
The Unique Properties of Placental Mesenchymal Stromal Cells: A Novel Source of Therapy for Congenital and Acquired Spinal Cord Injury

 ,
, 
Abstract
:1. Introduction
2. Spinal Cord Injury Pathogenesis
3. Properties of Placental Mesenchymal Stem/Stromal Cells
3.1. Isolation and Culture
3.2. PMSC Characterization
3.3. PMSC-Secretome—Free Proteins
3.4. PMSC Secretome—Extracellular Vesicles (EVs)
4. Potential PMSC Therapeutic Mechanisms
4.1. Neuroprotection
4.2. Immunomodulation
4.3. Myelin Regeneration
4.4. Angiogenesis
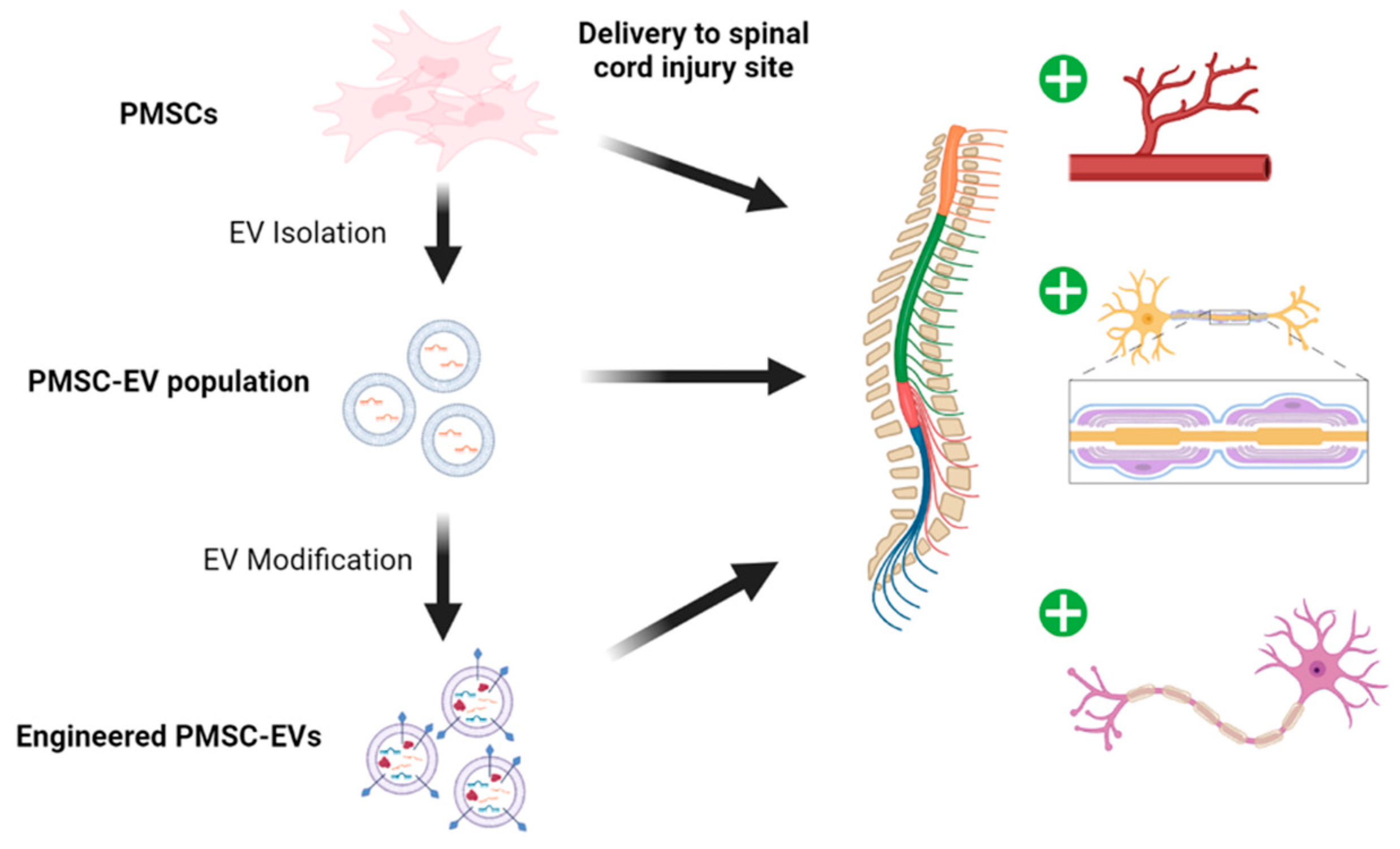
5. PMSC and PMSC-EV Bioengineering
5.1. Delivery
5.2. Surface Modification
5.3. Cargo Loading
6. Applications of PMSCs in SCI
6.1. Congenital SCI
6.2. Acquired SCI
7. MSCs and PMSCs in Clinical Trials
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cofano, F.; Boido, M.; Monticelli, M.; Zenga, F.; Ducati, A.; Vercelli, A.; Garbossa, D. Molecular Sciences Mesenchymal Stem Cells for Spinal Cord Injury: Current Options, Limitations, and Future of Cell Therapy. Int. J. Mol. Sci. 2019, 20, 2698. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.C.; Medress, Z.A.; Azad, T.D.; Doulames, V.M.; Veeravagu, A. Stem Cell Therapies for Acute Spinal Cord Injury in Humans: A Review. Neurosurg. Focus 2018. [Google Scholar] [CrossRef] [PubMed]
- Marketing, U.D.H. Public Affairs and First Stem Cell Clinical Trial for Spina Bifida Treatment Announced. Available online: https://health.ucdavis.edu/health-news/newsroom/first-stem-cell-clinical-trial-for-spina-bifida-treatment-announced/2021/03 (accessed on 14 August 2021).
- Mitchell, L.E.; Adzick, N.S.; Melchionne, J.; Pasquariello, P.S.; Sutton, L.N.; Whitehead, A.S. Spina Bifida. Lancet 2004, 364, 1885–1895. [Google Scholar] [CrossRef]
- Dewan, M.C.; Wellons, J.C. Fetal Surgery for Spina Bifida: JNSPG 75th Anniversary Invited Review Article. J. Neurosurg. Pediatr. 2019, 24, 105–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adzick, N.S.; Thom, E.A.; Spong, C.Y.; Brock, J.W.; Burrows, P.K.; Johnson, M.P.; Howell, L.J.; Farrell, J.A.; Dabrowiak, M.E.; Sutton, L.N.; et al. A Randomized Trial of Prenatal versus Postnatal Repair of Myelomeningocele. N. Engl. J. Med. 2011, 364, 993–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parolini, O.; Alviano, F.; Bagnara, G.P.; Bilic, G.; Bühring, H.-J.; Evangelista, M.; Hennerbichler, S.; Liu, B.; Magatti, M.; Mao, N.; et al. Concise Review: Isolation and Characterization of Cells from Human Term Placenta: Outcome of the First International Workshop on Placenta Derived Stem Cells. Stem Cells 2008, 26, 300–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowland, J.W.; Hawryluk, G.W.J.; Kwon, B.; Fehlings, M.G. Current Status of Acute Spinal Cord Injury Pathophysiology and Emerging Therapies: Promise on the Horizon. Neurosurg, Focus 2008, 25, E2. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.; Dyck, S.M.; Karimi-Abdolrezaee, S. Traumatic Spinal Cord Injury: An Overview of Pathophysiology, Models and Acute Injury Mechanisms. Front. Neurol. 2019, 10, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatesh, K.; Ghosh, S.K.; Mullick, M.; Manivasagam, G.; Sen, D. Spinal Cord Injury: Pathophysiology, Treatment Strategies, Associated Challenges, and Future Implications. Cell Tissue Res. 2019, 377, 125–151. [Google Scholar] [CrossRef] [PubMed]
- Essentials of Spinal Cord Injury. Available online: https://www.thieme.com/books-main/neurosurgery/product/1390-essentials-of-spinal-cord-injury (accessed on 5 September 2021).
- Wilson, J.R.; Hashimoto, R.E.; Dettori, J.R.; Fehlings, M.G. Spinal Cord Injury and Quality of Life: A Systematic Review of Outcome Measures. Evid. Based Spine Care J. 2011, 2, 37–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figley, S.A.; Khosravi, R.; Legasto, J.M.; Tseng, Y.-F.; Fehlings, M.G. Characterization of Vascular Disruption and Blood-Spinal Cord Barrier Permeability Following Traumatic Spinal Cord Injury. J. Neurotrauma 2014, 31, 541–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pineau, I.; Lacroix, S. Proinflammatory Cytokine Synthesis in the Injured Mouse Spinal Cord: Multiphasic Expression Pattern and Identification of the Cell Types Involved. J. Comp. Neurol. 2007, 500, 267–285. [Google Scholar] [CrossRef] [PubMed]
- David, S.; López-Vales, R.; Wee Yong, V. Harmful and Beneficial Effects of Inflammation after Spinal Cord Injury: Potential Therapeutic Implications. Handb. Clin. Neurol. 2012, 109, 485–502. [Google Scholar] [CrossRef] [PubMed]
- Huntemer-Silveira, A.; Patil, N.; Brickner, M.A.; Parr, A.M. Strategies for Oligodendrocyte and Myelin Repair in Traumatic CNS Injury. Front. Cell. Neurosci. 2021, 14, 458. [Google Scholar] [CrossRef] [PubMed]
- Kakulas, B.A. Neuropathology: The Foundation for New Treatments in Spinal Cord Injury. Spinal Cord 2004, 42, 549–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tator, C.H.; Koyanagi, I. Vascular Mechanisms in the Pathophysiology of Human Spinal Cord Injury. J. Neurosurg. 1997, 86, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, T.; Hida, K.; Akino, M.; Yano, S.; Iwasaki, Y.; Saito, H. Ultra-Early MRI Showing No Abnormality in a Fall Victim Presenting with Tetraparesis. Spinal Cord 2007, 45, 695–699. [Google Scholar] [CrossRef] [PubMed]
- Norenberg, M.D.; Smith, J.; Marcillo, A. The Pathology of Human Spinal Cord Injury: Defining the Problems. J. Neurotrauma 2004, 21, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Bunge, R.P.; Puckett, W.R.; Becerra, J.L.; Marcillo, A.; Quencer, R.M. Observations on the Pathology of Human Spinal Cord Injury. A Review and Classification of 22 New Cases with Details from a Case of Chronic Cord Compression with Extensive Focal Demyelination. Adv. Neurol. 1993, 59, 75–89. [Google Scholar] [PubMed]
- McDonald, J.W.; Belegu, V. Demyelination and Remyelination after Spinal Cord Injury. J. Neurotrauma 2006, 23, 345–359. [Google Scholar] [CrossRef] [PubMed]
- Nashmi, R.; Fehlings, M.G. Changes in Axonal Physiology and Morphology after Chronic Compressive Injury of the Rat Thoracic Spinal Cord. Neuroscience 2001, 104, 235–251. [Google Scholar] [CrossRef]
- Radojicic, M.; Reier, P.J.; Steward, O.; Keirstead, H.S. Septations in Chronic Spinal Cord Injury Cavities Contain Axons. Exp. Neurol. 2005, 196, 339–341. [Google Scholar] [CrossRef] [PubMed]
- Fehlings, M.G.; Tator, C.H. The Relationships among the Severity of Spinal Cord Injury, Residual Neurological Function, Axon Counts, and Counts of Retrogradely Labeled Neurons after Experimental Spinal Cord Injury. Exp. Neurol. 1995, 132, 220–228. [Google Scholar] [CrossRef]
- Lu, P.; Jones, L.L.; Snyder, E.Y.; Tuszynski, M.H. Neural Stem Cells Constitutively Secrete Neurotrophic Factors and Promote Extensive Host Axonal Growth after Spinal Cord Injury. Exp. Neurol. 2003, 181, 115–129. [Google Scholar] [CrossRef]
- Wilson, J.R.; Tetreault, L.A.; Kwon, B.K.; Arnold, P.M.; Mroz, T.E.; Shaffrey, C.; Harrop, J.S.; Chapman, J.R.; Casha, S.; Skelly, A.C.; et al. Timing of Decompression in Patients with Acute Spinal Cord Injury: A Systematic Review. Glob. Spine J. 2017, 7, 95S–115S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehlings, M.G.; Vaccaro, A.; Wilson, J.R.; Singh, A.; W Cadotte, D.; Harrop, J.S.; Aarabi, B.; Shaffrey, C.; Dvorak, M.; Fisher, C.; et al. Early versus Delayed Decompression for Traumatic Cervical Spinal Cord Injury: Results of the Surgical Timing in Acute Spinal Cord Injury Study (STASCIS). PLoS ONE 2012, 7, e32037. [Google Scholar] [CrossRef] [PubMed]
- Schanne, F.A.; Kane, A.B.; Young, E.E.; Farber, J.L. Calcium Dependence of Toxic Cell Death: A Final Common Pathway. Science 1979, 206, 700–702. [Google Scholar] [CrossRef] [PubMed]
- Lipton, S.A.; Rosenberg, P.A. Excitatory Amino Acids as a Final Common Pathway for Neurologic Disorders. N. Engl. J. Med. 1994, 330, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, D.J.; Popovich, P.G. Inflammation and Its Role in Neuroprotection, Axonal Regeneration and Functional Recovery after Spinal Cord Injury. Exp. Neurol. 2008, 209, 378–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Rabchevsky, A.G.; Hall, E.D. Role of Peroxynitrite in Secondary Oxidative Damage after Spinal Cord Injury. J. Neurochem. 2007, 100, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Noble, L.J.; Wrathall, J.R. Distribution and Time Course of Protein Extravasation in the Rat Spinal Cord after Contusive Injury. Brain Res. 1989, 482, 57–66. [Google Scholar] [CrossRef]
- Fleming, J.C.; Norenberg, M.D.; Ramsay, D.A.; Dekaban, G.A.; Marcillo, A.E.; Saenz, A.D.; Pasquale-Styles, M.; Dietrich, W.D.; Weaver, L.C. The Cellular Inflammatory Response in Human Spinal Cords after Injury. Brain 2006, 129, 3249–3269. [Google Scholar] [CrossRef] [PubMed]
- Popovich, P.G.; Wei, P.; Stokes, B.T. Cellular Inflammatory Response after Spinal Cord Injury in Sprague-Dawley and Lewis Rats. J. Comp. Neurol. 1997, 377, 443–464. [Google Scholar] [CrossRef]
- Beattie, M.S.; Hermann, G.E.; Rogers, R.C.; Bresnahan, J.C. Cell Death in Models of Spinal Cord Injury. Prog. Brain Res. 2002, 137, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Emery, E.; Aldana, P.; Bunge, M.B.; Puckett, W.; Srinivasan, A.; Keane, R.W.; Bethea, J.; Levi, A.D. Apoptosis after Traumatic Human Spinal Cord Injury. J. Neurosurg. 1998, 89, 911–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keane, R.W.; Kraydieh, S.; Lotocki, G.; Bethea, J.R.; Krajewski, S.; Reed, J.C.; Dietrich, W.D. Apoptotic and Anti-Apoptotic Mechanisms Following Spinal Cord Injury. J. Neuropathol. Exp. Neurol. 2001, 60, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, J.E.; Imura, T.; Song, B.; Qi, J.; Ao, Y.; Nguyen, T.K.; Korsak, R.A.; Takeda, K.; Akira, S.; Sofroniew, M.V. STAT3 Is a Critical Regulator of Astrogliosis and Scar Formation after Spinal Cord Injury. J. Neurosci. 2008, 28, 7231–7243. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.E.; Beattie, M.S.; Bresnahan, J.C. Degeneration and Sprouting of Identified Descending Supraspinal Axons after Contusive Spinal Cord Injury in the Rat. Exp. Neurol. 2001, 171, 153–169. [Google Scholar] [CrossRef] [PubMed]
- Stoodley, M.A. Pathophysiology of Syringomyelia. J. Neurosurg. 2000, 92, 1069–1073. [Google Scholar]
- McDonald, J.W.; Becker, D.; Sadowsky, C.L.; Jane, J.A.; Conturo, T.E.; Schultz, L.M. Late Recovery Following Spinal Cord Injury. Case Report and Review of the Literature. J. Neurosurg. 2002, 97, 252–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimi-Abdolrezaee, S.; Eftekharpour, E.; Wang, J.; Morshead, C.M.; Fehlings, M.G. Delayed Transplantation of Adult Neural Precursor Cells Promotes Remyelination and Functional Neurological Recovery after Spinal Cord Injury. J. Neurosci. 2006, 26, 3377–3389. [Google Scholar] [CrossRef] [PubMed]
- Keirstead, H.S.; Nistor, G.; Bernal, G.; Totoiu, M.; Cloutier, F.; Sharp, K.; Steward, O. Human Embryonic Stem Cell-Derived Oligodendrocyte Progenitor Cell Transplants Remyelinate and Restore Locomotion after Spinal Cord Injury. J. Neurosci. 2005, 25, 4694–4705. [Google Scholar] [CrossRef] [PubMed]
- Hagg, T.; Oudega, M. Degenerative and Spontaneous Regenerative Processes after Spinal Cord Injury. J. Neurotrauma 2006, 23, 264–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meuli, M.; Meuli-Simmen, C.; Hutchins, G.M.; Seller, M.J.; Harrison, M.R.; Adzick, N.S. The Spinal Cord Lesion in Human Fetuses with Myelomeningocele: Implications for Fetal Surgery. J. Pediatr. Surg. 1997, 32, 448–452. [Google Scholar] [CrossRef]
- Hutchins, G.M.; Meuli, M.; Meuli-Simmen, C.; Jordan, M.A.; Heffez, D.S.; Blakemore, K.J. Acquired Spinal Cord Injury in Human Fetuses with Myelomeningocele. Pediatr. Pathol. Lab. Med. 1996, 16, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Stiefel, D.; Copp, A.J.; Meuli, M. Fetal Spina Bifida: Loss of Neural Function in utero. J. Neurosurg. 2007, 106, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heffez, D.S.; Aryanpur, J.; Hutchins, G.M.; Freeman, J.M. The Paralysis Associated with Myelomeningocele: Clinical and Experimental Data Implicating a Preventable Spinal Cord Injury. Neurosurgery 1990, 26, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Copp, A.J.; Stanier, P.; Greene, N.D.E. Neural Tube Defects–Recent Advances, Unsolved Questions and Controversies. Lancet Neurol. 2013, 12, 799–810. [Google Scholar] [CrossRef] [Green Version]
- Adzick, N.S. Fetal Surgery for Myelomeningocele: Trials and Tribulations. Isabella Forshall Lecture. J. Pediatr. Surg. 2012, 47, 273–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oria, M.; Figueira, R.L.; Scorletti, F.; Sbragia, L.; Owens, K.; Li, Z.; Pathak, B.; Corona, M.U.; Marotta, M.; Encinas, J.L.; et al. CD200-CD200R Imbalance Correlates with Microglia and pro-Inflammatory Activation in Rat Spinal Cords Exposed to Amniotic Fluid in Retinoic Acid-Induced Spina Bifida. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Danzer, E.; Zhang, L.; Radu, A.; Bebbington, M.W.; Liechty, K.W.; Adzick, N.S.; Flake, A.W. Amniotic Fluid Levels of Glial Fibrillary Acidic Protein in Fetal Rats with Retinoic Acid Induced Myelomeningocele: A Potential Marker for Spinal Cord Injury. Am. J. Obs. Gynecol. 2011, 204, 178.e1-11. [Google Scholar] [CrossRef] [PubMed]
- Tomko, P.; Farkaš, D.; Čížková, D.; Vanický, I. Longitudinal Enlargement of the Lesion after Spinal Cord Injury in the Rat: A Consequence of Malignant Edema? Spinal Cord 2017, 55, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Lankford, L.; Chen, Y.J.; Saenz, Z.; Kumar, P.; Long, C.; Farmer, D.; Wang, A. Manufacture and Preparation of Human Placenta-Derived Mesenchymal Stromal Cells for Local Tissue Delivery. Cytotherapy 2017, 19, 680–688. [Google Scholar] [CrossRef] [PubMed]
- Lankford, L. Early Gestation Chorionic Villi-Derived Stromal Cells for Fetal Tissue Engineering. World J. Stem Cells 2015, 7, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.S.; Choi, Y.; Kim, H.-S.; Kim, H.O. Comparison of Molecular Profiles of Human Mesenchymal Stem Cells Derived from Bone Marrow, Umbilical Cord Blood, Placenta and Adipose Tissue. Int. J. Mol. Med. 2016, 37, 115–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barlow, S.; Brooke, G.; Chatterjee, K.; Price, G.; Pelekanos, R.; Rossetti, T.; Doody, M.; Venter, D.; Pain, S.; Gilshenan, K.; et al. Comparison of Human Placenta- and Bone Marrow–Derived Multipotent Mesenchymal Stem Cells. Stem Cells Dev. 2008, 17, 1095–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portmann-Lanz, C.B.; Schoeberlein, A.; Portmann, R.; Mohr, S.; Rollini, P.; Sager, R.; Surbek, D.V. Turning Placenta into Brain: Placental Mesenchymal Stem Cells Differentiate into Neurons and Oligodendrocytes. Am. J. Obs. Gynecol. 2010, 202. [Google Scholar] [CrossRef] [PubMed]
- Martini, M.M.; da S. Jeremias, T.; Kohler, M.C.; Marostica, L.L.; Trentin, A.G.; Alvarez-Silva, M. Human Placenta-Derived Mesenchymal Stem Cells Acquire Neural Phenotype Under the Appropriate Niche Conditions. DNA Cell Biol. 2013, 32, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; He, D.-M.; Zhang, Y. The Differentiation of Human Placenta-Derived Mesenchymal Stem Cells into Dopaminergic Cells in Vitro. Cell Mol. Biol. Lett. 2009, 14, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Brown, E.G.; Lankford, L.; Keller, B.A.; Pivetti, C.D.; Sitkin, N.A.; Beattie, M.S.; Bresnahan, J.C.; Farmer, D.L. Placental Mesenchymal Stromal Cells Rescue Ambulation in Ovine Myelomeningocele. Stem Cells Transl. Med. 2015, 4, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, H.; Erkers, T.; Nava, S.; Ruhm, S.; Westgren, M.; Ringdén, O. Stromal Cells from Term Fetal Membrane Are Highly Suppressive in Allogeneic Settings in Vitro. Clin. Exp. Immunol. 2012, 167, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-R.; Kyung, J.W.; Kumar, H.; Kwon, S.P.; Song, S.Y.; Han, I.-B.; Kim, B.-S. Targeted Delivery of Mesenchymal Stem Cell-Derived Nanovesicles for Spinal Cord Injury Treatment. Int. J. Mol. Sci. 2020, 21, 4185. [Google Scholar] [CrossRef] [PubMed]
- Park, W.S.; Ahn, S.Y.; Sung, S.I.; Ahn, J.-Y.; Chang, Y.S. Strategies to Enhance Paracrine Potency of Transplanted Mesenchymal Stem Cells in Intractable Neonatal Disorders. Pediatr. Res. 2018, 83, 214–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cízková, D.; Rosocha, J.; Vanický, I.; Jergová, S.; Cízek, M. Transplants of Human Mesenchymal Stem Cells Improve Functional Recovery after Spinal Cord Injury in the Rat. Cell Mol. Neurobiol. 2006, 26, 1167–1180. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Park, J.-S. Usage of Human Mesenchymal Stem Cells in Cell-Based Therapy: Advantages and Disadvantages. Dev. Reprod 2017, 21, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Ding, Y.; Zhang, Y.; Tse, H.-F.; Lian, Q. Paracrine Mechanisms of Mesenchymal Stem Cell-Based Therapy: Current Status and Perspectives. Cell Transplant. 2014, 23, 1045–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Xuan, A.; Chen, Y.; Zhang, J.; Xu, L.; Yan, Q.; Long, D. Combined Effect of Nerve Growth Factor and Brain-derived Neurotrophic Factor on Neuronal Differentiation of Neural Stem Cells and the Potential Molecular Mechanisms. Mol. Med. Rep. 2014, 10, 1739–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Chen, J.; Chen, X.G.; Wang, L.; Gautam, S.C.; Xu, Y.X.; Katakowski, M.; Zhang, L.J.; Lu, M.; Janakiraman, N.; et al. Human Marrow Stromal Cell Therapy for Stroke in Rat: Neurotrophins and Functional Recovery. Neurology 2002, 59, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Phan, J.; Kumar, P.; Hao, D.; Gao, K.; Farmer, D.; Wang, A. Engineering Mesenchymal Stem Cells to Improve Their Exosome Efficacy and Yield for Cell-Free Therapy. J. Extracell. Vesicles 2018, 7, 1522236. [Google Scholar] [CrossRef] [PubMed]
- Luarte, A.; Bátiz, L.F.; Wyneken, U.; Lafourcade, C. Potential Therapies by Stem Cell-Derived Exosomes in CNS Diseases: Focusing on the Neurogenic Niche. Stem Cells Int. 2016, 2016, 5736059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorabi, A.M.; Kiaie, N.; Barreto, G.E.; Read, M.I.; Tafti, H.A.; Sahebkar, A. The Therapeutic Potential of Mesenchymal Stem Cell-Derived Exosomes in Treatment of Neurodegenerative Diseases. Mol. Neurobiol. 2019, 56, 8157–8167. [Google Scholar] [CrossRef] [PubMed]
- Mendt, M.; Rezvani, K.; Shpall, E. Mesenchymal Stem Cell-Derived Exosomes for Clinical Use. Bone Marrow Transplant. 2019, 54, 789–792. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.S.Y. Mesenchymal Stem Cells: Angels or Demons? J. Biomed. Biotechnol. 2011, 2011, 459510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, P.; Han, T.; Xiang, X.; Wang, Y.; Fang, H.; Niu, Y.; Shen, C. The Role of Hepatocyte Growth Factor in Mesenchymal Stem Cell-Induced Recovery in Spinal Cord Injured Rats. Stem Cell Res. Ther. 2020, 11, 178. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Radtke, C.; Tan, A.M.; Zhao, P.; Hamada, H.; Houkin, K.; Honmou, O.; Kocsis, J.D. BDNF-Hypersecreting Human Mesenchymal Stem Cells Promote Functional Recovery, Axonal Sprouting, and Protection of Corticospinal Neurons after Spinal Cord Injury. J. Neurosci. 2009, 29, 14932–14941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, P.; Becker, J.C.; Gao, K.; Carney, R.P.; Lankford, L.; Keller, B.A.; Herout, K.; Lam, K.S.; Farmer, D.L.; Wang, A. Neuroprotective Effect of Placenta-Derived Mesenchymal Stromal Cells: Role of Exosomes. FASEB J. 2019, 33, 5836–5849. [Google Scholar] [CrossRef] [PubMed]
- Casha, S.; Zygun, D.; McGowan, M.D.; Bains, I.; Yong, V.W.; John Hurlbert, R. Results of a Phase II Placebo-Controlled Randomized Trial of Minocycline in Acute Spinal Cord Injury. Brain 2012, 135, 1224–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehlings, M.G.; Theodore, N.; Harrop, J.; Maurais, G.; Kuntz, C.; Shaffrey, C.I.; Kwon, B.K.; Chapman, J.; Yee, A.; Tighe, A.; et al. A Phase I/IIa Clinical Trial of a Recombinant Rho Protein Antagonist in Acute Spinal Cord Injury. J. Neurotrauma 2011, 28, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Papa, S.; Caron, I.; Erba, E.; Panini, N.; De Paola, M.; Mariani, A.; Colombo, C.; Ferrari, R.; Pozzer, D.; Zanier, E.R.; et al. Early Modulation of Pro-Inflammatory Microglia by Minocycline Loaded Nanoparticles Confers Long Lasting Protection after Spinal Cord Injury. Biomaterials 2016, 75, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Biglari, B.; Swing, T.; Child, C.; Büchler, A.; Westhauser, F.; Bruckner, T.; Ferbert, T.; Jürgen Gerner, H.; Moghaddam, A. A Pilot Study on Temporal Changes in IL-1β and TNF-α Serum Levels after Spinal Cord Injury: The Serum Level of TNF-α in Acute SCI Patients as a Possible Marker for Neurological Remission. Spinal Cord 2015, 53, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Kwon, B.K.; Streijger, F.; Fallah, N.; Noonan, V.K.; Bélanger, L.M.; Ritchie, L.; Paquette, S.J.; Ailon, T.; Boyd, M.C.; Street, J.; et al. Cerebrospinal Fluid Biomarkers To Stratify Injury Severity and Predict Outcome in Human Traumatic Spinal Cord Injury. J. Neurotrauma 2017, 34, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Ma, Z.; Li, J.; Kang, X. Mesenchymal Stem Cell-Derived Exosomes: Therapeutic Opportunities and Challenges for Spinal Cord Injury. Stem Cell Res. Ther. 2021, 12, 102. [Google Scholar] [CrossRef] [PubMed]
- Colton, C.A. Heterogeneity of Microglial Activation in the Innate Immune Response in the Brain. J. Neuroimmune Pharm. 2009, 4, 399–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popovich, P.G.; Guan, Z.; Wei, P.; Huitinga, I.; van Rooijen, N.; Stokes, B.T. Depletion of Hematogenous Macrophages Promotes Partial Hindlimb Recovery and Neuroanatomical Repair after Experimental Spinal Cord Injury. Exp. Neurol. 1999, 158, 351–365. [Google Scholar] [CrossRef] [PubMed]
- Romanelli, P.; Bieler, L.; Scharler, C.; Pachler, K.; Kreutzer, C.; Zaunmair, P.; Jakubecova, D.; Mrowetz, H.; Benedetti, B.; Rivera, F.J.; et al. Extracellular Vesicles Can Deliver Anti-Inflammatory and Anti-Scarring Activities of Mesenchymal Stromal Cells After Spinal Cord Injury. Front. Neurol. 2019, 10, 1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, S.; Kroner, A. Repertoire of Microglial and Macrophage Responses after Spinal Cord Injury. Nat. Rev. Neurosci. 2011, 12, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Kigerl, K.A.; Gensel, J.C.; Ankeny, D.P.; Alexander, J.K.; Donnelly, D.J.; Popovich, P.G. Identification of Two Distinct Macrophage Subsets with Divergent Effects Causing Either Neurotoxicity or Regeneration in the Injured Mouse Spinal Cord. J. Neurosci. 2009, 29, 13435–13444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Wang, Y.; Gong, F.; Rong, Y.; Luo, Y.; Tang, P.; Zhou, Z.; Zhou, Z.; Xu, T.; Jiang, T.; et al. Exosomes Derived from Bone Mesenchymal Stem Cells Repair Traumatic Spinal Cord Injury by Suppressing the Activation of A1 Neurotoxic Reactive Astrocytes. J. Neurotrauma 2019, 36, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Zhou, X.; Qiu, J.; Xin, D.; Li, T.; Chu, X.; Yuan, H.; Wang, H.; Wang, Z.; Wang, D. Exosomes Derived From Bone Marrow Mesenchymal Stem Cells Inhibit Complement Activation In Rats With Spinal Cord Injury. Drug Des. Dev. 2019, 13, 3693–3704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.M.; Jung, J.; Lee, H.-J.; Jeong, S.J.; Cho, K.J.; Hwang, S.-G.; Kim, G.J. Comparison of Immunomodulatory Effects of Placenta Mesenchymal Stem Cells with Bone Marrow and Adipose Mesenchymal Stem Cells. Int. Immunopharmacol. 2012, 13, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-J.; Yen, M.-L.; Chen, Y.-C.; Chien, C.-C.; Huang, H.-I.; Bai, C.-H.; Yen, B.L. Placenta-Derived Multipotent Cells Exhibit Immunosuppressive Properties That Are Enhanced in the Presence of Interferon-Gamma. Stem Cells 2006, 24, 2466–2477. [Google Scholar] [CrossRef] [PubMed]
- Abumaree, M.H.; Abomaray, F.M.; Alshabibi, M.A.; AlAskar, A.S.; Kalionis, B. Immunomodulatory Properties of Human Placental Mesenchymal Stem/Stromal Cells. Placenta 2017, 59, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.J.; Kim, H.-S.; Lee, H.-G.; Kim, G.J.; Jeon, H.G.; Shin, H.-S.; Chang, S.-K.; Hur, G.-H.; Chong, S.Y.; Oh, D.; et al. Placenta-Derived Mesenchymal Stem Cells Have an Immunomodulatory Effect That Can Control Acute Graft-versus-Host Disease in Mice. Acta Haematol. 2013, 129, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.M.; Ritter, T.; Ceredig, R.; Griffin, M.D. Mesenchymal Stem Cell Effects on T-Cell Effector Pathways. Stem Cell Res. 2011, 2, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amorim, R.M.; Clark, K.C.; Walker, N.J.; Kumar, P.; Herout, K.; Borjesson, D.L.; Wang, A. Placenta-Derived Multipotent Mesenchymal Stromal Cells: A Promising Potential Cell-Based Therapy for Canine Inflammatory Brain Disease. Stem Cell Res. Ther. 2020, 11, 304. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhang, W.; Jiang, X.; Mao, N. Human-Placenta-Derived Mesenchymal Stem Cells Inhibit Proliferation and Function of Allogeneic Immune Cells. Cell Tissue Res. 2007, 330, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Xue, Q.; Chen, Y.; Yu, G.-H.; Qing, M.; Shen, Y.; Wang, M.; Shi, Q.; Zhang, X.-G. Different Roles of PD-L1 and FasL in Immunomodulation Mediated by Human Placenta-Derived Mesenchymal Stem Cells. Hum. Immunol. 2013, 74, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Dombrowski, Y.; O’Hagan, T.; Dittmer, M.; Penalva, R.; Mayoral, S.R.; Bankhead, P.; Fleville, S.; Eleftheriadis, G.; Zhao, C.; Naughton, M.; et al. Regulatory T Cells Promote Myelin Regeneration in the Central Nervous System. Nat. Neurosci. 2017, 20, 674–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, R.J.M.; Ffrench-Constant, C. Remyelination in the CNS: From Biology to Therapy. Nat. Rev. Neurosci. 2008, 9, 839–855. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.; Zhang, S.; Barthe, S.; Kumar, P.; Pivetti, C.; Kreutzberg, N.; Reed, C.; Wang, Y.; Paxton, Z.; Farmer, D.; et al. Placental Mesenchymal Stem Cell-Derived Extracellular Vesicles Promote Myelin Regeneration in an Animal Model of Multiple Sclerosis. Cells 2019, 8, 1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, L.; Lennon, D.P.; Caplan, A.I.; DeChant, A.; Hecker, J.; Kranso, J.; Zaremba, A.; Miller, R.H. Hepatocyte Growth Factor Mediates Mesenchymal Stem Cell–Induced Recovery in Multiple Sclerosis Models. Nat. Neurosci. 2012, 15, 862–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giunti, D.; Marini, C.; Parodi, B.; Usai, C.; Milanese, M.; Bonanno, G.; Kerlero de Rosbo, N.; Uccelli, A. Role of MiRNAs Shuttled by Mesenchymal Stem Cell-Derived Small Extracellular Vesicles in Modulating Neuroinflammation. Sci. Rep. 2021, 11, 1740. [Google Scholar] [CrossRef] [PubMed]
- Osorio-Querejeta, I.; Alberro, A.; Muñoz-Culla, M.; Mäger, I.; Otaegui, D. Therapeutic Potential of Extracellular Vesicles for Demyelinating Diseases; Challenges and Opportunities. Front. Mol. Neurosci. 2018, 11, 434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, M.T.; Stammers, A.T.; Kwon, B.K. Vascular Disruption and the Role of Angiogenic Proteins after Spinal Cord Injury. Transl. Stroke Res. 2011, 2, 474–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauch, M.F.; Hynes, S.R.; Bertram, J.; Redmond, A.; Robinson, R.; Williams, C.; Xu, H.; Madri, J.A.; Lavik, E.B. Engineering Angiogenesis Following Spinal Cord Injury: A Coculture of Neural Progenitor and Endothelial Cells in a Degradable Polymer Implant Leads to an Increase in Vessel Density and Formation of the Blood-Spinal Cord Barrier. Eur. J. Neurosci. 2009, 29, 132–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofer, H.R.; Tuan, R.S. Secreted Trophic Factors of Mesenchymal Stem Cells Support Neurovascular and Musculoskeletal Therapies. Stem Cell Res. 2016, 7, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, D.; He, C.; Ma, B.; Lankford, L.; Reynaga, L.; Farmer, D.L.; Guo, F.; Wang, A. Hypoxic Preconditioning Enhances Survival and Proangiogenic Capacity of Human First Trimester Chorionic Villus-Derived Mesenchymal Stem Cells for Fetal Tissue Engineering. Stem Cells Int. 2019, 2019, 9695239. [Google Scholar] [CrossRef] [PubMed]
- Bian, X.; Ma, K.; Zhang, C.; Fu, X. Therapeutic Angiogenesis Using Stem Cell-Derived Extracellular Vesicles: An Emerging Approach for Treatment of Ischemic Diseases. Stem Cell Res. Ther. 2019, 10, 158. [Google Scholar] [CrossRef] [PubMed]
- Hao, D.; Swindell, H.S.; Ramasubramanian, L.; Liu, R.; Lam, K.S.; Farmer, D.L.; Wang, A. Extracellular Matrix Mimicking Nanofibrous Scaffolds Modified with Mesenchymal Stem Cell-Derived Extracellular Vesicles for Improved Vascularization. Front. Bioeng. Biotechnol. 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Rahimi-Sherbaf, F.; Nadri, S.; Rahmani, A.; Dabiri Oskoei, A. Placenta Mesenchymal Stem Cells Differentiation toward Neuronal-like Cells on Nanofibrous Scaffold. Bioimpacts 2020, 10, 117–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.; Robinson, M.; Petrie, T.; Spandler, V.; Boyd, W.D.; Sondergaard, C.S. Small Intestinal Submucosa-Derived Extracellular Matrix Bioscaffold Significantly Enhances Angiogenic Factor Secretion from Human Mesenchymal Stromal Cells. Stem Cell Res. 2015, 6, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaussaud, J.; Biais, M.; Calderon, J.; Chevaleyre, J.; Duchez, P.; Ivanovic, Z.; Couffinhal, T.; Barandon, L. Hypoxia-Preconditioned Mesenchymal Stromal Cells Improve Cardiac Function in a Swine Model of Chronic Myocardial Ischaemia. Eur. J. Cardiothorac. Surg. 2013, 43, 1050–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.S.; Noh, M.Y.; Cho, K.A.; Kim, H.; Kwon, M.-S.; Kim, K.S.; Kim, J.; Koh, S.-H.; Kim, S.H. Hypoxia/Reoxygenation-Preconditioned Human Bone Marrow-Derived Mesenchymal Stromal Cells Rescue Ischemic Rat Cortical Neurons by Enhancing Trophic Factor Release. Mol. Neurobiol. 2015, 52, 792–803. [Google Scholar] [CrossRef] [PubMed]
- Wiklander, O.P.B.; Nordin, J.Z.; O’Loughlin, A.; Gustafsson, Y.; Corso, G.; Mäger, I.; Vader, P.; Lee, Y.; Sork, H.; Seow, Y.; et al. Extracellular Vesicle in Vivo Biodistribution Is Determined by Cell Source, Route of Administration and Targeting. J. Extracell. Vesicles 2015, 4, 26316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Wang, Z.Y.; Zhang, J.; Zhang, Y.; Huo, H.; Wang, T.; Jiang, T.; Wang, S. RVG-Peptide-Linked Trimethylated Chitosan for Delivery of SiRNA to the Brain. Biomacromolecules 2014, 15, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, X.; Chen, X.; Wang, L.; Yang, G. Exosome Mediated Delivery of MiR-124 Promotes Neurogenesis after Ischemia. Mol. Ther. -Nucleic Acids 2017, 7, 278–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J.A. Delivery of SiRNA to the Mouse Brain by Systemic Injection of Targeted Exosomes. Nat. Biotechnol. 2011, 29, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Sellers, D.L.; Bergen, J.M.; Johnson, R.N.; Back, H.; Ravits, J.M.; Horner, P.J.; Pun, S.H. Targeted Axonal Import (TAxI) Peptide Delivers Functional Proteins into Spinal Cord Motor Neurons after Peripheral Administration. Proc. Natl. Acad. Sci. USA 2016, 113, 2514–2519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Z.; Xu, Y.; Deng, W.; Zhang, L.; Zhu, H.; Yu, P.; Qu, Y.; Zhao, W.; Han, Y.; Qin, C. Brain Derived Exosomes Are a Double-Edged Sword in Alzheimer’s Disease. Front. Mol. Neurosci. 2020, 13, 79. [Google Scholar] [CrossRef] [PubMed]
- Hao, D.; Ma, B.; He, C.; Liu, R.; Farmer, D.L.; Lam, K.S.; Wang, A. Surface Modification of Polymeric Electrospun Scaffolds via a Potent and High-Affinity Integrin A4β1 Ligand Improved the Adhesion, Spreading and Survival of Human Chorionic Villus-Derived Mesenchymal Stem Cells: A New Insight for Fetal Tissue Engineering. J. Mater. Chem. B 2020, 8, 1649–1659. [Google Scholar] [CrossRef] [PubMed]
- Reza-Zaldivar, E.E.; Hernández-Sapiéns, M.A.; Minjarez, B.; Gutiérrez-Mercado, Y.K.; Márquez-Aguirre, A.L.; Canales-Aguirre, A.A. Potential Effects of MSC-Derived Exosomes in Neuroplasticity in Alzheimer’s Disease. Front. Cell Neurosci. 2018, 12, 317. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wu, J.; Wu, J.; Fan, Q.; Zhou, J.; Wu, J.; Liu, S.; Zang, J.; Ye, J.; Xiao, M.; et al. Exosome-Mediated Targeted Delivery of MiR-210 for Angiogenic Therapy after Cerebral Ischemia in Mice. J. Nanobiotechnol. 2019, 17, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chivero, E.T.; Liao, K.; Niu, F.; Tripathi, A.; Tian, C.; Buch, S.; Hu, G. Engineered Extracellular Vesicles Loaded With MiR-124 Attenuate Cocaine-Mediated Activation of Microglia. Front. Cell Dev. Biol. 2020, 0. [Google Scholar] [CrossRef] [PubMed]
- Vanover, M.; Pivetti, C.; Lankford, L.; Kumar, P.; Galganski, L.; Kabagambe, S.; Keller, B.; Becker, J.; Chen, Y.J.; Chung, K.; et al. High Density Placental Mesenchymal Stromal Cells Provide Neuronal Preservation and Improve Motor Function Following In utero Treatment of Ovine Myelomeningocele. J. Pediatr. Surg. 2019, 54, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Kabagambe, S.; Keller, B.; Becker, J.; Goodman, L.; Pivetti, C.; Lankford, L.; Chung, K.; Lee, C.; Chen, Y.J.; Kumar, P.; et al. Placental Mesenchymal Stromal Cells Seeded on Clinical Grade Extracellular Matrix Improve Ambulation in Ovine Myelomeningocele. J. Pediatr. Surg. 2017, 53, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Xiao, Z.; Li, X.; Zhao, H.; Wang, B.; Qiu, Z.; Li, Z.; Mei, X.; Xu, B.; Fan, C.; et al. Human Placenta-Derived Mesenchymal Stem Cells Loaded on Linear Ordered Collagen Scaffold Improves Functional Recovery after Completely Transected Spinal Cord Injury in Canine. Sci. China Life Sci. 2018, 61, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Silva, M.; Feng, C.; Zhao, S.; Liu, L.; Li, S.; Zhong, J.; Zheng, W. Exosomes Derived from Human Placental Mesenchymal Stem Cells Enhanced the Recovery of Spinal Cord Injury by Activating Endogenous Neurogenesis. Stem Cell Res. Ther. 2021, 12, 174. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, Y.; Mu, J.; Chen, J.; Zhang, C.; Cao, H.; Gao, J. Transplantation of Human Mesenchymal Stem-Cell-Derived Exosomes Immobilized in an Adhesive Hydrogel for Effective Treatment of Spinal Cord Injury. Nano Lett. 2020, 20, 4298–4305. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, C.; Xu, Y.; Li, C.; Cao, Y.; Li, P. Exosomes Derived from Human Placenta-Derived Mesenchymal Stem Cells Improve Neurologic Function by Promoting Angiogenesis after Spinal Cord Injury. Neurosci. Lett. 2020, 739, 135399. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Qi, Y.; Sun, S.; Tao, Y.; Shi, R. Mesenchymal Stem Cell-Derived Exosomes: Hope for Spinal Cord Injury Repair. Stem Cells Dev. 2020, 29, 1467–1478. [Google Scholar] [CrossRef] [PubMed]
- Bydon, M. CELLTOP Part. II: A Phase II Clinical Trial of Autologous Adipose Derived Mesenchymal Stem Cells in the Treatment of Paralysis Due to Traumatic Spinal Cord Injury. Available online: http://www.clinicaltrials.govgov (accessed on 10 September 2021).
- Santos, R.R. dos Randomized Clinical Trial for the Evaluation of Autologous Mesenchymal Stem Cells Transplantation in Thoracolumbar Chronic and Complete Spinal Cord Injury. Available online: http://www.clinicaltrials.govgov (accessed on 10 September 2021).
- The Foundation for Orthopaedics and Regenerative Medicine Safety of Cultured Autologous Adult Adipose Derived Mesenchymal Stem Cell Intrathecal Injection for the Treatment of Spinal Cord Injury. Available online: https://clinicaltrials.gov/ct2/show/NCT05018793 (accessed on 10 September 2021).
- Wu, M.; Zhang, R.; Zou, Q.; Chen, Y.; Zhou, M.; Li, X.; Ran, R.; Chen, Q. Comparison of the Biological Characteristics of Mesenchymal Stem Cells Derived from the Human Placenta and Umbilical Cord. Sci. Rep. 2018, 8, 5014. [Google Scholar] [CrossRef] [PubMed]
- Moinuddin, F.M.; Yolcu, Y.U.; Wahood, W.; Siddiqui, A.M.; Chen, B.K.; Alvi, M.A.; Goyal, A.; Nesbitt, J.J.; Windebank, A.J.; Yeh, J.; et al. Early and Sustained Improvements in Motor Function in Rats after Infusion of Allogeneic Umbilical Cord-Derived Mesenchymal Stem Cells Following Spinal Cord Injury. Spinal Cord 2021, 59, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Mebarki, M.; Abadie, C.; Larghero, J.; Cras, A. Human Umbilical Cord-Derived Mesenchymal Stem/Stromal Cells: A Promising Candidate for the Development of Advanced Therapy Medicinal Products. Stem Cell Res. Ther. 2021, 12, 152. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Li, W.; Rong, D.; Xu, Z.; Zhang, Z.; Ye, H.; Xie, L.; Wu, Y.; Zhang, Y.; Wang, X. Human Umbilical Cord Mesenchymal Stem Cells-Derived Extracellular Vesicles Facilitate the Repair of Spinal Cord Injury via the MiR-29b-3p/PTEN/Akt/MTOR Axis. Cell Death Discov. 2021, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Fu, C.; Xiong, F.; He, C.; Wei, Q. Stem Cell Therapy for Spinal Cord Injury. Cell Transplant. 2021, 30. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Liu, H.; Zhang, K.; Xiao, D.-J.; Wang, C.; Wang, Y.-S. Adipose-Derived Stromal Cells Improve Functional Recovery after Spinal Cord Injury through TGF-Β1/Smad3/PLOD2 Pathway Activation. Aging 2021, 13, 4370–4387. [Google Scholar] [CrossRef] [PubMed]
- Farmer, D.L. Phase 1/2a Trial of Placental Mesenchymal Stem Cells for Repair of Fetal Myelomeningocele. Available online: http://www.clinicaltrials.govgov (accessed on 10 September 2021).
- Ringden, O.; Baygan, A.; Remberger, M.; Gustafsson, B.; Winiarski, J.; Khoein, B.; Moll, G.; Klingspor, L.; Westgren, M.; Sadeghi, B. Placenta-Derived Decidua Stromal Cells for Treatment of Severe Acute Graft-Versus-Host Disease. Stem Cells Transl. Med. 2018, 7, 325–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nevala-Plagemann, C.; Lee, C.; Tolar, J. Placenta-Based Therapies for the Treatment of Epidermolysis Bullosa. Cytotherapy 2015, 17, 786–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Institute of Cell Therapy Treatment of Coronavirus COVID-19 Pneumonia (Pathogen SARS-CoV-2) with Cryopreserved Allogeneic Multipotent Mesenchymal Stem Cells of the Placenta and Umbilical Cord. Available online: http://www.clinicaltrials.govgov (accessed on 10 September 2021).
- Beijing Tongren Hospital Phase I Clinical Trials of Human Placental Mesenchymal Stem Cells Treatment on Diabetic Foot Ulcer. Available online: http://www.clinicaltrials.govgov (accessed on 10 September 2021).
- Institute of Cell Therapy Efficacy of Bone-Marrow-Derived and Placenta-Derived Multipotent Mesenchymal Stem/Stromal Cells for Osteoarthritis. Available online: http://www.clinicaltrials.gov (accessed on 10 September 2021).



{kind=link}
{kind=link}
| Trial Identifier | Name | Phase | Key Inclusion Criteria | Therapy |
|---|---|---|---|---|
| NCT03505034 | Intrathecal Transplantation of UC-MSC in Patients with Late Stage of Chronic Spinal Cord Injury | 2 | Age 18–65 ASIA Scale A–D Injury more than 12 months | Intrathecal Transplantation of UC-MSCs, 106 cells/kg, once a month for 4 months |
| NCT03521336 | Intrathecal Transplantation of UC-MSC in Patients with Sub-Acute Spinal Cord Injury | 2 | Age 18–65 ASIA Scale A–D Injury 2 weeks to 2 months | Intrathecal Transplantation of UC-MSCs, 106 cells/kg, once a month for 4 months |
| NCT03521323 | Intrathecal Transplantation of UC-MSC in Patients with Early Stage of Chronic Spinal Cord Injury | 2 | Age 18–65 ASIA Scale A–D Injury 2 months to 12 months | Intrathecal Transplantation of UC-MSCs 106 cells/kg, once a month for 4 months |
| NCT02574585 | Autologous Mesenchymal Stem Cells Transplantation in Thoracolumbar Chronic and Complete Spinal Cord Injury Spinal Cord Injury | 2 | Age 18–65 ASIA Scale A Injury T1-L2 with at least 12 months | Two percutaneous injections of MSCs, with a 3-month interval between the injections |
| NCT05018793 | Safety of Cultured Autologous Adult Adipose-Derived Mesenchymal Stem Cell Intrathecal Injection for SCI | 1 | Diagnosis of Spinal Cord Injury | Single intrathecal injection of 100 million cells |
| NCT02688049 | NeuroRegen Scaffold™ Combined with Stem Cells for Chronic Spinal Cord Injury Repair | 1, 2 | Age 18–65 ASIA Scale A Injury C5-T12 | Patients with chronic SCI (ASIA grade A) will receive NeuroRegen Scaffold with 10 million MSCs transplantation after localized scars cleared |
| NCT04520373 | Autologous Adipose-Derived Mesenchymal Stem Cells for Spinal Cord Injury Patients | 2 | Age >18 ASIA Scale A, B | Patients will receive a single dose of autologous, adipose-derived MSCs one time |
| NCT03308565 | Adipose Stem Cells for Traumatic Spinal Cord Injury (CELLTOP) | 1 | Age >18 ASIA Scale A, B Injury 2 weeks to 1 year | Patients will receive a single dose of 100 million autologous, adipose derived MSCs |
| NCT02917291 | Safety and Preliminary Efficacy of FAB117-HC in Patients with Acute Traumatic Spinal Cord Injury | 1, 2 | Age 16–70 Phase 1—ASIA A, Injury 72–120 h prior Phase 2—ASIA A, B, Injury up to 96 h prior | Intramedullary administration of FAB117-HC (Adipose Derived Adult MSCs Expanded and Pulsed with H2O2) |
| NCT04213131 | Efficacy and Safety of hUC-MSCs and hUCB-MSCs in the Treatment of Chronic Spinal Cord Injury | n/a | Age 20–65 ASIA A-D with no change Injury over 1 year | Intravenous, lumbar, and local administration of hUC-MSCs/hUCB-MSCs |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulubya, E.S.; Clark, K.; Hao, D.; Lazar, S.; Ghaffari-Rafi, A.; Karnati, T.; Ebinu, J.O.; Zwienenberg, M.; Farmer, D.L.; Wang, A. The Unique Properties of Placental Mesenchymal Stromal Cells: A Novel Source of Therapy for Congenital and Acquired Spinal Cord Injury. Cells 2021, 10, 2837. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10112837
Kulubya ES, Clark K, Hao D, Lazar S, Ghaffari-Rafi A, Karnati T, Ebinu JO, Zwienenberg M, Farmer DL, Wang A. The Unique Properties of Placental Mesenchymal Stromal Cells: A Novel Source of Therapy for Congenital and Acquired Spinal Cord Injury. Cells. 2021; 10(11):2837. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10112837
Chicago/Turabian StyleKulubya, Edwin S, Kaitlin Clark, Dake Hao, Sabrina Lazar, Arash Ghaffari-Rafi, Tejas Karnati, Julius Okudu Ebinu, Marike Zwienenberg, Diana L Farmer, and Aijun Wang. 2021. "The Unique Properties of Placental Mesenchymal Stromal Cells: A Novel Source of Therapy for Congenital and Acquired Spinal Cord Injury" Cells 10, no. 11: 2837. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10112837