
Comparison of Spatial Structures and Packaging of Phosphorybosil Pyrophosphate Synthetase 2 from Thermus thermophilus HB27 in Rhombohedral and Tetragonal Crystals

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
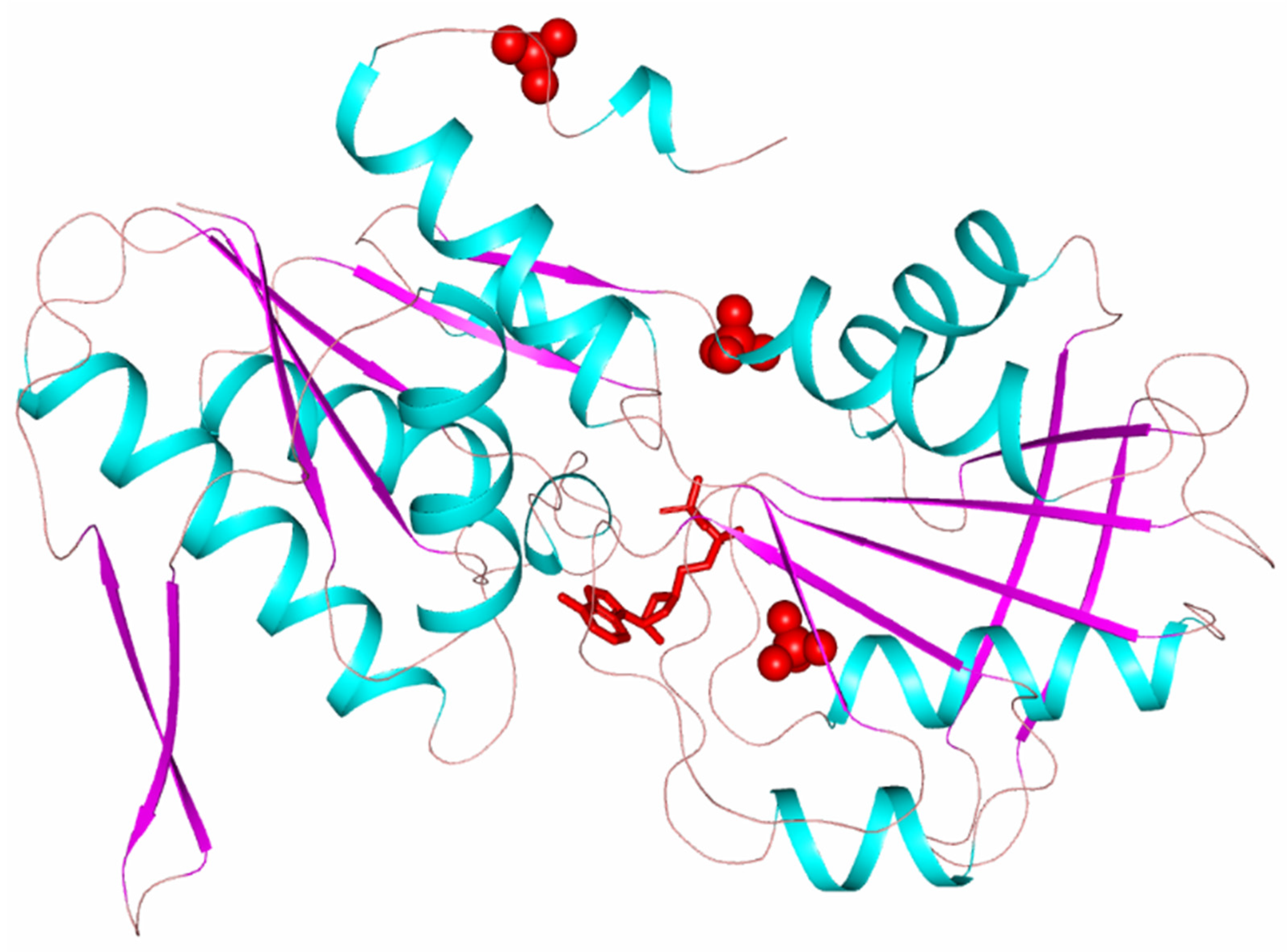
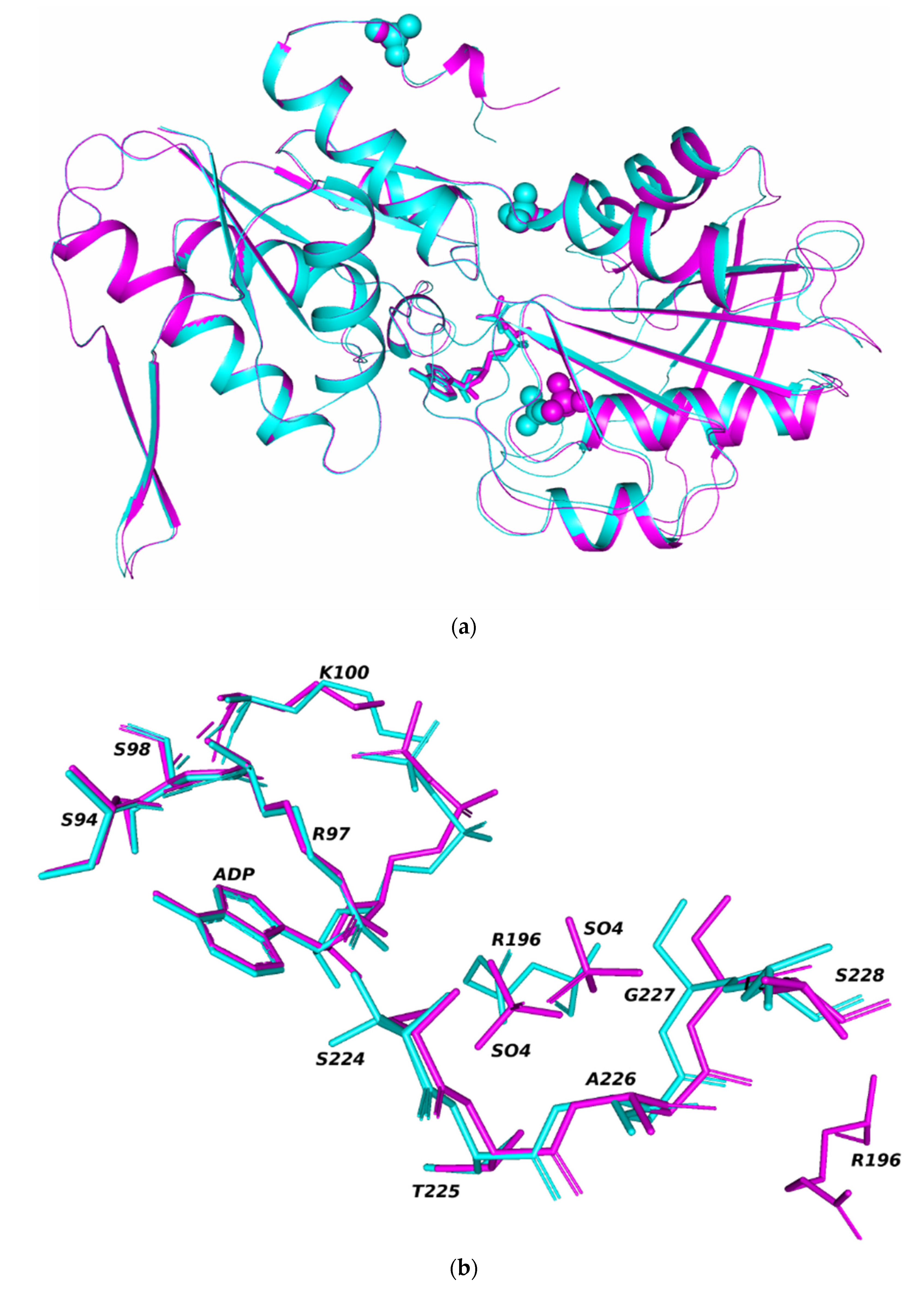
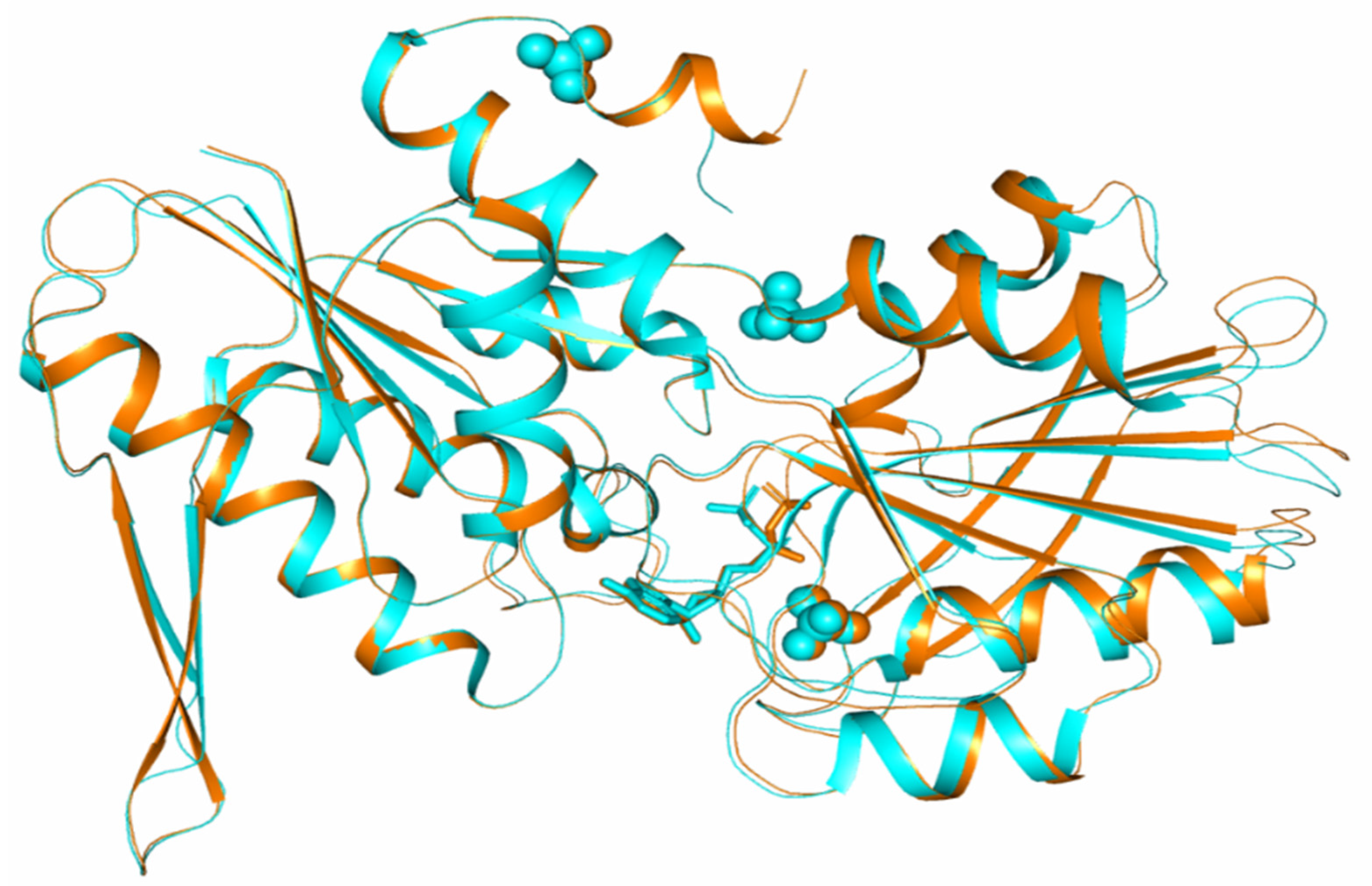
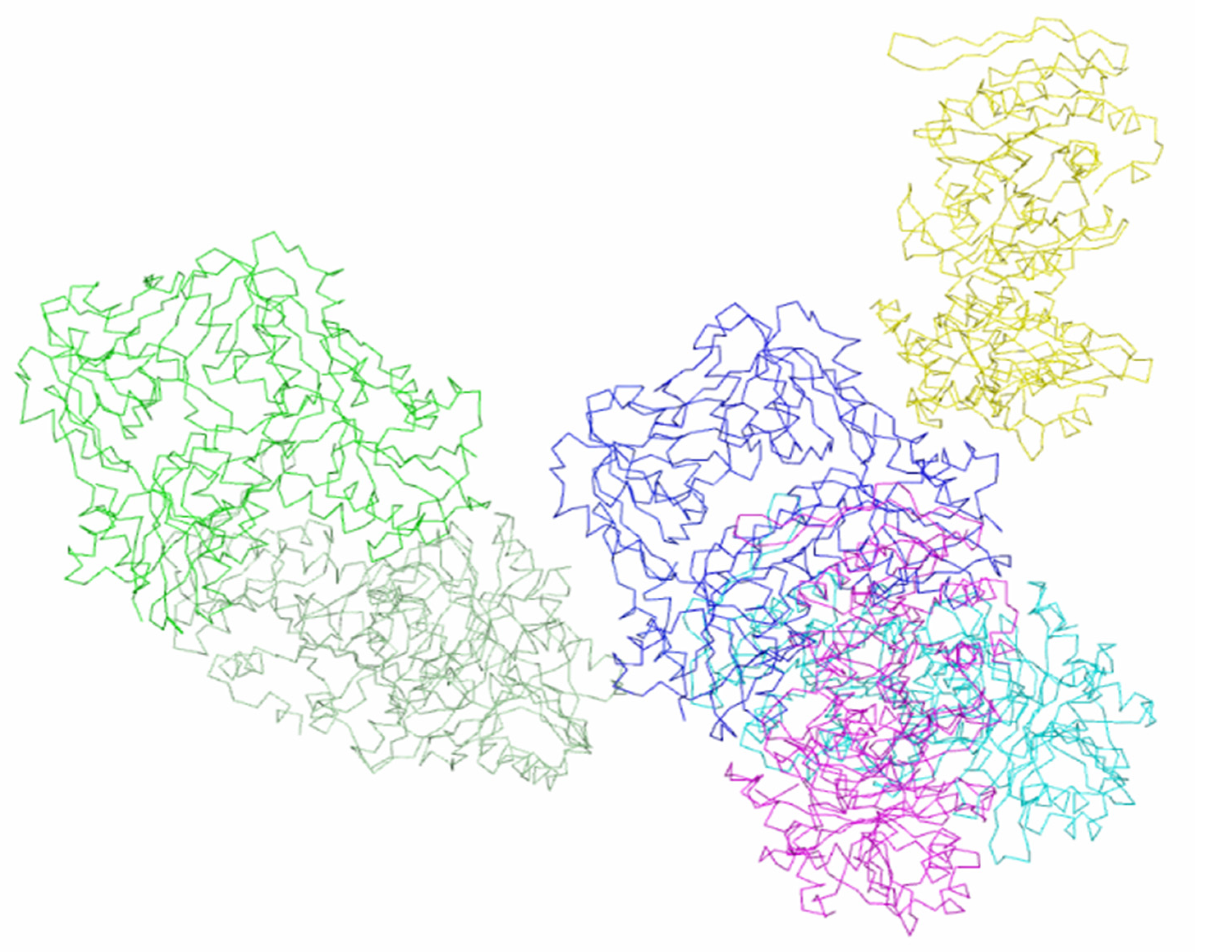
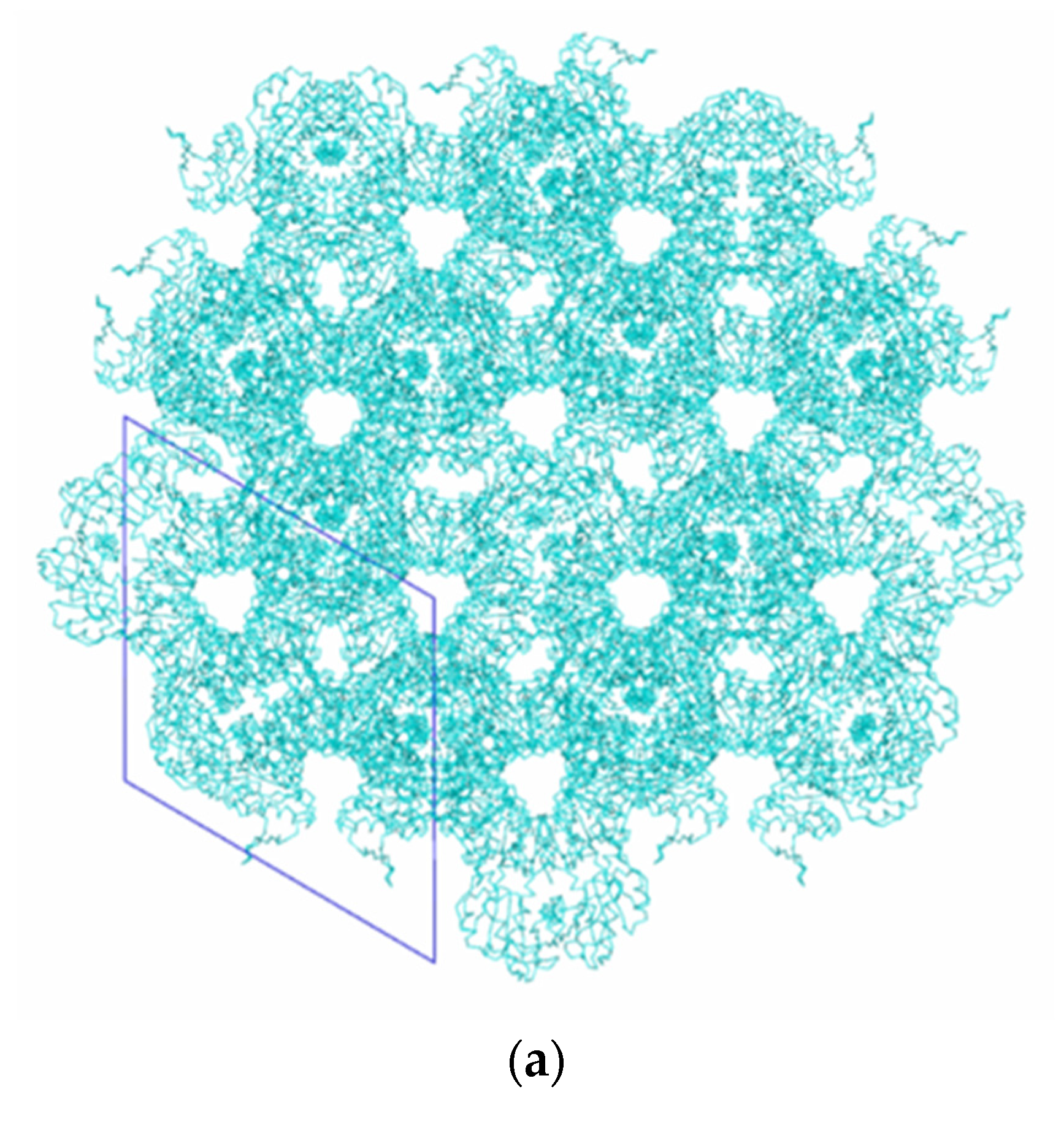
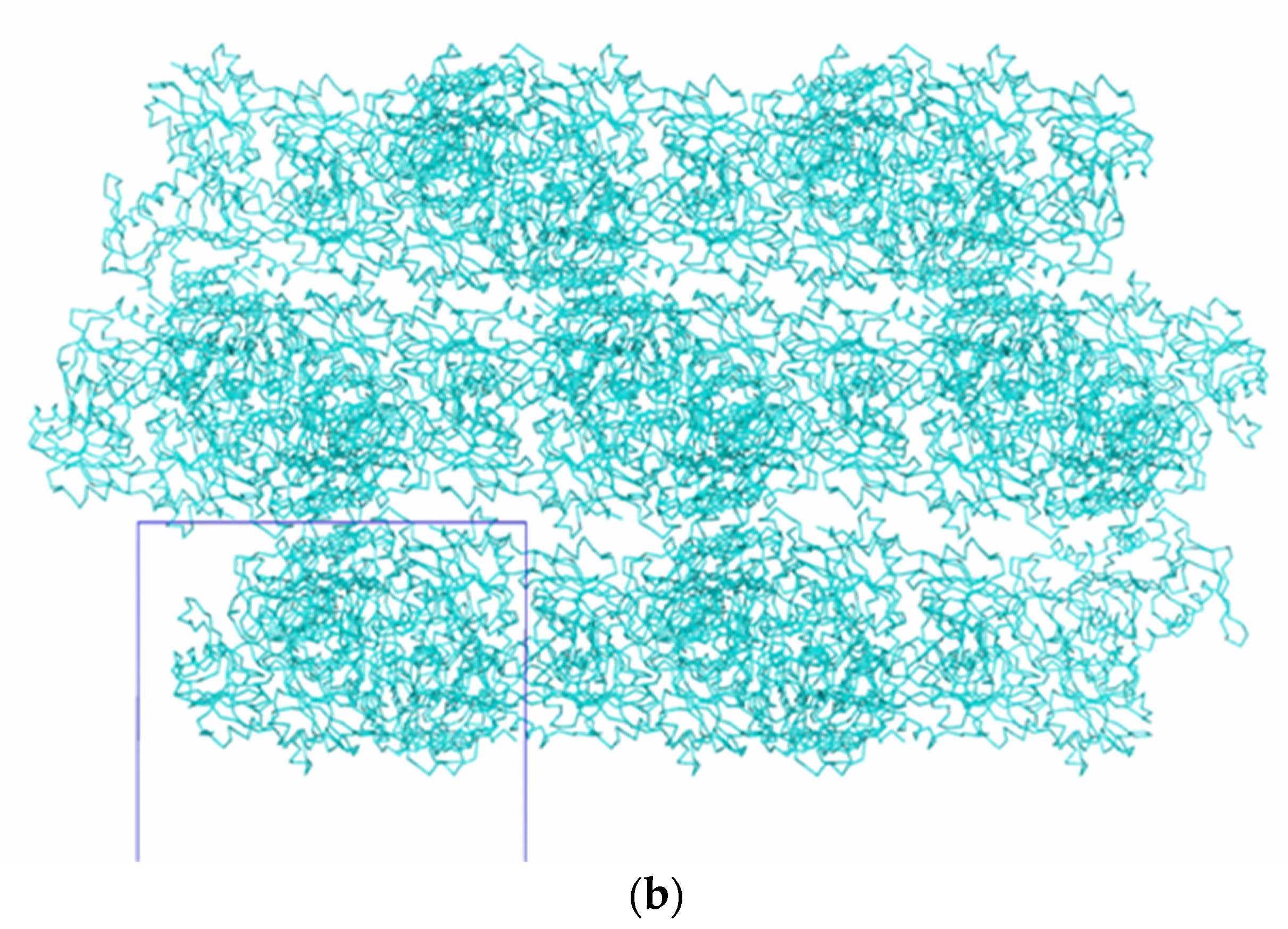
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Switzer, R.L. Regulation and mechanism of phosphoribosylpyrophosphate synthetase. I. Purification and properties of the enzyme from Salmonella typhimurium. J. Biol. Chem. 1969, 244, 2854–2863. [Google Scholar] [CrossRef]
- Becker, M.A.; Raivio, K.O.; Bakay, B.; Adams, W.B.; Nyhan, W.L. Variant human phosphoribosylpyrophosphate synthetase altered in regulatory and catalytic functions. J. Clin. Investig. 1980, 65, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Esipov, R.S.; Abramchik, Y.A.; Fateev, I.V.; Konstantinova, I.D.; Kostromina, M.A.; Muravyova, T.I.; Artemova, K.G.; Miroshnikov, A.I. A Cascade of Thermophilic Enzymes As an Approach to the Synthesis of Modified Nucleotides. Acta Nat. 2016, 8, 82–90. [Google Scholar] [CrossRef]
- Fateev, I.V.; Kostromina, M.A.; Abramchik, Y.A.; Eletskaya, B.Z.; Mikheeva, O.O.; Lukoshin, D.D.; Zayats, E.A.; Berzina, M.Y.; Dorofeeva, E.V.; Paramonov, A.S.; et al. Multi-Enzymatic Cascades in the Synthesis of Modified Nucleosides: Comparison of the Thermophilic and Mesophilic Pathways. Biomolecules 2021, 11, 586. [Google Scholar] [CrossRef] [PubMed]
- Hove-Jensen, B. Mutation in the phosphoribosylpyrophosphate synthetase gene (prs) that results in simultaneous requirements for purine and pyrimidine nucleosides, nicotinamide nucleotide, histidine, and tryptophan in Escherichia coli. J. Bacteriol. 1988, 170, 1148–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hove-Jensen, B. Phosphoribosylpyrophosphate (PRPP)-less mutants of Escherichia coli. Mol. Microbiol. 1989, 3, 1487–1492. [Google Scholar] [CrossRef]
- Hove-Jensen, B.; Nygaard, P. Phosphoribosylpyrophosphate synthetase of Escherichia coli, Identification of a mutant enzyme. Eur. J. Biochem. 1982, 126, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Switzer, R.L.; Sogin, D.C. Binding of the substrates and the allosteric inhibitor adenosine 5′-diphosphate to phosphoribosylpyrophosphate synthetase from Salmonella typhimurium. J. Biol. Chem. 1973, 248, 1063–1073. [Google Scholar] [CrossRef]
- Becker, M.A.; Kostel, P.J.; Meyer, L.J. Human erythrocyte phosphoribosylpyrophosphate synthetase. Subunit analysis and states of subunit association. J. Biol. Chem. 1975, 250, 6822–6830. [Google Scholar] [CrossRef]
- Gibson, K.J.; Schubert, K.R.; Switzer, R.L. Binding of the substrates and the allosteric inhibitor adenosine 5′-diphosphate to phosphoribosylpyrophosphate synthetase from Salmonella typhimurium. J. Biol. Chem. 1982, 257, 2391–2396. [Google Scholar] [CrossRef]
- Krath, B.N.; Hove-Jensen, B. Implications of secondary structure prediction and amino acid sequence comparison of class I and class II phosphoribosyl diphosphate synthases on catalysis, regulation, and quaternary structure. Protein Sci. 2001, 10, 2317–2324. [Google Scholar] [CrossRef]
- Li, S.; Lu, Y.; Peng, B.; Ding, J. Crystal structure of human phosphoribosylpyrophosphate synthetase 1 reveals a novel allosteric site. Biochem. J. 2007, 401, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Kadziola, A.; Jepsen, C.H.; Johansson, E.; McGuire, J.; Larsen, S.; Hove-Jensen, B. Novel class III phosphoribosyl diphosphate synthase: Structure and properties of the tetrameric, phosphate-activated, non-allosterically inhibited enzyme from Methanocaldococcus jannaschii. J. Mol. Biol. 2005, 354, 815–828. [Google Scholar] [CrossRef]
- Esipov, R.S.; Abramchik, Y.A.; Fateev, I.V.; Muravyova, T.I.; Artemova, K.G.; Konstantinova, I.D.; Kuranova, I.P.; Miroshnikov, A.I. Recombinant phosphoribosyl pyrophosphate synthetases from Thermus thermophilus HB27: Isolation and properties. Bioorg. Chem. 2016, 42, 512–521. [Google Scholar] [CrossRef]
- Timofeev, V.I.; Sinitsyna, E.V.; Kostromina, M.A.; Muravieva, T.I.; Makarov, D.A.; Mikheeva, O.O.; Kuranova, I.P.; Esipov, R.S. Crystal structure of recombinant phosphoribosylpyrophosphate synthetase 2 from Thermus thermophilus HB27 complexed with ADP and sulfate ions. Acta Cryst. 2017, F73, 369–375. [Google Scholar] [CrossRef]
- Timofeev, V.I.; Smirnova, E.A.; Chupova LAEsipov, R.S.; Kuranova, I.P. X-ray study of the conformational changes in the molecule of phosphopantetheine adenylyltransferase from Mycobacterium tuberculosis during the catalyzed reaction. Acta Cryst. 2012, D68, 1660–1670. [Google Scholar] [CrossRef]
- Takahashi, S.; Tsurumura, T.; Aritake, K.; Furubayashi, N.; Sato, M.; Yamanaka, M.; Hirota, E.; Sano, S.; Kobayashi, T.; Tanaka, T.; et al. High-quality crystals of human haematopoietic prostaglandin D synthase with novel inhibitors. Acta Cryst. 2010, F66, 846–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuranova, I.P.; Smirnova, E.A.; Abramchik, Y.A.; Timofeev, V.I.; Kovalchuk, M.V.; Chupova, L.A.; Esipov, R.S.; Akparov, V.K. Crystal growth of phosphopantetheine adenylyltransferase, carboxypeptidase t, and thymidine phosphorylase on the international space station by the capillary counter-diffusion method. Crystallogr. Rep. 2011, 56, 884–891. [Google Scholar] [CrossRef]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J.J. Phaser (CCP4: Supported Program) NAME phaser-2.5. 0-Maximum Likelihood Analysis and Phasing. SYNOPSIS phaser. Appl. Cryst. 2007, 40, 658–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eriksen, T.A.; Kadziola, A.; Bentsen, A.K.; Harlow, K.W.; Larsen, S. Structural basis for the function of Bacillus subtilis phosphoribosyl-pyrophosphate synthetase. Nat. Struct. Biol. 2000, 7, 303–308. [Google Scholar] [CrossRef]
- Murshudov, G.N.; Skubák, P.; Lebedev, A.A.; Pannu, N.S.; Steiner, R.A.; Nicholls, R.A.; Winn, M.D.; Long, F.; Vagin, A.A. REFMAC5 for the refinement of macromolecular crystal structures. Acta Cryst. 2011, D67, 355–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrodinger, L.L.C. The PyMOL Molecular Graphics System; Version 1.8; Schrodinger, L.L.C: New York, NY, USA, 2015. [Google Scholar]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef] [PubMed]
- Matthews, B.W. Solvent content of protein crystals. J. Mol. Biol. 1968, 33, 491–497. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value |
|---|---|
| Space group | R32-h |
| a, c, Å | 106.29, 333.29 |
| α, γ, ° | 90, 120 |
| Resolution, Å | 20.0−1.85 (1.89−1.85) * |
| Number of independent reflexes | 59412 |
| Completeness, % | 100 (99.8) |
| Redundancy | 7.01 (5.96) |
| I/σ(I) | 14.31 (3.53) |
| CC1/2 | 0.999 (0.905) |
| Rmrgd–F, % | 10.5 (54.6) |
| PDB_ID | 7PN0 |
| Rcryst, % | 18.7 (28.5) |
| Rfree, % | 22.1 (31.9) |
| Number of refined atoms | |
| Protein | 4662 |
| Solvent | 257 |
| Ligands | 84 |
| Average B-Factor, Å2 | 23.331 |
| rmsd | |
| Bonds, Å | 0.011 |
| Angles, ° | 1.795 |
| Ramachandran plot | |
| The most favorable areas | 97% |
| Acceptable areas | 3% |
| Tetragonal Crystals, Space Group P41212 (PDB_ID: 5T3O) | ||||
|---|---|---|---|---|
| Atom (x, y, z) | Secondary Structure Elements | Distance, Å | Atom (x−1/2, −y−1/2, −z−1/4) | Secondary Structure Elements |
| N_THR285/ B | β14 | 3.00 | O_ALA23/C | α1 |
| NH1_ARG184/B | α5 | 3.15 | OG1_THR158/C | α4 |
| ND2_ASN154/B | α4 | 2.93 | O_THR287/C | L |
| NH1_ARG284/B | β14 | 2.89 | OE1_GLU294/C | α8 |
| NH2_ARG159/B | α4 | 3.78 | OE2_GLU294/C | α8 |
| O_SER307/B | η4 | 3.89 | NH2_ARG284/C | β14 |
| O_SER307/B | η4 | 3.20 | NH1_ARG159/C | α4 |
| OG_SER307/B | η4 | 3.84 | NH2_ARG284/C | β14 |
| O_LEU308/B | η4 | 3.02 | NH2_ARG159/C | α4 |
| Rhombohedral crystals, space group R32 h (PDB_ID: 7PN0) | ||||
| Atom (x, y, z) | Secondary structure elements | Distance, Å | Atom (−x-2/3, −x + y−1/3, −z−1/3) | Secondary structure elements |
| NH1_ARG159/A | α4 | 3.99 | OE1_GLU303/A | L |
| NH1_ARG159/A | α4 | 3.75 | OE2_GLU303/A | L |
| NE_ARG284/A | β14 | 3.65 | OE1_GLU294/A | α8 |
| NE_ARG284/A | β14 | 3.17 | OE2_GLU294/A | α8 |
| NH1_ARG284/A | β14 | 3.99 | OE1_GLU303/A | L |
| NH2_ARG284/A | β14 | 3.23 | OE2_GLU294/A | α8 |
| NH2_ARG284/A | β14 | 3.02 | OE1_GLU303/A | L |
| OG1_THR285/A | β14 | 3.69 | O_ALA23/A | α1 |
| NH1_ARG298/A | α8 | 2.84 | O_THR158/A | α4 |
| NH2_ARG298/A | α8 | 3.17 | OG1_THR158/A | α4 |
| OE2_GLU303/A | L | 3.75 | NH1_ARG159/A | α4 |
| OE1_GLU303/A | L | 3.99 | NH1_ARG159/A | α4 |
| OE2_GLU294/A | α8 | 3.17 | NE_ARG284/A | β14 |
| OE1_GLU294/A | α8 | 3.65 | NE_ARG284/A | β14 |
| OE2_GLU294/A | α8 | 3.23 | NH2_ARG284/A | β14 |
| OE1_GLU303/A | L | 3.99 | NH1_ARG284/A | β14 |
| OE1_GLU303/A | L | 3.02 | NH2_ARG284/A | β14 |
| O_ALA23/A | α1 | 3.69 | OG1_THR285/A | β14 |
| O_THR158/A | α4 | 2.84 | NH1_ARG298/A | α8 |
| OG1_THR158/A | α4 | 3.17 | NH2_ARG298/A | α8 |
| Molecules (x, y, z) | Molecule (y, x, −z) | |||
| NH2_ARG298/B | α8 | 2.83 | OD1_ASN154/B | α4 |
| NE1_TRP297/B | α8 | 3.30 | O_THR158/B | α4 |
| NE_ARG298/B | α8 | 3.33 | OG1_THR158/B | α4 |
| NH2_ARG301/B | α8 | 3.46 | O_ARG159/B | α4 |
| OG1_THR158/B | α4 | 3.30 | OE2_GLU294/B | α8 |
| OD1_ASN154/B | α4 | 2.83 | NH2_ARG298/B | α8 |
| O_THR158/B | α4 | 3.30 | NE1_TRP297/B | α8 |
| OG1_THR158/B | α4 | 3.33 | NE_ARG298/B | α8 |
| O_ARG159/B | α4 | 3.46 | NH2_ARG301/B | α8 |
| OE2_GLU294/B | α8 | 3.30 | OG1_THR158/B | α4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abramchik, Y.; Zayats, E.; Kostromina, M.; Lykoshin, D.; Fateev, I.; Konstantinova, I.; Zhukhlistova, N.; Timofeev, V.; Kuranova, I.; Esipov, R. Comparison of Spatial Structures and Packaging of Phosphorybosil Pyrophosphate Synthetase 2 from Thermus thermophilus HB27 in Rhombohedral and Tetragonal Crystals. Crystals 2021, 11, 1128. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst11091128
Abramchik Y, Zayats E, Kostromina M, Lykoshin D, Fateev I, Konstantinova I, Zhukhlistova N, Timofeev V, Kuranova I, Esipov R. Comparison of Spatial Structures and Packaging of Phosphorybosil Pyrophosphate Synthetase 2 from Thermus thermophilus HB27 in Rhombohedral and Tetragonal Crystals. Crystals. 2021; 11(9):1128. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst11091128
Chicago/Turabian StyleAbramchik, Yulia, Evgeniy Zayats, Maria Kostromina, Dmitry Lykoshin, Ilya Fateev, Irina Konstantinova, Nadezda Zhukhlistova, Vladimir Timofeev, Inna Kuranova, and Roman Esipov. 2021. "Comparison of Spatial Structures and Packaging of Phosphorybosil Pyrophosphate Synthetase 2 from Thermus thermophilus HB27 in Rhombohedral and Tetragonal Crystals" Crystals 11, no. 9: 1128. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst11091128