
Activity of Cerebellar Nuclei Neurons Correlates with ZebrinII Identity of Their Purkinje Cell Afferents

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Surgery for In Vivo Awake Recordings
2.3. In Vivo Awake Extracellular Recordings
2.4. Slice Preparation for In Vitro Whole-Cell Recordings
2.5. In Vitro Whole-Cell Recordings
2.6. Measurement of In Vitro Electrophysiological Parameters
2.7. Immunofluorescence for In Vitro Recordings
2.8. Fluorescence Microscopy for 2D Sholl Analysis
2.9. Immunofluorescence for In Vivo Recordings
2.10. Acquisition of Confocal Images
2.11. Spike Analysis
2.12. Statistical Analysis
3. Results
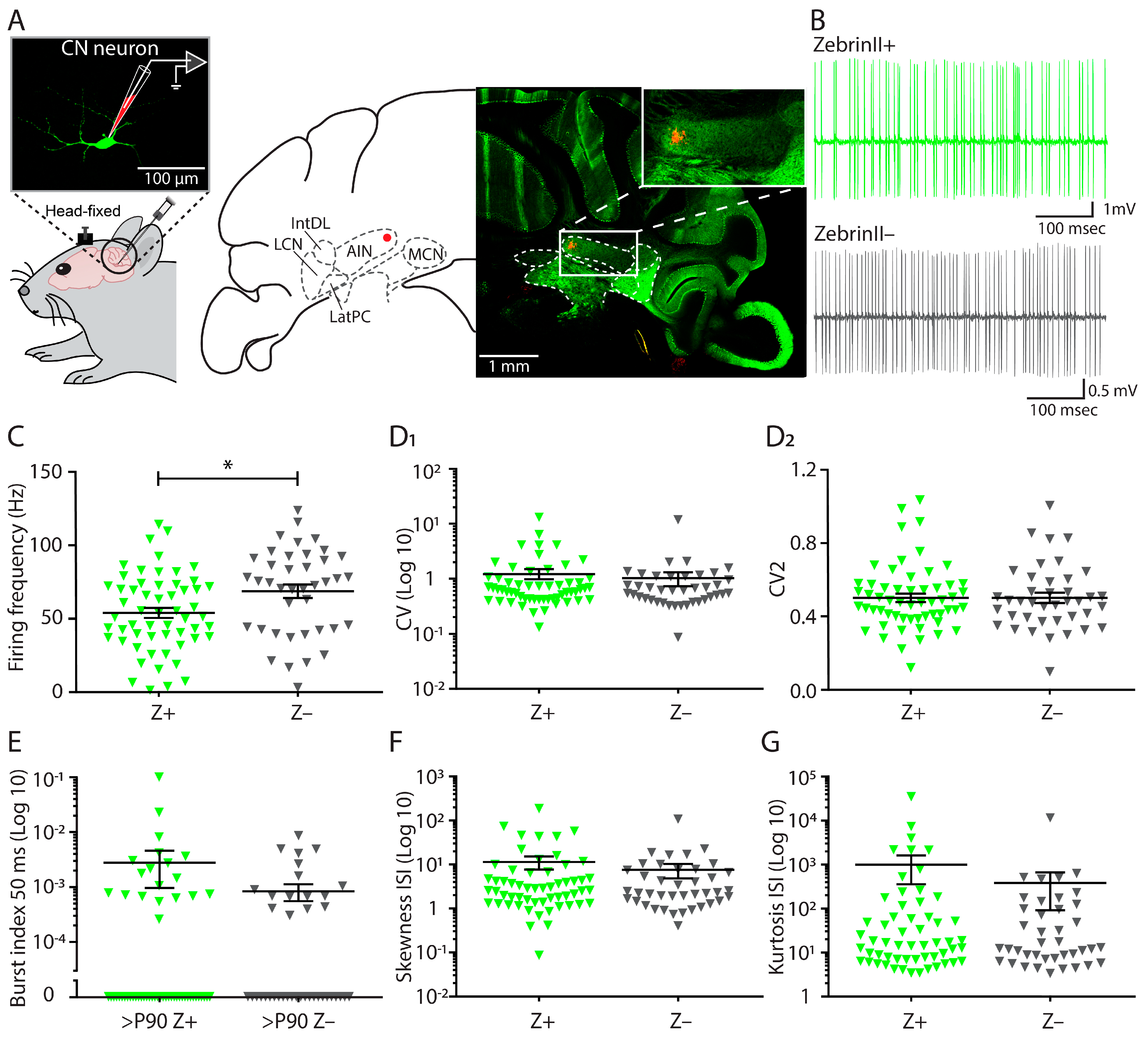
3.1. Firing Frequency Differs between ZebrinII Domains in Cerebellar Nuclei during Adulthood
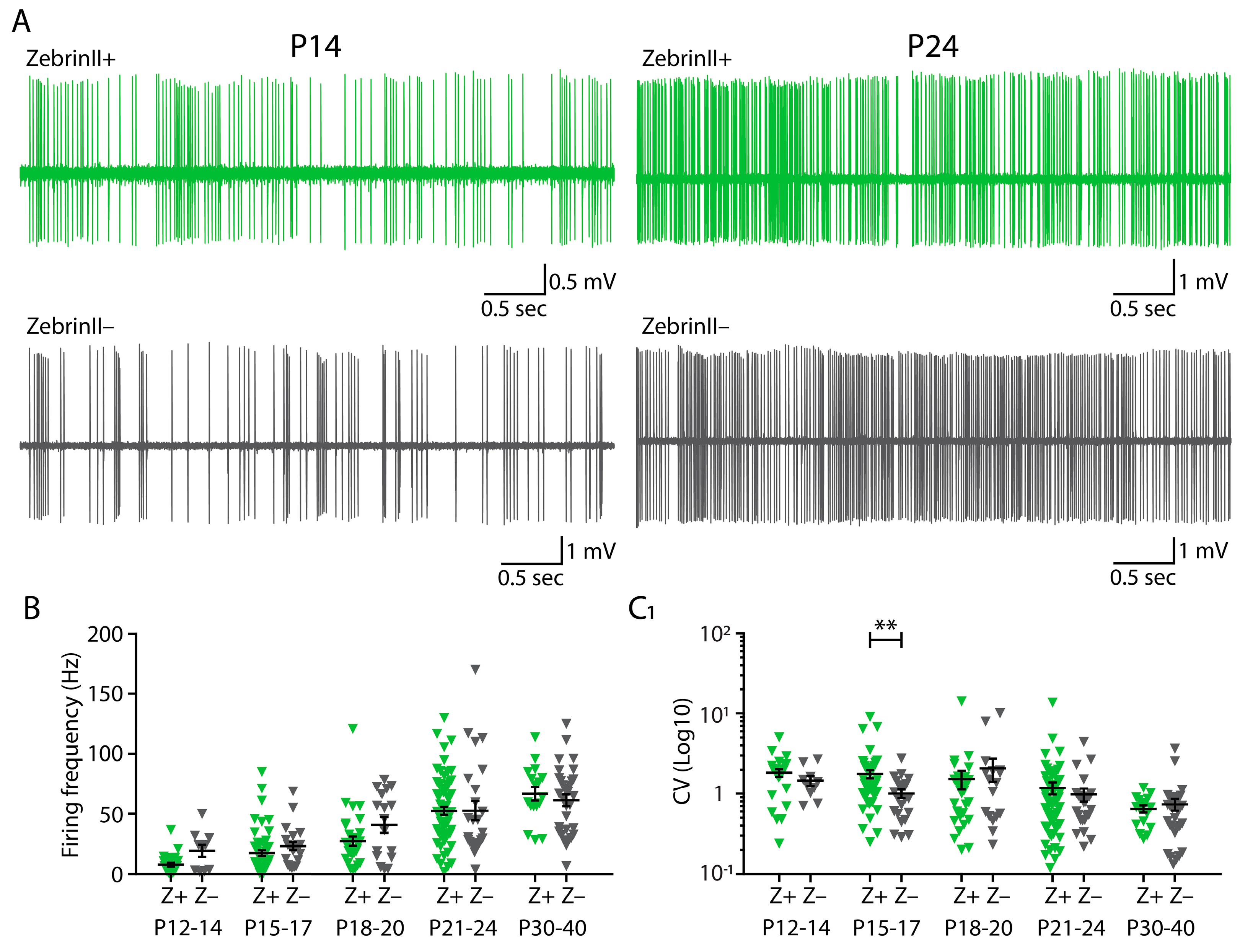
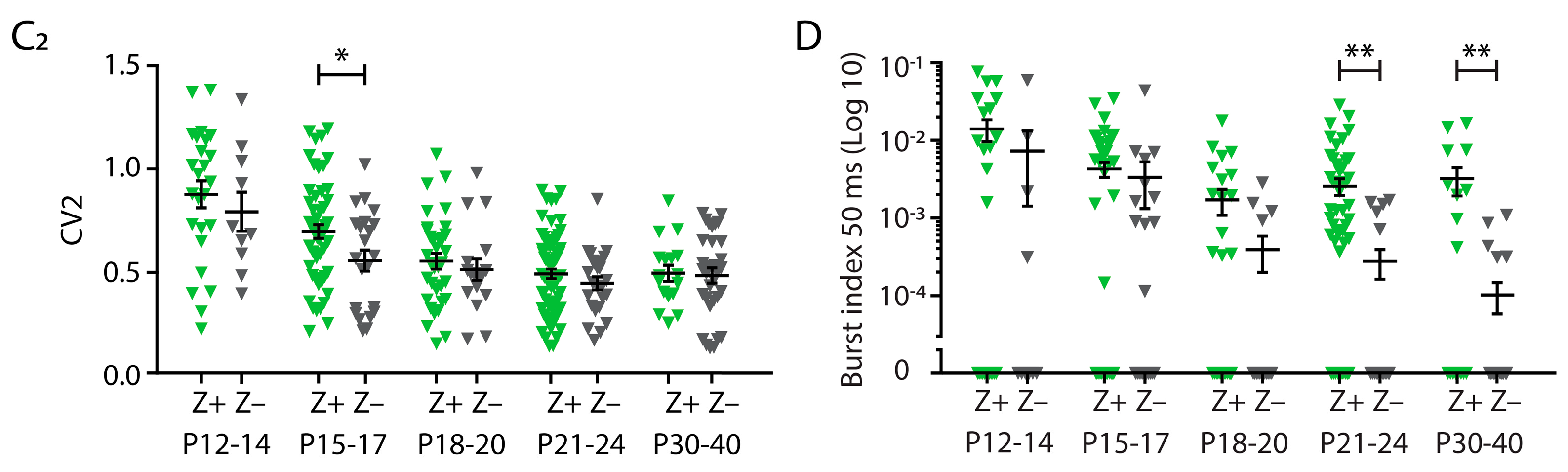
3.2. No Differences in Firing Frequency between ZebrinII Domains in Cerebellar Nuclei (CN) during Development
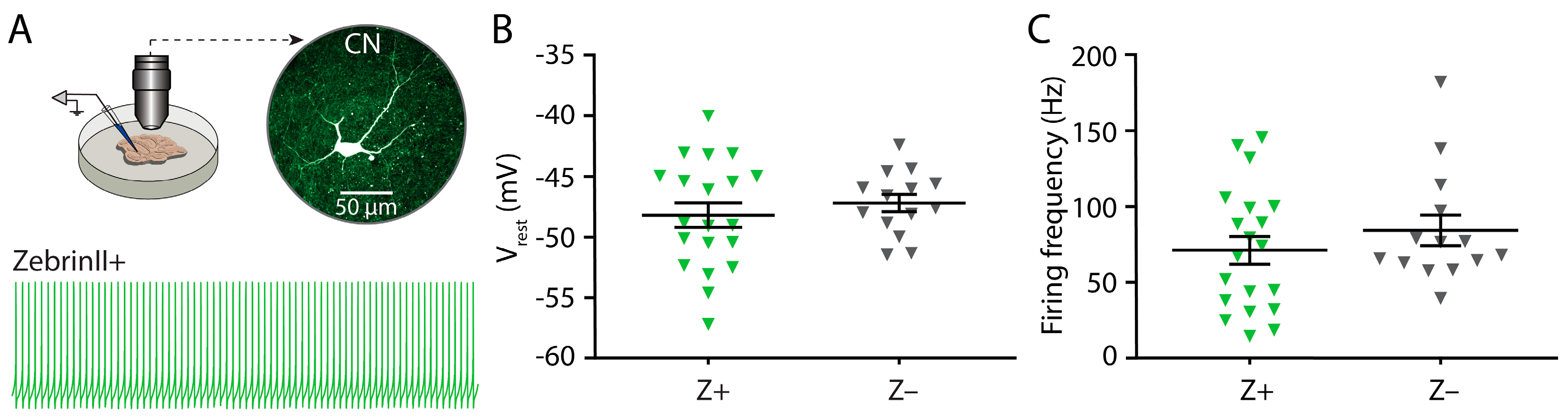
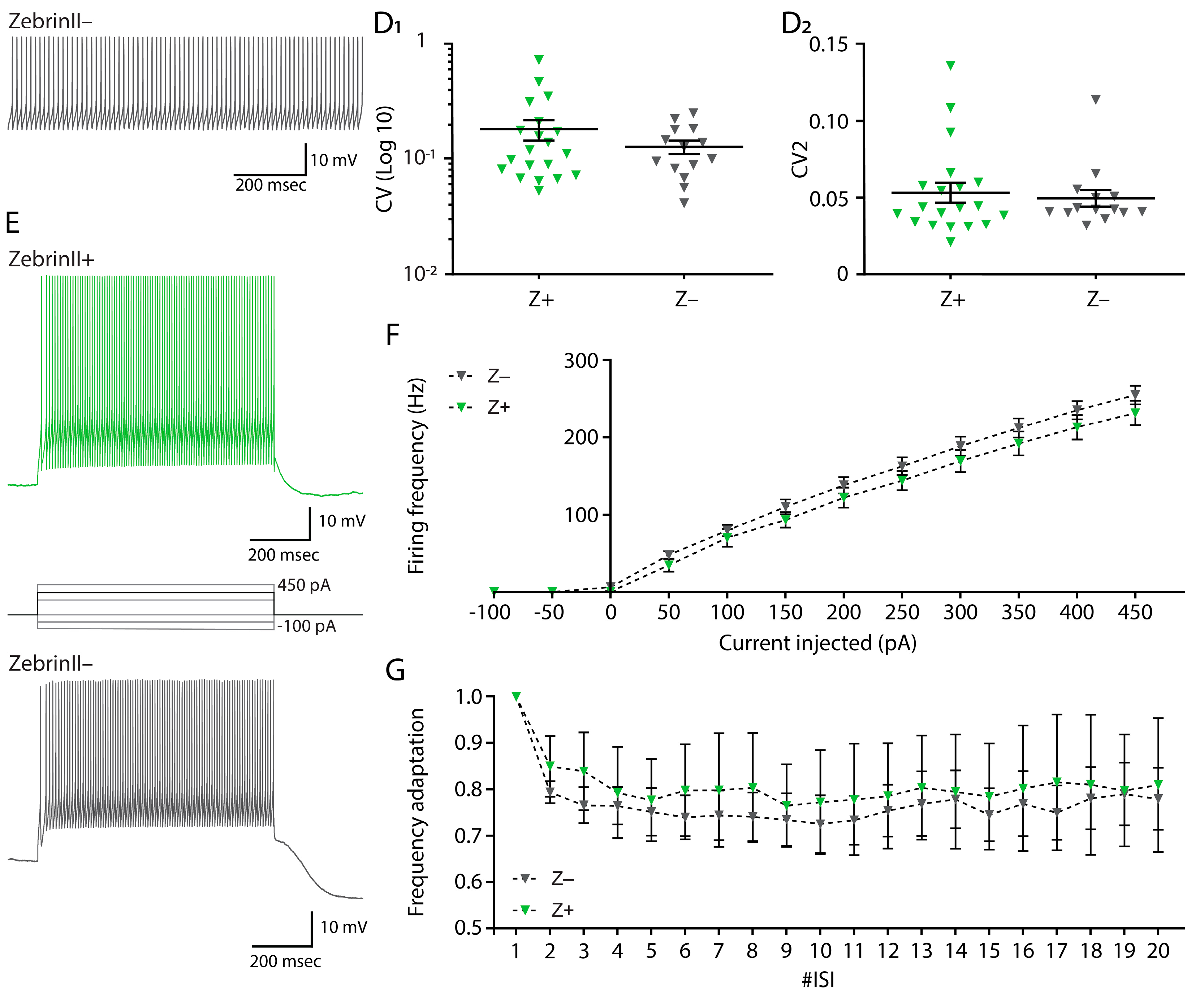
3.3. Similar Excitability and Pacemaker Activity of Cerebellar Nuclei Neurons in ZebrinII Domains
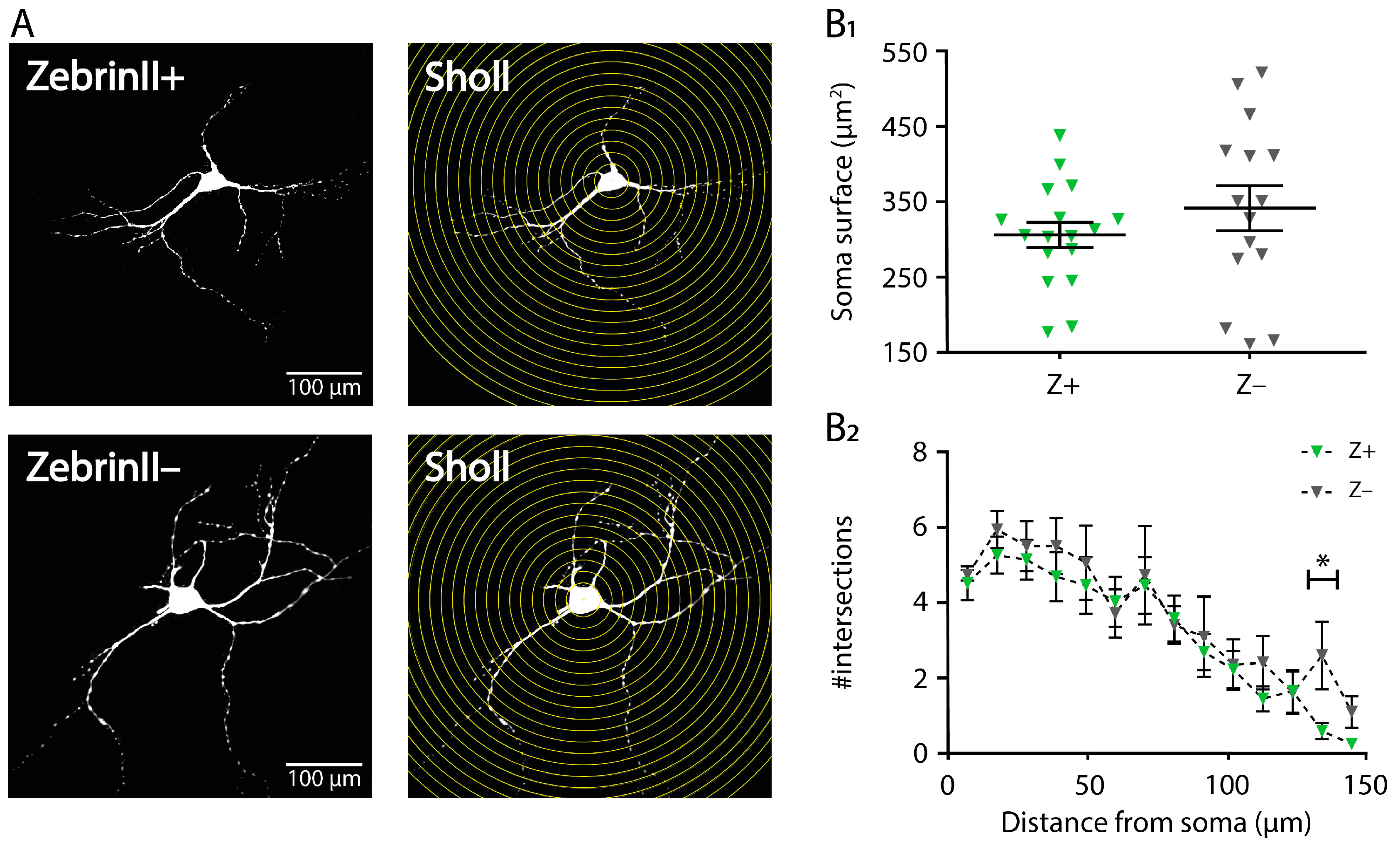
3.4. Morphology of Cerebellar Nuclei Neurons in ZebrinII Domains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stoodley, C.J.; Schmahmann, J.D. The cerebellum and language: Evidence from patients with cerebellar degeneration. Brain Lang. 2009, 110, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Sathyanesan, A.; Zhou, J.; Scafidi, J.; Heck, D.H.; Sillitoe, R.V.; Gallo, V. Emerging connections between cerebellar development, behaviour and complex brain disorders. Nat. Rev. Neurosci. 2019, 20, 298–313. [Google Scholar] [CrossRef]
- De Zeeuw, C.I.; Ten Brinke, M.M. Motor learning and the cerebellum. Cold Spring Harb. Perspect. Biol. 2015, 7, a021683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raman, I.M.; Gustafson, A.E.; Padgett, D. Ionic currents and spontaneous firing in neurons isolated from the cerebellar nuclei. J. Neurosci. 2000, 20, 9004–9016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garin, N.; Escher, G. The development of inhibitory synaptic specializations in the mouse deep cerebellar nuclei. Neuroscience 2001, 105, 431–441. [Google Scholar] [CrossRef]
- Person, A.L.; Raman, I.M. Purkinje neuron synchrony elicits time-locked spiking in the cerebellar nuclei. Nature 2011, 481, 502–505. [Google Scholar] [CrossRef]
- Hoebeek, F.E.; Witter, L.; Ruigrok, T.J.; De Zeeuw, C.I. Differential olivo-cerebellar cortical control of rebound activity in the cerebellar nuclei. Proc. Natl. Acad. Sci. USA 2010, 107, 8410–8415. [Google Scholar] [CrossRef] [Green Version]
- Thach, W.T. Discharge of purkinje and cerebellar nuclear neurons during rapidly alternating arm movements in the monkey. J. Neurophysiol. 1968, 31, 785–797. [Google Scholar] [CrossRef]
- Ohmae, S.; Uematsu, A.; Tanaka, M. Temporally specific sensory signals for the detection of stimulus omission in the primate deep cerebellar nuclei. J. Neurosci. 2013, 33, 15432–15441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antziferova, L.I.; Arshavsky, Y.I.; Orlovsky, G.N.; Pavlova, G.A. Activity of neurons of cerebellar nuclei during fictitious scratch reflex in cat. I. Fastigial nucleus. Brain Res. 1980, 200, 239–248. [Google Scholar] [CrossRef]
- Armstrong, D.M.; Edgley, S.A. Discharges of nucleus interpositus neurones during locomotion in the cat. J. Physiol. 1984, 351, 411–432. [Google Scholar] [CrossRef]
- Ten Brinke, M.M.; Boele, H.J.; Spanke, J.K.; Potters, J.W.; Kornysheva, K.; Wulff, P.; Ijpelaar, A.C.; Koekkoek, S.K.; De Zeeuw, C.I. Evolving models of pavlovian conditioning: Cerebellar cortical dynamics in awake behaving mice. Cell Rep. 2015, 13, 1977–1988. [Google Scholar] [CrossRef] [Green Version]
- Sarnaik, R.; Raman, I.M. Control of voluntary and optogenetically perturbed locomotion by spike rate and timing of neurons of the mouse cerebellar nuclei. eLife 2018, 7, e29546. [Google Scholar] [CrossRef] [PubMed]
- Gauck, V.; Jaeger, D. The control of rate and timing of spikes in the deep cerebellar nuclei by inhibition. J. Neurosci. 2000, 20, 3006–3016. [Google Scholar] [CrossRef] [PubMed]
- De Zeeuw, C.I.; Hoebeek, F.E.; Bosman, L.W.; Schonewille, M.; Witter, L.; Koekkoek, S.K. Spatiotemporal firing patterns in the cerebellum. Nat. Rev. Neurosci. 2011, 12, 327–344. [Google Scholar] [CrossRef] [PubMed]
- Najac, M.; Raman, I.M. Integration of purkinje cell inhibition by cerebellar nucleo-olivary neurons. J. Neurosci. 2015, 35, 544–549. [Google Scholar] [CrossRef] [Green Version]
- Najac, M.; Raman, I.M. Synaptic excitation by climbing fibre collaterals in the cerebellar nuclei of juvenile and adult mice. J. Physiol. 2017, 595, 6703–6718. [Google Scholar] [CrossRef]
- Lu, H.; Yang, B.; Jaeger, D. Cerebellar nuclei neurons show only small excitatory responses to optogenetic olivary stimulation in transgenic mice: In vivo and in vitro studies. Front. Neural Circuits 2016, 10, 21. [Google Scholar] [CrossRef] [Green Version]
- Chung, S.H.; Marzban, H.; Hawkes, R. Compartmentation of the cerebellar nuclei of the mouse. Neuroscience 2009, 161, 123–138. [Google Scholar] [CrossRef]
- Sugihara, I. Compartmentalization of the deep cerebellar nuclei based on afferent projections and aldolase c expression. Cerebellum 2011, 10, 449–463. [Google Scholar] [CrossRef]
- Sugihara, I.; Shinoda, Y. Molecular, topographic, and functional organization of the cerebellar nuclei: Analysis by three-dimensional mapping of the olivonuclear projection and aldolase c labeling. J. Neurosci. 2007, 27, 9696–9710. [Google Scholar] [CrossRef]
- Zhou, H.; Lin, Z.; Voges, K.; Ju, C.; Gao, Z.; Bosman, L.W.; Ruigrok, T.J.; Hoebeek, F.E.; De Zeeuw, C.I.; Schonewille, M. Cerebellar modules operate at different frequencies. eLife 2014, 3, e02536. [Google Scholar] [CrossRef] [PubMed]
- Beekhof, G.C.; Osório, C.; White, J.J.; van Zoomeren, S.; van der Stok, H.; Xiong, B.; Nettersheim, I.H.M.S.; Mak, W.A.; Runge, M.; Fiocchi, F.R.; et al. Differential spatiotemporal development of purkinje cell populations and cerebellum-dependent sensorimotor behaviors. eLife 2021, 10, e63668. [Google Scholar] [CrossRef] [PubMed]
- Ten Brinke, M.M.; Heiney, S.A.; Wang, X.; Proietti-Onori, M.; Boele, H.J.; Bakermans, J.; Medina, J.F.; Gao, Z.; De Zeeuw, C.I. Dynamic modulation of activity in cerebellar nuclei neurons during pavlovian eyeblink conditioning in mice. eLife 2017, 6, e28132. [Google Scholar] [CrossRef]
- Fujita, H.; Kodama, T.; du Lac, S. Modular output circuits of the fastigial nucleus for diverse motor and nonmotor functions of the cerebellar vermis. eLife 2020, 9, e58613. [Google Scholar] [CrossRef] [PubMed]
- Dehnes, Y.; Chaudhry, F.A.; Ullensvang, K.; Lehre, K.P.; Storm-Mathisen, J.; Danbolt, N.C. The glutamate transporter eaat4 in rat cerebellar purkinje cells: A glutamate-gated chloride channel concentrated near the synapse in parts of the dendritic membrane facing astroglia. J. Neurosci. 1998, 18, 3606–3619. [Google Scholar] [CrossRef] [Green Version]
- Canto, C.B.; Onuki, Y.; Bruinsma, B.; van der Werf, Y.D.; De Zeeuw, C.I. The sleeping cerebellum. Trends Neurosci. 2017, 40, 309–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates, 2nd ed.; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Ankri, L.; Yarom, Y.; Uusisaari, M.Y. Slice it hot: Acute adult brain slicing in physiological temperature. J. Vis. Exp. JoVE 2014, 92, e52068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekerli, M.; Del Negro, C.A.; Lee, R.H.; Butera, R.J. Estimating action potential thresholds from neuronal time-series: New metrics and evaluation of methodologies. IEEE Trans. Biomed. Eng. 2004, 51, 1665–1672. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. Nih image to imagej: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Ahn, A.H.; Dziennis, S.; Hawkes, R.; Herrup, K. The cloning of zebrin ii reveals its identity with aldolase c. Development 1994, 120, 2081–2090. [Google Scholar] [CrossRef] [PubMed]
- Aminov, Y.; Donchin, O.; Frens, M.A.; Winkelman, B.H.J. P2s2—“Ping-pong” Spike Sorting: An Interactive Method for Sorting Spikes Based on Their Shape and Inter-Spike Intervals. In Federation of European Neuroscience Societues; University of Barcelona: Barcelona, Spain, 2012. [Google Scholar]
- Shin, S.L.; Hoebeek, F.E.; Schonewille, M.; De Zeeuw, C.I.; Aertsen, A.; De Schutter, E. Regular patterns in cerebellar purkinje cell simple spike trains. PLoS ONE 2007, 2, e485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gincel, D.; Regan, M.R.; Jin, L.; Watkins, A.M.; Bergles, D.E.; Rothstein, J.D. Analysis of cerebellar purkinje cells using eaat4 glutamate transporter promoter reporter in mice generated via bacterial artificial chromosome-mediated transgenesis. Exp. Neurol. 2007, 203, 205–212. [Google Scholar] [CrossRef]
- Gong, S.; Zheng, C.; Doughty, M.L.; Losos, K.; Didkovsky, N.; Schambra, U.B.; Nowak, N.J.; Joyner, A.; Leblanc, G.; Hatten, M.E.; et al. A gene expression atlas of the central nervous system based on bacterial artificial chromosomes. Nature 2003, 425, 917–925. [Google Scholar] [CrossRef]
- Brochu, G.; Maler, L.; Hawkes, R. Zebrin ii: A polypeptide antigen expressed selectively by purkinje cells reveals compartments in rat and fish cerebellum. J. Comp. Neurol. 1990, 291, 538–552. [Google Scholar] [CrossRef]
- Lannoo, M.J.; Brochu, G.; Maler, L.; Hawkes, R. Zebrin II immunoreactivity in the rat and in the weakly electric teleost eigenmannia (gymnotiformes) reveals three modes of purkinje cell development. J. Comp. Neurol. 1991, 310, 215–233. [Google Scholar] [CrossRef]
- Uusisaari, M.; Obata, K.; Knopfel, T. Morphological and electrophysiological properties of gabaergic and non-gabaergic cells in the deep cerebellar nuclei. J. Neurophysiol. 2007, 97, 901–911. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Cerminara, N.L.; Kotsurovskyy, Y.; Aoki, H.; Burroughs, A.; Wise, A.K.; Luo, Y.; Marshall, S.P.; Sugihara, I.; Apps, R.; et al. Systematic regional variations in purkinje cell spiking patterns. PLoS ONE 2014, 9, e105633. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Voges, K.; Lin, Z.; Ju, C.; Schonewille, M. Differential purkinje cell simple spike activity and pausing behavior related to cerebellar modules. J. Neurophysiol. 2015, 113, 2524–2536. [Google Scholar] [CrossRef] [Green Version]
- Uusisaari, M.; De Schutter, E. The mysterious microcircuitry of the cerebellar nuclei. J. Physiol. 2011, 589, 3441–3457. [Google Scholar] [CrossRef] [PubMed]
- Pugh, J.R.; Raman, I.M. Potentiation of mossy fiber epscs in the cerebellar nuclei by nmda receptor activation followed by postinhibitory rebound current. Neuron 2006, 51, 113–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Gruijl, J.R.; Hoogland, T.M.; De Zeeuw, C.I. Behavioral correlates of complex spike synchrony in cerebellar microzones. J. Neurosci. 2014, 34, 8937–8947. [Google Scholar] [CrossRef] [Green Version]
- Beitzel, C.S.; Houck, B.D.; Lewis, S.M.; Person, A.L. Rubrocerebellar feedback loop isolates the interposed nucleus as an independent processor of corollary discharge information in mice. J. Neurosci. 2017, 37, 10085–10096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Raman, I.M. Facilitation of mossy fibre-driven spiking in the cerebellar nuclei by the synchrony of inhibition. J. Physiol. 2017, 595, 5245–5264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarden-Rabinowitz, Y.; Yarom, Y. In vivo analysis of synaptic activity in cerebellar nuclei neurons unravels the efficacy of excitatory inputs. J. Physiol. 2017, 595, 5945–5963. [Google Scholar] [CrossRef] [Green Version]
- Schonewille, M.; Khosrovani, S.; Winkelman, B.H.; Hoebeek, F.E.; De Jeu, M.T.; Larsen, I.M.; Van der Burg, J.; Schmolesky, M.T.; Frens, M.A.; De Zeeuw, C.I. Purkinje cells in awake behaving animals operate at the upstate membrane potential. Nat. Neurosci. 2006, 9, 459–461. [Google Scholar] [CrossRef]
- Adamson, T.W.; Kendall, L.V.; Goss, S.; Grayson, K.; Touma, C.; Palme, R.; Chen, J.Q.; Borowsky, A.D. Assessment of carprofen and buprenorphine on recovery of mice after surgical removal of the mammary fat pad. J. Am. Assoc. Lab. Anim. Sci. 2010, 49, 610–616. [Google Scholar] [PubMed]
- Mast, T.G.; Griff, E.R. The effects of analgesic supplements on neural activity in the main olfactory bulb of the mouse. Comp. Med. 2007, 57, 167–174. [Google Scholar] [PubMed]
- Pert, C.B.; Pert, A.; Tallman, J.F. Isolation of a novel endogenous opiate analgesic from human blood. Proc. Natl. Acad. Sci. USA 1976, 73, 2226–2230. [Google Scholar] [CrossRef] [Green Version]
- Ikai, Y.; Takada, M.; Mizuno, N. Single neurons in the ventral tegmental area that project to both the cerebral and cerebellar cortical areas by way of axon collaterals. Neuroscience 1994, 61, 925–934. [Google Scholar] [CrossRef]
- Grant, S.J.; Sonti, G. Buprenorphine and morphine produce equivalent increases in extracellular single unit activity of dopamine neurons in the ventral tegmental area in vivo. Synapse 1994, 16, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Ikai, Y.; Takada, M.; Shinonaga, Y.; Mizuno, N. Dopaminergic and non-dopaminergic neurons in the ventral tegmental area of the rat project, respectively, to the cerebellar cortex and deep cerebellar nuclei. Neuroscience 1992, 51, 719–728. [Google Scholar] [CrossRef]
- Aizenman, C.D.; Huang, E.J.; Linden, D.J. Morphological correlates of intrinsic electrical excitability in neurons of the deep cerebellar nuclei. J. Neurophysiol. 2003, 89, 1738–1747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canto, C.B.; Witter, L.; De Zeeuw, C.I. Whole-cell properties of cerebellar nuclei neurons in vivo. PLoS ONE 2016, 11, e0165887. [Google Scholar] [CrossRef] [PubMed]
- Gardette, R.; Debono, M.; Dupont, J.L.; Crepel, F. Electrophysiological studies on the postnatal development of intracerebellar nuclei neurons in rat cerebellar slices maintained in vitro. I. Postsynaptic potentials. Brain Res. 1985, 351, 47–55. [Google Scholar] [CrossRef]
- Gardette, R.; Debono, M.; Dupont, J.L.; Crepel, F. Electrophysiological studies on the postnatal development of intracerebellar nuclei neurons in rat cerebellar slices maintained in vitro. II. Membrane conductances. Brain Res. 1985, 352, 97–106. [Google Scholar] [CrossRef]
- Alvina, K.; Walter, J.T.; Kohn, A.; Ellis-Davies, G.; Khodakhah, K. Questioning the role of rebound firing in the cerebellum. Nat. Neurosci. 2008, 11, 1256–1258. [Google Scholar] [CrossRef] [Green Version]
- Alvina, K.; Tara, E.; Khodakhah, K. Developmental change in the contribution of voltage-gated Ca2+ channels to the pacemaking of deep cerebellar nuclei neurons. Neuroscience 2016, 322, 171–177. [Google Scholar] [CrossRef] [Green Version]
- LeDoux, M.S.; Hurst, D.C.; Lorden, J.F. Single-unit activity of cerebellar nuclear cells in the awake genetically dystonic rat. Neuroscience 1998, 86, 533–545. [Google Scholar] [CrossRef]
- Arancillo, M.; White, J.J.; Lin, T.; Stay, T.L.; Sillitoe, R.V. In vivo analysis of purkinje cell firing properties during postnatal mouse development. J. Neurophysiol. 2015, 113, 578–591. [Google Scholar] [CrossRef] [Green Version]
- Ruigrok, T.J. Ins and outs of cerebellar modules. Cerebellum 2011, 10, 464–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.; Proietti-Onori, M.; Lin, Z.; Ten Brinke, M.M.; Boele, H.J.; Potters, J.W.; Ruigrok, T.J.; Hoebeek, F.E.; De Zeeuw, C.I. Excitatory cerebellar nucleocortical circuit provides internal amplification during associative conditioning. Neuron 2016, 89, 645–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houck, B.D.; Person, A.L. Cerebellar premotor output neurons collateralize to innervate the cerebellar cortex. J. Comp. Neurol. 2015, 523, 2254–2271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Zeeuw, C.I. Bidirectional learning in upbound and downbound microzones of the cerebellum. Nat. Rev. Neurosci. 2021, 22, 92–110. [Google Scholar] [CrossRef]
- De Zeeuw, C.I.; Lisberger, S.G.; Raymond, J.L. Diversity and dynamism in the cerebellum. Nat. Neurosci. 2021, 24, 160–167. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beekhof, G.C.; Gornati, S.V.; Canto, C.B.; Libster, A.M.; Schonewille, M.; De Zeeuw, C.I.; Hoebeek, F.E. Activity of Cerebellar Nuclei Neurons Correlates with ZebrinII Identity of Their Purkinje Cell Afferents. Cells 2021, 10, 2686. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10102686
Beekhof GC, Gornati SV, Canto CB, Libster AM, Schonewille M, De Zeeuw CI, Hoebeek FE. Activity of Cerebellar Nuclei Neurons Correlates with ZebrinII Identity of Their Purkinje Cell Afferents. Cells. 2021; 10(10):2686. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10102686
Chicago/Turabian StyleBeekhof, Gerrit C., Simona V. Gornati, Cathrin B. Canto, Avraham M. Libster, Martijn Schonewille, Chris I. De Zeeuw, and Freek E. Hoebeek. 2021. "Activity of Cerebellar Nuclei Neurons Correlates with ZebrinII Identity of Their Purkinje Cell Afferents" Cells 10, no. 10: 2686. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10102686