
Hypoxic Incubation Conditions for Optimized Manufacture of Tenocyte-Based Active Pharmaceutical Ingredients of Homologous Standardized Transplant Products in Tendon Regenerative Medicine

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Primary Cell Sources and In Vitro Culture Conditions
2.2. Timecourse of HIF-1 Induction with Western Blotting
2.3. hFPT Comparative Proliferation Assays
2.4. hFPT Comparative Surface Marker Panel Characterization by Flow Cytometry
- CD90 (PE Mouse Anti-Human CD90, BD Biosciences, USA, ref. 561970);
- CD73 (PE Mouse Anti-Human CD73, BD Biosciences, USA, ref. 550257);
- CD105 (PE Mouse Anti-Human CD105, BD Biosciences, USA, ref. 560839);
- CD19 (PE Mouse Anti-Human CD19, BD Biosciences, USA, ref. 555413);
- CD34 (PE Mouse Anti-Human CD34, BD Biosciences, USA, ref. 560941);
- CD14 (FITC Mouse Anti-Human CD14, BD Biosciences, USA, ref. 555397);
- CD45 (FITC Mouse Anti-Human CD45, BD Biosciences, USA, ref. 560976);
- CD44 (PE Mouse Anti-Human CD44, BD Biosciences, USA, ref. 550989);
- CD26 (PE Mouse Anti-Human CD26, BD Biosciences, USA, ref. 555437);
- CD166 (PE Mouse Anti-Human CD166, BD Biosciences, USA, ref. 559263);
- MHC class I HLA-ABC (PE Mouse Anti-Human HLA-ABC, BD Biosciences, USA, ref. 560964);
- MHC class II HLA-DR,-DP,-DQ (FITC Mouse Anti-Human HLA-DR,-DP,-DQ, BD Biosciences, USA, ref. 555558);
- IgG1 isotype (PE Mouse IgG1, κ Isotype Control, BD Biosciences, USA, ref. 555749);
- IgG1 isotype (FITC Mouse IgG1, κ Isotype Control, BD Biosciences, USA, ref. 554679);
- IgG2a isotype (FITC Mouse IgG2a, κ Isotype Control, BD Biosciences, USA, ref. 555573).
2.5. hFPT Comparative Phenotypic Stability in Chemical Adipogenesis and Osteogenesis Induction Models
2.5.1. Comparative Chemical Adipogenic Differentiation Assay
2.5.2. Comparative Chemical Osteogenic Differentiation Assay
2.6. hFPT Comparative Proteomic Analysis by Quantitative Mass Spectrometry
2.6.1. Sample Preparation for LC-MS/MS
2.6.2. LC-MS/MS Processing of Samples
2.6.3. LC-MS/MS Data Analysis
2.7. Specific ECM Protein Quantification in hFPT Lysates
2.7.1. Collagen I Western Blotting
2.7.2. hFPT Freeze-Thaw Lysate Preparation
2.7.3. Comparative Endpoint Quantification of Elastin by Colorimetry
2.7.4. Comparative Endpoint Quantification of Fibronectin by ELISA
2.7.5. Comparative Endpoint Quantification of Total GAGs by Colorimetry
2.8. Statistical Analysis
3. Results
3.1. Hypoxia Transiently Induces HIF-1α Expression in hFPTs
3.2. hFPT Comparative Proliferation Assays and Cell Size Distribution Analysis
3.3. hFPT Comparative Surface Marker Panel Characterization by Flow Cytometry
3.4. hFPT Comparative Phenotypic Stability in Chemical Adipogenesis and Osteogenesis Induction Models
3.5. hFPT Comparative Proteomic Analysis by Quantitative Mass Spectrometry
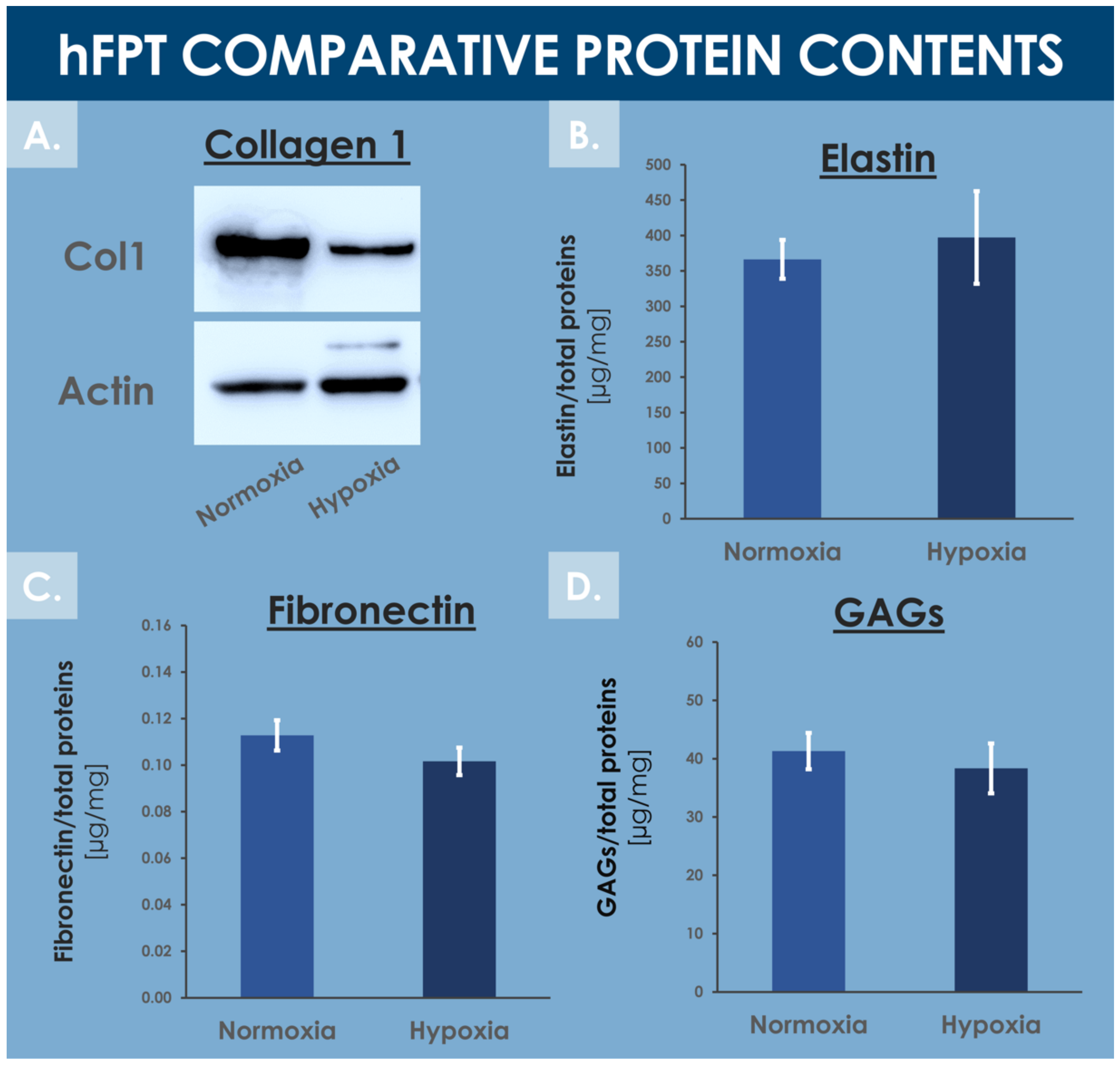
3.6. hFPT Specific ECM Protein Comparative Endpoint Analysis
4. Discussion
4.1. Conservation of hFPT Critical Quality Attributes in Hypoxic Culture Conditions as Compared to Normoxia Culture Conditions
4.2. Optimization of hFPT Key Quality Attributes in Hypoxic Culture Conditions as Compared to Normoxia Culture Conditions
4.3. hFPT HIF-1α Pathway Transient Induction and Collagen Downregulation in Hypoxia Culture Conditions
4.4. Specificities of hFPT Hypoxic Culture Conditions for Potential Next-Generation Manufacture of Cell-Free APIs in Tendon Regenerative Medicine
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| API | active pharmaceutical ingredient |
| ASC | adipose-derived stem cells |
| CD | cluster of differentiation |
| CHUV | Centre hospitalier universitaire vaudois |
| CM-FBS | complete medium with FBS |
| CM-HPL | complete medium with HPL |
| CPP | critical process parameter |
| CQA | critical quality attribute |
| DMEM | Dulbecco’s modified Eagle medium |
| DTT | dithiothreitol |
| ECM | extracellular matrix |
| EPFL | École polytechnique fédérale de Lausanne |
| ETC | electron transport chain |
| FACS | fluorescence-activated cell sorting |
| FASP | filter-aided sample preparation |
| FBS | fetal bovine serum |
| FC | fold change |
| FDR | false discovery rate |
| FITC | fluorescein isothiocyanate |
| FSC | forward scatter |
| GAG | glycosaminoglycan |
| GMP | good manufacturing practices |
| GO | gene ontology |
| HCD | high energy collision dissociation |
| hFPT | human fetal progenitor tenocytes |
| HPL | human platelet lysate |
| IBMX | 3-isobutyl-1-methylxanthine |
| IPC | in-process control |
| ITS | Insulin–Transferrin–Selenium |
| KPP | key process parameter |
| KQA | key quality attribute |
| MCB | master cell bank |
| MHC | major histocompatibility complex |
| MS | mass spectrometry |
| MSC | mesenchymal stem cell |
| NCE | normalized collision energy |
| PBS | phosphate-buffered saline |
| PCA | principal component analysis |
| PCB | parental cell bank |
| PE | phycoerythrin |
| PPC | post-process control |
| PRP | platelet-rich plasma |
| ROS | reactive oxygen species |
| SD | standard deviation |
| SDS | sodium dodecyl sulfate |
| SSC | side scatter |
| TCA | tricarboxylic acid cycle |
| TMT | tandem mass tag |
| USA | United States of America |
| VitCp | L-Ascorbic acid 2-phosphate sesquimagnesium salt hydrate |
| WCB | working cell bank |
References
- Grognuz, A.; Scaletta, C.; Farron, A.; Raffoul, W.; Applegate, L.A. Human fetal progenitor tenocytes for regenerative medicine. Cell Transplant. 2016, 25, 463–479. [Google Scholar] [CrossRef] [Green Version]
- Grognuz, A.; Scaletta, C.; Farron, A.; Pioletti, D.P.; Raffoul, W.; Applegate, L.A. Stability enhancement using hyaluronic acid gels for delivery of human fetal progenitor tenocytes. Cell Med. 2016, 8, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Laurent-Applegate, L.; Grognuz, A.; Hirt-Burri, N.; Petrou, I.G.; Raffoul, W. Cell therapies for tendons: Old cell choice for modern innovation. Swiss Med. Wkly. 2014, 144, 13989. [Google Scholar] [CrossRef] [PubMed]
- Aeberhard, P.; Grognuz, A.; Peneveyre, C.; McCallin, S.; Hirt-Burri, N.; Antons, J.; Pioletti, D.; Raffoul, W.; Applegate, L.A. Efficient decellularization of equine tendon with preserved biomechanical properties and cytocompatibility for human tendon surgery indications. Artif. Organs 2020, 44, E161–E171. [Google Scholar] [CrossRef]
- Laurent, A.; Abdel-Sayed, P.; Grognuz, A.; Scaletta, C.; Hirt-Burri, N.; Michetti, M.; Roessingh, A.D.B.; Raffoul, W.; Kronen, P.; Nuss, K.; et al. Industrial development of standardized fetal progenitor cell therapy for tendon regenerative medicine: Preliminary safety in xenogeneic transplantation. Biomedicines 2021, 9, 380. [Google Scholar] [CrossRef]
- Fujikawa, K.; Ohtani, T.; Matsumoto, H.; Seedhom, B.B. Reconstruction of the extensor apparatus of the knee with the Leeds-Keio ligament. J. Bone Joint Surg. Br. 1994, 76, 200–203. [Google Scholar] [CrossRef] [Green Version]
- Laurent, A.; Lin, P.; Scaletta, C.; Hirt-Burri, N.; Michetti, M.; Roessingh, A.S.D.B.; Raffoul, W.; She, B.-R.; Applegate, L.A. Bringing safe and standardized cell therapies to industrialized processing for burns and wounds. Front. Bioeng. Biotechnol. 2020, 8, 581. [Google Scholar] [CrossRef] [PubMed]
- Laurent, A.; Scaletta, C.; Michetti, M.; Hirt-Burri, N.; Roessingh, A.S.D.B.; Raffoul, W.; Applegate, L.A. GMP tiered cell banking of non-enzymatically isolated dermal progenitor fibroblasts for allogenic regenerative medicine. In Methods in Molecular Biology; Springer: Berlin/Heidelberg, Germany, 2020; pp. 25–48. [Google Scholar]
- Grayson, W.L.; Zhao, F.; Izadpanah, R.; Bunnell, B.; Ma, T. Effects of hypoxia on human mesenchymal stem cell expansion and plasticity in 3D constructs. J. Cell. Physiol. 2006, 207, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.-M.; Won, C.-H.; Sung, J.-H. Responses of adipose-derived stem cells during hypoxia: Enhanced skin-regenerative potential. Expert Opin. Biol. Ther. 2009, 9, 1499–1508. [Google Scholar] [CrossRef]
- Das, R.; Jahr, H.; Van Osch, G.J.; Farrell, E. The role of hypoxia in bone marrow–derived mesenchymal stem cells: Considerations for regenerative medicine approaches. Tissue Eng. Part B Rev. 2010, 16, 159–168. [Google Scholar] [CrossRef]
- He, J.; Genetos, D.C.; Yellowley, C.E.; Leach, J.K. Oxygen tension differentially influences osteogenic differentiation of human adipose stem cells in 2D and 3D cultures. J. Cell. Biochem. 2010, 110, 87–96. [Google Scholar] [CrossRef]
- Verloop, R.E. Progenitor Cells and Hypoxia in Angiogenesis. Ph.D. Thesis, Vrije Universiteit Amsterdam, Amsterdam, The Netherlands, 2011. [Google Scholar]
- Frazier, T.P.; Gimble, J.M.; Kheterpal, I.; Rowan, B.G. Impact of low oxygen on the secretome of human adipose-derived stromal/stem cell primary cultures. Biochimie 2013, 95, 2286–2296. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.R.; Pingguan-Murphy, B.; Abas, W.A.B.W.; Azmi, M.A.N.; Omar, S.Z.; Chua, K.H.; Safwani, W.K.Z.W. Impact of low oxygen tension on stemness, proliferation and differentiation potential of human adipose-derived stem cells. Biochem. Biophys. Res. Commun. 2014, 448, 218–224. [Google Scholar] [CrossRef]
- Galeano-Garces, C.; Camilleri, E.; Riester, S.M.; Dudakovic, A.; Larson, D.R.; Qu, W.; Smith, J.; Dietz, A.B.; Im, H.-J.; Krych, A.J.; et al. Molecular validation of chondrogenic differentiation and hypoxia responsiveness of platelet-lysate expanded adipose tissue–derived human mesenchymal stromal cells. Cartilage 2017, 8, 283–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.C.; Kim, K.-H.; Kim, O.-H.; Lee, S.K.; Hong, H.-E.; Won, S.S.; Jeon, S.-J.; Choi, B.J.; Jeong, W.; Kim, S.-J. Determination of optimized oxygen partial pressure to maximize the liver regenerative potential of the secretome obtained from adipose-derived stem cells. Stem Cell Res. Ther. 2017, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.-H.; Kim, M.-Y.; Yoon, Y.-S.; Koh, D.-I.; Cho, S.-Y.; Kim, K.-S.; Hur, M.-W. Hypoxia-induced RelA/p65 derepresses SLC16A3 (MCT4) by downregulating ZBTB7A. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Deep, G. Hypoxia in tumor microenvironment regulates exosome biogenesis: Molecular mechanisms and translational opportunities. Cancer Lett. 2020, 479, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Mazzatti, D.; Lim, F.-L.; O’Hara, A.; Wood, I.S.; Trayhurn, P. A microarray analysis of the hypoxia-induced modulation of gene expression in human adipocytes. Arch. Physiol. Biochem. 2012, 118, 112–120. [Google Scholar] [CrossRef] [Green Version]
- D’Alessandro, S.; Magnavacca, A.; Perego, F.; Fumagalli, M.; SanGiovanni, E.; Prato, M.; Dell’Agli, M.; Basilico, N. Effect of hypoxia on gene expression in cell populations involved in wound healing. BioMed Res. Int. 2019, 2019, 2626374. [Google Scholar] [CrossRef] [Green Version]
- Lee, P.; Chandel, N.S.; Simon, M.C. Cellular adaptation to hypoxia through hypoxia inducible factors and beyond. Nat. Rev. Mol. Cell Biol. 2020, 21, 268–283. [Google Scholar] [CrossRef] [PubMed]
- Wheaton, W.W.; Chandel, N.S. Hypoxia. 2. Hypoxia regulates cellular metabolism. Am. J. Physiol. Physiol. 2011, 300, C385–C393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.; Kim, S.-M.; Sung, J.-H. Cellular and molecular stimulation of adipose-derived stem cells under hypoxia. Cell Biol. Int. 2014, 38, 553–562. [Google Scholar] [CrossRef]
- Beegle, J.; Lakatos, K.; Kalomoiris, S.; Stewart, H.; Isseroff, R.R.; Nolta, J.A.; Fierro, F.A. Hypoxic preconditioning of mesenchymal stromal cells induces metabolic changes, enhances survival, and promotes cell retention in vivo. Stem Cells 2015, 33, 1818–1828. [Google Scholar] [CrossRef]
- Riis, S.; Stensballe, A.; Emmersen, J.; Pennisi, C.P.; Birkelund, S.; Zachar, V.; Fink, T. Mass spectrometry analysis of adipose-derived stem cells reveals a significant effect of hypoxia on pathways regulating extracellular matrix. Stem Cell Res. Ther. 2016, 7, 52. [Google Scholar] [CrossRef] [Green Version]
- Safwani, W.K.Z.W.; Choi, J.R.; Yong, K.W.; Ting, I.; Adenan, N.A.M.; Pingguan-Murphy, B. Hypoxia enhances the viability, growth and chondrogenic potential of cryopreserved human adipose-derived stem cells. Cryobiology 2017, 75, 91–99. [Google Scholar] [CrossRef]
- Hu, X.; Yu, S.P.; Fraser, J.L.; Lu, Z.; Ogle, M.E.; Wang, J.-A.; Wei, L. Transplantation of hypoxia-preconditioned mesenchymal stem cells improves infarcted heart function via enhanced survival of implanted cells and angiogenesis. J. Thorac. Cardiovasc. Surg. 2008, 135, 799–808. [Google Scholar] [CrossRef] [Green Version]
- Abdollahi, H.; Harris, L.J.; Zhang, P.; McIlhenny, S.; Srinivas, V.; Tulenko, T.; DiMuzio, P.J. The role of hypoxia in stem cell differentiation and therapeutics. J. Surg. Res. 2011, 165, 112–117. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.-S.; Sung, J.-H. Hypoxic culturing enhances the wound-healing potential of adipose-derived stem cells. Adv. Wound Care 2012, 1, 172–176. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.Y.; Lui, P.P.Y.; Rui, Y.F. Hypoxia-mediated efficient expansion of human tendon-derived stem cells in vitro. Tissue Eng. Part A 2012, 18, 484–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, N.; Rahman, M.T.; Abu Kasim, N.H.; Alabsi, A. Hypoxic culture conditions as a solution for mesenchymal stem cell based regenerative therapy. Sci. World J. 2013, 2013, 632972. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Fujita, M.; Tanaka, Y.; Kojima, I.; Kanatani, Y.; Ishihara, M.; Tachibana, S. Low oxygen tension enhances proliferation and maintains stemness of adipose tissue-derived stromal cells. BioResearch Open Access 2013, 2, 199–205. [Google Scholar] [CrossRef]
- Hsiao, S.; Dilley, R.J.; Dusting, G.J.; Lim, S.Y. Ischemic preconditioning for cell-based therapy and tissue engineering. Pharmacol. Ther. 2014, 142, 141–153. [Google Scholar] [CrossRef]
- Korski, K.I.; Kubli, D.A.; Wang, B.J.; Khalafalla, F.G.; Monsanto, M.M.; Firouzi, F.; Echeagaray, O.H.; Kim, T.; Adamson, R.M.; Dembitsky, W.P.; et al. Hypoxia prevents mitochondrial dysfunction and senescence in human c-Kit+ cardiac progenitor cells. Stem Cells 2019, 37, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, B.; Zhang, W.J.; Zhou, G.; Cao, Y.; Liu, W. Enhanced proliferation capacity of porcine tenocytes in low O2 tension culture. Biotechnol. Lett. 2009, 32, 181–187. [Google Scholar] [CrossRef]
- Martin-Rendon, E.; Hale, S.J.; Ryan, D.; Baban, D.; Forde, S.P.; Roubelakis, M.G.; Sweeney, D.; Moukayed, M.; Harris, A.L.; Davies, K.; et al. Transcriptional profiling of human cord blood CD133+ and cultured bone marrow mesenchymal stem cells in response to hypoxia. Stem Cells 2007, 25, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Luo, F.; Yang, L.; Wu, W.; Liu, X. Hypoxia stimulates the expression of macrophage migration inhibitory factor in human vascular smooth muscle cells via HIF-1α dependent pathway. BMC Cell Biol. 2010, 11, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tello, D.; Balsa, E.; Acosta-Iborra, B.; Fuertes-Yebra, E.; Elorza, A.; Ordóñez, Á.; Corral-Escariz, M.; Soro, I.; López-Bernardo, E.; Perales-Clemente, E.; et al. Induction of the mitochondrial NDUFA4L2 protein by HIF-1α decreases oxygen consumption by inhibiting complex I activity. Cell Metab. 2011, 14, 768–779. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. HIF-1 mediates metabolic responses to intratumoral hypoxia and oncogenic mutations. J. Clin. Investig. 2013, 123, 3664–3671. [Google Scholar] [CrossRef] [Green Version]
- Dengler, V.L.; Galbraith, M.; Espinosa, J.M. Transcriptional regulation by hypoxia inducible factors. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morotti, M.; Bridges, E.; Valli, A.; Choudhry, H.; Sheldon, H.; Wigfield, S.; Gray, N.; Zois, C.E.; Grimm, F.; Jones, D.; et al. Hypoxia-induced switch in SNAT2/SLC38A2 regulation generates endocrine resistance in breast cancer. Proc. Natl. Acad. Sci. USA 2019, 116, 12452–12461. [Google Scholar] [CrossRef] [Green Version]
- Thangarajah, H.; Vial, I.N.; Chang, E.; El-Ftesi, S.; Januszyk, M.; Chang, E.I.; Paterno, J.; Neofytou, E.; Longaker, M.T.; Gurtner, G.C. IFATS collection: Adipose stromal cells adopt a proangiogenic phenotype under the influence of hypoxia. Stem Cells 2009, 27, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-A.; He, A.; Hu, X.; Jiang, Y.; Sun, Y.; Jiang, J.; Gui, C.; Wang, Y.; Chen, H. Anoxic preconditioning: A way to enhance the cardioprotection of mesenchymal stem cells. Int. J. Cardiol. 2009, 133, 410–412. [Google Scholar] [CrossRef]
- Zhao, F.; Grayson, W.L.; Ma, T.; Irsigler, A. Perfusion affects the tissue developmental patterns of human mesenchymal stem cells in 3D scaffolds. J. Cell. Physiol. 2009, 219, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.-F.; Yew, T.-L.; Chiang, E.-R.; Ma, H.-L.; Hsu, C.-Y.; Hsu, S.-H.; Hsu, Y.-T.; Hung, S.-C. Mesenchymal stem cells from a hypoxic culture improve and engraft achilles tendon repair. Am. J. Sports Med. 2013, 41, 1117–1125. [Google Scholar] [CrossRef]
- Koziel, A.; Jarmuszkiewicz, W. Hypoxia and aerobic metabolism adaptations of human endothelial cells. Pflugers Arch. 2017, 469, 815–827. [Google Scholar] [CrossRef] [Green Version]
- Laurent, A.; Hirt-Burri, N.; Scaletta, C.; Michetti, M.; Roessingh, A.S.D.B.; Raffoul, W.; Applegate, L.A. Holistic approach of Swiss fetal progenitor cell banking: Optimizing safe and sustainable substrates for regenerative medicine and biotechnology. Front. Bioeng. Biotechnol. 2020, 8, 557758. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Kulak, N.; Pichler, G.; Paron, I.; Nagaraj, N.; Mann, M. Minimal, encapsulated proteomic-sample processing applied to copy-number estimation in eukaryotic cells. Nat. Methods 2014, 11, 319–324. [Google Scholar] [CrossRef]
- Dorfer, V.; Pichler, P.; Stranzl, T.; Stadlmann, J.; Taus, T.; Winkler, S.; Mechtler, K. MS Amanda, a universal identification algorithm optimized for high accuracy tandem mass spectra. J. Proteome Res. 2014, 13, 3679–3684. [Google Scholar] [CrossRef]
- Kong, A.T.; LePrevost, F.V.; Avtonomov, D.M.; Mellacheruvu, D.; Nesvizhskii, A.I. MS Fragger: Ultrafast and comprehensive peptide identification in mass spectrometry–based proteomics. Nat. Methods 2017, 14, 513–520. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 22 September 2021).
- Plubell, D.; Wilmarth, P.A.; Zhao, Y.; Fenton, A.M.; Minnier, J.; Reddy, A.P.; Klimek, J.; Yang, X.; David, L.L.; Pamir, N. Extended multiplexing of tandem mass tags (TMT) labeling reveals age and high fat diet specific proteome changes in mouse epididymal adipose tissue. Mol. Cell. Proteom. 2017, 16, 873–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Klopfenstein, D.V.; Zhang, L.; Pedersen, B.S.; Ramírez, F.; Vesztrocy, A.W.; Naldi, A.; Mungall, C.J.; Yunes, J.M.; Botvinnik, O.; Weigel, M.; et al. GOATOOLS: A python library for gene ontology analyses. Sci. Rep. 2018, 8, 10872. [Google Scholar] [CrossRef]
- Uchida, T.; Rossignol, F.; Matthay, M.A.; Mounier, R.; Couette, S.; Clottes, E.; Clerici, C. Prolonged hypoxia differentially regulates hypoxia-inducible factor (HIF)-1α and HIF-2α expression in lung epithelial cells. J. Biol. Chem. 2004, 279, 14871–14878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-W.; Tchernyshyov, I.; Semenza, G.L.; Dang, C.V. HIF-1-mediated expression of pyruvate dehydrogenase kinase: A metabolic switch required for cellular adaptation to hypoxia. Cell Metab. 2006, 3, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Wood, S.M.; Wiesener, M.S.; Yeates, K.M.; Okada, N.; Pugh, C.; Maxwell, P.H.; Ratcliffe, P. Selection and analysis of a mutant cell line defective in the hypoxia-inducible factor-1 α-subunit (HIF-1α). J. Biol. Chem. 1998, 273, 8360–8368. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Choi, J.S.; Park, C.; Jeong, J.W. Ethyl pyruvate stabilizes hypoxia-inducible factor 1 alpha via stimulation of the TCA cycle. Cancer Let. 2010, 295, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zhang, M.; Weng, H. Induction of the mitochondrial NDUFA4L2 protein by HIF-1a regulates heart regeneration by promoting the survival of cardiac stem cell. Biochem. Biophys. Res. Commun. 2018, 503, 2226–2233. [Google Scholar] [CrossRef]
- Smits, V.A.; Cabrera, E.; Freire, R.; Gillespie, D. Claspin—Checkpoint adaptor and DNA replication factor. FEBS J. 2018, 286, 441–455. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Lam, W.H.; Zhai, Y.; Cheng, J.; Cheng, E.; Zhao, Y.; Gao, N.; Tye, B.-K. Structure of the origin recognition complex bound to DNA replication origin. Nat. Cell Biol. 2018, 559, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Nikonova, A.S.; Astsaturov, I.; Serebriiskii, I.G.; Dunbrack, R.; Golemis, E. Aurora A kinase (AURKA) in normal and pathological cell division. Cell. Mol. Life Sci. 2013, 70, 661–687. [Google Scholar] [CrossRef]
- Fabbro, M.; Zhou, B.-B.; Takahashi, M.; Sarcevic, B.; Lal, P.; Graham, M.; Gabrielli, B.; Robinson, P.J.; Nigg, E.; Ono, Y.; et al. Cdk1/Erk2- and Plk1-dependent phosphorylation of a centrosome protein, Cep55, is required for its recruitment to midbody and cytokinesis. Dev. Cell 2005, 9, 477–488. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Bu, Y. SKA2/FAM33A: A novel gene implicated in cell cycle, tumorigenesis, and psychiatric disorders. Genes Dis. 2019, 6, 25–30. [Google Scholar] [CrossRef]
- Nakamura, K.; Sakaue, H.; Nishizawa, A.; Matsuki, Y.; Gomi, H.; Watanabe, E.; Hiramatsua, R.; Tamamori-Adachi, M.; Kitajima, S.; Noda, T.; et al. PDK1 regulates cell proliferation and cell cycle progression through control of cyclin D1 and p27Kip1 expression. J. Biol. Chem. 2008, 283, 17702–17711. [Google Scholar] [CrossRef] [Green Version]
- Politis, P.K.; Makri, G.; Thomaidou, D.; Geissen, M.; Rohrer, H.; Matsas, R. BM88/CEND1 coordinates cell cycle exit and differentiation of neuronal precursors. Proc. Natl. Acad. Sci. USA 2007, 104, 17861–17866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, M. Structure and metabolism of tendons. Scand. J. Med. Sci. Sports 2007, 7, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, C.T.; Screen, H.R.C. Tendon structure and composition. Adv. Exp. Med. Biol. 2016, 920, 3–10. [Google Scholar] [CrossRef]
- Vetro, S.W.; Bellanti, J.A. Fetal and neonatal immunoincompetence. Fetal Diagn. Ther. 1989, 4, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, J.H.-C. Human tendon stem cells better maintain their stemness in hypoxic culture conditions. PLoS ONE 2013, 8, e61424. [Google Scholar] [CrossRef] [Green Version]
- D’Ippolito, G.; Diabira, S.; Howard, G.A.; Roos, B.A.; Schiller, P.C. Low oxygen tension inhibits osteogenic differentiation and enhances stemness of human MIAMI cells. Bone 2006, 39, 513–522. [Google Scholar] [CrossRef]
- Kabat, M.; Bobkov, I.; Kumar, S.; Grumet, M. Trends in mesenchymal stem cell clinical trials 2004–2018: Is efficacy optimal in a narrow dose range? Stem Cells Transl. Med. 2020, 9, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Rezakhani, L.; Kelishadrokhi, A.F.; Soleimanizadeh, A.; Rahmati, S. Mesenchymal stem cell (MSC)-derived exosomes as a cell-free therapy for patients infected with COVID-19: Real opportunities and range of promises. Chem. Phys. Lipids 2021, 234, 105009. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]
- World Medical Association (WMA). Declaration of helsinki: Ethical principles for medical research involving human subjects. JAMA 2013, 310, 233–238. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population Doubling Values (Mean ± SD) | |||||
|---|---|---|---|---|---|
| Passage 6 Cells | p-Values | Passage 7 Cells | p-Values | ||
| Day 5 harvest | 21% O2 | 3.52 ± 0.22 | 0.012 * | 2.98 ± 0.24 | 0.092 |
| 2% O2 | 4.05 ± 0.12 | 3.30 ± 0.15 | |||
| Day 6 harvest | 21% O2 | 3.83 ± 0.32 | 0.020 * | 3.85 ± 0.12 | 0.165 |
| 2% O2 | 4.37 ± 0.27 | 4.08 ± 0.37 | |||
| Day 7 harvest | 21% O2 | 4.24 ± 0.23 | 0.016 * | 4.29 ± 0.12 | 0.015 * |
| 2% O2 | 4.59 ± 0.10 | 4.74 ± 0.19 | |||
| Protein Name | Accession Number | Protein Symbol | Log2 FC Hypoxia vs. Normoxia |
|---|---|---|---|
| NADH dehydrogenase [ubiquinone] 1 alpha subcomplex subunit 4-like 2 | Q9NRX3 | NDUFA4L2 | 2.1464 |
| Syndecan binding protein (Syntenin), isoform CRA_a | G5EA09 | SDCBP | 1.9118 |
| Alpha-2-macroglobulin | P01023 | A2M | 1.8585 |
| Ferritin heavy chain | P02794 | FTH1 | 1.7748 |
| Ferritin light chain | P02792 | FTL | 1.6041 |
| Nicotinate-nucleotide pyrophosphorylase [carboxylating] | Q15274 | QPRT | 1.4797 |
| Monocarboxylate transporter 4 | O15427 | SLC16A3 | 1.4795 |
| Lysosomal-associated transmembrane protein 4A | Q15012 | LAPTM4A | 1.4593 |
| Solute carrier family 2, facilitated glucose transporter member 1 | P11166 | SLC2A1 | 1.3763 |
| Spindle and kinetochore-associated protein 2 | J3KSP0 | SKA2 | 1.3565 |
| Transforming growth factor-beta-induced protein ig-h3 | Q15582 | TGFBI | 1.3464 |
| Lactadherin | Q08431 | MFGE8 | 1.3313 |
| Zinc finger protein 185 | O15231 | ZNF185 | 1.3219 |
| Mitogen-activated protein kinase 13 | O15264 | MAPK13 | 1.3148 |
| [Pyruvate dehydrogenase (acetyl-transferring)] kinase isozyme 1, mitochondrial | Q15118 | PDK1 | 1.2980 |
| Apolipoprotein B-100 | P04114 | APOB | 1.2616 |
| Metalloreductase STEAP1 | Q9UHE8 | STEAP1 | 1.2398 |
| Tetraspanin (fragment) | F8VWK8 | CD63 | 1.2375 |
| Fos-related antigen 1 | P15407 | FOSL1 | 1.2331 |
| Gamma-enolase | P09104 | ENO2 | 1.1619 |
| G-protein coupled receptor 39 | O43194 | GPR39 | 1.1402 |
| Cytoplasmic aconitate hydratase | P21399 | ACO1 | 1.1232 |
| Podocalyxin | O00592 | PODXL | 1.1188 |
| RNA-binding protein EWS (fragment) | H7BY36 | EWSR1 | 1.1169 |
| NPC intracellular cholesterol transporter 1 | O15118 | NPC1 | 1.1023 |
| Tetraspanin-3 | O60637 | TSPAN3 | 1.0965 |
| Repetin | Q6XPR3 | RPTN | 1.0841 |
| EGF-like repeat and discoidin I-like domain-containing protein 3 | O43854 | EDIL3 | 1.0480 |
| Sodium-coupled neutral amino acid transporter 2 | Q96QD8 | SLC38A2 | 1.0393 |
| Fructose-bisphosphate aldolase C | P09972 | ALDOC | 1.0192 |
| Desmocollin-2 | A0A3B3ISU0 | DSC2 | 1.0026 |
| Inter-alpha-trypsin inhibitor heavy chain H3 | A0A087WW43 | ITIH3 | 0.9987 |
| Hepatocyte growth factor activator | D6RAR4 | HGFAC | 0.9831 |
| Centrosomal protein of 55 kDa | Q53EZ4 | CEP55 | 0.9777 |
| Claspin | Q9HAW4 | CLSPN | 0.9771 |
| Macrophage migration inhibitory factor | P14174 | MIF | 0.9708 |
| Rho GTPase-activating protein 7 | Q96QB1 | DLC1 | 0.9628 |
| Aurora kinase A | O14965 | AURKA | 0.9528 |
| Leucine-rich repeat and fibronectin type-III domain-containing protein 5 | Q96NI6 | LRFN5 | 0.9520 |
| Proenkephalin-A | P01210 | PENK | 0.9503 |
| Scavenger receptor class B member 1 | B7ZKQ9 | SCARB1/SR-B1 | 0.9385 |
| Sortilin | Q99523 | SORT1 | 0.9375 |
| Tyrosine-protein kinase Fes/Fps | P07332 | FES | 0.9296 |
| Tetraspanin-6 | O43657 | TSPAN6 | 0.9283 |
| WD repeat and HMG-box DNA-binding protein 1 | O75717 | WDHD1 | 0.9185 |
| Monoglyceride lipase | Q99685 | MGLL | 0.9183 |
| Gamma-aminobutyric acid receptor-associated protein-like 2 | P60520 | GABARAPL2 | 0.9041 |
| Protein Name | Accession Number | Protein Symbol | Log2 FC Hypoxia vs. Normoxia |
|---|---|---|---|
| All-trans-retinol dehydrogenase [NAD (+)] ADH1B | ADH1B | P00325 | −2.2908 |
| Ubiquilin-1 (fragment) | UBQLN1 | H0YDS0 | −2.1746 |
| Collagen alpha-1(I) chain | COL1A1 | P02452 | −1.9743 |
| Aldehyde dehydrogenase, dimeric NADP-preferring | ALDH3A1 | P30838 | −1.9191 |
| Collagen alpha-1(XII) chain | COL12A1 | Q99715 | −1.8542 |
| Limbic system-associated membrane protein | LSAMP | Q13449 | −1.6349 |
| Collagen alpha-2(I) chain | COL1A2 | A0A087WTA8 | −1.4767 |
| Collagen alpha-1(XII) chain (fragment) | COL12A1 | H0Y5N9 | −1.4188 |
| Collagen alpha-1(III) chain | COL3A1 | P02461 | −1.3271 |
| Tropomodulin-1 | TMOD1 | P28289 | −1.3166 |
| Collagen triple helix repeat-containing protein 1 | CTHRC1 | Q96CG8 | −1.2917 |
| Collagen alpha-1(IV) chain | COL4A1 | P02462 | −1.2683 |
| Isoform 2 of Collagen alpha-1 (V) chain | COL5A1 | P20908-2 | −1.2281 |
| Neuroserpin | SERPINI1 | Q99574 | −1.2189 |
| Transducin-like enhancer protein 4 | TLE4 | Q04727 | −1.2179 |
| BH3-interacting domain death agonist 1 | BID | P55957 | −1.1611 |
| NAD(P)H dehydrogenase [quinone] 1 | NQO1 | P15559 | −1.1425 |
| Isoform TrkB-T1 of BDNF/NT-3 growth factors receptor | NTRK2 | Q16620-2 | −1.1343 |
| Protein phosphatase 1L | PPM1L | Q5SGD2 | −1.0958 |
| Collagen alpha-2(V) chain | COL5A2 | P05997 | −1.0816 |
| Collagen alpha-1(XIV) chain | COL14A1 | Q05707 | −1.0782 |
| 10-formyltetrahydrofolate dehydrogenase | ALDH1L2 | A0A494C1M4 | −1.0602 |
| Cytochrome c oxidase subunit NDUFA4 | NDUFA4 | O00483 | −1.0348 |
| Peptidyl-prolyl cis-trans isomerase FKBP9 | FKBP9 | O95302 | −1.0305 |
| Fibroblast growth factor 1 | FGF1 | P05230 | −1.0188 |
| Cell cycle exit and neuronal differentiation protein 1 | CEND1 | Q8N111 | −1.0105 |
| Tissue alpha-L-fucosidase | FUCA1 | P04066 | −1.0054 |
| Peptidyl-prolyl cis-trans isomerase FKBP10 | FKBP10 | Q96AY3 | −0.9833 |
| Alkaline phosphatase, tissue-nonspecific isozyme | ALPL | P05186 | −0.9812 |
| Glypican-4 | GPC4 | O75487 | −0.9779 |
| Peptidyl-prolyl cis-trans isomerase C | PPIC | P45877 | −0.9732 |
| Cystathionine beta-synthase-like protein | CBS | P0DN79 | −0.9432 |
| Protein disulfide-isomerase A5 | PDIA5 | Q14554 | −0.9414 |
| Pyrroline-5-carboxylate reductase 1, mitochondrial | PYCR1 | P32322 | −0.9383 |
| Isoform 6 of Dystrophin | DMD | P11532-6 | −0.9348 |
| Glycerol-3-phosphate dehydrogenase [NAD (+)], cytoplasmic | GPD1 | P21695 | −0.9348 |
| Ectonucleotide pyrophosphatase/phosphodiesterase family member 2 | ENPP2 | E5RIA2 | −0.9187 |
| Ubiquitin carboxyl-terminal hydrolase isozyme L1 | UCHL1 | P09936 | −0.9146 |
| Mitochondrial carnitine/acylcarnitine carrier protein | SLC25A20 | O43772 | −0.9108 |
| Peptidyl-prolyl cis-trans isomerase FKBP11 | FKBP11 | Q9NYL4 | −0.9097 |
| Phosphoserine aminotransferase | PSAT1 | Q9Y617 | −0.9057 |
| Delta-1-pyrroline-5-carboxylate synthase | ALDH18A1 | P54886 | −0.9056 |
| Collagen Types | Log2 FC Hypoxia vs. Normoxia |
|---|---|
| COL1A1 | −1.9743 |
| COL12A1 | −1.8542 |
| COL1A2 | −1.4767 |
| COL3A1 | −1.3271 |
| COL4A1 | −1.2683 |
| COL5A1 | −1.2281 |
| COL5A2 | −1.0816 |
| COL14A1 | −1.0782 |
| COL18A1 | −0.6919 |
| COL11A1 | −0.3977 |
| COL6A6 | −0.3544 |
| COL16A1 | −0.3072 |
| COL6A1 | −0.2727 |
| COL6A3 | 0.1690 |
| COL2A1 | 0.2588 |
| COL8A1 | 0.2618 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeannerat, A.; Peneveyre, C.; Armand, F.; Chiappe, D.; Hamelin, R.; Scaletta, C.; Hirt-Burri, N.; de Buys Roessingh, A.; Raffoul, W.; Applegate, L.A.; et al. Hypoxic Incubation Conditions for Optimized Manufacture of Tenocyte-Based Active Pharmaceutical Ingredients of Homologous Standardized Transplant Products in Tendon Regenerative Medicine. Cells 2021, 10, 2872. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10112872
Jeannerat A, Peneveyre C, Armand F, Chiappe D, Hamelin R, Scaletta C, Hirt-Burri N, de Buys Roessingh A, Raffoul W, Applegate LA, et al. Hypoxic Incubation Conditions for Optimized Manufacture of Tenocyte-Based Active Pharmaceutical Ingredients of Homologous Standardized Transplant Products in Tendon Regenerative Medicine. Cells. 2021; 10(11):2872. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10112872
Chicago/Turabian StyleJeannerat, Annick, Cédric Peneveyre, Florence Armand, Diego Chiappe, Romain Hamelin, Corinne Scaletta, Nathalie Hirt-Burri, Anthony de Buys Roessingh, Wassim Raffoul, Lee Ann Applegate, and et al. 2021. "Hypoxic Incubation Conditions for Optimized Manufacture of Tenocyte-Based Active Pharmaceutical Ingredients of Homologous Standardized Transplant Products in Tendon Regenerative Medicine" Cells 10, no. 11: 2872. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10112872