al., 2019 |
| No. | Source | Phytase | Approaches | Location of Mutation | Contributing Factor in Acid Stability | References |
|---|---|---|---|---|---|---|
| 1 | Aspergillus neoniger | PhyAn | Site-directed mutagenesis | Catalytic motif | Positively charged residues | Zhou et al., 2019 |
| 2 | Escherichia coli | AppA | Site-directed mutagenesis | Surface of pocket edge | Decreased isoelectric point through substitution K24E | Zhang et al., 2016 |
| 3 | Yersinia mollaretii | Ymphytase | Directed evolution, multi-site saturation mutagenesis | Close to active site loop, surface area, α-domain, α/β-domain | Epistatic effect of four substitutions which introduced H-bonds and aromatic interactions | Shivange et al., 2014 |
| 4 | Yersinia kristensenii, Yersinia enterocolitica | YkAPPA, YeAPPA | Site-directed mutagenesis | Phytase surface | N-glycosylation | Niu et al., 2016 |
| 5 | Yersinia kristensenii | YkAPPA | Site-directed mutagenesis | Phytase surface | Decreased hydrophobicity, increased positive surface charge | Niu et al., 2017 |
| No. | Source | Phytase | Approaches | Location of Mutation | Contributing Factors in Proteolytic Stability | References |
|---|---|---|---|---|---|---|
| 1 | Yersinia kristensenii, Yersinia rohdei | YkAPPA, YrAPPA | Site-directed mutagenesis | Vicinity of N-glycosylation motifs | Substituting pepsin cleavage sites | Niu et al., 2016 |
| 2 | Yersinia kristensenii | YkAPPA | Site-directed mutagenesis | Phytase surface | Remove hydrophobic interactions phenyl F group and pepsin, increased positive surface charge | Niu et al., 2017 |
| 3 | Yersinia kristensenii, Yersinia enterocolitica | YkAPPA, YeAPPA | Site-directed mutagenesis | Phytase surface | Reducing pepsin cleavage sites | Niu et al., 2017 |
| 4 | Escherichia coli | AppA | Site-directed mutagenesis | Loops | Substitution of trypsin cleavage sites, H-bond, N-glycosylation | Wang et al., 2018 |
| No. | Source | Phytase | Approaches | Location of Mutations | Contributing Factors | Improved Properties | References |
|---|---|---|---|---|---|---|---|
| 1 | Aspergillus niger N25 | PhyA | Site-directed mutagenesis | Loop | Remodeling the active site through substituting longer side chain | Catalytic efficiency | Liao et al., 2013 |
| 2 | Aspergillus niger | PhyA | Site-directed mutagenesis | Far from active site, surface coil | Introducing positively charged polar amino acid (Arg), increase conformational flexibility | Catalytic efficiency | Hesampour et al., 2014 |
| 3 | Aspergillus niger N25 | phyA | Site-directed mutagenesis | Surface region | Electrostatic interactions improved conformational flexibility | Catalytic efficiency, specific activity | Tang et al., 2018 |
| 4 | Aspergillus neoniger | PhyAn | Site-directed mutagenesis | Edge of binding pocket | Surface charge modification | Catalytic efficiency | Zhou et al., 2019 |
| 5 | Escherichia coli | EcAppA | Site-directed mutagenesis | - | Stabilized loop near to binding pocket | Specific activity | Wu et al., 2014 |
| 6 | Escherichia coli | AppA | Site-directed mutagenesis | Supporting point on the surface of the pocket edge | Electrostatic interaction | Catalytic efficiency | Zhang et al., 2016 |
| 7 | Escherichia coli | AppA | Site-directed mutagenesis | - | Hydrophobic interaction | Catalytic efficiency | Fakhravar and Hesampour, 2018 |
| 8 | Bacillus amyloliquefaciens | PhyC | Site-directed mutagenesis | Surface of phytase | Surface flexibility | Specific activity | Xu et al., 2014 |
| 9 | Bacillus amyloliquefaciens DSM 1061 | PhyC | Site-directed mutagenesis | Surface of phytase | Reduced free energies of activation | Catalytic efficiency, specific activity | Xu et al., 2015 |
| 10 | Bacillus subtilis 168 | Phy168 | Directed evolution | Loops close to active site | Synergistic effects between mutations, broadening the size of active pocket, increase positive charge density, removing steric hindrance for phytate binding | Catalytic efficiency, specific activity | Chen et al., 2015 |
| 11 | Yersinia kristensenii | YkAPPA | Site-directed mutagenesis | Surface of phytase | Substitutions with larger steric space in the catalytic pocket | Catalytic efficiency | Niu et al., 2017 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nezhad, N.G.; Raja Abd Rahman, R.N.Z.; Normi, Y.M.; Oslan, S.N.; Shariff, F.M.; Leow, T.C. Integrative Structural and Computational Biology of Phytases for the Animal Feed Industry. Catalysts 2020, 10, 844. https://0-doi-org.brum.beds.ac.uk/10.3390/catal10080844
Nezhad NG, Raja Abd Rahman RNZ, Normi YM, Oslan SN, Shariff FM, Leow TC. Integrative Structural and Computational Biology of Phytases for the Animal Feed Industry. Catalysts. 2020; 10(8):844. https://0-doi-org.brum.beds.ac.uk/10.3390/catal10080844
Chicago/Turabian StyleNezhad, Nima Ghahremani, Raja Noor Zaliha Raja Abd Rahman, Yahaya M. Normi, Siti Nurbaya Oslan, Fairolniza Mohd Shariff, and Thean Chor Leow. 2020. "Integrative Structural and Computational Biology of Phytases for the Animal Feed Industry" Catalysts 10, no. 8: 844. https://0-doi-org.brum.beds.ac.uk/10.3390/catal10080844